
Effects of Natural Products on Enzymes Involved in Ferroptosis: Regulation and Implications

 ,
,  ,
,
Abstract
:1. Introduction
2. Results and Discussion
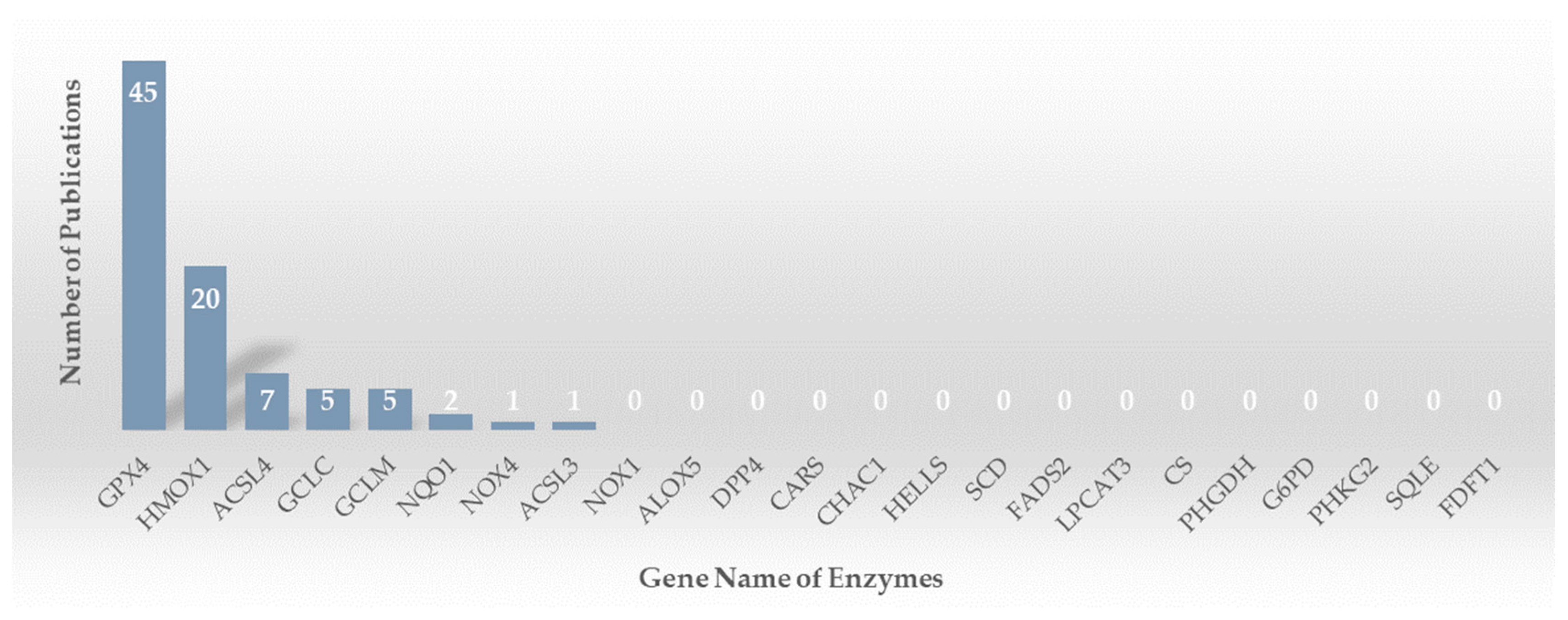
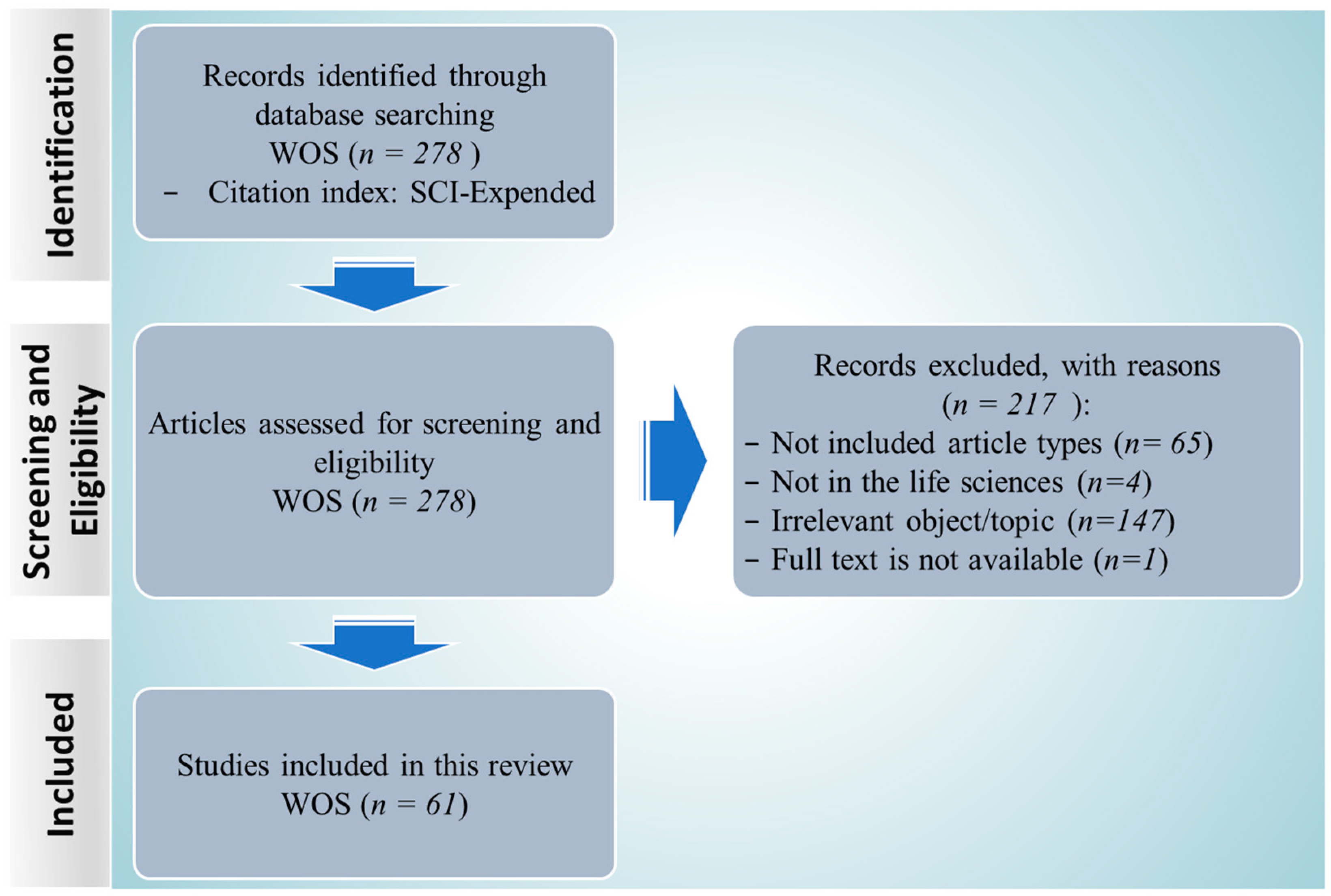
2.1. Search Results and Study Inclusion
2.2. Study Characteristics
2.3. The Molecular Mechanisms of Ferroptosis
2.4. Enzymes-Mediated Phospholipid Peroxidation
2.5. The Effects of Natural Products on the Enzymes Involving Ferroptosis
2.6. The Effects of Natural Products on the Enzymes Involving Glutathione Metabolic Pathway
2.6.1. GPX4
2.6.2. HO-1
2.6.3. NQO1
2.6.4. NOX4
2.6.5. GCLC and GCLM
2.7. The Effects of Natural Products on Enzymes Involving Lipid Metabolism
3. Materials and Methods
3.1. Data Retrieval
3.2. Screening and Eligibility
3.3. Annotated Bibliography
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Tang, D.; Chen, X.; Kang, R.; Kroemer, G. Ferroptosis: Molecular mechanisms and health implications. Cell Res. 2021, 31, 107–125. [Google Scholar] [CrossRef] [PubMed]
- Dixon, S.J.; Lemberg, K.M.; Lamprecht, M.R.; Skouta, R.; Zaitsev, E.M.; Gleason, C.E.; Patel, D.N.; Bauer, A.J.; Cantley, A.M.; Yang, W.S. Ferroptosis: An iron-dependent form of nonapoptotic cell death. Cell 2012, 149, 1060–1072. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Cao, F.; Yin, H.L.; Huang, Z.J.; Lin, Z.T.; Mao, N.; Sun, B.; Wang, G. Ferroptosis: Past, present and future. Cell Death Dis. 2020, 11, 88. [Google Scholar] [CrossRef] [PubMed]
- Dixon, S.J.; Stockwell, B.R. The Hallmarks of Ferroptosis. Annu. Rev. Cancer Biol. 2019, 3, 35–54. [Google Scholar] [CrossRef]
- Zhang, C.; Liu, X.Y.; Jin, S.D.; Chen, Y.; Guo, R.H. Ferroptosis in cancer therapy: A novel approach to reversing drug resistance. Mol. Cancer 2022, 21, 47. [Google Scholar] [CrossRef] [PubMed]
- Lei, G.; Zhuang, L.; Gan, B.Y. Targeting ferroptosis as a vulnerability in cancer. Nat. Rev. Cancer 2022, 22, 381–396. [Google Scholar] [CrossRef]
- Ren, J.X.; Sun, X.; Yan, X.L.; Guo, Z.N.; Yang, Y. Ferroptosis in neurological diseases. Front. Cell. Neurosci. 2020, 14, 218. [Google Scholar] [CrossRef]
- Zhou, L.X.; Han, S.T.; Guo, J.Y.; Qiu, T.; Zhou, J.Q.; Shen, L. Ferroptosis—A New Dawn in the Treatment of Organ Ischemia–Reperfusion Injury. Cells 2022, 11, 3653. [Google Scholar] [CrossRef]
- Peleman, C.; Van Coillie, S.; Ligthart, S.; Choi, S.M.; De Waele, J.; Depuydt, P.; Benoit, D.; Schaubroeck, H.; Francque, S.M.; Dams, K. Ferroptosis and pyroptosis signatures in critical COVID-19 patients. Cell Death Differ. 2023, 30, 2066–2077. [Google Scholar] [CrossRef]
- Wang, Y.; Zhong, J.; Bai, J.J.; Tong, R.S.; An, F.F.; Jiao, P.C.; He, L.; Zeng, D.W.; Long, E.W.; Yan, J.F. The application of natural products in cancer therapy by targeting apoptosis pathways. Curr. Drug Metab. 2018, 19, 739–749. [Google Scholar] [CrossRef]
- Arulselvan, P.; Fard, M.T.; Tan, W.S.; Gothai, S.; Fakurazi, S.; Norhaizan, M.E.; Kumar, S.S. Role of antioxidants and natural products in inflammation. Oxidative Med. Cell. Longev. 2016, 2016, 5276130. [Google Scholar] [CrossRef]
- Rehman, M.U.; Wali, A.F.; Ahmad, A.; Shakeel, S.; Rasool, S.; Ali, R.; Rashid, S.M.; Madkhali, H.; Ganaie, M.A.; Khan, R. Neuroprotective strategies for neurological disorders by natural products: An update. Curr. Neuropharmacol. 2019, 17, 247–267. [Google Scholar] [CrossRef]
- Zheng, K.; Dong, Y.; Yang, R.; Liang, Y.F.; Wu, H.Q.; He, Z.D. Regulation of ferroptosis by bioactive phytochemicals: Implications for medical nutritional therapy. Pharmacol. Res. 2021, 168, 105580. [Google Scholar] [CrossRef] [PubMed]
- Kibble, M.; Saarinen, N.; Tang, J.; Wennerberg, K.; Mäkelä, S.; Aittokallio, T. Network pharmacology applications to map the unexplored target space and therapeutic potential of natural products. Nat. Prod. Rep. 2015, 32, 1249–1266. [Google Scholar] [CrossRef]
- Ma, T.Y.; Du, J.T.; Zhang, Y.F.; Wang, Y.Y.; Wang, B.X.; Zhang, T.H. GPX4-independent ferroptosis—A new strategy in disease’s therapy. Cell Death Discov. 2022, 8, 434. [Google Scholar] [CrossRef]
- Ding, Y.H.; Chen, X.P.; Liu, C.; Ge, W.Z.; Wang, Q.; Hao, X.; Wang, M.M.; Chen, Y.; Zhang, Q. Identification of a small molecule as inducer of ferroptosis and apoptosis through ubiquitination of GPX4 in triple negative breast cancer cells. J. Hematol. Oncol. 2021, 14, 1–21. [Google Scholar] [CrossRef]
- Amos, A.; Amos, A.; Wu, L.R.; Xia, H. The Warburg effect modulates DHODH role in ferroptosis: A review. Cell Commun. Signal. 2023, 21, 100. [Google Scholar] [CrossRef] [PubMed]
- Kuang, F.M.; Liu, J.; Tang, D.L.; Kang, R. Oxidative damage and antioxidant defense in ferroptosis. Front. Cell Dev. Biol. 2020, 8, 586578. [Google Scholar] [CrossRef] [PubMed]
- Park, M.W.; Cha, H.W.; Kim, J.; Kim, J.H.; Yang, H.; Yoon, S.; Boonpraman, N.; Yi, S.S.; Yoo, I.D.; Moon, J.S. NOX4 promotes ferroptosis of astrocytes by oxidative stress-induced lipid peroxidation via the impairment of mitochondrial metabolism in Alzheimer’s diseases. Redox Biol. 2021, 41, 101947. [Google Scholar] [CrossRef]
- Lin, Z.; Liu, J.; Kang, R.; Yang, M.H.; Tang, D.L. Lipid metabolism in ferroptosis. Adv. Biol. 2021, 5, 2100396. [Google Scholar] [CrossRef]
- Gnanapradeepan, K.; Basu, S.; Barnoud, T.; Budina-Kolomets, A.; Kung, C.-P.; Murphy, M.E. The p53 tumor suppressor in the control of metabolism and ferroptosis. Front. Endocrinol. 2018, 9, 124. [Google Scholar] [CrossRef]
- Wei, R.R.; Zhao, Y.Q.; Wang, J.; Yang, X.; Li, S.L.; Wang, Y.Y.; Yang, X.Z.; Fei, J.M.; Hao, X.J.; Zhao, Y.H. Tagitinin C induces ferroptosis through PERK-Nrf2-HO-1 signaling pathway in colorectal cancer cells. Int. J. Biol. Sci. 2021, 17, 2703. [Google Scholar] [CrossRef]
- Yang, J.W.; Mo, J.; Dai, J.J.; Ye, C.Q.; Cen, W.; Zheng, X.Z.; Jiang, L.; Ye, L.C. Cetuximab promotes RSL3-induced ferroptosis by suppressing the Nrf2/HO-1 signalling pathway in KRAS mutant colorectal cancer. Cell Death Dis. 2021, 12, 1079. [Google Scholar] [CrossRef]
- Kang, Y.P.; Mockabee-Macias, A.; Jiang, C.; Falzone, A.; Prieto-Farigua, N.; Stone, E.; Harris, I.S.; DeNicola, G.M. Non-canonical Glutamate-Cysteine Ligase Activity Protects against Ferroptosis. Cell Metab. 2021, 33, 174–189. [Google Scholar] [CrossRef]
- Hayano, M.; Yang, W.S.; Corn, C.K.; Pagano, N.C.; Stockwell, B.R. Loss of cysteinyl-tRNA synthetase (CARS) induces the transsulfuration pathway and inhibits ferroptosis induced by cystine deprivation. Cell Death Differ. 2016, 23, 270–278. [Google Scholar] [CrossRef]
- Liu, Y.; Wu, D.; Fu, Q.L.; Hao, S.J.; Gu, Y.Z.; Zhao, W.; Chen, S.Y.; Sheng, F.Y.; Xu, Y.L.; Chen, Z.Q.; et al. CHAC1 as a Novel Contributor of Ferroptosis in Retinal Pigment Epithelial Cells with Oxidative Damage. Int. J. Mol. Sci. 2023, 24, 1582. [Google Scholar] [CrossRef]
- Chen, X.; Li, J.B.; Kang, R.; Klionsky, D.J.; Tang, D.L. Ferroptosis: Machinery and regulation. Autophagy 2021, 17, 2054–2081. [Google Scholar] [CrossRef] [PubMed]
- Xuan, Y.; Wang, H.; Yung, M.M.; Chen, F.; Chan, W.S.; Chan, Y.S.; Tsui, S.K.; Ngan, H.Y.; Chan, K.K.; Chan, D.W. SCD1/FADS2 fatty acid desaturases equipoise lipid metabolic activity and redox-driven ferroptosis in ascites-derived ovarian cancer cells. Theranostics 2022, 12, 3534–3552. [Google Scholar] [CrossRef] [PubMed]
- Yuan, Z.H.; Liu, T.; Wang, H.; Xue, L.X.; Wang, J.J. Fatty acids metabolism: The bridge between ferroptosis and ionizing radiation. Front. Cell Dev. Biol. 2021, 9, 675617. [Google Scholar] [CrossRef]
- Yang, Y.F.; Zhu, T.; Wang, X.; Xiong, F.; Hu, Z.M.; Qiao, X.H.; Yuan, X.; Wang, D.Q. ACSL3 and ACSL4, distinct roles in ferroptosis and cancers. Cancers 2022, 14, 5896. [Google Scholar] [CrossRef] [PubMed]
- Kagan, V.E.; Mao, G.W.; Qu, F.; Angeli, J.P.F.; Doll, S.; Croix, C.S.; Dar, H.H.; Liu, B.; Tyurin, V.A.; Ritov, V.B. Oxidized arachidonic and adrenic PEs navigate cells to ferroptosis. Nat. Chem. Biol. 2017, 13, 81–90. [Google Scholar] [CrossRef]
- Xie, Y.; Hou, W.; Song, X.; Yu, Y.; Huang, J.; Sun, X.; Kang, R.; Tang, D. Ferroptosis: Process and function. Cell Death Differ. 2016, 23, 369–379. [Google Scholar] [CrossRef] [PubMed]
- Grant, G.A. D-3-Phosphoglycerate dehydrogenase. Front. Mol. Biosci. 2018, 5, 110. [Google Scholar] [CrossRef] [PubMed]
- Meng, Q.F.; Zhang, Y.H.; Hao, S.M.; Sun, H.H.; Liu, B.; Zhou, H.L.; Wang, Y.S.; Xu, Z.X. Recent findings in the regulation of G6PD and its role in diseases. Front. Pharmacol. 2022, 13, 932154. [Google Scholar] [CrossRef]
- Lei, P.X.; Bai, T.; Sun, Y.L. Mechanisms of ferroptosis and relations with regulated cell death: A review. Front. Physiol. 2019, 10, 139. [Google Scholar] [CrossRef] [PubMed]
- Forcina, G.C.; Dixon, S.J. GPX4 at the crossroads of lipid homeostasis and ferroptosis. Proteomics 2019, 19, 1800311. [Google Scholar] [CrossRef]
- Picón, D.F.; Skouta, R. Unveiling the therapeutic potential of squalene synthase: Deciphering its biochemical mechanism, disease implications, and intriguing ties to ferroptosis. Cancers 2023, 15, 3731. [Google Scholar] [CrossRef] [PubMed]
- Maiorino, M.; Conrad, M.; Ursini, F. GPx4, lipid peroxidation, and cell death: Discoveries, rediscoveries, and open issues. Antioxid. Redox Signal. 2018, 29, 61–74. [Google Scholar] [CrossRef] [PubMed]
- Stockwell, B.R. Ferroptosis turns 10: Emerging mechanisms, physiological functions, and therapeutic applications. Cell 2022, 185, 2401–2421. [Google Scholar] [CrossRef]
- Yang, W.S.; SriRamaratnam, R.; Welsch, M.E.; Shimada, K.; Skouta, R.; Viswanathan, V.S.; Cheah, J.H.; Clemons, P.A.; Shamji, A.F.; Clish, C.B. Regulation of ferroptotic cancer cell death by GPX4. Cell 2014, 156, 317–331. [Google Scholar] [CrossRef]
- Yang, W.S.; Kim, K.J.; Gaschler, M.M.; Patel, M.; Shchepinov, M.S.; Stockwell, B.R. Peroxidation of polyunsaturated fatty acids by lipoxygenases drives ferroptosis. Proc. Natl. Acad. Sci. USA 2016, 113, E4966–E4975. [Google Scholar] [CrossRef]
- Xie, Y.C.; Song, X.X.; Sun, X.F.; Huang, J.; Zhong, M.Z.; Lotze, M.T.; Zeh, H.J., 3rd; Kang, R.; Tang, D.L. Identification of baicalein as a ferroptosis inhibitor by natural product library screening. Biochem. Biophys. Res. Commun. 2016, 473, 775–780. [Google Scholar] [CrossRef]
- Liu, Y.; Song, Z.; Liu, Y.J.; Ma, X.B.; Wang, W.; Ke, Y.; Xu, Y.C.; Yu, D.Q.; Liu, H.M. Identification of ferroptosis as a novel mechanism for antitumor activity of natural product derivative a2 in gastric cancer. Acta Pharm. Sin. B 2021, 11, 1513–1525. [Google Scholar] [CrossRef]
- Batbold, U.; Liu, J.J. Novel Insights of Herbal Remedy into NSCLC Suppression through Inducing Diverse Cell Death Pathways via Affecting Multiple Mediators. Appl. Sci. 2022, 12, 4868. [Google Scholar] [CrossRef]
- He, C.Y.; Wang, C.Z.; Liu, H.S.; Shan, B.E. Kayadiol exerted anticancer effects through p53-mediated ferroptosis in NKTCL cells. BMC Cancer 2022, 22, 724. [Google Scholar] [CrossRef]
- Zhang, M.Y.; Zhang, T.; Song, C.L.; Qu, J.; Gu, Y.P.; Liu, S.J.; Li, H.B.; Xiao, W.; Kong, L.D.; Sun, Y. Guizhi Fuling Capsule ameliorates endometrial hyperplasia through promoting p62-Keap1-NRF2-mediated ferroptosis. J. Ethnopharmacol. 2021, 274, 114064. [Google Scholar] [CrossRef]
- Greco, G.; Schnekenburger, M.; Catanzaro, E.; Turrini, E.; Ferrini, F.; Sestili, P.; Diederich, M.; Fimognari, C. Discovery of sulforaphane as an inducer of ferroptosis in u-937 leukemia cells: Expanding its anticancer potential. Cancers 2021, 14, 76. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Jiang, L.; Chen, H.; Wei, S.; Yao, K.; Sun, X.; Yang, G.; Jiang, L.; Zhang, C.; Wang, N. Resveratrol protected acrolein-induced ferroptosis and insulin secretion dysfunction via ER-stress-related PERK pathway in MIN6 cells. Toxicology 2022, 465, 153048. [Google Scholar] [CrossRef]
- Zhang, S.; Cao, S.; Zhou, H.; Li, L.; Hu, Q.; Mao, X.; Ji, S. Realgar-induced nephrotoxicity via ferroptosis in mice. J. Appl. Toxicol. 2022, 42, 1843–1853. [Google Scholar] [CrossRef]
- Wang, Y.; Wang, Z.; Wu, Z.; Chen, M.; Dong, D.; Yu, P.; Lu, D.; Wu, B. Involvement of REV-ERBα dysregulation and ferroptosis in aristolochic acid I-induced renal injury. Biochem. Pharmacol. 2021, 193, 114807. [Google Scholar] [CrossRef] [PubMed]
- Chang, W.T.; Bow, Y.D.; Fu, P.J.; Li, C.Y.; Wu, C.Y.; Chang, Y.H.; Teng, Y.N.; Li, R.N.; Lu, M.C.; Liu, Y.C.; et al. A Marine Terpenoid, Heteronemin, Induces Both the Apoptosis and Ferroptosis of Hepatocellular Carcinoma Cells and Involves the ROS and MAPK Pathways. Oxid. Med. Cell. Longev. 2021, 2021, 7689045. [Google Scholar] [CrossRef] [PubMed]
- Wen, Y.; Chen, H.; Zhang, L.; Wu, M.; Zhang, F.; Yang, D.; Shen, J.; Chen, J. Glycyrrhetinic acid induces oxidative/nitrative stress and drives ferroptosis through activating NADPH oxidases and iNOS, and depriving glutathione in triple-negative breast cancer cells. Free Radic. Biol. Med. 2021, 173, 41–51. [Google Scholar] [CrossRef]
- Wang, B.; Ma, W.; Di, Y.L. Activation of the Nrf2/GPX4 Signaling by Pratensein From Trifolium pretense Mitigates Ferroptosis in OGD/R-Insulted H9c2 Cardiomyocytes. Nat. Prod. Commun. 2022, 17, 1934578X221115313. [Google Scholar] [CrossRef]
- Li, X.P.; Li, W.; Yang, P.; Zhou, H.G.; Zhang, W.D.; Ma, L. Anticancer effects of Cryptotanshinone against lung cancer cells through ferroptosis. Arab. J. Chem. 2021, 14, 103177. [Google Scholar] [CrossRef]
- Tian, L.; Ji, H.; Wang, W.; Han, X.; Zhang, X.; Li, X.; Guo, L.; Huang, L.; Gao, W. Mitochondria-targeted pentacyclic triterpenoid carbon dots for selective cancer cell destruction via inducing autophagy, apoptosis, as well as ferroptosis. Bioorg. Chem. 2023, 130, 106259. [Google Scholar] [CrossRef]
- Wang, X.; Wang, Y.; Huang, D.; Shi, S.; Pei, C.; Wu, Y.; Shen, Z.; Wang, F.; Wang, Z. Astragaloside IV regulates the ferroptosis signaling pathway via the Nrf2/SLC7A11/GPX4 axis to inhibit PM2.5-mediated lung injury in mice. Int. Immunopharmacol. 2022, 112, 109186. [Google Scholar] [CrossRef]
- Li, J.; Tian, X.; Liu, J.; Mo, Y.; Guo, X.; Qiu, Y.; Liu, Y.; Ma, X.; Wang, Y.; Xiong, Y. Therapeutic material basis and underling mechanisms of Shaoyao Decoction-exerted alleviation effects of colitis based on GPX4-regulated ferroptosis in epithelial cells. Chin. Med. 2022, 17, 96. [Google Scholar] [CrossRef]
- Xu, X.; Wei, Y.; Hua, H.; Jing, X.; Zhu, H.; Xiao, K.; Zhao, J.; Liu, Y. Polyphenols Sourced from Ilex latifolia Thunb. Relieve Intestinal Injury via Modulating Ferroptosis in Weanling Piglets under Oxidative Stress. Antioxidants 2022, 11, 966. [Google Scholar] [CrossRef]
- Wang, C.; Su, Z.; Xu, J.H.; Ko, C.Y. Danshensu attenuated lipopolysaccharide-induced LX-2 and T6 cells activation through regulation of ferroptosis. Food Sci. Nutr. 2023, 11, 344–349. [Google Scholar] [CrossRef]
- Lan, B.; Ge, J.W.; Cheng, S.W.; Zheng, X.L.; Liao, J.; He, C.; Rao, Z.Q.; Wang, G.Z. Extract of Naotaifang, a compound Chinese herbal medicine, protects neuron ferroptosis induced by acute cerebral ischemia in rats. J. Integr. Med. 2020, 18, 344–350. [Google Scholar] [CrossRef]
- Huang, S.; Cao, B.; Zhang, J.; Feng, Y.; Wang, L.; Chen, X.; Su, H.; Liao, S.; Liu, J.; Yan, J.; et al. Induction of ferroptosis in human nasopharyngeal cancer cells by cucurbitacin B: Molecular mechanism and therapeutic potential. Cell Death Dis. 2021, 12, 237. [Google Scholar] [CrossRef] [PubMed]
- Han, N.; Li, L.G.; Peng, X.C.; Ma, Q.L.; Yang, Z.Y.; Wang, X.Y.; Li, J.; Li, Q.R.; Yu, T.T.; Xu, H.Z.; et al. Ferroptosis triggered by dihydroartemisinin facilitates chlorin e6 induced photodynamic therapy against lung cancerthrough inhibiting GPX4 and enhancing ROS. Eur. J. Pharmacol. 2022, 919, 174797. [Google Scholar] [CrossRef] [PubMed]
- Catanzaro, E.; Turrini, E.; Kerre, T.; Sioen, S.; Baeyens, A.; Guerrini, A.; Bellau, M.L.A.; Sacchetti, G.; Paganetto, G.; Krysko, D.V.; et al. Perillaldehyde is a new ferroptosis inducer with a relevant clinical potential for acute myeloid leukemia therapy. Biomed. Pharmacother. 2022, 154, 113662. [Google Scholar] [CrossRef] [PubMed]
- Du, X.; Zhang, J.; Liu, L.; Xu, B.; Han, H.; Dai, W.; Pei, X.; Fu, X.; Hou, S. A novel anticancer property of Lycium barbarum polysaccharide in triggering ferroptosis of breast cancer cells. J. Zhejiang Univ. Sci. B 2022, 23, 286–299. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.B.; Tang, P.L.; Cai, Q.; Xie, S.F.; Duan, X.Y.; Pan, Y.Z. Matrine can inhibit the growth of colorectal cancer cells by inducing ferroptosis. Nat. Prod. Commun. 2020, 15, 1934578X20982779. [Google Scholar] [CrossRef]
- Liu, G.; Wei, C.; Yuan, S.; Zhang, Z.; Li, J.; Zhang, L.; Wang, G.; Fang, L. Wogonoside attenuates liver fibrosis by triggering hepatic stellate cell ferroptosis through SOCS1/P53/SLC7A11 pathway. Phytother. Res. 2022, 36, 4230–4243. [Google Scholar] [CrossRef] [PubMed]
- Riccio, G.; Nuzzo, G.; Zazo, G.; Coppola, D.; Senese, G.; Romano, L.; Costantini, M.; Ruocco, N.; Bertolino, M.; Fontana, A.; et al. Bioactivity Screening of Antarctic Sponges Reveals Anticancer Activity and Potential Cell Death via Ferroptosis by Mycalols. Mar. Drugs 2021, 19, 459. [Google Scholar] [CrossRef]
- Huang, Q.; Li, J.; Ma, M.; Lv, M.; Hu, R.; Sun, J.; Zhong, X.; Sun, X.; Feng, W.; Ma, W.; et al. High-throughput screening identification of a small-molecule compound that induces ferroptosis and attenuates the invasion and migration of hepatocellular carcinoma cells by targeting the STAT3/GPX4 axis. Int. J. Oncol. 2023, 62, 42. [Google Scholar] [CrossRef]
- Tsai, Y.; Xia, C.; Sun, Z. The Inhibitory Effect of 6-Gingerol on Ubiquitin-Specific Peptidase 14 Enhances Autophagy-Dependent Ferroptosis and Anti-Tumor in vivo and in vitro. Front. Pharmacol. 2020, 11, 598555. [Google Scholar] [CrossRef]
- Yang, R.; Chen, F.; Xu, H.; Guo, Z.; Cao, C.; Zhang, H.; Zhang, C. Exploring the Mechanism of Realgar against Esophageal Cancer Based on Ferroptosis Induced by ROS-ASK1-p38 MAPK Signaling Pathway. Evid. Based Complement. Alternat. Med. 2022, 2022, 3698772. [Google Scholar] [CrossRef]
- Li, L.; Li, W.J.; Zheng, X.R.; Liu, Q.L.; Du, Q.; Lai, Y.J.; Liu, S.Q. Eriodictyol ameliorates cognitive dysfunction in APP/PS1 mice by inhibiting ferroptosis via vitamin D receptor-mediated Nrf2 activation. Mol. Med. 2022, 28, 11. [Google Scholar] [CrossRef] [PubMed]
- Chen, P.; Li, X.J.; Zhang, R.N.; Liu, S.P.; Xiang, Y.; Zhang, M.M.; Chen, X.Y.; Pan, T.; Yan, L.L.; Feng, J. Combinative treatment of β-elemene and cetuximab is sensitive to KRAS mutant colorectal cancer cells by inducing ferroptosis and inhibiting epithelial-mesenchymal transformation. Theranostics 2020, 10, 5107. [Google Scholar] [CrossRef] [PubMed]
- Song, C.Y.; Feng, M.X.; Li, L.; Wang, P.; Lu, X.; Lu, Y.Q. Tripterygium wilfordii Hook.f. ameliorates paraquat-induced lung injury by reducing oxidative stress and ferroptosis via Nrf2/HO-1 pathway. Ecotoxicol. Environ. Saf. 2023, 252, 114575. [Google Scholar] [CrossRef]
- Dai, W.; Pang, X.; Peng, W.; Zhan, X.; Chen, C.; Zhao, W.; Zeng, C.; Mei, Q.; Chen, Q.; Kuang, W.; et al. Liver Protection of a Low-Polarity Fraction from Ficus pandurata Hance, Prepared by Supercritical CO(2) Fluid Extraction, on CCl(4)-Induced Acute Liver Injury in Mice via Inhibiting Apoptosis and Ferroptosis Mediated by Strengthened Antioxidation. Molecules 2023, 28, 2078. [Google Scholar] [CrossRef] [PubMed]
- He, Q.; Yang, J.; Pan, Z.; Zhang, G.; Chen, B.; Li, S.; Xiao, J.; Tan, F.; Wang, Z.; Chen, P.; et al. Biochanin A protects against iron overload associated knee osteoarthritis via regulating iron levels and NRF2/System xc-/GPX4 axis. Biomed. Pharmacother. 2023, 157, 113915. [Google Scholar] [CrossRef]
- Liu, X.J.; Lv, Y.F.; Cui, W.Z.; Li, Y.; Liu, Y.; Xue, Y.T.; Dong, F. Icariin inhibits hypoxia/reoxygenation-induced ferroptosis of cardiomyocytes via regulation of the Nrf2/HO-1 signaling pathway. FEBS Open Bio 2021, 11, 2966–2976. [Google Scholar] [CrossRef]
- Feng, S.; Zhou, Y.; Huang, H.; Lin, Y.; Zeng, Y.; Han, S.; Huang, K.; Liu, Q.; Zhu, W.; Yuan, Z.; et al. Nobiletin Induces Ferroptosis in Human Skin Melanoma Cells Through the GSK3β-Mediated Keap1/Nrf2/HO-1 Signalling Pathway. Front. Genet. 2022, 13, 865073. [Google Scholar] [CrossRef]
- Yang, J.H.; Nguyen, C.D.; Lee, G.; Na, C.S. Insamgobonhwan Protects Neuronal Cells from Lipid ROS and Improves Deficient Cognitive Function. Antioxidants 2022, 11, 295. [Google Scholar] [CrossRef]
- Shen, Y.; Shen, X.; Wang, S.; Zhang, Y.; Wang, Y.; Ding, Y.; Shen, J.; Zhao, J.; Qin, H.; Xu, Y.; et al. Protective effects of Salvianolic acid B on rat ferroptosis in myocardial infarction through upregulating the Nrf2 signaling pathway. Int. Immunopharmacol. 2022, 112, 109257. [Google Scholar] [CrossRef]
- Zhang, J.; Wang, N.; Zhou, Y.; Wang, K.; Sun, Y.; Yan, H.; Han, W.; Wang, X.; Wei, B.; Ke, Y.; et al. Oridonin induces ferroptosis by inhibiting gamma-glutamyl cycle in TE1 cells. Phytother. Res. 2021, 35, 494–503. [Google Scholar] [CrossRef]
- Li, J.; Yang, J.; Zhu, B.; Fan, J.; Hu, Q.; Wang, L. Tectorigenin protects against unilateral ureteral obstruction by inhibiting Smad3-mediated ferroptosis and fibrosis. Phytother. Res. 2022, 36, 475–487. [Google Scholar] [CrossRef] [PubMed]
- Li, P.; Zhang, L.; Guo, Z.J.; Kang, Q.J.; Chen, C.; Liu, X.Y.; Ma, Q.T.; Zhang, J.X.; Hu, Y.J.; Wang, T. Epimedium koreanum Nakai–Induced Liver Injury—A Mechanistic Study Using Untargeted Metabolomics. Front. Pharmacol. 2022, 13, 934057. [Google Scholar] [CrossRef] [PubMed]
- Wang, D.; Liu, J.; Chen, X.; Chen, J.; Zhao, T.; Du, J.; Wang, C.; Meng, Q.; Sun, H.; Wang, F.; et al. Renal transporter OAT1 and PPAR-α pathway co-contribute to icaritin-induced nephrotoxicity. Phytother. Res. 2023, 37, 549–562. [Google Scholar] [CrossRef] [PubMed]
- Lin, J.H.; Yang, K.T.; Ting, P.C.; Luo, Y.P.; Lin, D.J.; Wang, Y.S.; Chang, J.C. Gossypol Acetic Acid Attenuates Cardiac Ischemia/Reperfusion Injury in Rats via an Antiferroptotic Mechanism. Biomolecules 2021, 11, 1667. [Google Scholar] [CrossRef]
- Jiao, H.; Yang, H.; Yan, Z.; Chen, J.; Xu, M.; Jiang, Y.; Liu, Y.; Xue, Z.; Ma, Q.; Li, X.; et al. Traditional Chinese Formula Xiaoyaosan Alleviates Depressive-Like Behavior in CUMS Mice by Regulating PEBP1-GPX4-Mediated Ferroptosis in the Hippocampus. Neuropsychiatr. Dis. Treat 2021, 17, 1001–1019. [Google Scholar] [CrossRef]
- Tang, X.; Li, X.; Zhang, D.; Han, W. Astragaloside-IV alleviates high glucose-induced ferroptosis in retinal pigment epithelial cells by disrupting the expression of miR-138-5p/Sirt1/Nrf2. Bioengineered 2022, 13, 8240–8254. [Google Scholar] [CrossRef]
- Li, J.; Lu, K.; Sun, F.; Tan, S.; Zhang, X.; Sheng, W.; Hao, W.; Liu, M.; Lv, W.; Han, W. Panaxydol attenuates ferroptosis against LPS-induced acute lung injury in mice by Keap1-Nrf2/HO-1 pathway. J. Transl. Med. 2021, 19, 96. [Google Scholar] [CrossRef]
- Hao, X.; Fan, H.; Yang, J.; Tang, J.; Zhou, J.; Zhao, Y.; Huang, L.; Xia, Y. Network Pharmacology Research and Dual-omic Analyses Reveal the Molecular Mechanism of Natural Product Nodosin Inhibiting Muscle-Invasive Bladder Cancer in Vitro and in Vivo. J. Nat. Prod. 2022, 85, 2006–2017. [Google Scholar] [CrossRef]
- Liu, X.; Tian, Y.; Yang, A.; Zhang, C.; Miao, X.; Yang, W. Antitumor Effects of Poplar Propolis on DLBCL SU-DHL-2 Cells. Foods 2023, 12, 283. [Google Scholar] [CrossRef]
- Sun, Y.; He, L.; Wang, W.; Xie, Z.; Zhang, X.; Wang, P.; Wang, L.; Yan, C.; Liu, Z.; Zhao, J.; et al. Activation of Atg7-dependent autophagy by a novel inhibitor of the Keap1-Nrf2 protein-protein interaction from Penthorum chinense Pursh. attenuates 6-hydroxydopamine-induced ferroptosis in zebrafish and dopaminergic neurons. Food Funct. 2022, 13, 7885–7900. [Google Scholar] [CrossRef]
- Shao, Y.; Sun, L.; Yang, G.; Wang, W.; Liu, X.; Du, T.; Chen, F.; Jing, X.; Cui, X. Icariin protects vertebral endplate chondrocytes against apoptosis and degeneration via activating Nrf-2/HO-1 pathway. Front. Pharmacol. 2022, 13, 937502. [Google Scholar] [CrossRef] [PubMed]
- Liu, Z.; Ma, H.; Lai, Z. Revealing the potential mechanism of Astragalus membranaceus improving prognosis of hepatocellular carcinoma by combining transcriptomics and network pharmacology. BMC Complement Med. Ther. 2021, 21, 263. [Google Scholar] [CrossRef] [PubMed]
- Ma, R.; Shimura, T.; Yin, C.; Okugawa, Y.; Kitajima, T.; Koike, Y.; Okita, Y.; Ohi, M.; Uchida, K.; Goel, A.; et al. Antitumor effects of Andrographis via ferroptosis-associated genes in gastric cancer. Oncol. Lett. 2021, 22, 523. [Google Scholar] [CrossRef] [PubMed]
- Sun, G.; Su, W.; Bao, J.; Teng, T.; Song, X.; Wang, J.; Shi, B. Dietary full-fat rice bran prevents the risk of heart ferroptosis and imbalance of energy metabolism induced by prolonged cold stimulation. Food Funct. 2023, 14, 1530–1544. [Google Scholar] [CrossRef]
- Shimura, T.; Sharma, P.; Sharma, G.G.; Banwait, J.K.; Goel, A. Enhanced anti-cancer activity of andrographis with oligomeric proanthocyanidins through activation of metabolic and ferroptosis pathways in colorectal cancer. Sci. Rep. 2021, 11, 7548. [Google Scholar] [CrossRef]
- Sharma, P.; Shimura, T.; Banwait, J.K.; Goel, A. Andrographis-mediated chemosensitization through activation of ferroptosis and suppression of β-catenin/Wnt-signaling pathways in colorectal cancer. Carcinogenesis 2020, 41, 1385–1394. [Google Scholar] [CrossRef] [PubMed]
- Xia, Y.; Yang, J.; Li, C.; Hao, X.; Fan, H.; Zhao, Y.; Tang, J.; Wan, X.; Lian, S.; Yang, J. TMT-Based Quantitative Proteomics Analysis Reveals the Panoramic Pharmacological Molecular Mechanism of β-Elemonic Acid Inhibition of Colorectal Cancer. Front. Pharmacol. 2022, 13, 830328. [Google Scholar] [CrossRef]
- Yan, W.; Wang, D.; Wan, N.; Wang, S.; Shao, C.; Zhang, H.; Zhao, Z.; Lu, W.; Tian, Y.; Ye, H.; et al. Living Cell-Target Responsive Accessibility Profiling Reveals Silibinin Targeting ACSL4 for Combating Ferroptosis. Anal. Chem. 2022, 94, 14820–14826. [Google Scholar] [CrossRef]
- Soriano-Castell, D.; Liang, Z.; Maher, P.; Currais, A. Profiling the chemical nature of anti-oxytotic/ferroptotic compounds with phenotypic screening. Free Radic. Biol. Med. 2021, 177, 313–325. [Google Scholar] [CrossRef]
- Loboda, A.; Damulewicz, M.; Pyza, E.; Jozkowicz, A.; Dulak, J. Role of Nrf2/HO-1 system in development, oxidative stress response and diseases: An evolutionarily conserved mechanism. Cell. Mol. Life Sci. 2016, 73, 3221–3247. [Google Scholar] [CrossRef]
- Chiang, S.K.; Chen, S.E.; Chang, L.C. A Dual Role of Heme Oxygenase-1 in Cancer Cells. Int. J. Mol. Sci. 2019, 20, 39. [Google Scholar] [CrossRef]
- Pan, F.; Lin, X.R.; Hao, L.P.; Wang, T.; Song, H.Z.; Wang, R. The critical role of ferroptosis in hepatocellular carcinoma. Front. Cell Dev. Biol. 2022, 10, 882571. [Google Scholar] [CrossRef]
- Pey, A.L.; Megarity, C.F.; Timson, D.J. NAD(P)H quinone oxidoreductase (NQO1): An enzyme which needs just enough mobility, in just the right places. Biosci. Rep. 2019, 39, BSR20180459. [Google Scholar] [CrossRef] [PubMed]
- Ross, D.; Siegel, D. NAD(P)H:Quinone Oxidoreductase 1 (NQO1, DT-Diaphorase), Functions and Pharmacogenetics. Methods Enzymol. 2004, 382, 115–144. [Google Scholar] [PubMed]
- Zhan, S.; Lu, L.; Pan, S.S.; Wei, X.Q.; Miao, R.Q.; Liu, X.H.; Xue, M.; Lin, X.K.; Xu, H.L. Targeting NQO1/GPX4-mediated ferroptosis by plumbagin suppresses in vitro and in vivo glioma growth. Br. J. Cancer 2022, 127, 364–376. [Google Scholar] [CrossRef] [PubMed]
- Yu, J.; Zhong, B.l.; Zhao, L.; Hou, Y.; Ai, N.n.; Lu, J.J.; Ge, W.; Chen, X.p. Fighting drug-resistant lung cancer by induction of NAD (P) H: Quinone oxidoreductase 1 (NQO1)-mediated ferroptosis. Drug Resist. Updates 2023, 70, 100977. [Google Scholar] [CrossRef] [PubMed]
- Wang, T.X.; Duan, K.L.; Huang, Z.X.; Xue, Z.A.; Liang, J.Y.; Dang, Y.J.; Zhang, A.; Xiong, Y.; Ding, C.Y.; Guan, K.L. Tanshinone functions as a coenzyme that confers gain of function of NQO1 to suppress ferroptosis. Life Sci. Alliance 2023, 6, e202201667. [Google Scholar] [CrossRef] [PubMed]
- Lin, L.; Li, X.N.; Zhu, S.D.; Long, Q.S.; Hu, Y.Z.; Zhang, L.Y.; Liu, Z.X.; Li, B.; Li, X.S. Ferroptosis-related NFE2L2 and NOX4 Genes are Potential Risk Prognostic Biomarkers and Correlated with Immunogenic Features in Glioma. Cell Biochem. Biophys. 2023, 81, 7–17. [Google Scholar] [CrossRef]
- Yang, X.P.; Yu, Y.; Wang, Z.R.; Wu, P.F.; Su, X.L.; Wu, Z.P.; Gan, J.X.; Zhang, D.K. NOX4 has the potential to be a biomarker associated with colon cancer ferroptosis and immune infiltration based on bioinformatics analysis. Front. Oncol. 2022, 12, 968043. [Google Scholar] [CrossRef] [PubMed]
- Li, D.S.; Li, Y.S. The interaction between ferroptosis and lipid metabolism in cancer. Signal Transduct. Target. Ther. 2020, 5, 108. [Google Scholar] [CrossRef]
- Liberati, A.; Altman, D.G.; Tetzlaff, J.; Mulrow, C.; Gøtzsche, P.C.; Ioannidis, J.P.; Clarke, M.; Devereaux, P.J.; Kleijnen, J.; Moher, D. The PRISMA statement for reporting systematic reviews and meta-analyses of studies that evaluate health care interventions: Explanation and elaboration. J. Clin. Epidemiol. 2009, 62, e1–e34. [Google Scholar] [CrossRef] [PubMed]
- Page, M.J.; McKenzie, J.E.; Bossuyt, P.M.; Boutron, I.; Hoffmann, T.C.; Mulrow, C.D.; Shamseer, L.; Tetzlaff, J.M.; Akl, E.A.; Brennan, S.E. The PRISMA 2020 statement: An updated guideline for reporting systematic reviews. Int. J. Surg. 2021, 88, 105906. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
{kind=link}
| Pathway | Effect on Ferroptosis | Gene Name | Uniprot ID | Protein Name | Ref. |
|---|---|---|---|---|---|
| Redox homeostasis | Inhibit | GPX4 | P36969 | Glutathione peroxidase 4 | [15,16] |
| NQO1 | P15559 | NAD(P)H Quinone Dehydrogenase 1 | [17] | ||
| Promote | NOX1 | Q9Y5S8 | NADPH oxidase 1 | [18] | |
| NOX4 | Q9NPH5 | NADPH oxidase 4 | [19] | ||
| ALOX5 | P09917 | Polyunsaturated fatty acid 5-lipoxygenase | [20] | ||
| DPP4 | Q9NPH5 | Dipeptidyl peptidase 4 | [21] | ||
| Iron homeostasis/redox homeostasis | Heme metabolism | HMOX1 | P09601 | Heme oxygenase 1 | [22,23] |
| GSH homeostasis | Inhibit | GCLC | P48506 | Glutamate-cysteine ligase catalytic subunit | [24] |
| GCLM | P48507 | Glutamate-cysteine ligase regulatory subunit | [24] | ||
| Promote | CARS | P49589 | Cysteine-tRNA ligase, cytoplasmic | [25] | |
| CHAC1 | Q9BUX1 | Glutathione-specific gamma-glutamylcyclotransferase 1 | [26] | ||
| Lipid metabolism | Inhibit | HELLS | Q9NRZ9 | Lymphoid-specific helicase | [27] |
| SCD | O00767 | Stearoyl-CoA desaturase | [20] | ||
| FADS2 | O95864 | Acyl-CoA 6-desaturase | [28] | ||
| Promote | ACSL4 | O60488 | Acyl-CoA Synthetase long chain family member 4 | [29] | |
| ACSL3 | O95573 | Acyl-CoA Synthetase long chain family member 3 | [30] | ||
| LPCAT3 | Q6P1A2 | Lysophosphatidylcholine acyltransferase 3 | [31] | ||
| CS | O75390 | Citrate synthase | [32] | ||
| Glucose metabolism | Inhibit | PHGDH | O43175 | D-3-phosphoglycerate dehydrogenase | [33] |
| G6PD | P11413 | Glucose-6-phosphate 1-dehydrogenase | [34] | ||
| Promote | PHKG2 | P15735 | Phosphorylase kinase subunit gamma-2 | [35] | |
| Mevalonate pathway | Promote | SQLE | Q14534 | Squalene monooxygenase | [36] |
| FDFT1 | P37268 | Squalene synthase | [37] |
| Enzyme | Natural Product (Compound) | Natural Product (Herb /Chinese Medicine) | Cell Line | Animal | Activity/Mechanism | Relevant Disease | Ref. |
|---|---|---|---|---|---|---|---|
| GPX4 | Baicalein (5,6,7-trihydroxyflavone) | Solanum nigrum L. | PANC1, BxPc-3 | — — | At the protein level, baicalein suppresses erastin-mediated degradation of GPX4. | Ferroptosis-associated tissue injury | [42] |
| GPX4 | Jiyuan oridonin A derivative (a2) | Jiyuan Rabdosia rubescens | HGC-27, MGC-803, BGC-823, AGS, GES1 | Gastric cancer cell line-derived xenograftmicemodels | Compound a2 decreased GPX4 expression | Gastric cancer | [43] |
| GPX4 | Artemisia santolinifolia ethanol extract (ASE) | Artemisia santolinifolia | A549 and H23 | — — | ASE decreased the GPX4 level at a significantly higher rate in ASE concentrations of 200 µg/mL. | Non-small cell lung cancer (NSCLC) | [44] |
| GPX4 | Kayadiol | Torreya nucifera | NKTCL | — — | Kayadiol decreased the expression of GPX4. | Cancer | [45] |
| GPX4 | DMOCPTL | A derivative of natural product parthenolide | MDAMB-231, SUM159, BT574,4T1, Hs578T, MDA-MB-468 | Mouse | DMOCPTL directly binding to GPX4 protein, inducing GPX4 ubiquitination. | Triple-negative breast cancer (TNBC) | [16] |
| GPX4 | — — | Guizhi Fuling Capsule (GFC) | — — | C57BL/6 mice | GFC could inhibit the expression of GPX4 at protein level. | Gynecological diseases | [46] |
| GPX4 | Isothiocyanate sulforaphane | Cruciferous vegetables | U-937, MV4-11 | — — | Isothiocyanate sulforaphane decreased GPX4 protein expression level. | Acute myeloid leukemia | [47] |
| GPX4 | Resveratrol (RSV) | — — | MIN6 | — — | RSV could increase the expression of GPX4, and decrease the expression of ACSL4. | Insulin secretion dysfunction | [48] |
| GPX4 | Arsenic | Realgar | HK2 | Mouse | Realgar reduced expression of SLC7A11 and GPX4. | Nephrotoxicity | [49] |
| GPX4 | Aristolactam I (ALI) | Aristolochia and Asarum plants | mRTECs | Wild-type C57BL/6 mice and kidney-specific Rev-erbα knockout mice with AAI-induced nephropathy | ALI treatment led to decreased GPX4 in mRNA and protein level. | Nephropathy | [50] |
| GPX4 | Heteronemin | Hippospongia sp. | HA22T/VGH, HA59T | — — | Heteronemin treatment reduced the expression of GPX4. | Hepatocellular carcinoma (HCC) | [51] |
| GPX4 | Glycyrrhetinic acid (GA) | Licorice (Glycyrrhiza glabra) | MDA-MB-231, BT-549, and MCF-10A | — — | GA down-regulated the expression of SLC7A11 at protein level; GA treatment (40 μM) significantly inhibited the activity of GPX4 but no influence on GPX4 expression. | Triple-negative breast cancer (TNBC) | [52] |
| GPX4 | Pratensein (PTS) | Trifolium pretense L. | H9c2 | — — | PTS treatment up-regulated Nrf2 expression and GPX4 expression at protein level in OGD/R-treated H9c2 cells. | Myocardial infarction (MI), myocardial ischemia-reperfusion (I/R) injury | [53] |
| GPX4 | Cryptotanshinone (CTN) | Salvia miltiorrhiza Bunge (Danshen) | A549, NCI-H520, and BAES-2B | — — | CTN can inhibit the activity of GPX4. | Lung cancer | [54] |
| GPX4 | Pentacyclic triterpenoids (PTs), including glycyrrhetinic acid (GA), ursolic acid (UA) and oleanolic acid (OA) | PTs are natural products that can be found in various plants, such as licorice, rosemary, olive, and loquat. | HCT116, HeLa, A549, L02, AT-II, and HEK293 | Nude mice with HCT116 tumor xenografts | GA treatment led the ferroptosis negative regulatory protein expression (GPX4, SLC40A1, and SLC7A11) decreased considerably. | Colorectal cancer (CRC) | [55] |
| GPX4 | Astragaloside IV (Ast-IV) | Astragalus membranaceus Bunge, a traditional Chinese herb | — — | C57BL/6J male mice, lung injury model induced by PM2.5 | The Ast-IV intervention increased the expression of GPX4, which was decreased in PM2.5 group. | Lung injury caused by PM2.5 exposure | [56] |
| GPX4 | Wogonoside, wogonin, palmatine, paeoniforin and liquiritin | Shaoyao Decoction (SYD) | Caco-2 | Sprague–Dawley male rats with TNBS-induced colitis model | SYD induced activation of GPX4 transcription and increase of protein expression. | Inflammatory bowel disease (IBD), especially ulcerative colitis (UC) | [57] |
| GPX4 | Polyphenols sourced from Ilex latifolia Thunb. (PIT) | Ilex latifolia Thunb. | Porcine intestinal epithelial cells (IPEC-J2) | Weanling piglets under oxidative stress induced by diquat injection | Supplementation with PIT significantly reduced jejunal TFR1 gene expressions and improved SLC7A11 and GPX4 gene expressions in the piglets under oxidative stress. | Intestinal injury caused by oxidative stress | [58] |
| GPX4 | Danshensu (Dan) | Dan is a pure molecule derived from the root of the Salvia miltiorrhiza herb, Danshen. | LX-2 human and T6 rat hepatic stellate cells (HSCs) | — — | Dan up-regulated GPX4, and SLC7A11. | Liver fibrosis and cirrhosis | [59] |
| GPX4 | — — | Naotaifang extract (NTE): a compound traditional Chinese herbal medicine composed of four herbs: Radix Astragali (Huangqi), Rhizoma chuanxiong (Chuangxiong), Pheretima (Dilong), and Bombyx batryticatus (Jiangcan) | — — | Sprague-Dawley (SD) rats with middle cerebral artery occlusion (MCAO) model of acute cerebral ischemia | NTE increased the expression levels of SLC7A11, GPX4. | Ischemic stroke | [60] |
| GPX4 | Cucurbitacin B (CuB) | Trichosanthes kirilowii Maximowicz | Human nasopharyngeal carcinoma CNE1 cells | BALB/c nude mice, human nasopharyngeal carcinoma murine xenograft model | CuB induced ferroptosis by increasing lipid peroxidation and reducing the expression of GPX4 in mRNA and protein level. | Nasopharyngeal carcinoma | [61] |
| GPX4 | Dihydroartemisinin (DHA) and Chlorin e6 (Ce6) | DHA derived from the natural plant Artemisia annua, is a sesquiterpene lactone compound | Lewis cells (LLC), a lung cancer cell model | — — | DHA could reduce GPX4 at mRNA and protein level. | Lung cancer | [62] |
| GPX4 | Perillaldehyde | Ammodaucus leucotrichus Coss. and Dur (A. leucotrichus), commonly known as hairy cumin | HL-60, Jurkat, DLD-1, SHSY5Y | — — | Perillaldehyde decreased GPX4 protein expression. | Acute myeloid leukemia | [63] |
| GPX4 | Lycium barbarum polysaccharide (LBP) | The fruits of the traditional Chinese herb L. barbarum. | MCF-7, MDA-MB-231 | — — | LBP down-regulated the protein level of xCT (SLC7A11) and GPX4. | Breast cancer | [64] |
| GPX4 | Matrine | Sophora flavescens | HCT116 | — — | Matrine down-regulated the protein level of GPX4. | Colorectal cancer (CRC) | [65] |
| GPX4 | Wogonoside (WG) | Scutellaria baicalensis Georgi, a perennial herb of the Labiatae family | HSC-T6, AML-12, RAW264.7 | C57BL/6 mice, induced by CCl4 for liver fibrosis model | WG down-regulated the protein levels of GPX4 and SLC7A11. | Liver fibrosis | [66] |
| GPX4 | Suberitenones A and B, mycalols | Hemimycale topsenti, Haliclona (Rhizoniera) dancoi | HepG2, A549, A2058, MRC5 | — — | Mycalols could reduce the level of GPX4 protein in HepG2 cells. | Hepatocellular carcinoma, lung carcinoma, melanoma, anaplastic thyroid carcinoma | [67] |
| GPX4 | Polyphyllin VI (PPVI) | Paris polyphylla | HCCLM3, Huh7 and THLE-2 | Male BALB/c nude mice with subcutaneous tumor model of Huh7 cells | The GPX4 protein expression levels in the HCCLM3 and Huh7 cells were down-regulated, and negatively associated with the concentration of PPVI. | Hepatocellular carcinoma (HCC) | [68] |
| GPX4 | 6-Gingerol | Ginger (Zingiber officinale Roscoe) | A549 | BALB/cNude mice, A549 tumor xenografts | 6-Gingerol down-regulate the expressions of GPX4 at protein level. | Lung cancer | [69] |
| GPX4 | Realgar (REA) | — — | Eca109, KYSE150 | — — | REA IC50 and 2IC50 caused significant down-regulation of GPX4. | Esophageal cancer | [70] |
| GPX4, HO-1 | Eriodictyol | Citrus fruits and some Chinese herbal medicines | HT-22 | APPswe/PS1E9 transgenic mice, a mouse model of Alzheimer’s disease | Eriodictyol treatment could increase the GPX4, and HO-1 at protein level in both the cortex and hippocampus of APP/PS1 mice. | Alzheimer’s disease (AD) | [71] |
| GPX4, HO-1 | β-elemene | Curcumae rhizoma | HCT116, LoVo, CaCO2 | β-elemene in combination with cetuximab was shown to upregulate the HO-1 and down-regulate GPX4. | Colorectal cancer | [72] | |
| GPX4, HO-1 | — — | Tripterygium wilfordii Hook.f. (TwHF) | — — | APPswe/PS1E9 transgenic mice, a mouse model of Alzheimer’s disease | TwHF treatment reduced the PQ up-regulated HO-1 level, and it up-regulated the expression of GPX4 compared to the PQ group. | Alzheimer’s disease (AD) | [73] |
| GPX4, HO-1 | FPHLP (low-polarity fraction from Ficus pandurata Hance) | Ficus pandurata Hance | HepG2 | C57BL6/J mice, CCl4-induced acute liver injury model | FPHLP significantly reduced the level of Fe2+ and expression of TfR1, xCT/SLC7A11, and Bcl2, while increasing the expression of GPX4. | Acute liver injury | [74] |
| GPX4, HO-1 | Biochanin A (BCA) | Huangqi | — — | C57BL/6 mice | The tendency of HO-1, and GPX4 expression was downregulated when treated with ferric ammonium citrate and upregulated after incubation with BCA. | knee osteoarthritis | [75] |
| GPX4, HO-1, ACSL4 | Icariin (ICA) | Epimedium | H9C2 | — — | The mRNA expressions and protein levels of Nrf2 and HO-1 were reduced notably after H/R stimulation but were elevated with the increase of ICA concentration. The expression of ACSL4 were decreased with the increase of ICA concentration, while the expression of GPX4 was increased with the increase of ICA concentration. | Myocardial ischemia /reperfusion injury | [76] |
| GPX4, HO-1 | Nobiletin | Citrus peel | SK-MEL-28 | — — | The mRNA and protein level of GPX4 and HO-1 was decreased in nobiletin-treated melanoma cells. | Melanoma | [77] |
| GPX4, HO-1 | — — | GBH (Insamgobonhwan) | HT22 | ICR mouse | GBH can reverse the RSL3 induced GPX4 reduction and HO-1 increase. | Alzheimer’s disease (AD) | [78] |
| GPX4, HO-1 | Salvianolic acid B(Sal B) | Salvia miltiorrhiza (Danshen in Chinese) | H9C2 | Sprague-Dawley rats, myocardial infarction model | Nrf2 was strongly activated in MI rats pretreated with Sal B, and resulted in the upregulation of its target genes including HO-1, xCT, Gpx4, Fth1, and Fpn1. | Myocardial infarction (MI) | [79] |
| GPX4, GCLC | Oridonin (Ori) | Rabdosia rubescens (Hemsl.) | TE1 | — — | Ori can decrease the enzymatic activity of GCLC and GPX4. | Esophageal cancer | [80] |
| GPX4, NOX4 | Tectorigenin | Belamcanda chinensis | Primary renal tubular epithelial cells | — — | Tectorigenin treatment greatly inhibited Smad3 phosphorylation, and the transcription and protein level of Nox4; while it indirectly restored the expression of GPX4. | Chronic kidney disease | [81] |
| GPX4, ACSL4 | — — | Epimedium koreanum Nakai | — — | Rat | Epimedium koreanum Nakai ethanol extract (EEE) downregulated GPX4, and up-regulated ACSL4 significantly. | Toxicity-natural products-induced liver injury | [82] |
| GPX4, ACSL3, ACSL4 | Icaritin | Epimedium, a traditional Chinese medicine | HK-2 cells, hOAT1-HEK293 cells, and mock-HEK293 cells | C57BL/6 mice, with icaritin-induced nephrotoxicity model | Icaritin significantly downregulated the protein expression of ACSL3, and ACSL4, and it can reduce the activity of GPX4. | Icaritin-induced nephrotoxicity | [83] |
| GPX4, ACSL4 | Gossypol acetic acid (GAA) | The seeds of cotton plants | H9C2 cardiomyoblast cells and neonatal rat cardiomyocytes | Sprague-Dawley | GAA significantly decreased the mRNA levels of Ptgs2 and Acsl4, decreased the protein levels of ACSL4 and NRF2, and increased the protein levels of GPX4 in I/R-induced ex vivo rat hearts. | Cardiac Ischmia/Reperfusion Injury | [84] |
| GPX4, ACSL4 | — — | Xiaoyaosan: Radix Angelicae sinensis, Radix Paeoniae alba, Poria, Radix bupleuri, Rhizoma Atractylodis macrocephalae, Radix glycyrrhizae, Herba menthae, Rhizoma Zingiberis recens | — — | C57BL/6 | The Xiaoyaosan treatment could upregulate the stress-induced down-regulation of GPX4; and down-regulate the stress-induced up-regulation of ACSL4. | Depressive-like behavior | [85] |
| GPX4, GCLC, GCLM | Astragaloside-IV (AS-IV) | Astragalus membranaceus | ARPE-19 cells | — — | AS-IV could restore the down-regulated expression of GPX4, GCLM, and GCLC under high glucose conditions. | Diabetic retinopathy (DR) | [86] |
| HO-1 | Panaxydol (PX) | Panax ginseng | BEAS-2B | Specific pathogen-free (SPF) male C57BL/6 mice, murine model of LPS-induced ALI. | PX increased Nrf2 and HO-1 expression in comparison with LPS group. | Acute lung injury (ALI)/acute respiratory distress syndrome (ARDS) | [87] |
| HO-1 | Tagitinin C | Tithonia diversifolia | HCT116 | — — | Tagitinin C increased mRNA expression level of HO-1 | Colorectal cancer | [22] |
| HO-1 | Nodosin | Isodon serra (Maxim.) Kudo | T24, UMUC3, SW780 | Xenograft tumor model | Nodosin up-regulated the expression of genes, including HMOX1, G0S2, SQSTM1, FTL and AIFM2 | Muscle-invasive bladder cancer (MIBC) | [50,88] |
| HO-1 | — — | Propolis | SU-DHL-2 | — — | The expressions of HO-1 were up-regulated at mRNA level when treated by propolis. | Diffuse large B-cell lymphoma (DLBCL) | [89] |
| HO-1 | Thonningianin A (Th A) | Penthorum chinense Pursh. | SH-SY5Y | Zebrafish (AB strain of wild-type zebrafish); 6-OHDA-induced zebrafish model of Parkinson’s disease | Th A treatment significantly facilitated the Nrf2 nuclear translocation and subsequently increased the HO-1 protein level. | Parkinson’s disease (PD) | [90] |
| HO-1 | Icariin (ICA) | Herba epimedii | Endplate chondrocytes, nucleus pulposus cells, annulus fibrosus cells | C57BL/6J | ICA treatment led to notably increased protein levels of Nrf-2 and its downstream HO-1. | Intervertebral disc degeneration | [91] |
| HO-1 | Astragalus membranaceus, Astragaloside IV, Astragalus polysaccharide, Swainsonine, Daidzein | Astragalus membranaceus, a kind of traditional Chinese medicine | HepG2 | — — | miRNA level of HMOX1 can be up-regulated by Astragalus membranaceus. | Hepatocellular carcinoma (HCC) | [92] |
| HO-1, GCLC, GCLM | Andrographolide | Andrographis paniculata, a traditional Chinese herbal medicine | MKN74 and NUGC4 | — — | Andrographis treatment upregulated the expression of HMOX1, GCLC, and GCLM | Gastric cancer | [93] |
| HO-1, NQO1 | Full-fat rice bran | A by-product of rice processing. | — — | Yorkshire pigs were used as experimental animals to establish a prolonged cold stimulation model. | Full-fat rice bran promoted the mRNA expression of Nrf2 and NQO1, as well as the protein content of Nrf2 and HO-1. | Cardiac injury and energy metabolism disturbance caused by prolonged cold stimulation. | [94] |
| HO-1, GCLC, GCLM | Andrographolide | Andrographis paniculata, a traditional herb | HCT116, HT29, NCM460 | athymic nude mice, subcutaneous xenograft model | HMOX1, GCLC, and GCLM were significantly up-regulated in the andrographis and the combination treatment group vs. the untreated group. | Colorectal cancer (CRC) | [95] |
| HO-1, GCLC, GCLM | Andrographis extract (standardized to 20% andrographolide content) | Andrographis paniculata | HCT116 and SW480 | Mouse | Andrographis could upregulate the HMOX1, GCLC, and GCLM, in mRNA and protein level individually or in combination with 5FU. | Colorectal cancer (CRC) | [96] |
| ACSL4 | β-Elemonic acid (EA) | EA is isolated from Boswellia papyrifera, a plant used in traditional medicine. | CRC lines SW480, HCT116 and HT29; the control colorectal cell line NCM460 | Female BALB/c nude mice; subcutaneous xenograft model of CRC | EA at high concentration induces ferroptosis by downregulating FTL and upregulating TF, CP, and ACSL4. | Colorectal cancer (CRC) | [97] |
| ACSL4 | Silibinin (SIL) | Silybum marianum | HepG2, HEK293T, Hep1-6 | — — | The enzymatic assays showed SIL inhibited ACSL4 enzymatic activity, thereby mitigating the ACSL4-mediated ferroptosis. | Liver diseases | [98] |
| NQO1 | Quinones: Cryptotanshinone, idebenone, menaquinone 4, dopamine, lipoic acid, ascorbic acid, baicalein, 3,3′-diindolylmethane, fisetin, triptophenolide, cynaroside (luteoloside), icaritin, gossypol, carnosol, honokiol, β-lapachone, dihydroisotanshinone I (DHIT I), dihydrotanshinone I (DHT I), tanshinone I and cryptotanshinone. | — — | HT22, MC65 | — — | All quinones that were substrates of these proteins as identified by the NADH decay assay displayed an increase in anti-lipid peroxidation activity in the liposomes in the presence of NQO1 or FSP1. | Alzheimer’s disease (AD) | [99] |
| No. | Eligibility Criteria |
|---|---|
| 1 | Not included article types. e.g., review, proceedings, feature, editorial material. |
| 2 | Not in the life Sciences |
| 3 | Irrelevant object/topic |
| 4 | Full text not available |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zuo, H.-L.; Huang, H.-Y.; Lin, Y.-C.-D.; Liu, K.-M.; Lin, T.-S.; Wang, Y.-B.; Huang, H.-D. Effects of Natural Products on Enzymes Involved in Ferroptosis: Regulation and Implications. Molecules 2023, 28, 7929. https://doi.org/10.3390/molecules28237929
Zuo H-L, Huang H-Y, Lin Y-C-D, Liu K-M, Lin T-S, Wang Y-B, Huang H-D. Effects of Natural Products on Enzymes Involved in Ferroptosis: Regulation and Implications. Molecules. 2023; 28(23):7929. https://doi.org/10.3390/molecules28237929
Chicago/Turabian StyleZuo, Hua-Li, Hsi-Yuan Huang, Yang-Chi-Dung Lin, Kun-Meng Liu, Ting-Syuan Lin, Yi-Bing Wang, and Hsien-Da Huang. 2023. "Effects of Natural Products on Enzymes Involved in Ferroptosis: Regulation and Implications" Molecules 28, no. 23: 7929. https://doi.org/10.3390/molecules28237929
APA StyleZuo, H.-L., Huang, H.-Y., Lin, Y.-C.-D., Liu, K.-M., Lin, T.-S., Wang, Y.-B., & Huang, H.-D. (2023). Effects of Natural Products on Enzymes Involved in Ferroptosis: Regulation and Implications. Molecules, 28(23), 7929. https://doi.org/10.3390/molecules28237929