
UHPLC-MS-Based Metabolomics Reveal the Potential Mechanism of Armillaria mellea Acid Polysaccharide in and Its Effects on Cyclophosphamide-Induced Immunosuppressed Mice

Abstract
:1. Introduction
2. Results
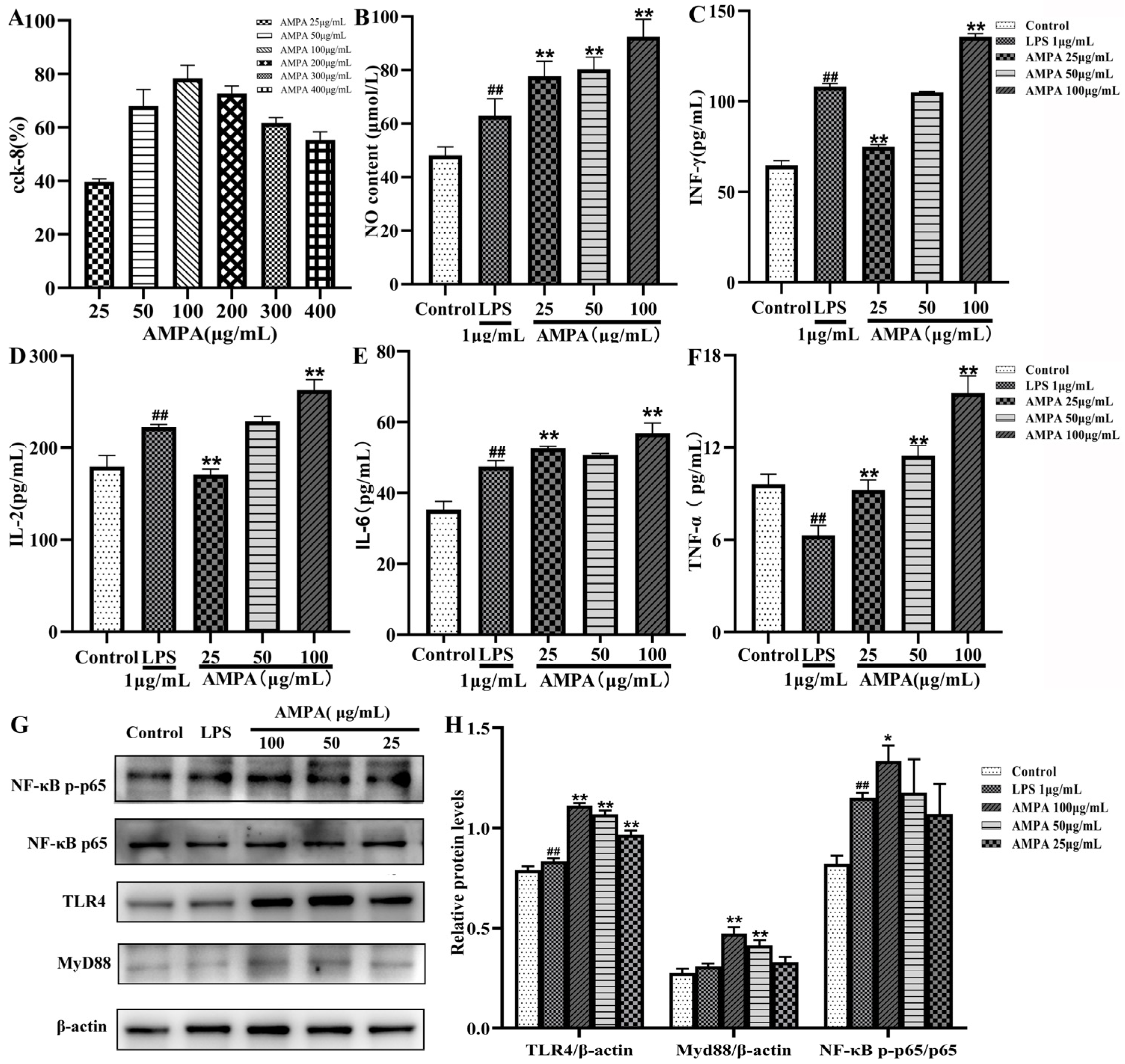
2.1. Effect of AMPA on RAW264.7 Cell Proliferation
2.2. Effects of AMPA on the Production of NO in RAW264.7 Cells
2.3. AMPA Promotes Cytokine Secretion by RAW264.7 Cells
2.4. AMPA Activates the TLR4/MyD88/NF-κB Signaling Pathway in RAW264.7 Cells
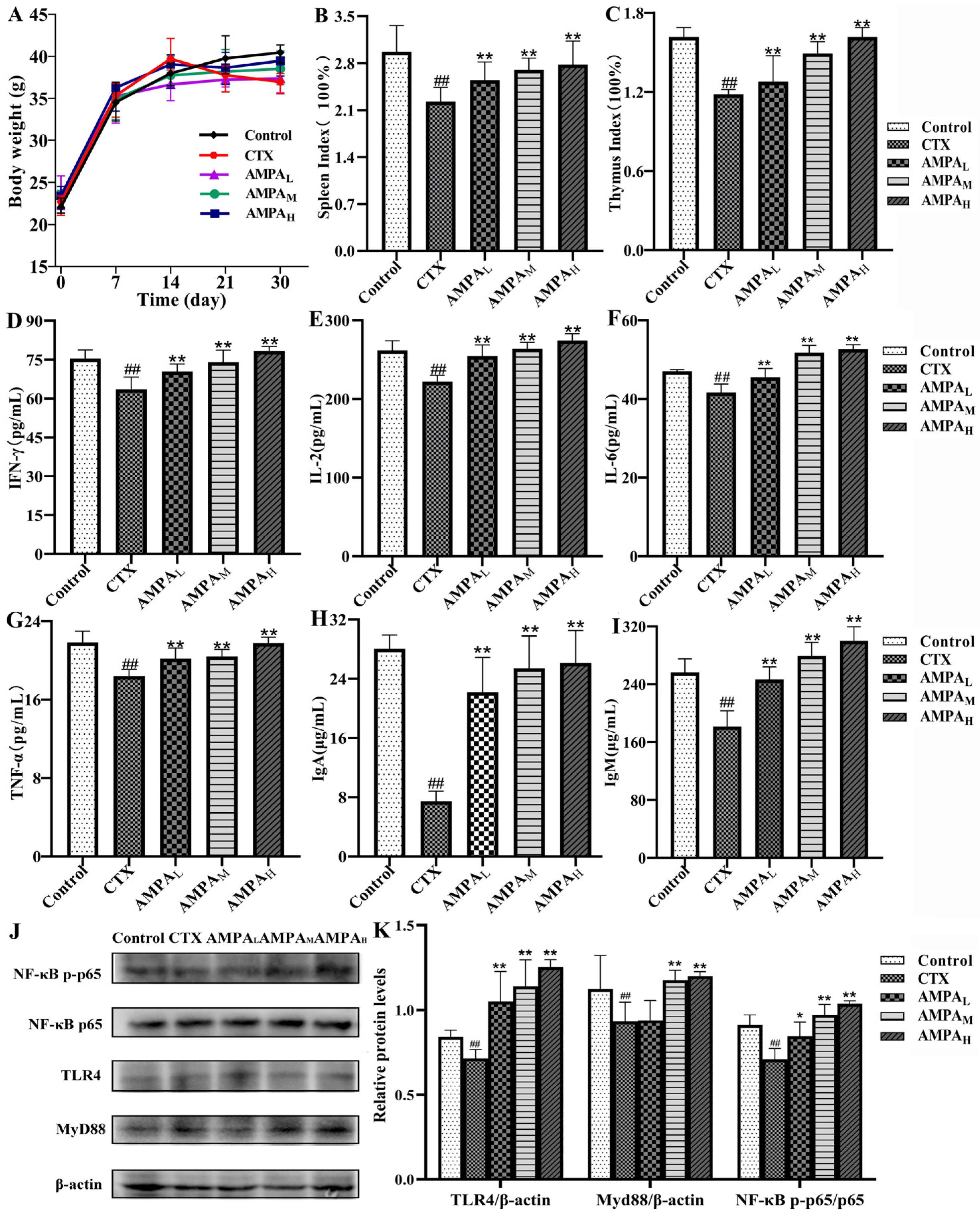
2.5. AMPA Improves the Physiological and Biochemical Parameters of CTX-Treated Mice
2.6. AMPA Activates the TLR4/MyD88/NF-κB Signal Pathway in CTX-Treated Mice
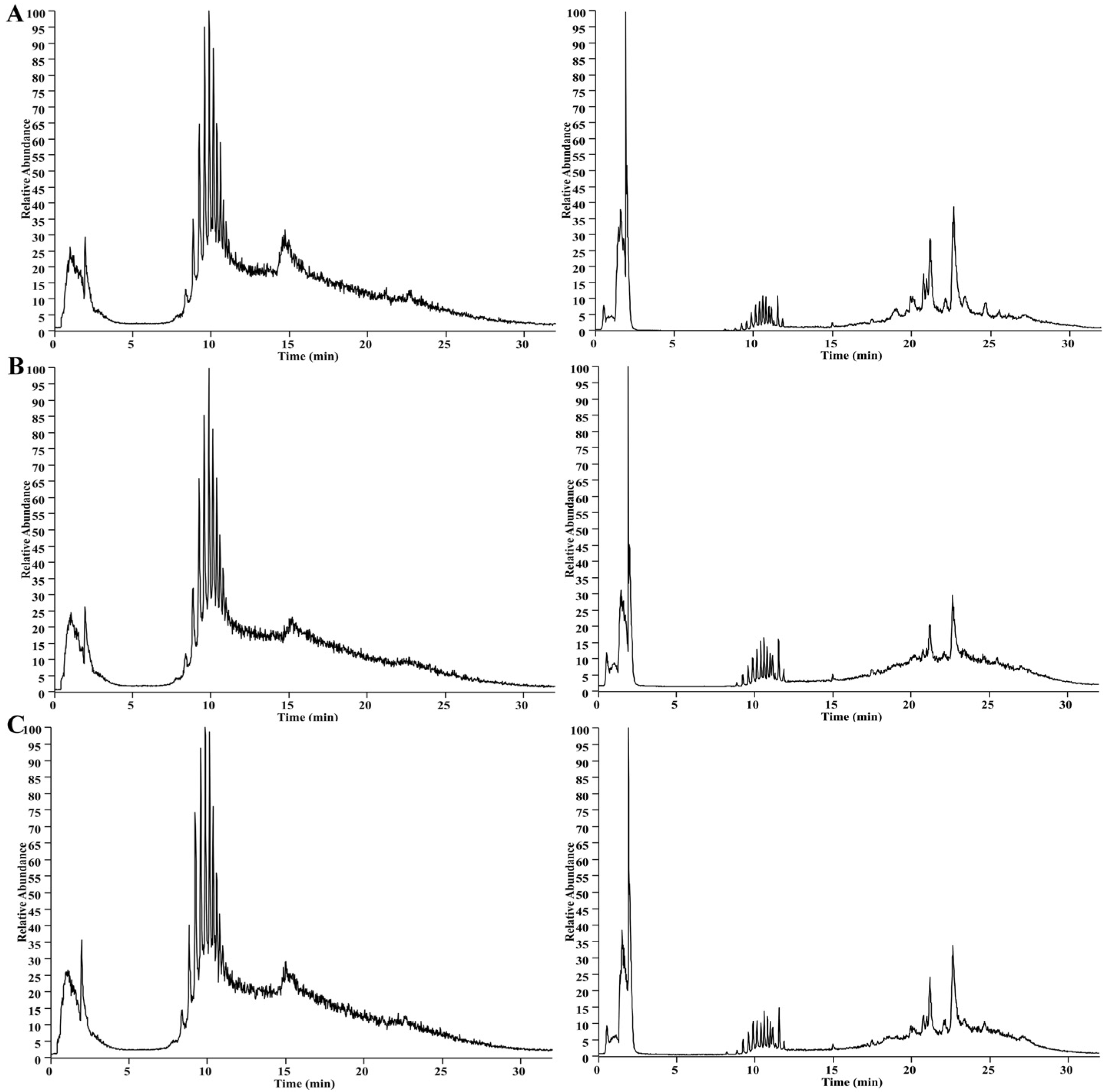
2.7. UHPLC-MS-Based Serum Metabolomics Analysis
2.8. Metabolic Pathway Analysis
3. Discussion
4. Materials and Methods
4.1. Preparation of AMPA
4.2. Materials and Instruments
4.3. RAW264.7 Cells for the Experiments
4.3.1. Evaluating the Optimal Concentration of AMPA and RAW264.7
4.3.2. Assessment of NO Production and ELISA Kits
4.3.3. Western Blot Analysis of the AMPA-Activated RAW264.7 Cells
4.4. Animals and Treatment
4.5. Serum UPLC-MS Metabolomic Analysis
4.6. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Bernal-Bayard, J.; Thiebaud, J.; Brossaud, M.; Beaussart, A.; Caillet, C.; Waldvogel, Y.; Travier, L.; Létoffé, S.; Fontaine, T.; Rokbi, B.; et al. Bacterial Capsular Polysaccharides with Antibiofilm Activity Share Common Biophysical and Electrokinetic Properties. Nat. Commun. 2023, 14, 2553. [Google Scholar] [CrossRef] [PubMed]
- Erbiai, E.H.; da Silva, L.P.; Saidi, R.; Lamrani, Z.; Esteves da Silva, J.C.G.; Maouni, A. Chemical Composition, Bioactive Compounds, and Antioxidant Activity of Two Wild Edible Mushrooms Armillaria Mellea and Macrolepiota Procera from Two Countries (Morocco and Portugal). Biomolecules 2021, 11, 575. [Google Scholar] [CrossRef] [PubMed]
- Nowacka-Jechalke, N.; Kanak, S.; Moczulski, M.; Martyna, A.; Kubiński, K.; Masłyk, M.; Szpakowska, N.; Kaczyński, Z.; Nowak, R.; Olech, M. Crude Polysaccharides from Wild-Growing Armillaria Mellea—Chemical Composition and Antidiabetic, Anti-Inflammatory, Antioxidant, and Antiproliferative Potential. Appl. Sci. 2023, 13, 3853. [Google Scholar] [CrossRef]
- Ooi, V.E.; Liu, F. Immunomodulation and Anti-Cancer Activity of Polysaccharide-Protein Complexes. Curr. Med. Chem. 2000, 7, 715–729. [Google Scholar] [CrossRef] [PubMed]
- Ying, M.; Yu, Q.; Zheng, B.; Wang, H.; Wang, J.; Chen, S.; Nie, S.; Xie, M. Cultured Cordyceps Sinensis Polysaccharides Modulate Intestinal Mucosal Immunity and Gut Microbiota in Cyclophosphamide-Treated Mice. Carbohydr. Polym. 2020, 235, 115957. [Google Scholar] [CrossRef] [PubMed]
- Niu, Y.; Dong, J.; Jiang, H.; Wang, J.; Liu, Z.; Ma, C.; Kang, W. Effects of Polysaccharide from Malus Halliana Koehne Flowers in Cyclophosphamide-Induced Immunosuppression and Oxidative Stress on Mice. Oxid. Med. Cell. Longev. 2020, 2020, 1603735. [Google Scholar] [CrossRef]
- Chen, R.; Ren, X.; Yin, W.; Lu, J.; Tian, L.; Zhao, L.; Yang, R.; Luo, S. Ultrasonic Disruption Extraction, Characterization and Bioactivities of Polysaccharides from Wild Armillaria Mellea. Int. J. Biol. Macromol. 2020, 156, 1491–1502. [Google Scholar] [CrossRef]
- Li, Y.; Zheng, J.; Wang, Y.; Yang, H.; Cao, L.; Gan, S.; Ma, J.; Liu, H. Immuno-Stimulatory Activity of Astragalus Polysaccharides in Cyclophosphamide-Induced Immunosuppressed Mice by Regulating Gut Microbiota. Int. J. Biol. Macromol. 2023, 242, 124789. [Google Scholar] [CrossRef]
- Feng, S.; Ding, H.; Liu, L.; Peng, C.; Huang, Y.; Zhong, F.; Li, W.; Meng, T.; Li, J.; Wang, X.; et al. Astragalus Polysaccharide Enhances the Immune Function of RAW264.7 Macrophages via the NF-κB P65/MAPK Signaling Pathway. Exp. Ther. Med. 2021, 21, 20. [Google Scholar] [CrossRef]
- Xie, Z.; Jiang, N.; Lin, M.; He, X.; Li, B.; Dong, Y.; Chen, S.; Lv, G. The Mechanisms of Polysaccharides from Tonic Chinese Herbal Medicine on the Enhancement Immune Function: A Review. Molecules 2023, 28, 7355. [Google Scholar] [CrossRef]
- Han, C.; Wang, Y.; Liu, R.; Ran, B.; Li, W. Structural Characterization and Protective Effect of Lonicerae Flos Polysaccharide on Cyclophosphamide-Induced Immunosuppression in Mice. Ecotoxicol. Environ. Saf. 2022, 230, 113174. [Google Scholar] [CrossRef] [PubMed]
- Cheong, K.-L.; Yu, B.; Chen, J.; Zhong, S. A Comprehensive Review of the Cardioprotective Effect of Marine Algae Polysaccharide on the Gut Microbiota. Foods 2022, 11, 3550. [Google Scholar] [CrossRef] [PubMed]
- Ren, J.-L.; Dong, H.; Han, Y.; Yang, L.; Zhang, A.-H.; Sun, H.; Li, Y.; Yan, G.; Wang, X.-J. Network Pharmacology Combined with Metabolomics Approach to Investigate the Protective Role and Detoxification Mechanism of Yunnan Baiyao Formulation. Phytomedicine 2020, 77, 153266. [Google Scholar] [CrossRef] [PubMed]
- O’Neill, L.A.J.; Kishton, R.J.; Rathmell, J. A Guide to Immunometabolism for Immunologists. Nat. Rev. Immunol. 2016, 16, 553–565. [Google Scholar] [CrossRef] [PubMed]
- Lou, X.; Ye, Y.; Wang, Y.; Sun, Y.; Pan, D.; Cao, J. Effect of High-Pressure Treatment on Taste and Metabolite Profiles of Ducks with Two Different Vinasse-Curing Processes. Food Res. Int. 2018, 105, 703–712. [Google Scholar] [CrossRef] [PubMed]
- Han, J.; Qin, W.-X.; Li, Z.-L.; Xu, A.-J.; Xing, H.; Wu, H.; Zhang, H.; Wang, M.; Li, C.; Liang, L.; et al. Tissue and Serum Metabolite Profiling Reveals Potential Biomarkers of Human Hepatocellular Carcinoma. Clin. Chim. Acta 2019, 488, 68–75. [Google Scholar] [CrossRef] [PubMed]
- Sun, Y.; Wang, F.; Liu, Y.; Liu, S.; An, Y.; Xue, H.; Wang, J.; Xia, F.; Chen, X.; Cao, Y. Microbiome-Metabolome Responses of Fuzhuan Brick Tea Crude Polysaccharides with Immune-Protective Benefit in Cyclophosphamide-Induced Immunosuppressive Mice. Food Res. Int. 2022, 157, 111370. [Google Scholar] [CrossRef]
- Xie, H.; Hu, M.; Yu, J.; Yang, X.; Li, J.; Yu, N.; Han, L.; Peng, D. Mass Spectrometry-Based Metabolomics Reveal Dendrobium Huoshanense Polysaccharide Effects and Potential Mechanism of N-Methyl-N’-Nitro-N-Nitrosoguanidine -Induced Damage in GES-1 Cells. J. Ethnopharmacol. 2023, 310, 116342. [Google Scholar] [CrossRef]
- Liu, X.; Xie, J.; Jia, S.; Huang, L.; Wang, Z.; Li, C.; Xie, M. Immunomodulatory Effects of an Acetylated Cyclocarya Paliurus Polysaccharide on Murine Macrophages RAW264.7. Int. J. Biol. Macromol. 2017, 98, 576–581. [Google Scholar] [CrossRef]
- Dai, J.; Chen, J.; Qi, J.; Ding, M.; Liu, W.; Shao, T.; Han, J.; Wang, G. Konjac Glucomannan from Amorphophallus Konjac Enhances Immunocompetence of the Cyclophosphamide-Induced Immunosuppressed Mice. Food Sci. Nutr. 2021, 9, 728–735. [Google Scholar] [CrossRef]
- Gong, A.G.; Zhang, L.M.; Lam, C.T.; Xu, M.L.; Wang, H.Y.; Lin, H.Q.; Dong, T.T.; Tsim, K.W. Polysaccharide of Danggui Buxue Tang, an Ancient Chinese Herbal Decoction, Induces Expression of Pro-Inflammatory Cytokines Possibly Via Activation of NFκB Signaling in Cultured RAW 264.7 Cells. Phytother. Res. 2017, 31, 274–283. [Google Scholar] [CrossRef] [PubMed]
- Martínez-Reyes, I.; Chandel, N.S. Mitochondrial TCA Cycle Metabolites Control Physiology and Disease. Nat. Commun. 2020, 11, 102. [Google Scholar] [CrossRef] [PubMed]
- Zasłona, Z.; O’Neill, L.A.J. Cytokine-like Roles for Metabolites in Immunity. Mol. Cell 2020, 78, 814–823. [Google Scholar] [CrossRef] [PubMed]
- Weis, G.C.C.; Assmann, C.E.; Cadoná, F.C.; Bonadiman, B. da S.R.; Alves, A. de O.; Machado, A.K.; Duarte, M.M.M.F.; da Cruz, I.B.M.; Costabeber, I.H. Immunomodulatory Effect of Mancozeb, Chlorothalonil, and Thiophanate Methyl Pesticides on Macrophage Cells. Ecotoxicol. Environ. Saf. 2019, 182, 109420. [Google Scholar] [CrossRef] [PubMed]
- Zhao, R.-H.; Yang, F.-X.; Bai, Y.-C.; Zhao, J.-Y.; Hu, M.; Zhang, X.-Y.; Dou, T.-F.; Jia, J.-J. Research Progress on the Mechanisms Underlying Poultry Immune Regulation by Plant Polysaccharides. Front. Vet. Sci. 2023, 10, 1175848. [Google Scholar] [CrossRef] [PubMed]
- Zhu, G.; Jiang, Y.; Yao, Y.; Wu, N.; Luo, J.; Hu, M.; Tu, Y.; Xu, M. Ovotransferrin Ameliorates the Dysbiosis of Immunomodulatory Function and Intestinal Microbiota Induced by Cyclophosphamide. Food Funct. 2019, 10, 1109–1122. [Google Scholar] [CrossRef] [PubMed]
- Liu, H.; Yan, Y.; Zhang, F.; Wu, Q. The Immuno-Enhancement Effects of Tubiechong (Eupolyphaga Sinensis) Lyophilized Powder in Cyclophosphamide-Induced Immunosuppressed Mice. Immunol. Investig. 2019, 48, 844–859. [Google Scholar] [CrossRef]
- Hayes, C.; Donohoe, C.L.; Davern, M.; Donlon, N.E. The Oncogenic and Clinical Implications of Lactate Induced Immunosuppression in the Tumour Microenvironment. Cancer Lett. 2021, 500, 75–86. [Google Scholar] [CrossRef]
- Dichtl, S.; Lindenthal, L.; Zeitler, L.; Behnke, K.; Schlösser, D.; Strobl, B.; Scheller, J.; El Kasmi, K.C.; Murray, P.J. Lactate and IL6 Define Separable Paths of Inflammatory Metabolic Adaptation. Sci. Adv. 2021, 7, eabg3505. [Google Scholar] [CrossRef]
- Hinder, L.M.; Vivekanandan-Giri, A.; McLean, L.L.; Pennathur, S.; Feldman, E.L. Decreased Glycolytic and Tricarboxylic Acid Cycle Intermediates Coincide with Peripheral Nervous System Oxidative Stress in a Murine Model of Type 2 Diabetes. J. Endocrinol. 2013, 216, 1–11. [Google Scholar] [CrossRef]
- Liu, Y.; Huang, R.; Liu, L.; Peng, J.; Xiao, B.; Yang, J.; Miao, Z.; Huang, H. Metabonomics Study of Urine from Sprague-Dawley Rats Exposed to Huang-Yao-Zi Using (1)H NMR Spectroscopy. J. Pharm. Biomed. Anal. 2010, 52, 136–141. [Google Scholar] [CrossRef] [PubMed]
- Roosterman, D.; Cottrell, G.S. Rethinking the Citric Acid Cycle: Connecting Pyruvate Carboxylase and Citrate Synthase to the Flow of Energy and Material. Int. J. Mol. Sci. 2021, 22, 604. [Google Scholar] [CrossRef] [PubMed]
- Conte, F.; van Buuringen, N.; Voermans, N.C.; Lefeber, D.J. Galactose in Human Metabolism, Glycosylation and Congenital Metabolic Diseases: Time for a Closer Look. Biochim. Biophys. Acta Gen. Subj. 2021, 1865, 129898. [Google Scholar] [CrossRef] [PubMed]
- Rolfsson, Ó.; Johannsson, F.; Magnusdottir, M.; Paglia, G.; Sigurjonsson, Ó.E.; Bordbar, A.; Palsson, S.; Brynjólfsson, S.; Guðmundsson, S.; Palsson, B. Mannose and Fructose Metabolism in Red Blood Cells during Cold Storage in SAGM. Transfusion 2017, 57, 2665–2676. [Google Scholar] [CrossRef] [PubMed]
- Morris, S.M. Arginine Metabolism Revisited12. J. Nutr. 2016, 146, 2579S–2586S. [Google Scholar] [CrossRef]
- Taddei, M.L.; Pardella, E.; Pranzini, E.; Raugei, G.; Paoli, P. Role of Tyrosine Phosphorylation in Modulating Cancer Cell Metabolism. Biochim. Biophys. Acta Rev. Cancer 2020, 1874, 188442. [Google Scholar] [CrossRef]
- Garavito, M.F.; Narváez-Ortiz, H.Y.; Zimmermann, B.H. Pyrimidine Metabolism: Dynamic and Versatile Pathways in Pathogens and Cellular Development. J. Genet. Genom. 2015, 42, 195–205. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| ID | Rt (min) | Query Mass | Metabolite Name | Formula | Ion Forms | Compound ID | KEGG | VIP | FC | Change Tend | ||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| CTX/ Control | AMPA/ CTX | CTX/ Control | AMPA/ CTX | |||||||||
| 1 | 6.8 | 90.5268 | D-4-Hydroxy -2-oxoglutarate | C5H6O6 | M+H+NH4 | HMDB 0060466 | C05946 | 1.21 | 0.95 | 0.86 | ↓ | ↓ |
| 2 | 9.7 | 90.5268 | Gentisate aldehyde | C7H6O3 | M+2H | HMDB 0004062 | C05585 | 1.14 | 0.96 | 1.13 | ↓ | ↑ |
| 3 | 11.2 | 90.5268 | 3-Hydroxybenzoic acid | C7H6O3 | M+2H | HMDB 0002466 | C00587 | 1.33 | 0.95 | 0.96 | ↓ | ↓ |
| 4 | 11.4 | 205.0609 | (±)-2-(1-Methylpropyl)-4,6-dinitrophenol | C10H12N2O5 | M+H-2H2O | HMDB 0032559 | C14302 | 1.21 | 0.98 | 0.97 | ↓ | ↓ |
| 5 | 13.4 | 205.0609 | Benzophenone | C13H10O | M+Na | HMDB 0032049 | C06354 | 1.02 | 0.99 | 0.97 | ↓ | ↓ |
| 6 | 16.9 | 90.5268 | Hydroxy -2-oxoglutaric acid | C5H6O6 | M+H+NH4 | HMDB 0002070 | C01127 | 1.09 | 0.96 | 1.09 | ↓ | ↑ |
| 7 | 18.4 | 90.5267 | L-Hypoglycin A | C7H11NO2 | M+H+K | HMDB 0029427 | C08287 | 1.21 | 0.96 | 0.98 | ↓ | ↓ |
| 8 | 29.8 | 166.0982 | 3-Hydroxyquinine | C20H24N2O3 | M+H-H2O | HMDB 0001091 | C07344 | 1.08 | 0.96 | 0.96 | ↓ | ↓ |
| 9 | 8.6 | 177.0077 | 3,5-Dihydroxybenzoic acid | C7H6O4 | M+Na | HMDB 0013677 | C00180 | 1.17 | 0.98 | 0.95 | ↓ | ↓ |
| 10 | 20.5 | 205.0609 | D-Sorbitol | C6H14O6 | M+Na | HMDB 0000247 | C00794 | 1.34 | 0.97 | 1.34 | ↓ | ↑ |
| 11 | 10.3 | 177.0077 | Protocatechuic acid | C7H6O4 | M+Na | HMDB 0001856 | C00230 | 1.14 | 0.97 | 0.95 | ↓ | ↓ |
| 12 | 13.1 | 177.0078 | Galactitol | C6H14O6 | M+Na | HMDB 0000107 | C01697 | 1.23 | 0.96 | 1.23 | ↓ | ↑ |
| 13 | 18.9 | 255.9450 | 2-Pyrocatechuic acid | C7H6O4 | M+Na | HMDB 0000397 | C00196 | 1.19 | 0.97 | 0.94 | ↓ | ↓ |
| 14 | 21.8 | 205.0607 | L-Proline | C5H9NO2 | M+H-2H2O | HMDB 0000162 | C00148 | 1.10 | 0.98 | 1.10 | ↓ | ↑ |
| 15 | 12.0 | 177.0076 | Orotic acid | C5H4N2O4 | M+H-H2O | HMD B0000226 | C00295 | 1.24 | 1.23 | 0.94 | ↑ | ↓ |
| Pathway Name | Match Status | p | −log(p) | Holm p | FDR | Impact |
|---|---|---|---|---|---|---|
| Galactose metabolism | 2/27 | 0.0061524 | 2.211 | 0.5168 | 0.50498 | 0.0 |
| Arginine and proline metabolism | 2/38 | 0.012023 | 1.92 | 0.99793 | 0.50498 | 0.10165 |
| Fructose and mannose metabolism | 1/18 | 0.08088 | 1.0922 | 1.0 | 1.0 | 0.03313 |
| Glyoxylate and dicarboxylate metabolism | 1/32 | 0.13985 | 0.85435 | 1.0 | 1.0 | 0.07937 |
| Pyrimidine metabolism | 1/39 | 0.16809 | 0.77445 | 1.0 | 1.0 | 0.04819 |
| Tyrosine metabolism | 1/42 | 0.17995 | 0.74484 | 1.0 | 1.0 | 0.0 |
| Aminoacyl-tRNA biosynthesis | 1/48 | 0.20324 | 0.69199 | 1.0 | 1.0 | 0.0 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, Y.; Li, Q.; Niu, H.; Li, H.; Jiao, L.; Wu, W. UHPLC-MS-Based Metabolomics Reveal the Potential Mechanism of Armillaria mellea Acid Polysaccharide in and Its Effects on Cyclophosphamide-Induced Immunosuppressed Mice. Molecules 2023, 28, 7944. https://doi.org/10.3390/molecules28247944
Li Y, Li Q, Niu H, Li H, Jiao L, Wu W. UHPLC-MS-Based Metabolomics Reveal the Potential Mechanism of Armillaria mellea Acid Polysaccharide in and Its Effects on Cyclophosphamide-Induced Immunosuppressed Mice. Molecules. 2023; 28(24):7944. https://doi.org/10.3390/molecules28247944
Chicago/Turabian StyleLi, Ying, Qingqing Li, Huazhou Niu, Hui Li, Lili Jiao, and Wei Wu. 2023. "UHPLC-MS-Based Metabolomics Reveal the Potential Mechanism of Armillaria mellea Acid Polysaccharide in and Its Effects on Cyclophosphamide-Induced Immunosuppressed Mice" Molecules 28, no. 24: 7944. https://doi.org/10.3390/molecules28247944
APA StyleLi, Y., Li, Q., Niu, H., Li, H., Jiao, L., & Wu, W. (2023). UHPLC-MS-Based Metabolomics Reveal the Potential Mechanism of Armillaria mellea Acid Polysaccharide in and Its Effects on Cyclophosphamide-Induced Immunosuppressed Mice. Molecules, 28(24), 7944. https://doi.org/10.3390/molecules28247944