
Azorean Black Tea (Camellia sinensis) Antidermatophytic and Fungicidal Properties


 , , , ,
, , , ,  and
and
Abstract
:
1. Introduction
2. Results
2.1. ABT Extract Preparation and Yield of the Extraction
2.2. Phytochemical Analysis of ABT Extract
2.2.1. Organic Acids
2.2.2. Gallic Acid Derivatives
2.2.3. Flavan 3-Ols
2.2.4. Flavones
2.2.5. Hydroxycinammate Quinic Esters
2.2.6. Flavonols
2.2.7. Theaflavins
2.2.8. Purine Alkaloids
2.3. Antidermatophytic Activity of ABT Aqueous Extract
2.4. Synergy between ABT and Antifungals Clinically Used
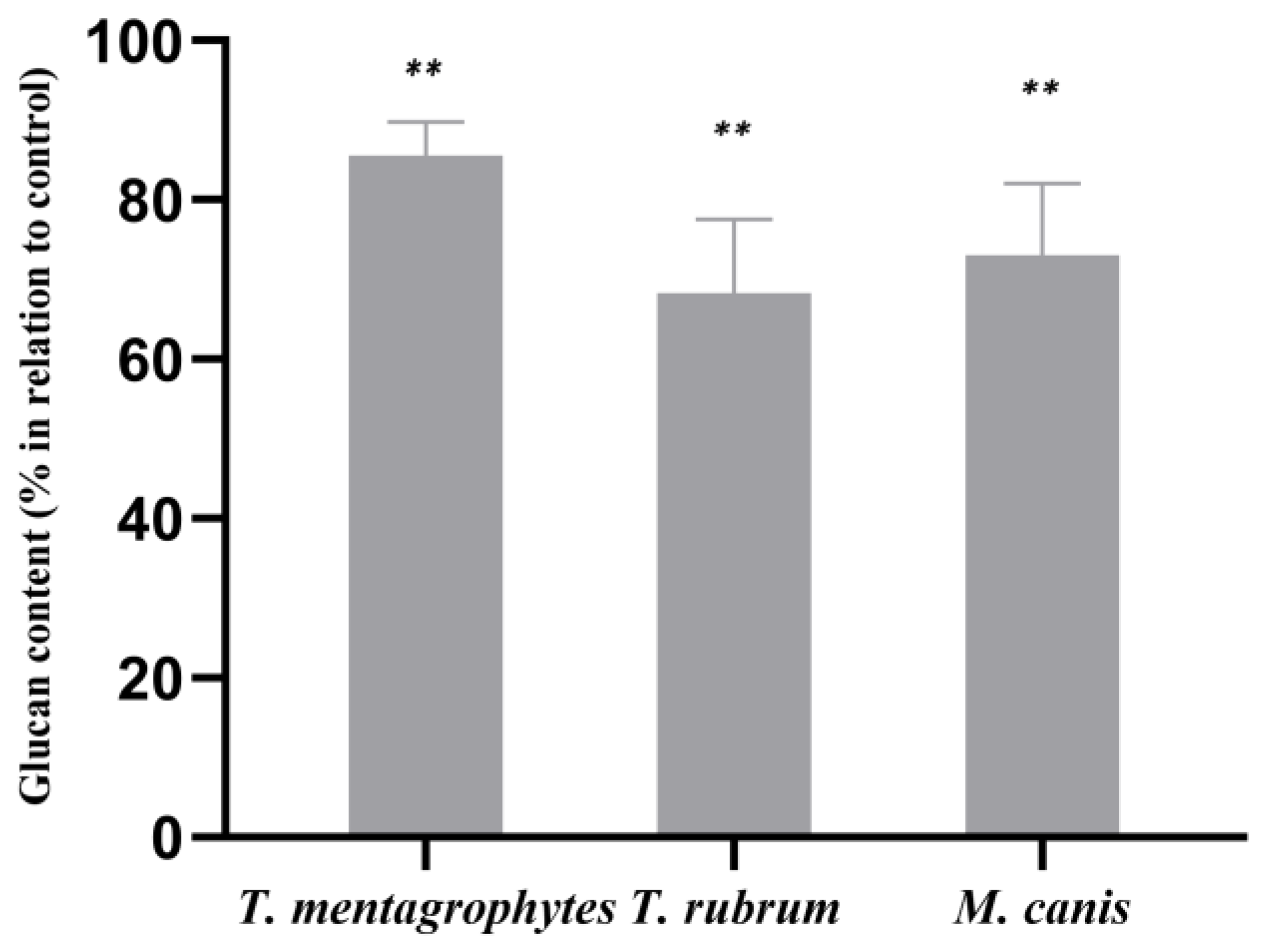
2.5. Modulation of Fungal Cell Wall ß-1,3-glucan and Chitin, and Fungal Cytoplasmic Membrane Ergosterol in Response to ABT
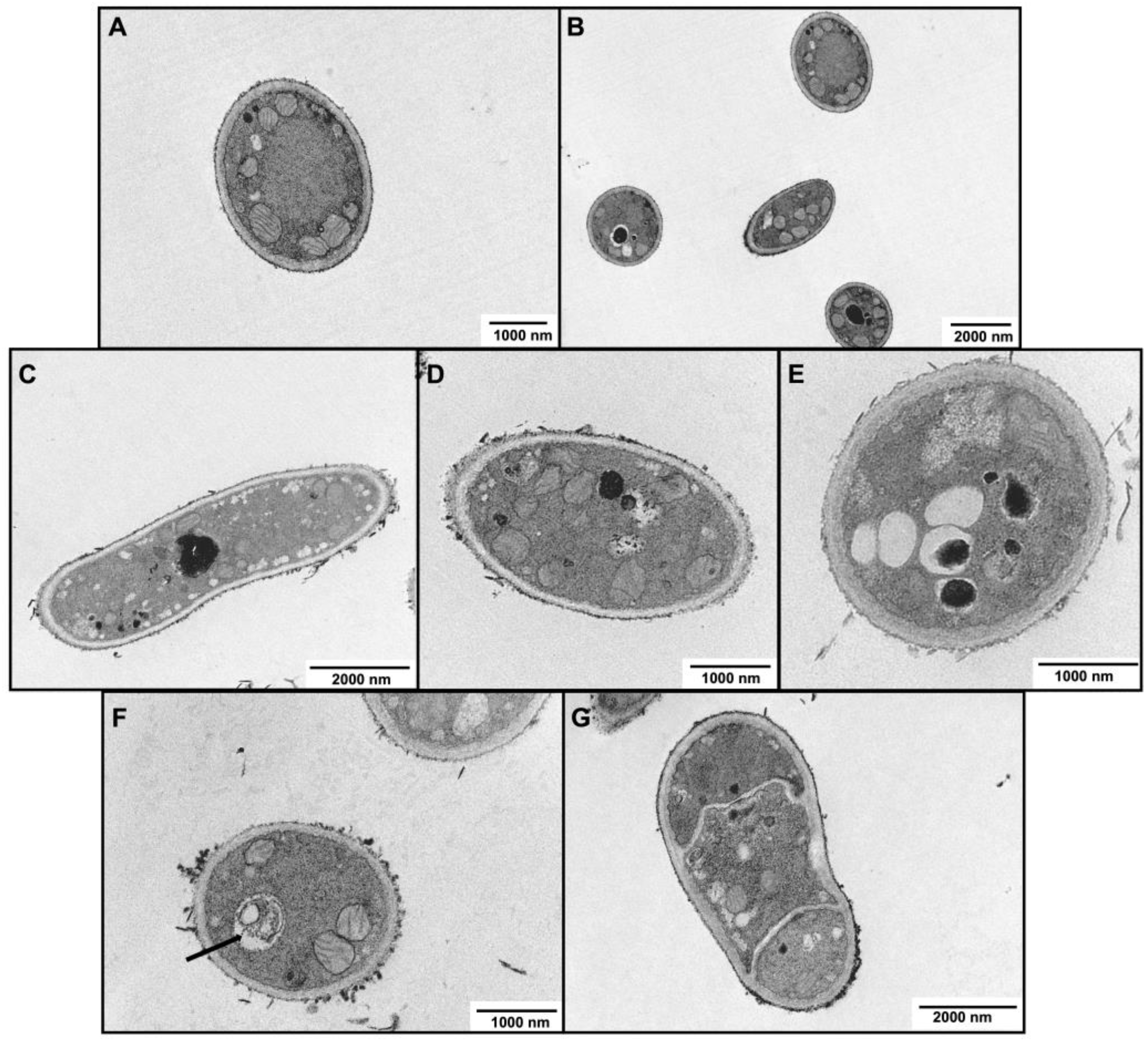
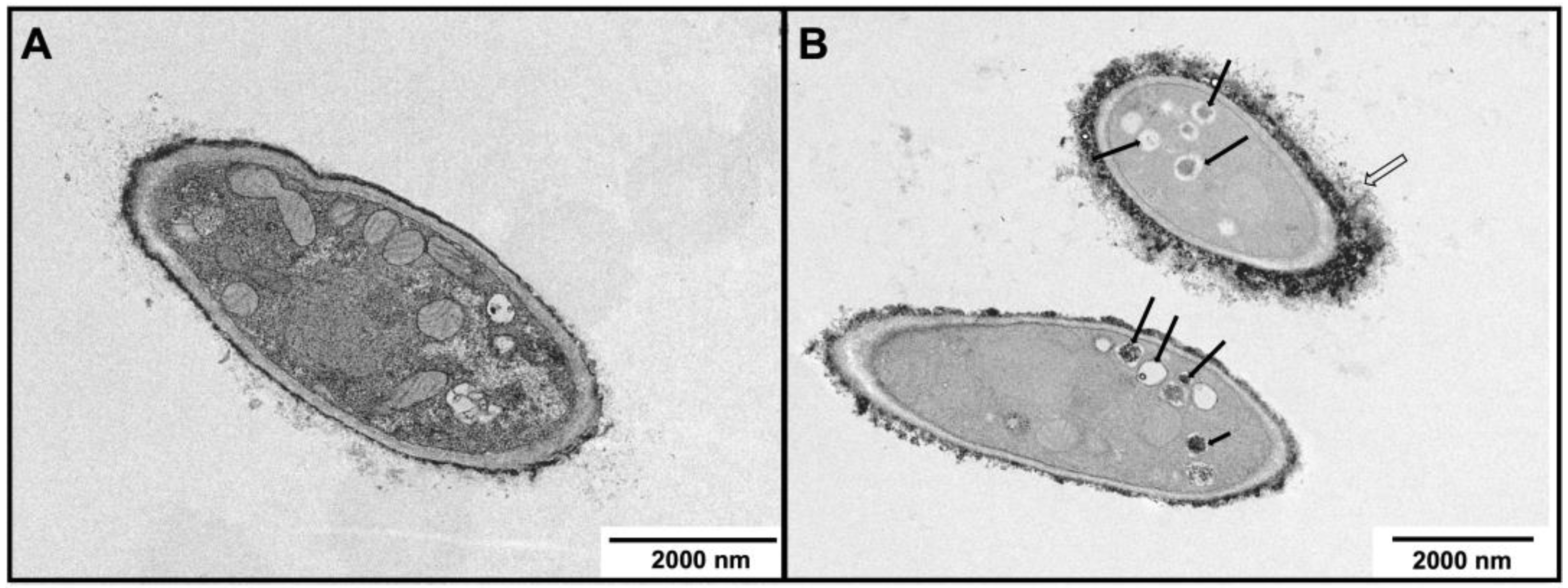
2.6. Characterization of Morphological and Ultrastructural Changes
3. Discussion
4. Materials and Methods
4.1. Crude Aqueous Extracts of Tea
4.2. Phytochemical Analysis
4.3. Fungal Strains and Culture Conditions
4.4. Antidermatophytic Activity of ABT Extract
4.5. Synergy Testing Assays
4.6. Determination of the Modulation of Fungal ß-1,3-glucan, Chitin and Ergosterol in Response to ABT
4.7. Characterization of Morphological and Ultrastructural Changes
4.8. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Moskaluk, A.E.; VandeWoude, S. Current Topics in Dermatophyte Classification and Clinical Diagnosis. Pathogens 2022, 11, 957. [Google Scholar] [CrossRef] [PubMed]
- Weitzman, I.; Summerbell, R.C. The dermatophytes. Clin. Microbiol. Rev. 1995, 8, 240–259. [Google Scholar] [CrossRef]
- Lipner, S.R.; Scher, R.K. Onychomycosis: Clinical Overview and Diagnosis. J. Am. Acad. Dermatol. 2019, 80, 835–851. [Google Scholar] [CrossRef]
- Reich, A.; Szepietowski, J.C. Healh-related quality of life in patients with nail disorders. Am. J. Clin. Dermatol. 2011, 12, 313–320. [Google Scholar] [CrossRef] [PubMed]
- Whittam, L.R.; Hay, R.J. The impact of onychomycosis on quality of life. Clin. Exp. Dermatol. 1997, 22, 87–89. [Google Scholar] [CrossRef] [PubMed]
- Benedict, K.; Jackson, B.R.; Chiller, T.; Beer, K.D. Estimation of Direct Healthcare Costs of Fungal Diseases in the United States. Clin. Infect. Dis. 2019, 68, 1791–1797. [Google Scholar] [CrossRef] [PubMed]
- Havlickova, B.; Czaika, V.A.; Friedrich, M. Epidemiological trends in skin mycoses worldwide. Mycoses 2008, 51, 2–15. [Google Scholar] [CrossRef]
- World Health Organization. Epidemiology and Management of Common Skin Diseases in Children in Developing Countries; World Health Organization: Geneva, Switzerland, 2005; Available online: https://apps.who.int/iris/handle/10665/69229 (accessed on 14 October 2022).
- Kreijkamp-Kaspers, S.; Hawke, K.; Guo, L.; Kerin, G.; Bell-Syer, S.E.; Magin, P.; Bell-Syer, S.V.; van Driel, M.L. Oral antifungal medication for toenail onychomycosis. Cochrane Database Syst. Rev. 2017, 2017, CD010031. [Google Scholar] [CrossRef]
- Sugiura, K.; Sugimoto, N.; Hosaka, S.; Katafuchi-Nagashima, M.; Arakawa, Y.; Tatsumi, Y.; Siu, W.J.; Pillai, R. The low keratin affinity of efinaconazole contributes to its nail penetration and fungicidal activity in topical onychomycosis treatment. Antimicrob. Agents Chemother. 2014, 58, 3837–3842. [Google Scholar] [CrossRef]
- Tatsum, Y.; Yokoo, M.; Arika, T.; Yamaguchi, H. KP–103, a novel triazole derivative, is effective in preventing relapse and successfully treating experimental interdigital tinea pedis and tinea corporis in guinea pigs. Microbiol. Immunol. 2002, 46, 425–432. [Google Scholar] [CrossRef]
- Ghannoum, M. Azole Resistance in Dermatophytes: Prevalence and Mechanism of Action. J. Am. Podiatr. Med. Assoc. 2016, 106, 79–86. [Google Scholar] [CrossRef] [PubMed]
- Yamada, T.; Maeda, M.; Alshahni, M.M.; Tanaka, R.; Yaguchi, T.; Bontems, O.; Salamin, K.; Fratti, M.; Monod, M. Terbinafine resistance of Trichophyton clinical isolates caused by specific point mutations in the squalene epoxidase gene. Antimicrob. Agents Chemother. 2017, 61, e00115-17. [Google Scholar] [CrossRef] [PubMed]
- Martinez-Rossi, N.M.; Bitencourt, T.A.; Peres, N.T.A.; Lang, E.A.S.; Gomes, E.V.; Quaresemin, N.R.; Martins, M.P.; Lopes, L.; Rossi, A. Dermatophyte Resistance to Antifungal Drugs: Mechanisms and Prospectus. Front. Microbiol. 2018, 9, 1108. [Google Scholar] [CrossRef] [PubMed]
- Gupta, A.K.; Cooper, E.A. Update in antifungal therapy of dermatophytosis. Mycopathologia 2008, 166, 353–367. [Google Scholar] [CrossRef]
- Sharma, A.; Gupta, S. Protective manifestation of herbonanoceuticals as antifungals: A possible drug candidate for dermatophytic infection. Health Sci. Rep. 2022, 5, e775. [Google Scholar] [CrossRef]
- Dou, Q.P. Tea in Health and Disease. Nutrients 2019, 11, 929. [Google Scholar] [CrossRef] [PubMed]
- Valadas, L.A.R.; de Oliveira Filho, R.D.; Rodrigues Neto, E.M.; Bandeira, M.A.M.; Fonteles, M.M.F.; Passos, V.F.; Fiallos, A.C.M.; Lotif, M.A.L.; de Sena, N.J.C.; Dantas, T.C.F.B.; et al. Camellia sinensis in Dentistry: Technological Prospection and Scientific Evidence. Evid.-Based Complement. Altern. Med. 2021, 2021, 9966738. [Google Scholar] [CrossRef]
- Paiva, L.; Rego, C.; Lima, E.; Marcone, M.; Baptista, J. Comparative Analysis of the Polyphenols, Caffeine, and Antioxidant Activities of Green Tea, White Tea, and Flowers from Azorean Camellia sinensis Varieties Affected by Different Harvested and Processing Conditions. Antioxidants 2021, 10, 183. [Google Scholar] [CrossRef]
- Friedman, M. Overview of antibacterial, antitoxin, antiviral, and antifungal activities of tea flavonoids and teas. Mol. Nutr. Food Res. 2007, 51, 116–134. [Google Scholar] [CrossRef]
- Inamdar, P.; Vazir, J.; Desai, S.; Patel, D.; Meshram, D. Phytochemical screening and in vitro antifungal activity of Camellia sinensis. Int. J. Pharm. Pharm. Sci. 2014, 6, 148–150. [Google Scholar]
- Chen, F.; Chen, Y.P.; Wu, H.; Li, Y.; Zhang, S.; Ke, J.; Yao, J.Y. Characterization of tea (Camellia sinensis L.) flower extract and insights into its antifungal susceptibilities of Aspergillus flavus. BMC Complement. Med. Ther. 2023, 23, 286. [Google Scholar] [CrossRef]
- Arora, D.S.; Ohlan, D. In vitro studies on antifungal activity of tea (Camellia sinensis) and coffee (Coffea arabica) against wood-rotting fungi. J. Basic Microbiol. 1997, 37, 159–165. [Google Scholar] [CrossRef]
- Akroum, S. Antifungal activity of Camellia sinensis crude extracts against four species of Candida and Microsporum persicolor. J. Med Mycol. 2018, 28, 424–427. [Google Scholar] [CrossRef] [PubMed]
- Dai, J.; Sameen, D.E.; Zeng, Y.; Li, S.; Qin, W.; Liu, Y. An overview of tea polyphenols as bioactive agents for food packaging applications. LWT 2022, 167, 113845. [Google Scholar] [CrossRef]
- Teixeira, A.M.; Sousa, C. A Review on the Biological Activity of Camellia Species. Molecules 2021, 26, 2178. [Google Scholar] [CrossRef]
- Wong, M.; Sirisena, S.; Ng, K. Phytochemical profile of differently processed tea: A review. J. Food Sci. 2022, 87, 1925–1942. [Google Scholar] [CrossRef] [PubMed]
- Naveed, M.; BiBi, J.; Kamboh, A.A.; Suheryani, I.; Kakar, I.; Fazlani, S.A.; FangFang, X.; Kalhoro, S.A.; Yunjuan, L.; Kakar, M.U.; et al. Pharmacological values and therapeutic properties of black tea (Camellia sinensis): A comprehensive overview. Biomed. Pharmacother. 2018, 100, 521–531. [Google Scholar] [CrossRef]
- Wilmot, M. Inhibition of Phytopathogenic Fungi on Selected Vegetable Crops by Catechins, Caffeine, Theanine and Extracts of Camellia sinensis (L.) O. Kuntze. (Magister Scientiae Dissertation). 2006. Available online: https://repository.up.ac.za/dspace/handle/2263/26854 (accessed on 16 January 2023).
- Sitheeque, M.A.; Panagoda, G.J.; Yau, J.; Amarakoon, A.M.; Udagama, U.R.; Samaranayake, L.P. Antifungal activity of black tea polyphenols (catechins and theaflavins) against Candida species. Chemotherapy 2009, 55, 189–196. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Bandara, H.M.H.N.; Mikkelsen, D.; Samaranayake, L.P. Effects of tea extracts on the colonization behaviour of Candida species: Attachment inhibition and biofilm enhancement. J. Med. Microbiol. 2017, 66, 1244–1252. [Google Scholar] [CrossRef]
- Camargo, L.E.; Pedroso, L.S.; Vendrame, S.C.; Mainardes, R.M.; Khalil, N.M. Antioxidant and antifungal activities of Camellia sinensis (L.) Kuntze leaves obtained by different forms of production. Braz. J. Biol. 2016, 76, 428–434. [Google Scholar] [CrossRef]
- Okubo, S.; Toda, M.; Hara, Y.; Shimamura, T. Antifungal and fungicidal activities of tea extract and catechin against Trichophyton. Nihon Saikingaku Zasshi 1991, 46, 509–514. [Google Scholar] [CrossRef] [PubMed]
- Park, B.J.; Taguchi, H.; Kamei, K.; Matsuzawa, T.; Hyon, S.H.; Park, J.C. In vitro antifungal activity of epigallocatechin 3-O-gallate against clinical isolates of dermatophytes. Yonsei Med. J. 2011, 52, 535–538. [Google Scholar] [CrossRef] [PubMed]
- Masike, K.; Mhlongo, M.I.; Mudau, S.P.; Nobela, O.; Ncube, E.N.; Tugizimana, F.; George, M.J.; Madala, N.E. Highlighting mass spectrometric fragmentation differences and similarities between hydroxycinnamoyl-quinic acids and hydroxycinnamoyl-isocitric acids. Chem. Cent. J. 2017, 11, 29. [Google Scholar] [CrossRef] [PubMed]
- Kelebek, H. LC-DAD–ESI-MS/MS characterization of phenolic constituents in Turkish black tea: Effect of infusion time and temperature. Food Chem. 2016, 204, 227–238. [Google Scholar] [CrossRef]
- Enomoto, H. Unique distribution of ellagitannins in ripe strawberry fruit revealed by mass spectrometry imaging. Curr. Res. Food Sci. 2021, 4, 821–828. [Google Scholar]
- Gomes, P.; Quirós-Guerrero, L.; Silva, C.; Pamplona, S.; Boutin, J.A.; Eberlin, M.; Wolfender, J.L.; Silva, M. Feature-Based Molecular Network-Guided Dereplication of Natural Bioactive Products from Leaves of Stryphnodendron pulcherrimum (Willd.) Hochr. Metabolites 2021, 11, 281. [Google Scholar] [CrossRef] [PubMed]
- Lee, M.K.; Kim, H.W.; Lee, S.H.; Kim, Y.J.; Asamenew, G.; Choi, J.; Lee, J.-W.; Jung, H.-A.; Yoo, S.M.; Kim, J.B. Characterization of catechins, theaflavins, and flavonols by leaf processing step in green and black teas (Camellia sinensis) using UPLC-DAD-QToF/MS. Eur. Food Res. Technol. 2019, 245, 997–1010. [Google Scholar] [CrossRef]
- AlEraky, D.M.; Abuohashish, H.M.; Gad, M.M.; Alshuyukh, M.H.; Bugshan, A.S.; Almulhim, K.S.; Mahmoud, M.M. The Antifungal and Antibiofilm Activities of Caffeine against Candida albicans on Polymethyl Methacrylate Denture Base Material. Biomedicines 2022, 10, 2078. [Google Scholar] [CrossRef]
- Barros, M.E.D.S.; Santos, D.A.; Hamdan, J.S. Evaluation of susceptibility of Trichophyton mentagrophytes and Trichophyton rubrum clinical isolates to antifungal drugs using a modified CLSI microdilution method (M38-A). J. Med. Microbiol. 2007, 56, 514–518. [Google Scholar] [CrossRef]
- Hofbauer, B.; Leitner, I.; Ryder, N.S. In vitro susceptibility of Microsporum canis and other dermatophyte isolates from veterinary infections during therapy with terbinafine or griseofulvin. Med. Mycol. 2002, 40, 179–183. [Google Scholar] [CrossRef]
- Hazen, K.C. Fungicidal versus fungistatic activity of terbinafine and itraconazole: An in vitro comparison. J. Am. Acad. Dermatol. 1998, 38, S37–S41. [Google Scholar] [CrossRef] [PubMed]
- Shaban, S.; Patel, M.; Ahmad, A. Improved efficacy of antifungal drugs in combination with monoterpene phenols against Candida auris. Sci. Rep. 2020, 10, 1162. [Google Scholar] [CrossRef] [PubMed]
- Aboulwafa, M.M.; Youssef, F.S.; Gad, H.A.; Altyar, A.E.; Al-Azizi, M.M.; Ashour, M.L. A Comprehensive Insight on the Health Benefits and Phytoconstituents of Camellia sinensis and Recent Approaches for Its Quality Control. Antioxidants 2019, 8, 455. [Google Scholar] [CrossRef] [PubMed]
- Paiva, L.; Lima, E.; Motta, M.; Marcone, M.; Baptista, J. Variability of antioxidant properties, catechins, caffeine, L-theanine and other amino acids in different plant parts of Azorean Camellia sinensis. Curr. Res. Food Sci. 2020, 3, 227–234. [Google Scholar] [CrossRef]
- Gonçalves Bortolini, D.; Windson Isidoro Haminiuk, C.; Cristina Pedro, A.; de Andrade Arruda Fernandes, I.; Maria Maciel, G. Processing, chemical signature and food industry applications of Camellia sinensis teas: An overview. Food Chem. X 2021, 12, 100160. [Google Scholar] [CrossRef] [PubMed]
- Baptista, J.; Lima, E.; Paiva, L.; Andrade, A.L.; Alves, M.G. Comparison of Azorean tea theanine to teas from other origins by HPLC/DAD/FD. Effect of fermentation, drying temperature, drying time and shoot maturity. Food Chem. 2012, 132, 2181–2187. [Google Scholar] [CrossRef]
- Paiva, L.; Lima, E.; Motta, M.; Marcone, M.; Baptista, J. Investigation of the Azorean Camellia sinensis Processing Conditions to Maximize the Theaflavin 3,3’-di-O-Gallate Content as a Potential Antiviral Compound. Antioxidants 2022, 11, 1066. [Google Scholar] [CrossRef]
- Cheruiyot, S.E.; Muturi, M.; Bii, C. Antifungal Activities of Camellia Sinensis Crude Extract on Selected Pathogenic and Mycotoxic Fungi. J. Bacteriol. Mycol. 2015, 2, 1015. [Google Scholar]
- Rodrigues, V.d.C.; da Silva, M.V.; dos Santos, A.R.; Zielinski, A.A.F.; Haminiuk, C.W.I. Evaluation of hot and cold extraction of bioactive compounds in teas. Int. J. Food Sci. Technol. 2015, 50, 2038–2045. [Google Scholar] [CrossRef]
- Boros, K.; Jedlinszki, N.; Csupor, D. Theanine and Caffeine Content of Infusions Prepared from Commercial Tea Samples. Pharmacogn. Mag. 2016, 12, 75–79. [Google Scholar]
- Wang, H.; Helliwell, K. Determination of flavonols in green and black tea leaves and green tea infusions by high-performance liquid chromatography. Food Res. Int. 2001, 34, 223–227. [Google Scholar] [CrossRef]
- Rocha, M.F.G.; Sales, J.A.; da Rocha, M.G.; Galdino, L.M.; de Aguiar, L.; Pereira-Neto, W.A.; de Aguiar Cordeiro, R.; Castelo-Branco, D.S.C.M.; Sidrim, J.J.C.; Brilhante, R.S.N. Antifungal effects of the flavonoids kaempferol and quercetin: A possible alternative for the control of fungal biofilms. Biofouling 2019, 35, 320–328. [Google Scholar] [CrossRef] [PubMed]
- Zhan, P.; Liu, W. The Changing Face of Dermatophytic Infections Worldwide. Mycopathologia 2017, 182, 77–86. [Google Scholar] [CrossRef] [PubMed]
- Brescini, L.; Fioriti, S.; Morroni, G.; Barchiesi, F. Antifungal Combinations in Dermatophytes. J. Fungi 2021, 7, 727. [Google Scholar] [CrossRef]
- Kane, A.; Carter, D.A. Augmenting Azoles with Drug Synergy to Expand the Antifungal Toolbox. Pharmaceuticals 2022, 15, 482. [Google Scholar] [CrossRef] [PubMed]
- Hirasawa, M.; Takada, K. Multiple effects of green tea catechin on the antifungal activity of antimycotics against Candida albicans. J. Antimicrob. Chemother. 2004, 53, 225–229. [Google Scholar] [CrossRef] [PubMed]
- Behbehani, J.M.; Irshad, M.; Shreaz, S.; Karched, M. Synergistic effects of tea polyphenol epigallocatechin 3-O-gallate and azole drugs against oral Candida isolates. J. Mycol. Med. 2019, 29, 158–167. [Google Scholar] [CrossRef]
- Yokokawa, D.; Tatematsu, S.; Takagi, R.; Saga, Y.; Roy, H.; Fischer, F.; Becker, H.D.; Kushiro, T. Synthesis of aminoacylated ergosterols: A new lipid component of fungi. Steroids 2021, 169, 108823. [Google Scholar] [CrossRef]
- Loi, M.; Paciolla, C.; Logrieco, A.F.; Mulè, G. Plant Bioactive Compounds in Pre- and Postharvest Management for Aflatoxins Reduction. Front. Microbiol. 2020, 11, 243. [Google Scholar] [CrossRef]
- Bitencourt, T.A.; Komoto, T.T.; Massaroto, B.G.; Miranda, C.E.; Beleboni, R.O.; Marins, M.; Fachin, A.L. Trans-chalcone and quercetin down-regulate fatty acid synthase gene expression and reduce ergosterol content in the human pathogenic dermatophyte Trichophyton rubrum. BMC Complement. Altern. Med. 2013, 13, 229. [Google Scholar] [CrossRef]
- Latgé, J.P. Tasting the fungal cell wall. Cell. Microbiol. 2010, 12, 863–872. [Google Scholar] [CrossRef] [PubMed]
- Bowman, S.M.; Free, S.J. The structure and synthesis of the fungal cell wall. BioEssays 2006, 28, 799–808. [Google Scholar] [CrossRef]
- Gow, N.A.R.; Latgé, J.P.; Munro, C.A. The fungal cell wall: Structure, biosynthesis, and function. Microbiol. Spectr. 2017, 5, 267–292. [Google Scholar] [CrossRef] [PubMed]
- Szymański, M.; Chmielewska, S.; Czyżewska, U.; Malinowska, M.; Tylicki, A. Echinocandins–structure, mechanism of action and use in antifungal therapy. J. Enzym. Inhib. Med. Chem. 2022, 37, 876–894. [Google Scholar] [CrossRef] [PubMed]
- Denning, D.W. Echinocandin antifungal drugs. Lancet 2003, 362, 1142–1151. [Google Scholar] [CrossRef] [PubMed]
- Ganesan, L.T.; Manavathu, E.K.; Cutright, J.L.; Alangaden, G.J.; Chandrasekar, P.H. In vitro activity of nikkomycin Z alone and in combination with polyenes, triazoles or echinocandins against Aspergillus fumigatus. Clin. Microbiol. Infect. 2004, 10, 961–966. [Google Scholar] [CrossRef]
- Fernandes, C.; Anjos, J.; Walker, L.A.; Silva, B.M.; Cortes, L.; Mota, M.; Munro, C.A.; Gow, N.A.; Gonçalves, T. Modulation of Alternaria infectoria cell wall chitin and glucan synthesis by cell wall synthase inhibitors. Antimicrob. Agents Chemother. 2014, 58, 2894–2904. [Google Scholar] [CrossRef]
- Chen, Y.; Pandiri, I.; Joe, Y.; Kim, H.J.; Kim, S.K.; Park, J.; Ryu, J.; Cho, G.J.; Park, J.W.; Ryter, S.W.; et al. Synergistic Effects of Cilostazol and Probucol on ER Stress-Induced Hepatic Steatosis via Heme Oxygenase-1-Dependent Activation of Mitochondrial Biogenesis. Oxidative Med. Cell. Longev. 2016, 2016, 3949813. [Google Scholar] [CrossRef]
- Shingu-Vazquez, M.; Traven, A. Mitochondria and fungal pathogenesis: Drug tolerance, virulence, and potential for antifungal therapy. Eukaryot. Cell 2011, 10, 1376–1383. [Google Scholar] [CrossRef]
- Müller, J.; Polak-Wyss, A.; Melchinger, W. Influence of amorolfine on the morphology of Candida albicans and Trichophyton mentagrophytes. Clin. Exp. Dermatol. 1992, 17, 18–25. [Google Scholar] [CrossRef]
- McCarty, T.P.; Pappas, P.G. Antifungal Pipeline. Front. Cell. Infect. Microbiol. 2021, 11, 732223. [Google Scholar] [CrossRef]
- Koch, W.; Zagórska, J.; Marzec, Z.; Kukula-Koch, W. Applications of Tea (Camellia sinensis) and its Active Constituents in Cosmetics. Molecules 2019, 24, 4277. [Google Scholar] [CrossRef] [PubMed]
- Espinel-Ingroff, A.; Chaturvedi, V.; Fothergill, A.; Rinaldi, M. Optimal testing conditions for determining MICs and minimum fungicidal concentrations of new and established antifungal agents for uncommon molds: NCCLS collaborative study. J. Clin. Microbiol. 2002, 40, 3776–3781. [Google Scholar] [CrossRef] [PubMed]
- Bellio, P.; Fagnani, L.; Nazzicone, L.; Celenza, G. New and simplified method for drug combination studies by checkerboard assay. MethodsX 2021, 8, 101543. [Google Scholar] [CrossRef]
- European Committee for Antimicrobial Susceptibility Testing (EUCAST) of the European Society of Clinical Microbiology and Infectious Dieases (ESCMID). EUCAST Definitive Document E.Def 1.2, May 2000: Terminology relating to methods for the determination of susceptibility of bacteria to antimicrobial agents. Clin. Microbiol. Infect. 2000, 6, 503–508. [Google Scholar] [CrossRef]
- Fernandes, C.; Mota, M.; Barros, L.; Dias, M.I.; Ferreira, I.C.F.R.; Piedade, A.P.; Casadevall, A.; Gonçalves, T. Pyomelanin Synthesis in Alternaria alternata Inhibits DHN-Melanin Synthesis and Decreases Cell Wall Chitin Content and Thickness. Front. Microbiol. 2021, 12, 691433. [Google Scholar] [CrossRef] [PubMed]
- Kahn, J.N.; Hsu, M.J.; Racine, F.; Giacobbe, R.; Motyl, M. Caspofungin susceptibility in Aspergillus and non-Aspergillus molds: Inhibition of glucan synthase and reduction of beta-D-1,3 glucan levels in culture. Antimicrob. Agents Chemother. 2006, 50, 2214–2216. [Google Scholar] [CrossRef]
- Kapteyn, J.C.; Hoyer, L.L.; Hecht, J.E.; Muller, W.H.; Andel, A.; Verkleij, A.J. The cell wall architecture of Candida albicans wild-type cells and cell wall-defective mutants. Mol. Microbiol. 2000, 35, 601–611. [Google Scholar] [CrossRef]
- Breivik, O.N.; Owades, J.L. Yeast analysis, spectrophotometric semimicro determination of ergosterol in yeast. J. Agric. Food Chem. 1957, 5, 360–363. [Google Scholar] [CrossRef]
- Calheiros, D.; Dias, M.I.; Calhelha, R.C.; Barros, L.; Ferreira, I.C.F.R.; Fernandes, C.; Gonçalves, T. Antifungal Activity of Spent Coffee Ground Extracts. Microorganisms 2023, 11, 242. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Peak | Identification | Rt (min.) | λmax. by HPLC/PDA (nm) | a [M − H]/ b [M − H]+ | MS2 | MS3 |
|---|---|---|---|---|---|---|
| 1 | Isocitric acid | 2.14 | 261 | a 191 (100) | [191]: 191 (75),173 (45), 155 (15), 111 (100) | [191 111]: 111 (100) |
| 2 | Galloylquinic acid | 3.06 | 271 | a 343(100) | [191]: 191 (100), 169 (25) | - |
| 3 | Gallic acid | 3.20 | 270 | a 169 (100) | [169]: 169 (70), 125 (100) | [169 125]: 125 (100) |
| 4 | Theobromine | 5.29 | 273 | b 181(100) | - | |
| 5 | galloyl-(epi)gallocatechin-(epi)gallocatechin | 5.35 | 273 | a 761 (100) | [761]: 609 (40), 591 (100) | [761 591]: 591 (50), 573 (100) |
| 6 | (Epi)gallocatechin | 6.27 | 278 | a 305 (100) | [305]: 261 (17), 221 (45), 219 (55), 179 (100) | - |
| 7 | Galloyl-HHDP-glucose | 7.55 | 276 | a 633 (100) | [633]: 633 (50), 463 (10), 301 (100) | [633 301]: 301(100) |
| 8 | 4-p-coumaroylquinic acid | 11.16 | 311, 296 | a 337 (100) | [337]: 173 (100) | [337 173]: 173 (85), 111 (100), 93 (83) |
| 9 | Epigallocatechin gallate | 12.17 | 280 | a 457 (100) | [457]: 331 (60), 305 (40), 287 (20), 169 (100) | [457 169]: 169 (100), 125 (70) |
| 10 | Gallocatechin gallate | 12.81 | 279 | a 457 (100) | [457]: 331 (60), 305 (40), 287 (20), 169 (100) | [457 169]: 169 (100), 125 (50) |
| 11 | Caffein | 13.38 | 273 | b 195 (100) | - | - |
| 12 | 4″-O-Glucosylvitexin | 13.95 | 272 | a 593 (100) | [593]: 593 (30), 575 (20), 503 (20), 473 (100) | [593 473]: 383 (20), 353 (100) |
| 13 | 6-pentosyl–8 hexosyl–apigenin | 18.43 | 273, 342 | a 563 (100) | [563]: 563 (60), 545 (40), 503 (80), 473 (100), 383 (35), 353 (40) | [563 473]: 383 (20), 353 (100) |
| 14 | 6-pentosyl–8 hexosyl–apigenin | 19.03 | 273, 342 | a 563 (100) | [563]: 563 (45), 545 (40), 503 (55), 473 (100), 443 (75), 383 (30), 353 (25) | [563 473]: 383 (15), 353 (100) |
| 15 | 6-pentosyl–8 hexosyl–apigenin | 19.68 | 273, 342 | a 563 (100) | [563]: 563 (50), 545 (20), 503 (25), 473 (50), 443 (75), 383 (100), 353 (20) | - |
| 16 | Epicatechin gallate | 20.04 | 275 | a 441 (100) | [441]: 289 (100), 169 (20) | [441 289]: 289 (40), 245 (100), 205 (25), 179 (20) |
| 17 | Catechin gallate | 20.85 | 274 | a 441 (100) | [441]: 289 (100), 169 (20) | [441 289]: 289 (15), 245 (100), 231 (15), 205 (25), 179 (15) |
| 18 | Quercetin-3-O-galactosyl-rhamnosyl-glucoside | 22.00 | 253sh, 266m 290sh, 350 | a 771 (100) | [771]: 771 (90), 609 (40), 301 (100) | - |
| 19 | Quercetin-3-O-galactosyl-rhamnosyl-glucoside | 23.01 | 253sh, 266m 290sh, 350 | a 771 (100) | [771]: 771 (100), 301 (40) | - |
| 20 | Quercetin-3-O-galactosyl-rhamnosyl-glucoside | 23.56 | 257sh, 264m 291sh, 352 | a 771 (100) | [771]: 771 (70), 301 (40) | - |
| 21 | Quercetin-3-O-rutinoside | 24.72 | 257sh, 264m 291sh, 352 | a 609 (100) | [609]: 609 (100), 301 (100) | - |
| 22 | Quercetin-3-O-rutinoside | 25.30 | 257sh, 264m 291sh, 352 | a 609 (100) | [609]: 609 (100), 301 (100) | - |
| 23 | Kaempferol-3-O-glucosyl-rhamnosyl-glucoside | 26.68 | 265, 299sh, 347 | a 755 (100) | [755]: 755 (40), 593 (15), 285 (100) | - |
| 24 | Kaempferol-3-O-glucosyl-rhamnosyl-glucoside | 27.13 | 265, 299sh, 347 | a 755 (100) | [755]: 755 (40), 593 (15), 285 (100) | - |
| 25 | Kaempferol-3-O-rutinoside | 28.67 | 265, 299sh, 347 | a 593 (100) | [593]: 593 (20), 285 (100) | - |
| 26 | Kaempferol-3-O-rutinoside | 29.32 | 265, 299sh, 347 | a 593 (100) | [593]: 593 (20), 285 (100) | - |
| 27 | Kaempferol-3-O-galactoside | 29.79 | 265, 299sh, 347 | a 447 (100) | [477]: 447 (100), 285 (60) | - |
| 28 | Kaempferol-3-O-glucoside | 30.33 | 265, 299sh, 347 | a 447 (100) | [477]: 447 (100), 285 (60) | - |
| 29 | Quercetin-3-(2G-p-coum-trans-3G)-2G-arabinosyl-3R-rhamnosyl rutinoside | 41.82 | 253sh, 269, 290sh, 359 | a 1033 (100) | [1033]: 887 (100) | [887 741]: 741 (20), 301 (100) |
| 30 | Theaflavin gallate | 44.67 | 270, 359 | a 715 (100) | [715]: 577 (15), 563 (50), 545 (75), 527 (100), 501 (30), 419 (30) | - |
| 31 | Theaflavin gallate | 45.98 | 270, 356 | a 715 (100) | [715]: 697 (40), 563 (100), 545 (75), 527 (55), 483 (55), 407 (45) | - |
| ABT | Terbinafine | Griseofulvin | ||||||
|---|---|---|---|---|---|---|---|---|
| Fungal Species | MIC | MFC | MIC | FICI * | Index | MIC | FICI ** | Index |
| T. mentagrophytes | 250.0 | 250.0 | 0.0312 | 0.75 | Additive | 0.5 | 1.5 | Indifference |
| T. rubrum | 125.0 | 125.0 | 0.0625 | 1 | Additive | 0.5 | 1.5 | Indifference |
| M. canis | 500.0 | 500.0 | 0.0312 | 0.75 | Additive | 0.5 | 1.5 | Indifference |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fernandes, C.; Sousa-Baptista, J.; Lenha-Silva, A.F.; Calheiros, D.; Correia, E.; Figueirinha, A.; Salgueiro, L.; Gonçalves, T. Azorean Black Tea (Camellia sinensis) Antidermatophytic and Fungicidal Properties. Molecules 2023, 28, 7775. https://doi.org/10.3390/molecules28237775
Fernandes C, Sousa-Baptista J, Lenha-Silva AF, Calheiros D, Correia E, Figueirinha A, Salgueiro L, Gonçalves T. Azorean Black Tea (Camellia sinensis) Antidermatophytic and Fungicidal Properties. Molecules. 2023; 28(23):7775. https://doi.org/10.3390/molecules28237775
Chicago/Turabian StyleFernandes, Chantal, José Sousa-Baptista, Ana Filipa Lenha-Silva, Daniela Calheiros, Edmilson Correia, Artur Figueirinha, Lígia Salgueiro, and Teresa Gonçalves. 2023. "Azorean Black Tea (Camellia sinensis) Antidermatophytic and Fungicidal Properties" Molecules 28, no. 23: 7775. https://doi.org/10.3390/molecules28237775
APA StyleFernandes, C., Sousa-Baptista, J., Lenha-Silva, A. F., Calheiros, D., Correia, E., Figueirinha, A., Salgueiro, L., & Gonçalves, T. (2023). Azorean Black Tea (Camellia sinensis) Antidermatophytic and Fungicidal Properties. Molecules, 28(23), 7775. https://doi.org/10.3390/molecules28237775