
Silencing LY6D Expression Inhibits Colon Cancer in Xenograft Mice and Regulates Colon Cancer Stem Cells’ Proliferation, Stemness, Invasion, and Apoptosis via the MAPK Pathway

 , , and
, , and {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
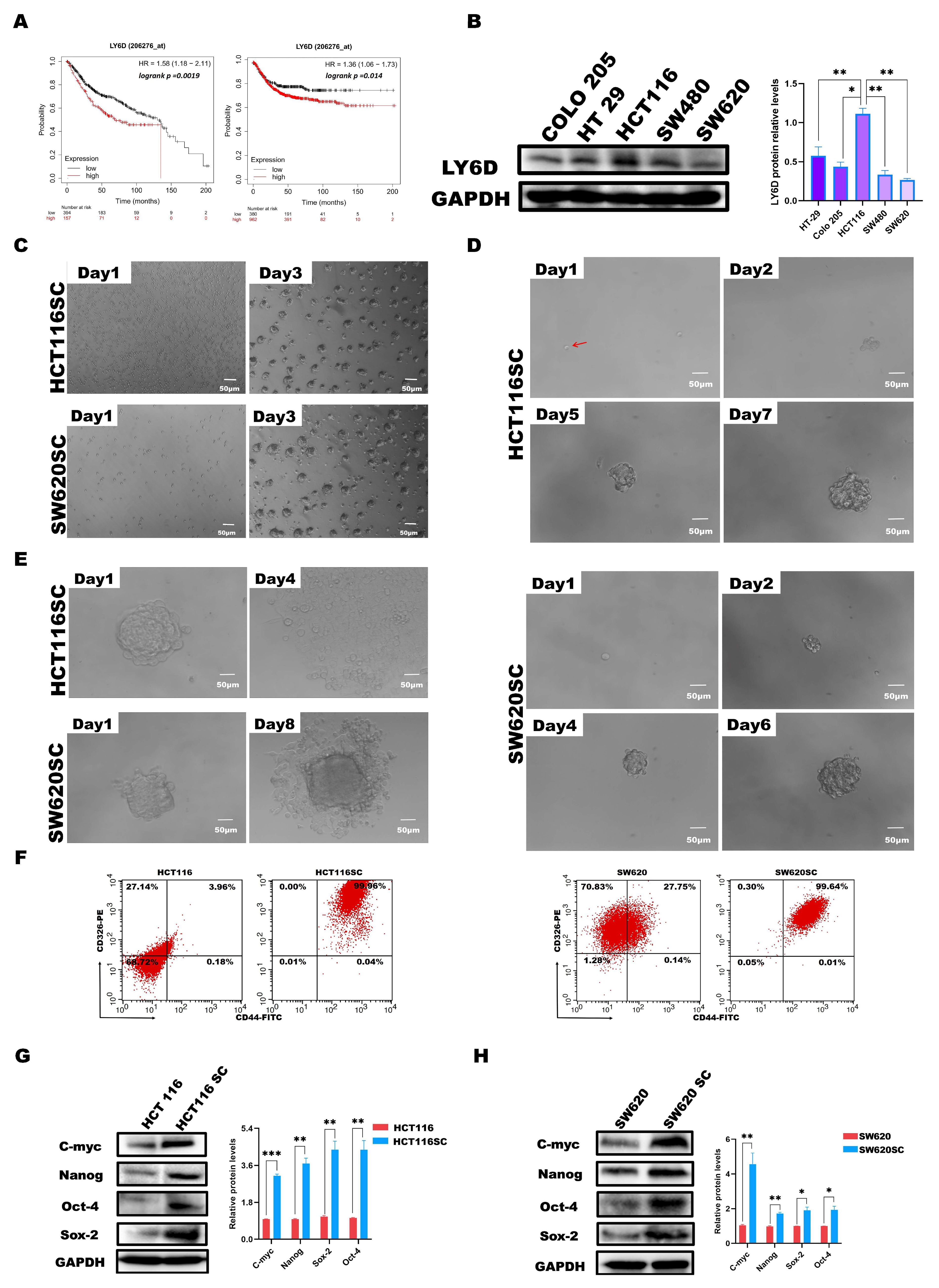
2.1. LY6D Expression in CC and CCSCs’ Screening and Identification
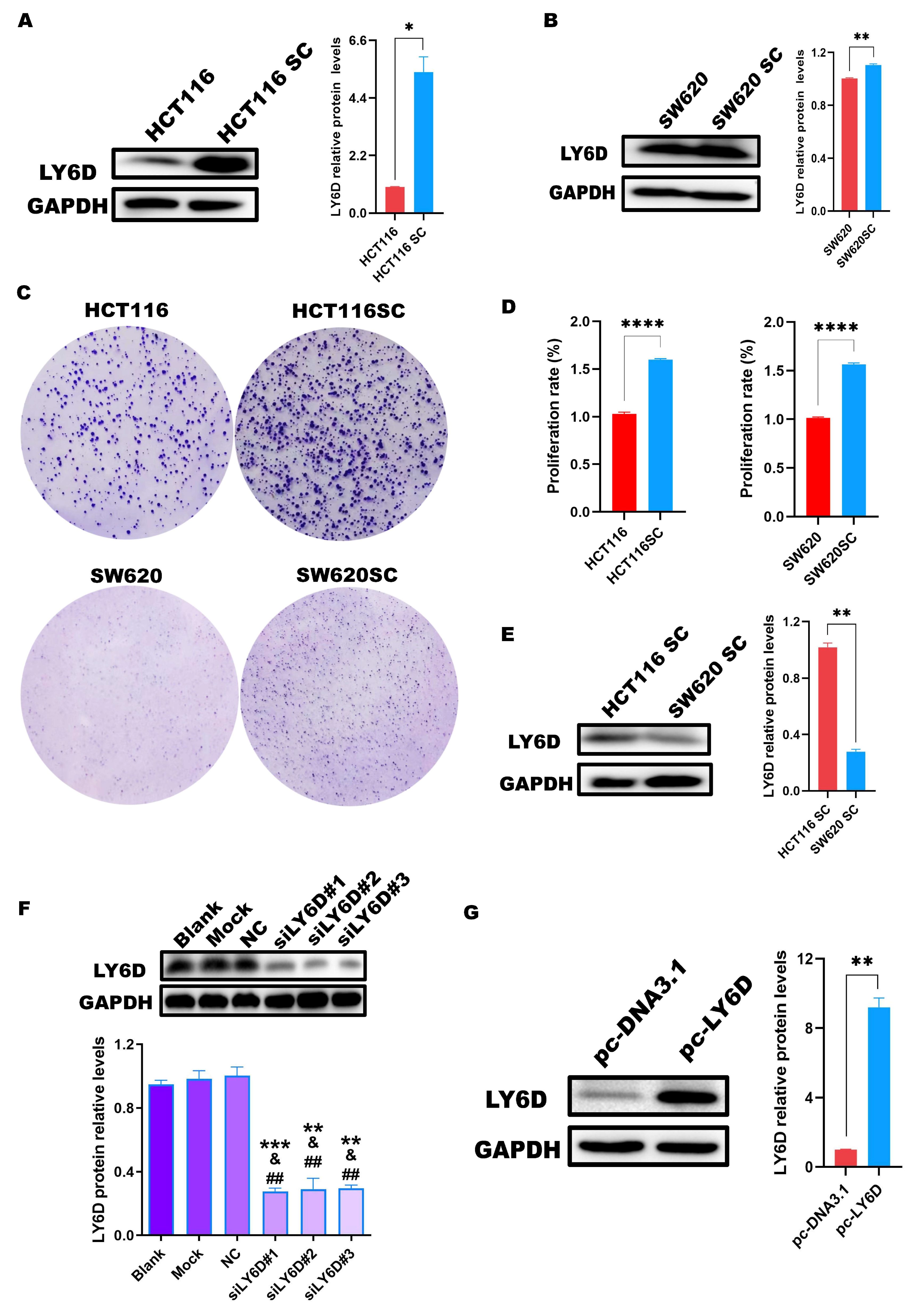
2.2. LY6D Was Expressed at High Levels in CCSCs
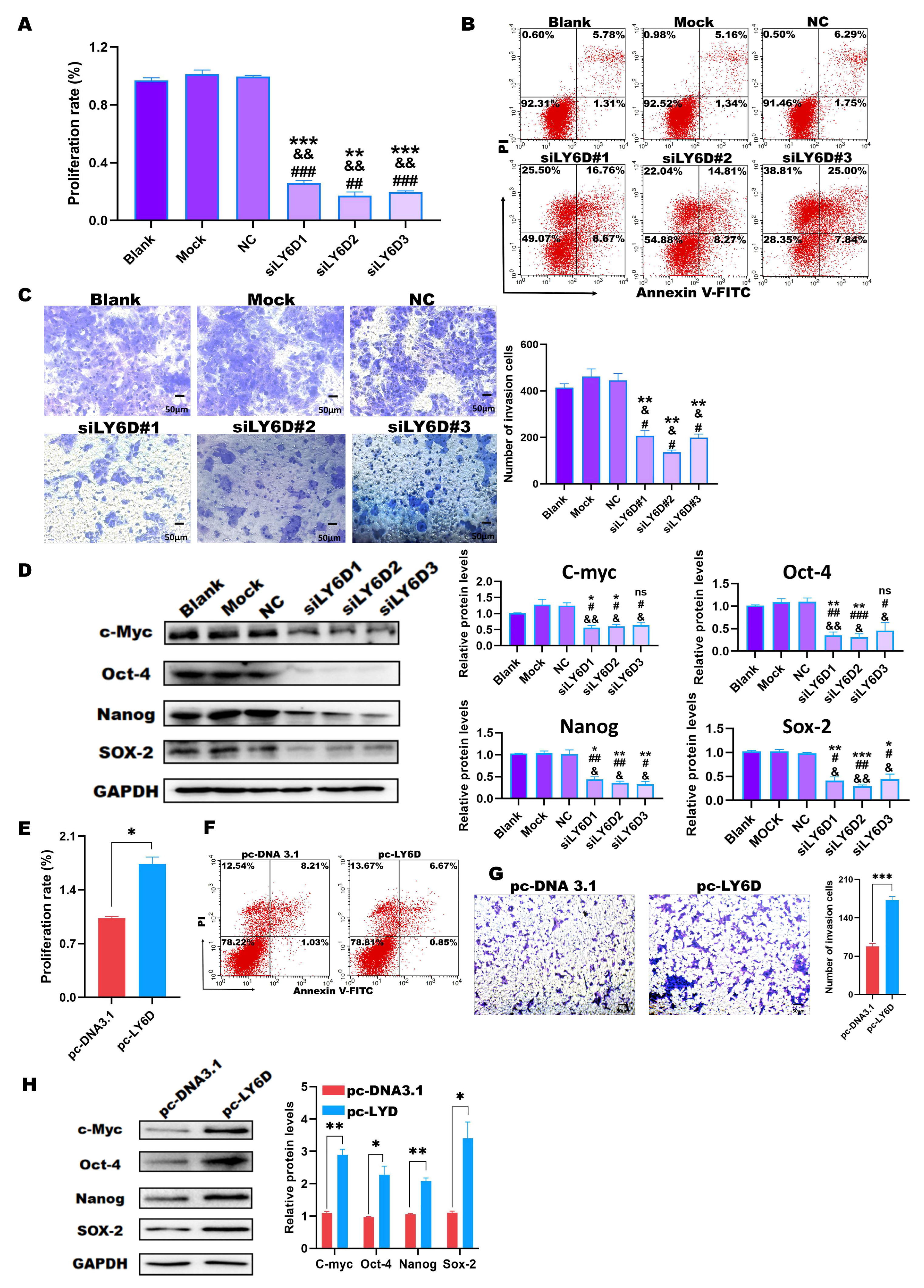
2.3. Down-Regulation of LY6D Expression Reduced HCT116SCs’ Proliferation, Invasion, and Stemness and Increased Apoptosis
2.4. Up-Regulation of LY6D Expression Increased the Proliferation, Invasion, and Stemness of SW620SCs
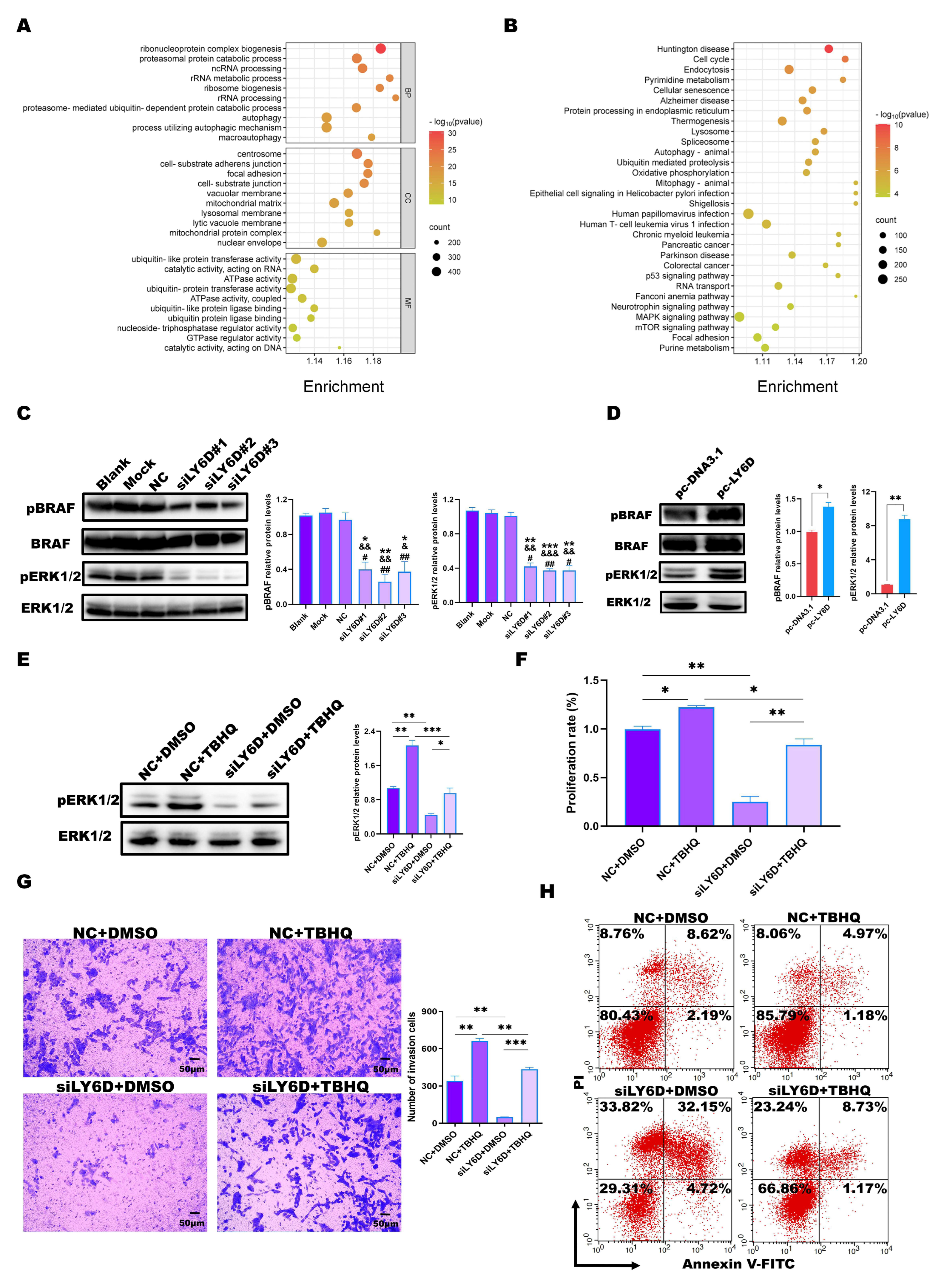
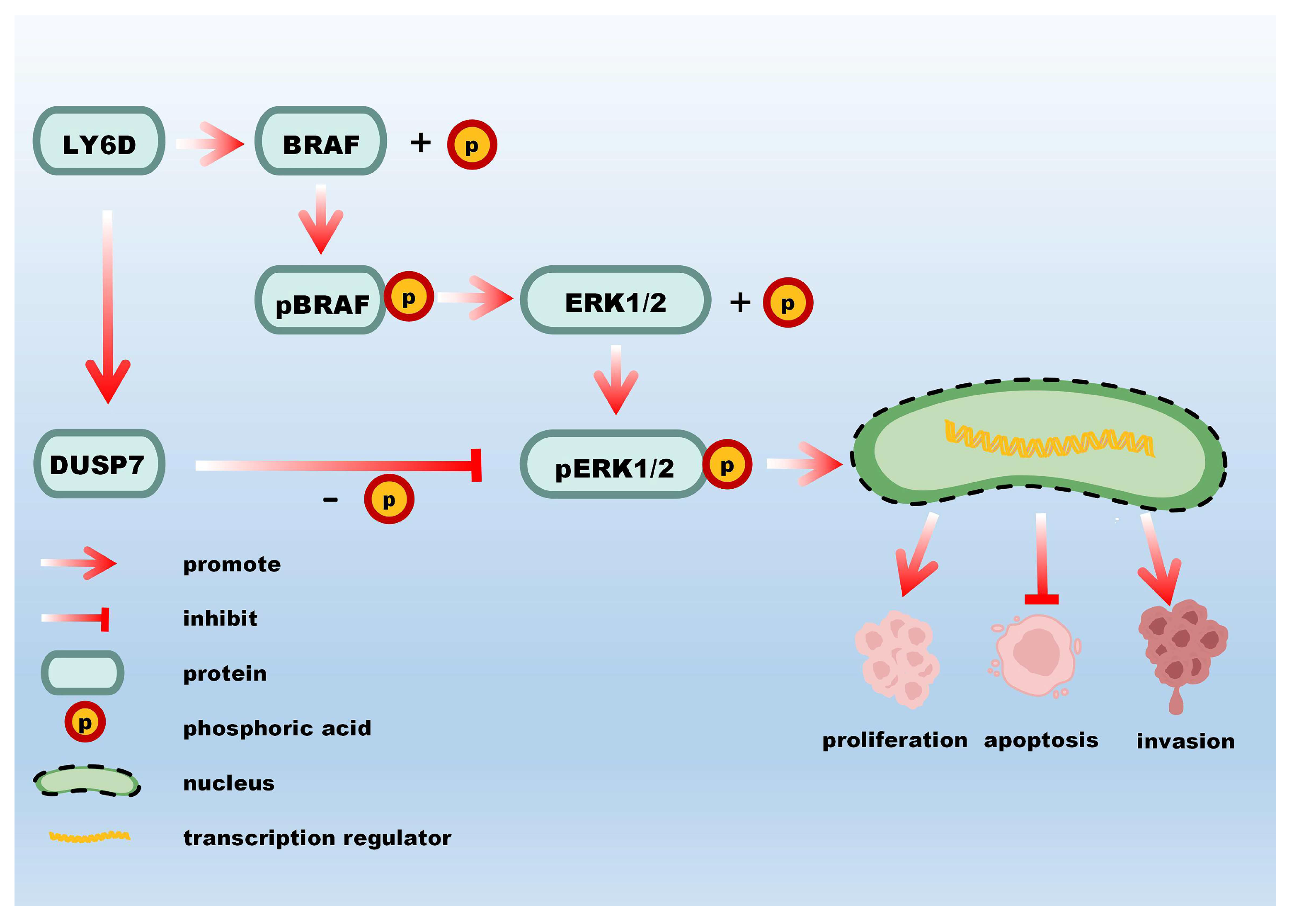
2.5. LY6D Regulated CCSCs’ Proliferation, Apoptosis, and Invasion via the MAPK Pathway
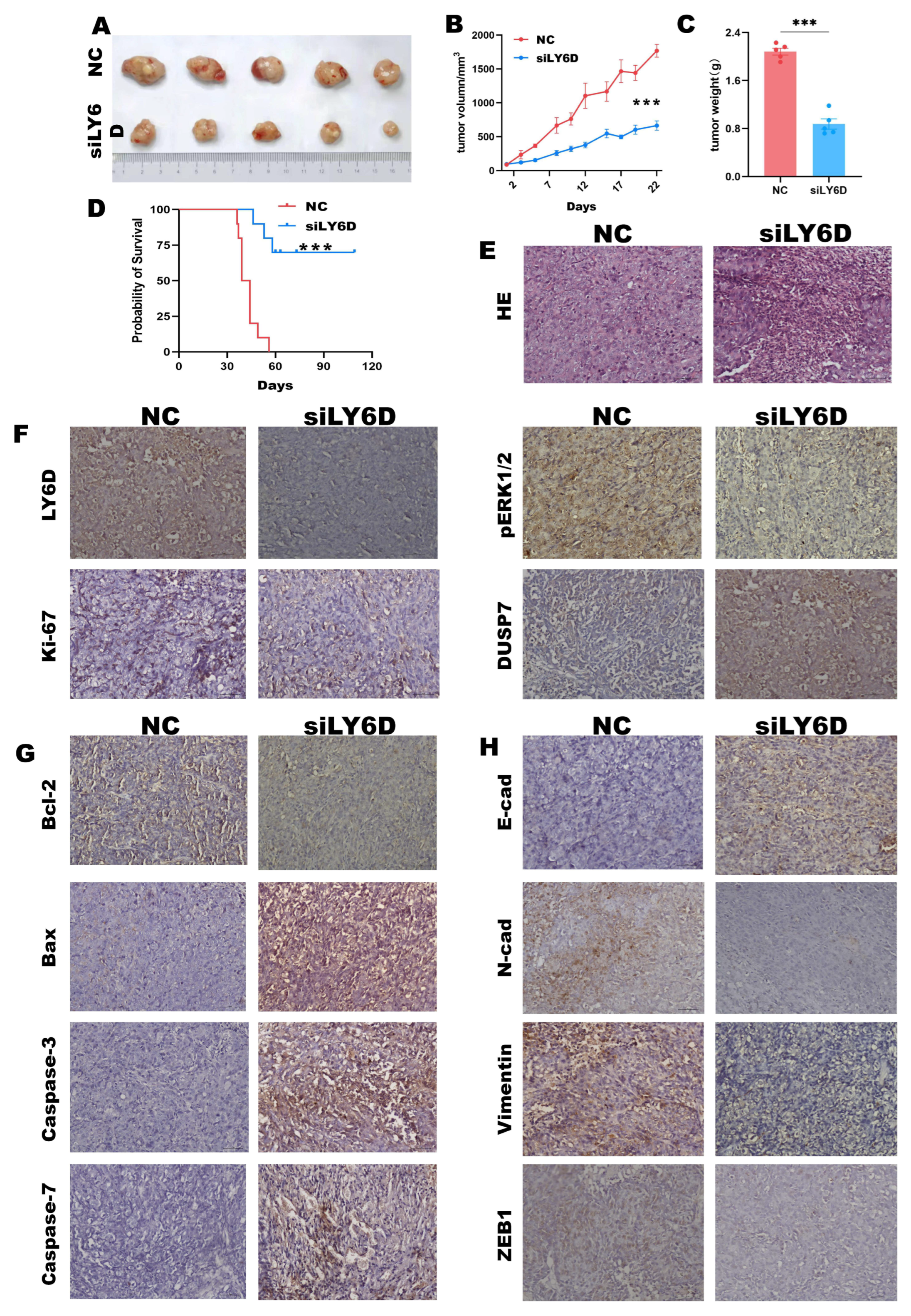
2.6. siLY6D-Mediated Inhibition of CC Xenograft Tumor Growth
3. Discussion
4. Materials and Method
4.1. Cell Lines and Cell Culture
4.1.1. CC Cell Lines
4.1.2. CCSCs
4.2. Cell Sorting
4.3. Cell Transfection
4.4. Cell Proliferation
4.5. Colony Formation Assay
4.6. Tert-Butyl Hydroquinone (TBHQ) Response Assay
4.7. Cell Apoptosis
4.8. Cell Invasion
4.9. RNA-Seq and Bioinformatics Analysis
4.9.1. RNA-Seq
4.9.2. GO and KEGG Enrichment Analysis
4.10. Western Blotting
4.11. Animals
4.12. Xenograft Assay
4.13. Hematoxylin-Eosin (HE) and Immunohistochemistry (IHC) Staining
4.14. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
References
- Sung, H.; Ferlay, J.; Siegel, R.L.; Laversanne, M.; Soerjomataram, I.; Jemal, A.; Bray, F.; Bsc, M.F.B.; Me, J.F.; Soerjomataram, M.I.; et al. Global cancer statistics 2020: GLOBOCAN estimates of incidence and mortality worldwide for 36 cancers in 185 countries. CA A Cancer J. Clin. 2021, 71, 209–249. [Google Scholar] [CrossRef] [PubMed]
- Uchino, T.; Ouchi, A.; Komori, K.; Kinoshita, T.; Oshiro, T.; Sano, T.; Shimizu, Y. The prognostic relevance of primary tumor sidedness to surgical treatment for recurrent colon cancer. Surg. Today 2020, 51, 94–100. [Google Scholar] [CrossRef]
- Spinoglio, G.; Bianchi, P.P.; Marano, A.; Priora, F.; Lenti, L.M.; Ravazzoni, F.; Petz, W.; Borin, S.; Ribero, D.; Formisano, G.; et al. Robotic Versus Laparoscopic Right Colectomy with Complete Mesocolic Excision for the Treatment of Colon Cancer: Perioperative Outcomes and 5-Year Survival in a Consecutive Series of 202 Patients. Ann. Surg. Oncol. 2018, 25, 3580–3586. [Google Scholar] [CrossRef] [PubMed]
- Xiong, W.; Qin, J.; Cai, X.; Xiong, W.; Liu, Q.; Li, C.; Ju, Y.; Wang, Q.; Li, Y.; Yang, Y. Overexpression LINC01082 suppresses the proliferation, migration and invasion of colon cancer. Mol. Cell Biochem. 2019, 462, 33–40. [Google Scholar] [CrossRef] [PubMed]
- O’Keefe, S.J.D. Diet, microorganisms and their metabolites, and colon cancer. Nat. Rev. Gastroenterol. Hepatol. 2016, 13, 691–706. [Google Scholar] [CrossRef]
- Guo, Q.; Zhou, Y.; Xie, T.; Yuan, Y.; Li, H.; Shi, W.; Zheng, L.; Li, X.; Zhang, W. Tumor microenvironment of cancer stem cells: Perspectives on cancer stem cell targeting. Genes Dis. 2023. [Google Scholar] [CrossRef]
- Chen, X.; Tang, K.; Li, X.; Zhang, C.; Xin, Y.; Li, K.; Tan, Y. Biomechanics of cancer stem cells. Essays Biochem. 2022, 66, 359–369. [Google Scholar]
- Bocci, F.; Gearhart-Serna, L.; Boareto, M.; Ribeiro, M.; Ben-Jacob, E.; Devi, G.R.; Levine, H.; Onuchic, J.N.; Jolly, M.K. Toward understanding cancer stem cell heterogeneity in the tumor microenvironment. Proc. Natl. Acad. Sci. USA 2018, 116, 148–157. [Google Scholar] [CrossRef]
- Li, J.; Zhou, Q.; Liu, L.; He, J. Effects of SMC1A on immune microenvironment and cancer stem cells in colon adenocarcinoma. Cancer Med. 2023, 12, 12881–12895. [Google Scholar] [CrossRef]
- Ibragimova, M.; Tsyganov, M.; Litviakov, N. Tumour Stem Cells in Breast Cancer. Int. J. Mol. Sci. 2022, 23, 5058. [Google Scholar] [CrossRef]
- Rowbotham, S.P.; Goruganthu, M.U.L.; Arasada, R.R.; Wang, W.Z.; Carbone, D.P.; Kim, C.F. Lung Cancer Stem Cells and Their Clinical Implications. Cold Spring Harb. Perspect. Med. 2022, 12, a041270. [Google Scholar] [CrossRef] [PubMed]
- Zhou, L.; Ma, S. Deciphering cancer stem cells in liver cancers: New tools with improved resolution. Carcinogenesis 2022, 43, 297–300. [Google Scholar] [CrossRef]
- Banerjee, P.; Kapse, P.; Siddique, S.; Kundu, M.; Choudhari, J.; Mohanty, V.; Malhotra, D.; Gosavi, S.W.; Gacche, R.N.; Kundu, G.C. Therapeutic implications of cancer stem cells in prostate cancer. Cancer Biol. Med. 2023, 20, 401–420. [Google Scholar] [CrossRef] [PubMed]
- Acikgoz, E.; Tatar, C.; Oktem, G. Triptolide inhibits CD133+/CD44+ colon cancer stem cell growth and migration through triggering apoptosis and represses epithelial-mesenchymal transition via downregulating expressions of snail, slug, and twist. J. Cell. Biochem. 2020, 121, 3313–3324. [Google Scholar] [CrossRef] [PubMed]
- Yasodha, K.; Mary, L.L.; Surajit, P.; Satish, R. Exosomes from metastatic colon cancer cells drive the proliferation and migration of primary colon cancer through increased expression of cancer stem cell markers CD133 and DCLK1. Tissue Cell 2023, 84, 102163. [Google Scholar] [CrossRef] [PubMed]
- McKenzie, I.F.; Gardiner, J.; Cherry, M.; Snell, G.D. Lymphocyte antigens: Ly-4, Ly-6, and Ly-7. Transpl. Proc. 1977, 9, 667–669. [Google Scholar]
- Loughner, C.L.; Bruford, E.A.; McAndrews, M.S.; Delp, E.E.; Swamynathan, S.; Swamynathan, S.K. Organization, evolution and functions of the human and mouse Ly6/uPAR family genes. Hum. Genom. 2016, 10, 19. [Google Scholar] [CrossRef]
- Gruet, M.; Cotton, D.; Coveney, C.; Boocock, D.J.; Wagner, S.; Komorowski, L.; Rees, R.C.; Pockley, A.G.; Garner, A.C.; Wallis, J.D. β2-Adrenergic Signalling Promotes Cell Migration by Upregulating Expression of the Metastasis-Associated Molecule LYPD3. Biology 2020, 9, 39. [Google Scholar] [CrossRef] [PubMed]
- Leth, J.M.; Leth-Espensen, K.Z.; Kristensen, K.K.; Kumari, A.; Winther, A.-M.L.; Young, S.G.; Ploug, M. Evolution and Medical Significance of LU Domain-Containing Proteins. Int. J. Mol. Sci. 2019, 20, 2760. [Google Scholar] [CrossRef]
- Miwa, J.M. Lynx1 prototoxins: Critical accessory proteins of neuronal nicotinic acetylcholine receptors. Curr. Opin. Pharmacol. 2020, 56, 46–51. [Google Scholar] [CrossRef]
- Shelukhina, I.; Siniavin, A.; Kasheverov, I.; Ojomoko, L.; Tsetlin, V.; Utkin, Y. α7- and α9-Containing Nicotinic Acetylcholine Receptors in the Functioning of Immune System and in Pain. Int. J. Mol. Sci. 2023, 24, 6524. [Google Scholar] [CrossRef] [PubMed]
- Shmerling, M.; Chalik, M.; Smorodinsky, N.I.; Meeker, A.; Roy, S.; Sagi-Assif, O.; Meshel, T.; Danilevsky, A.; Shomron, N.; Levinger, S.; et al. LY6S, a New IFN-Inducible Human Member of the Ly6a Subfamily Expressed by Spleen Cells and Associated with Inflammation and Viral Resistance. ImmunoHorizons 2022, 6, 253–272. [Google Scholar] [CrossRef]
- Jung, H. LY6E (Lymphocyte Antigen 6 Complex Locus) is Associated with Invasion and Poor Prognosis in Colorectal Cancer. Ann. Surg. Oncol. 2017, 24, S89. [Google Scholar]
- Kim, H.J.; Hong, I.; Roh, S.; Kim, S.; Kim, H.; Oh, S.; Ahn, T.S.; Kang, D.H.; Baek, M.-J.; Jeong, D. High expression of LY6E is an independent prognostic factor of colorectal cancer patients. Oncol. Rep. 2023, 49, 80. [Google Scholar] [CrossRef] [PubMed]
- Geng, L.; Wang, Z.; Tian, Y. Down-regulation of ZNF252P-AS1 alleviates ovarian cancer progression by binding miR-324-3p to downregulate LY6K. J. Ovarian Res. 2022, 15, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Kong, H.K.; Son, D.S.; Park, J.H. The regulatory mechanism of human LY6K related to carcinogenesis and metastasis in breast cancer. Cancer Res. 2016, 76, 2926. [Google Scholar] [CrossRef]
- Kong, H.K.; Yoon, S.; Park, J.H. The Regulatory Mechanism of the LY6K Gene Expression in Human Breast Cancer Cells. J. Biol. Chem. 2012, 287, 38889–38900. [Google Scholar] [CrossRef]
- Matsuda, R.; Enokida, H.; Chiyomaru, T.; Kikkawa, N.; Sugimoto, T.; Kawakami, K.; Tatarano, S.; Yoshino, H.; Toki, K.; Uchida, Y.; et al. LY6K is a novel molecular target in bladder cancer on basis of integrate genome-wide profiling. Br. J. Cancer 2010, 104, 376–386. [Google Scholar] [CrossRef] [PubMed]
- Selvanesan, B.C.; Varghese, S.; Andrys-Olek, J.; Arriaza, R.H.; Prakash, R.; Tiwari, P.B.; Hupalo, D.; Gusev, Y.; Patel, M.N.; Contente, S. Lymphocyte antigen 6K signaling to aurora kinase promotes advancement of the cell cycle and the growth of cancer cells, which is inhibited by LY6K-NSC243928 interaction. Cancer Lett. 2023, 558, 216094. [Google Scholar] [CrossRef]
- Kim, Y.; Son, D.; Yoo, K.H.; Park, J.H. miRNAs involved in LY6K and estrogen receptor-alpha contribute to tamoxifen susceptibility in breast cancer. Cancer Res. 2017, 77, 42261. [Google Scholar] [CrossRef]
- Haensel, D.; Gaddam, S.; Li, N.Y.; Gonzalez, F.; Patel, T.; Cloutier, J.M.; Sarin, K.Y.; Tang, J.Y.; Rieger, K.E.; Aasi, S.Z.; et al. LY6D marks pre-existing resistant basosquamous tumor subpopulations. Nat. Commun. 2022, 13, 7520. [Google Scholar] [CrossRef]
- Barros-Silva, J.D.; Linn, D.E.; Steiner, I.; Guo, G.; Ali, A.; Pakula, H.; Ashton, G.; Peset, I.; Brown, M.; Clarke, N.W.; et al. Single-Cell Analysis Identifies LY6D as a Marker Linking Castration-Resistant Prostate Luminal Cells to Prostate Progenitors and Cancer. Cell Rep. 2018, 25, 3504–3518.e6. [Google Scholar] [CrossRef]
- Brakenhoff, R.H.; Gerretsen, M.; Knippels, E.M.; van Dijk, M.; van Essen, H.; Weghuis, D.O.; Sinke, R.J.; Snow, G.B.; van Dongen, G.A. The human E48 antigen, highly homologous to the murine Ly-6 antigen ThB, is a GPI-anchored molecule apparently involved in keratinocyte cell-cell adhesion. J. Cell Biol. 1995, 129, 1677–1689. [Google Scholar] [CrossRef]
- Torenbeek, R.; Blomjous, C.E.; Quak, J.J.; Ybema, S.; Meijer, C.J. Use of monoclonal antibody E48 in diagnosing transitional cell carcinoma of urinary bladder. J. Clin. Pathol. 1992, 45, 303–307. [Google Scholar] [CrossRef]
- Colnot, D.R.; Nieuwenhuis, E.J.; Kuik, D.J.; Leemans, C.R.; Dijkstra, J.; Snow, G.B.; van Dongen, G.A.; Brakenhoff, R.H. Clinical significance of micrometastatic cells detected by E48 (Ly-6D) reverse transcription-polymerase chain reaction in bone marrow of head and neck cancer patients. Clin. Cancer Res. 2004, 10, 7827–7833. [Google Scholar] [CrossRef]
- Luo, L.; McGarvey, P.; Madhavan, S.; Kumar, R.; Gusev, Y.; Upadhyay, G. Distinct lymphocyte antigens 6 (Ly6) family members Ly6D, Ly6E, Ly6K and Ly6H drive tumorigenesis and clinical outcome. Oncotarget 2016, 7, 11165–11193. [Google Scholar] [CrossRef] [PubMed]
- Romeu, C.; Farre, X.; Cardesa, A.; Nadal, A. Expression of Ep-CAM, but not of E48, associates with nodal involvement in advanced squamous cell carcinomas of the larynx. Histopathology 2013, 62, 954–961. [Google Scholar] [CrossRef] [PubMed]
- Mayama, A.; Takagi, K.; Suzuki, H.; Sato, A.; Onodera, Y.; Miki, Y.; Sakurai, M.; Watanabe, T.; Sakamoto, K.; Yoshida, R.; et al. OLFM4, LY6D and S100A7 as potent markers for distant metastasis in estrogen receptor-positive breast carcinoma. Cancer Sci. 2018, 109, 3350–3359. [Google Scholar] [CrossRef] [PubMed]
- Osz, A.; Lanczky, A.; Gyorffy, B. Survival analysis in breast cancer using proteomic data from four independent datasets. Sci. Rep. 2021, 11, 16787. [Google Scholar] [CrossRef] [PubMed]
- Chang, S.H.; Mezzano-Robinson, V.; Zhou, H.; Moreira, A.; Pillai, R.; Ramaswami, S.; Loomis, C.; Heguy, A.; Tsirigos, A.; Pass, H.I. Digital Spatial Profiling to Predict Recurrence in Grade 3 Stage I Lung Adenocarcinoma. J. Thorac. Cardiovasc. Surg. 2023, S0022-5223, 01004–01008. [Google Scholar] [CrossRef]
- Liu, M.; Di, J.; Liu, Y.; Su, Z.; Jiang, B.; Wang, Z.; Su, X. Comparison of EpCAM high CD44 + cancer stem cells with EpCAM high CD44—Tumor cells in colon cancer by single-cell sequencing. Cancer Biol. Ther. 2018, 19, 939–947. [Google Scholar] [CrossRef]
- Jing, F.; Kim, H.J.; Kim, C.H.; Kim, Y.J.; Lee, J.H.; Kim, H.R. Colon cancer stem cell markers CD44 and CD133 in patients with colorectal cancer and synchronous hepatic metastases. Int. J. Oncol. 2015, 46, 1582–1588. [Google Scholar] [CrossRef]
- Zakaria, S.; Elsebaey, S.; Allam, S.; El-Sisi, A. Modulating the Siah2-PHD3-HIF1α axis and/or autophagy potentially retard colon cancer proliferation possibly, due to the damping of colon cancer stem cells. Biomed. Pharmacother. Biomed. Pharmacother. 2022, 154, 113562. [Google Scholar] [CrossRef] [PubMed]
- AbdelMageed, M.; Ismail, H.T.H.; Olsson, L.; Lindmark, G.; Hammarström, M.-L.; Hammarström, S.; Sitohy, B. Clinical Significance of Stem Cell Biomarkers EpCAM, LGR5 and LGR4 mRNA Levels in Lymph Nodes of Colon Cancer Patients. Int. J. Mol. Sci. 2021, 23, 403. [Google Scholar] [CrossRef]
- Upadhyay, G. Emerging Role of Lymphocyte Antigen-6 Family of Genes in Cancer and Immune Cells. Front. Immunol. 2019, 10, 819. [Google Scholar] [CrossRef]
- Bhuvaneshwar, K. Variant analysis of LY6 genes in TCGA ovarian cancer. Cancer Res. 2018, 78, LB-392. [Google Scholar] [CrossRef]
- Liu, D.; Xu, C.; Liu, Y.; Ouyang, W.; Lin, S.; Xu, A.; Zhang, Y.; Xie, Y.; Huang, Q.; Zhao, W.; et al. A systematic survey of LU domain-containing proteins reveals a novel human gene, LY6A, which encodes the candidate ortholog of mouse Ly-6A/Sca-1 and is aberrantly expressed in pituitary tumors. Front. Med. 2023, 17, 458–475. [Google Scholar] [CrossRef] [PubMed]
- Semba, T.; Sato, R.; Kasuga, A.; Suina, K.; Shibata, T.; Kohno, T.; Suzuki, M.; Saya, H.; Arima, Y. Lung Adenocarcinoma Mouse Models Based on Orthotopic Transplantation of Syngeneic Tumor-Initiating Cells Expressing EpCAM, SCA-1, and Ly6d. Cancers 2020, 12, 3805. [Google Scholar] [CrossRef] [PubMed]
- Ali, D.; Zepp, M.; Bozza, M.; Nikolova, M.; Harbottle, R.; Berger, M.R. Ly6-neurotoxin1 knockout in PDAC cells inhibits their growth in vitro and in vivo. Cancer Res. 2020, 80, 4717. [Google Scholar] [CrossRef]
- Tian, J.-H.; Chang, S.-J.; Guo, H.-X.; Ji, H.; Wang, Y.-H. Inhibition of proliferation of breast cancer cells by down-regulation of cyclin d1 and survivin protein expression by erk inhibitor u0126(Article). Chin. Pharmacol. Bull. 2019, 35, 1061–1066. [Google Scholar]
- Yang, C.; Wang, M.; Zhou, J.; Chi, Q. MicroRNA-335 targets the MEK/ERK pathway to regulate the proliferation and metastasis of colon cancer. Am. J. Transl. Res. 2020, 12, 7899–7907. [Google Scholar]
- Hong, C.S.; Sun, E.G.; Choi, J.N.; Kim, D.H.; Kim, J.H.; Ryu, K.H.; Shim, H.J.; Hwang, J.E.; Bae, W.K.; Kim, H.R.; et al. Fibroblast growth factor receptor 4 increases epidermal growth factor receptor (EGFR) signaling by inducing amphiregulin expression and attenuates response to EGFR inhibitors in colon cancer. Cancer Sci. 2020, 111, 3268–3278. [Google Scholar] [CrossRef] [PubMed]
- Cheng, B.; Rong, A.; Zhou, Q.; Li, W. LncRNA LINC00662 promotes colon cancer tumor growth and metastasis by competitively binding with miR-340-5p to regulate CLDN8/IL22 co-expression and activating ERK signaling pathway. J. Exp. Clin. Cancer Res. 2020, 39, 5. [Google Scholar] [CrossRef]
- Qi, Y.; Zou, H.; Zhao, X.; Kapeleris, J.; Monteiro, M.; Li, F.; Xu, Z.P.; Deng, Y.; Wu, Y.; Tang, Y.; et al. Inhibition of colon cancer K-Ras(G13D) mutation reduces cancer cell proliferation but promotes stemness and inflammation via RAS/ERK pathway. Front. Pharmacol. 2022, 13, 996053. [Google Scholar] [CrossRef] [PubMed]
- Yu, M.; Xu, C.; Zhang, H.; Lun, J.; Wang, L.; Zhang, G.; Fang, J. The tyrosine phosphatase SHP2 promotes proliferation and oxaliplatin resistance of colon cancer cells through AKT and ERK. Biochem. Biophys. Res. Commun. 2021, 563, 1–7. [Google Scholar] [CrossRef]
- Ullah, R.; Yin, Q.; Snell, A.H.; Wan, L. RAF-MEK-ERK pathway in cancer evolution and treatment. Semin. Cancer Biol. 2022, 85, 123–154. [Google Scholar] [CrossRef] [PubMed]
- Fan, J.; Ren, D.; Wang, J.; Liu, X.; Zhang, H.; Wu, M.; Yang, G. Bruceine D induces lung cancer cell apoptosis and autophagy via the ROS/MAPK signaling pathway in vitro and in vivo. Cell Death Dis. 2020, 11, 126. [Google Scholar] [CrossRef]
- Lv, Y.; Wang, X.; Li, X.; Xu, G.; Bai, Y.; Wu, J.; Piao, Y.; Shi, Y.; Xiang, R.; Wang, L. Nucleotide de novo synthesis increases breast cancer stemness and metastasis via cGMP-PKG-MAPK signaling pathway. PLoS Biol. 2020, 18, e3000872. [Google Scholar] [CrossRef]
- Montico, B.; Giurato, G.; Colizzi, F.; Covre, A.; Comaro, E.; Pivetta, E.; Benedetti, D.; Weisz, A.; Steffan, A.; Maio, M.; et al. Abstract B06: Loss of SPRY1 expression in BRAF-mutant cutaneous melanoma inhibits cell proliferation and improves response to targeted therapy. Cancer Res. 2020, 80, B06. [Google Scholar] [CrossRef]
- Hua, Z.; Wei, R.; Guo, M.; Lin, Z.; Yu, X.; Li, X.; Gu, C.; Yang, Y. YTHDF2 promotes multiple myeloma cell proliferation via STAT5A/MAP2K2/p-ERK axis. Oncogene 2022, 41, 1482–1491. [Google Scholar] [CrossRef]
- Zhang, Q.; Jia, Y.; Pan, P.; Zhang, X.; Jia, Y.; Zhu, P.; Chen, X.; Jiao, Y.; Kang, G.; Zhang, L.; et al. α5-nAChR associated with Ly6E modulates cell migration via TGF-β1/Smad signaling in non-small cell lung cancer. Carcinogenesis 2022, 43, 393–404. [Google Scholar] [CrossRef] [PubMed]
- Hu, H.; Dong, Z.; Wang, X.; Bai, L.; Lei, Q.; Yang, J.; Li, L.; Li, Q.; Liu, L.; Zhang, Y. Dehydrocorydaline inhibits cell proliferation, migration and invasion via suppressing MEK1/2-ERK1/2 cascade in melanoma. OncoTargets Ther. 2019, 12, 5163–5175. [Google Scholar] [CrossRef] [PubMed]
- Zhang, D.; Aldrich, K.; Dexheimer, T.; Ellsworth, E.; Odom, A.; Liby, K. Abstract C063: A novel Nrf2 inhibitor suppresses proliferation and enhances the sensitivity of cancer cells to chemotherapy. Mol. Cancer Ther. 2019, 18, C63. [Google Scholar] [CrossRef]
- Li, R.; Li, H.; Lan, J.; Yang, D.; Lin, X.; Xu, H.; Han, B.; Yang, M.; Su, B.; Liu, F.; et al. Damnacanthal isolated from morinda species inhibited ovarian cancer cell proliferation and migration through activating autophagy. Phytomedicine 2022, 100, 154084. [Google Scholar] [CrossRef]
- Carlsson, G.; Gullberg, B.; Hafstrom, L. Estimation of liver tumor volume using different formulas—An experimental study in rats. J. Cancer Res. Clin. Oncol. 1983, 105, 20–23. [Google Scholar] [CrossRef]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Duan, J.; Wang, Y.; Chen, Y.; Wang, Y.; Li, Q.; Liu, J.; Fu, C.; Cao, C.; Cong, Z.; Su, M. Silencing LY6D Expression Inhibits Colon Cancer in Xenograft Mice and Regulates Colon Cancer Stem Cells’ Proliferation, Stemness, Invasion, and Apoptosis via the MAPK Pathway. Molecules 2023, 28, 7776. https://doi.org/10.3390/molecules28237776
Duan J, Wang Y, Chen Y, Wang Y, Li Q, Liu J, Fu C, Cao C, Cong Z, Su M. Silencing LY6D Expression Inhibits Colon Cancer in Xenograft Mice and Regulates Colon Cancer Stem Cells’ Proliferation, Stemness, Invasion, and Apoptosis via the MAPK Pathway. Molecules. 2023; 28(23):7776. https://doi.org/10.3390/molecules28237776
Chicago/Turabian StyleDuan, Jinyue, Yi Wang, Yuanyuan Chen, Yujue Wang, Qisen Li, Jinrui Liu, Changhao Fu, Chenyu Cao, Zhongyi Cong, and Manman Su. 2023. "Silencing LY6D Expression Inhibits Colon Cancer in Xenograft Mice and Regulates Colon Cancer Stem Cells’ Proliferation, Stemness, Invasion, and Apoptosis via the MAPK Pathway" Molecules 28, no. 23: 7776. https://doi.org/10.3390/molecules28237776
APA StyleDuan, J., Wang, Y., Chen, Y., Wang, Y., Li, Q., Liu, J., Fu, C., Cao, C., Cong, Z., & Su, M. (2023). Silencing LY6D Expression Inhibits Colon Cancer in Xenograft Mice and Regulates Colon Cancer Stem Cells’ Proliferation, Stemness, Invasion, and Apoptosis via the MAPK Pathway. Molecules, 28(23), 7776. https://doi.org/10.3390/molecules28237776