
How to Target Small-Molecule Fluorescent Imaging Probes to the Plasma Membrane—The Influence and QSAR Modelling of Amphiphilicity, Lipophilicity, and Flip-Flop


Abstract
:1. Introduction
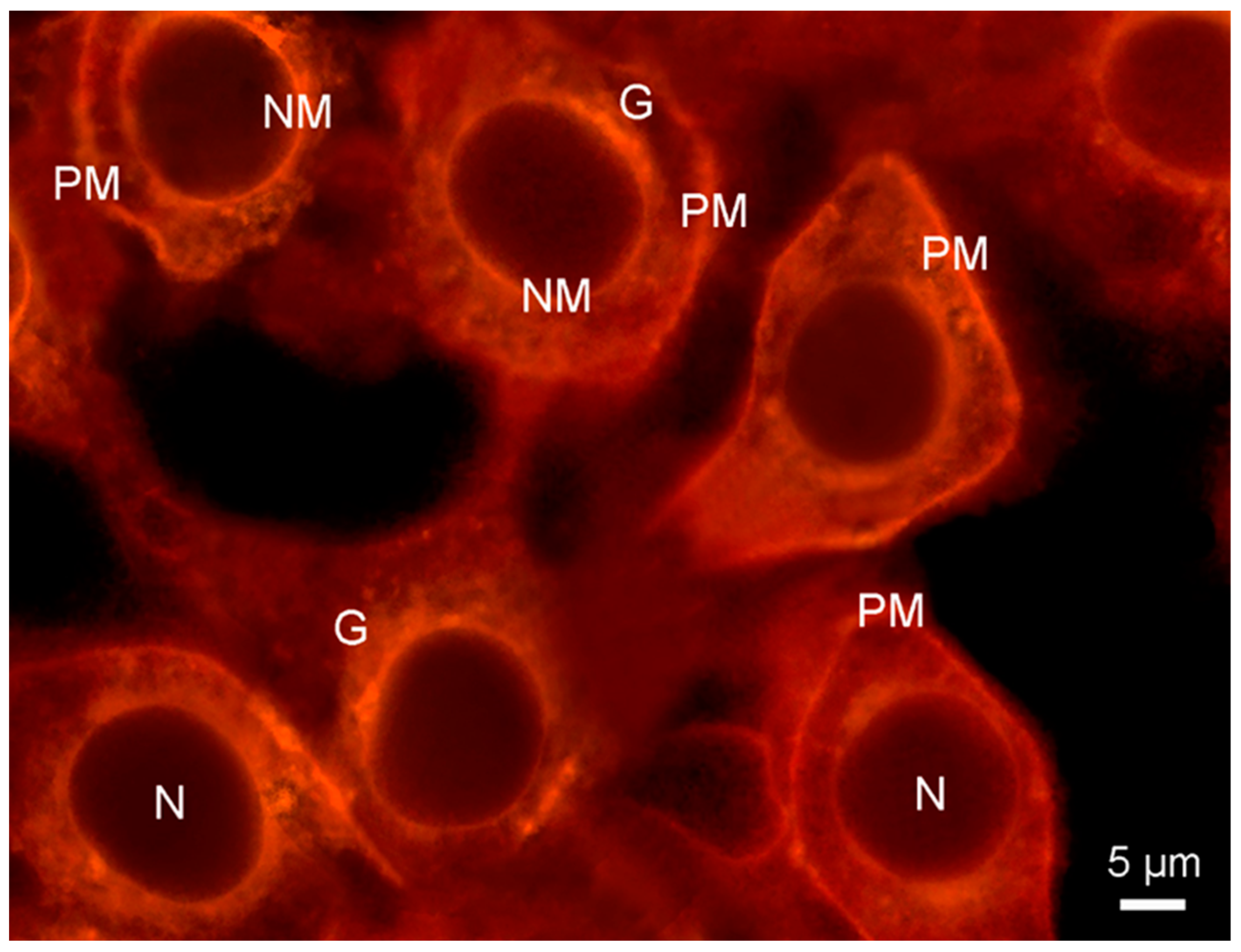
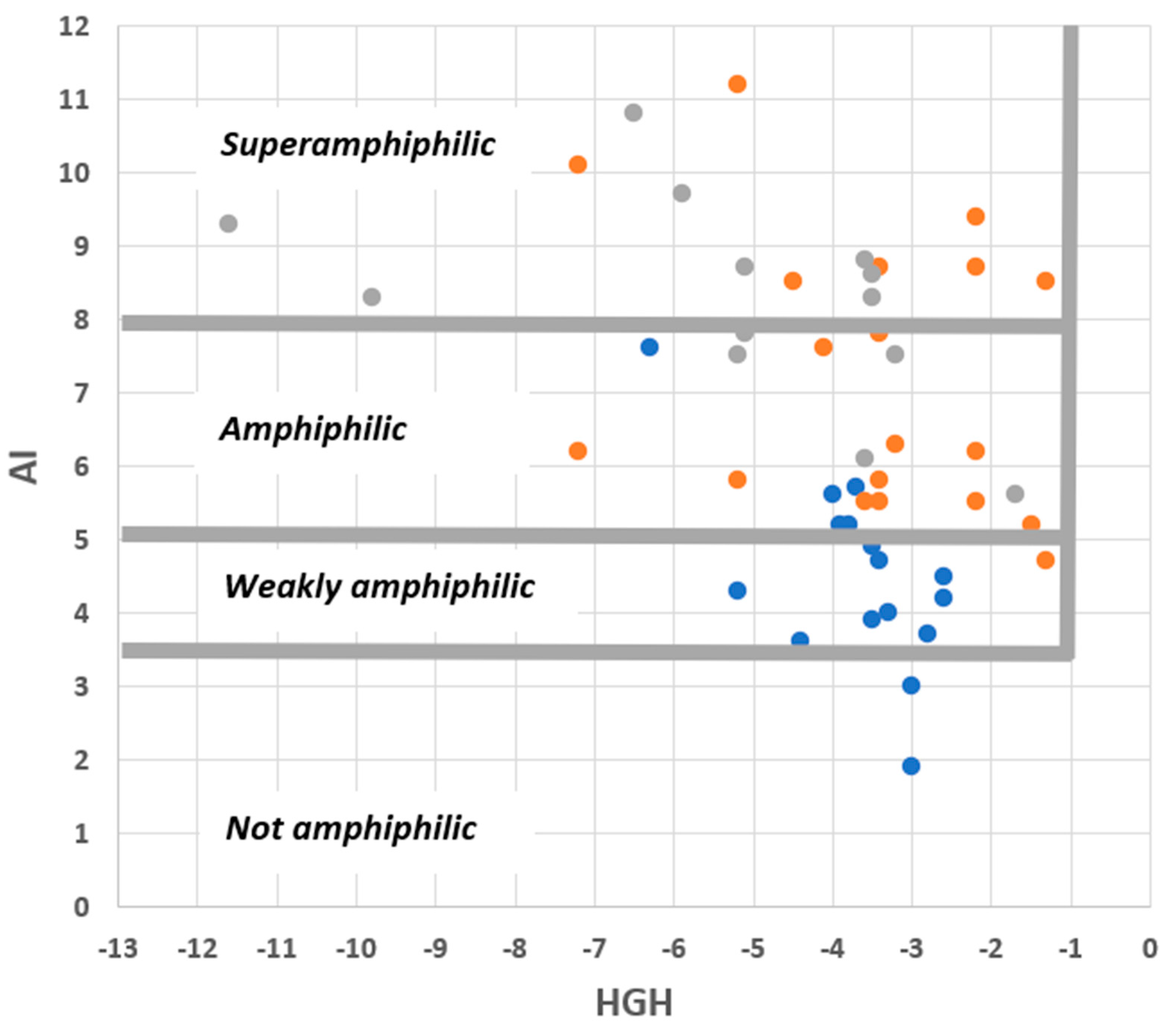
2. Results

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Dye Name | Structure Parameters | Activity Dependence | |
|---|---|---|---|
| AI | Log P | ||
| FM 14-68 | 2.2 | −2.5 | + |
| FM 2-10 | 2.7 | −2.0 | ++ |
| FM 1-43 | 4.2 | −0.5 | ++++ |
| FM 4-84 | 4,2 | −3.3 | ++++ |
| FM 1-84 | 5.0 | 0.3 | ++++ |
| FM 14-27 | 6.6 | 1.9 | − |
| FM 14-29 | 8.2 | 3.5 | − |
| FM 3-25 | 15.9 | 11.2 | − |
| RH 414 | 3.4 | −1.3 | +++ |
| FM 6-55 | 4.2 | −0.5 | +++ |
| FM 10-75 | 3.6 | −1.1 | ++ |
| FM 4-64 | 4.1 | −0.6 | ++++ |
| FM 1-81 | 5.7 | 1.0 | − |
| FM 9-49 | 2.5 | −0.5 | − |
| FM 4-95 | 5.3 | 0.6 | − |
| FM 5-27 | 7.0 | −2.4 | − |
| FM 4-59 | 1.3 | −3.4 | − |
3. Discussion
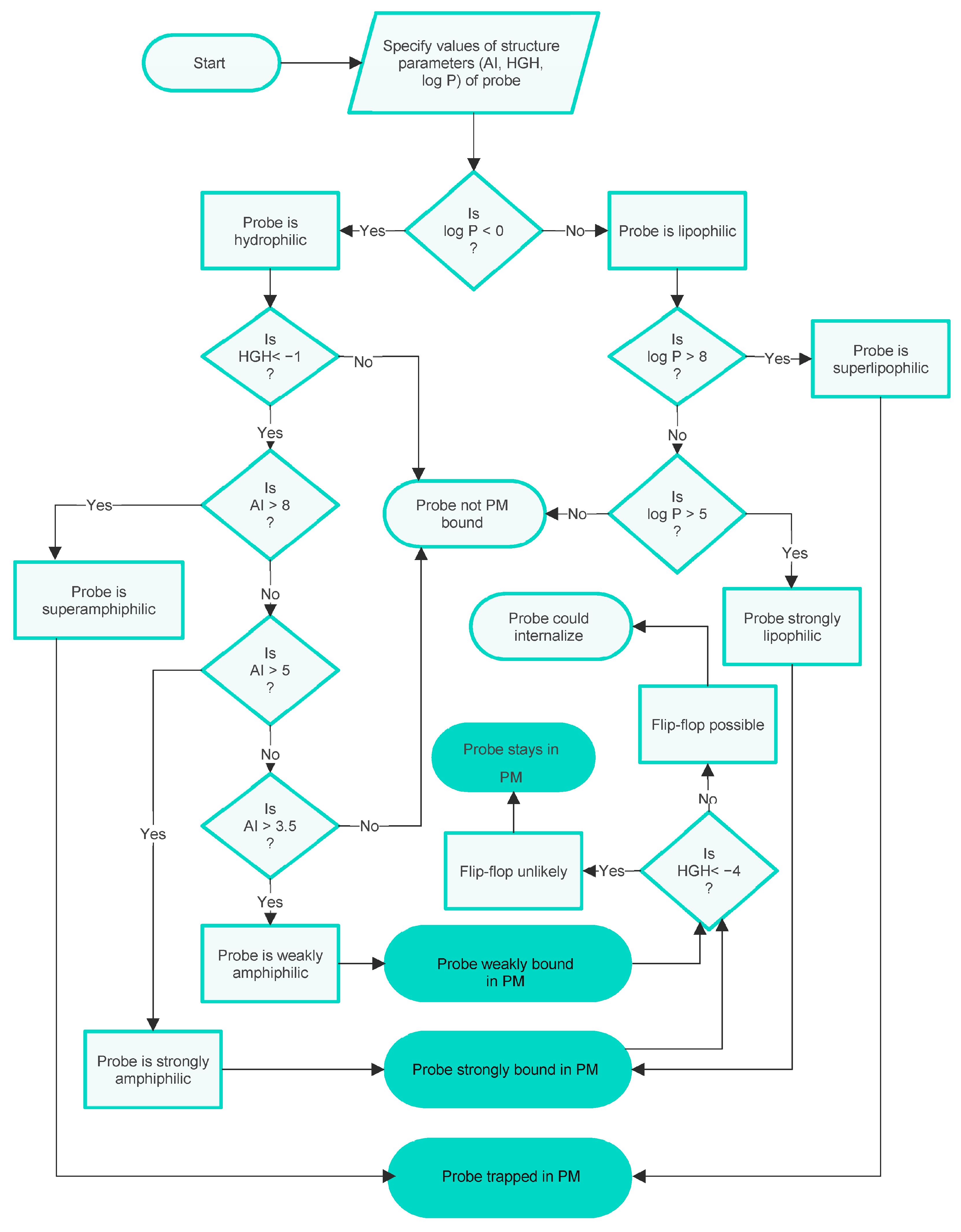
3.1. Clarifications Concerning Amphiphilicity and Lipophilicity
3.2. Core QSAR Models: Describing Membrane Uptake of Probes
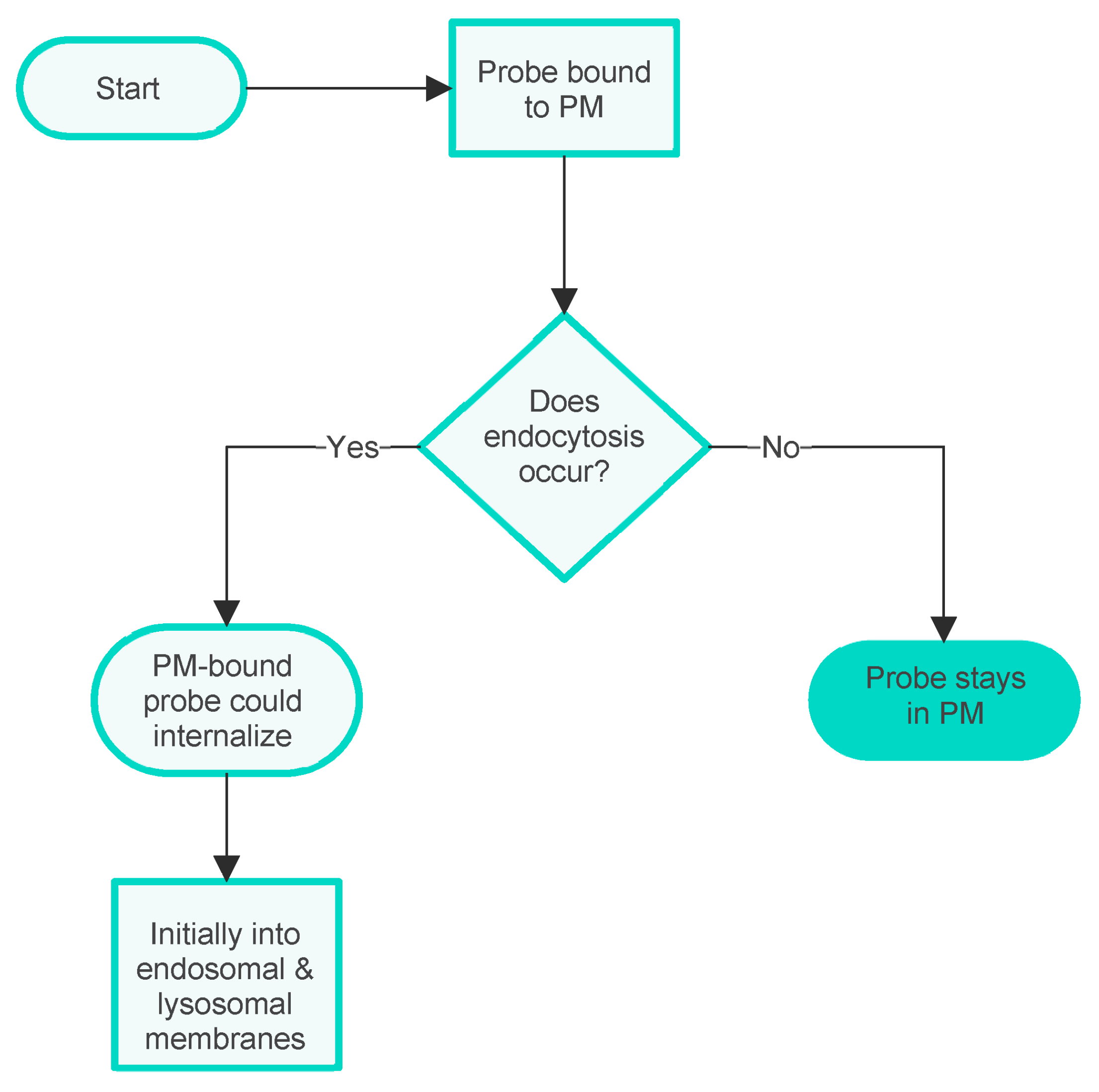
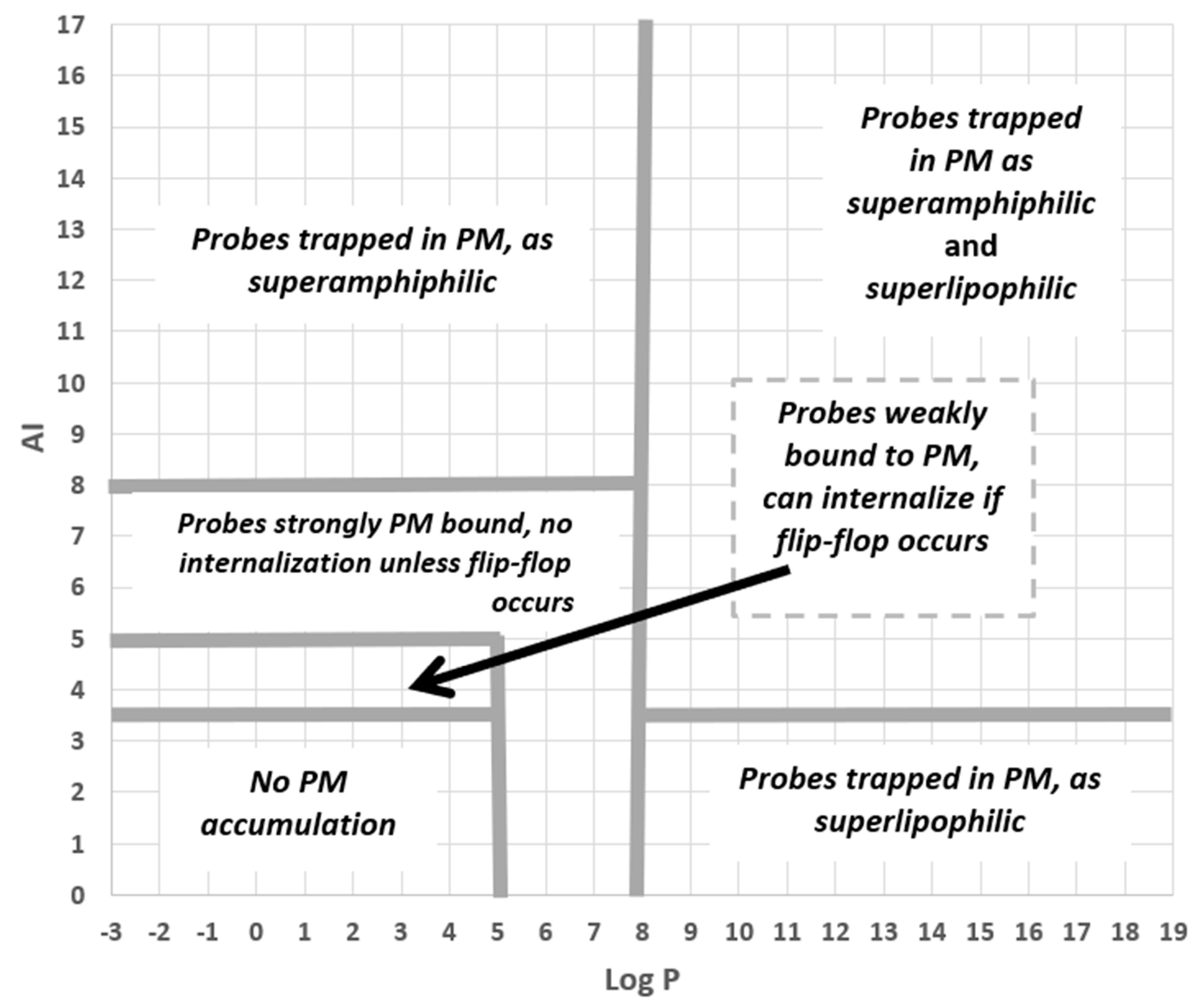
3.3. An Integrated QSAR Model of PM Probes
- To be effectively trapped in the PM.
- To be strongly but not irreversibly bound to the PM, so able to internalise via flip-flop.
- To be weakly bound to the PM, so able to internalise via flip-flop.
- Not bound to the PM.
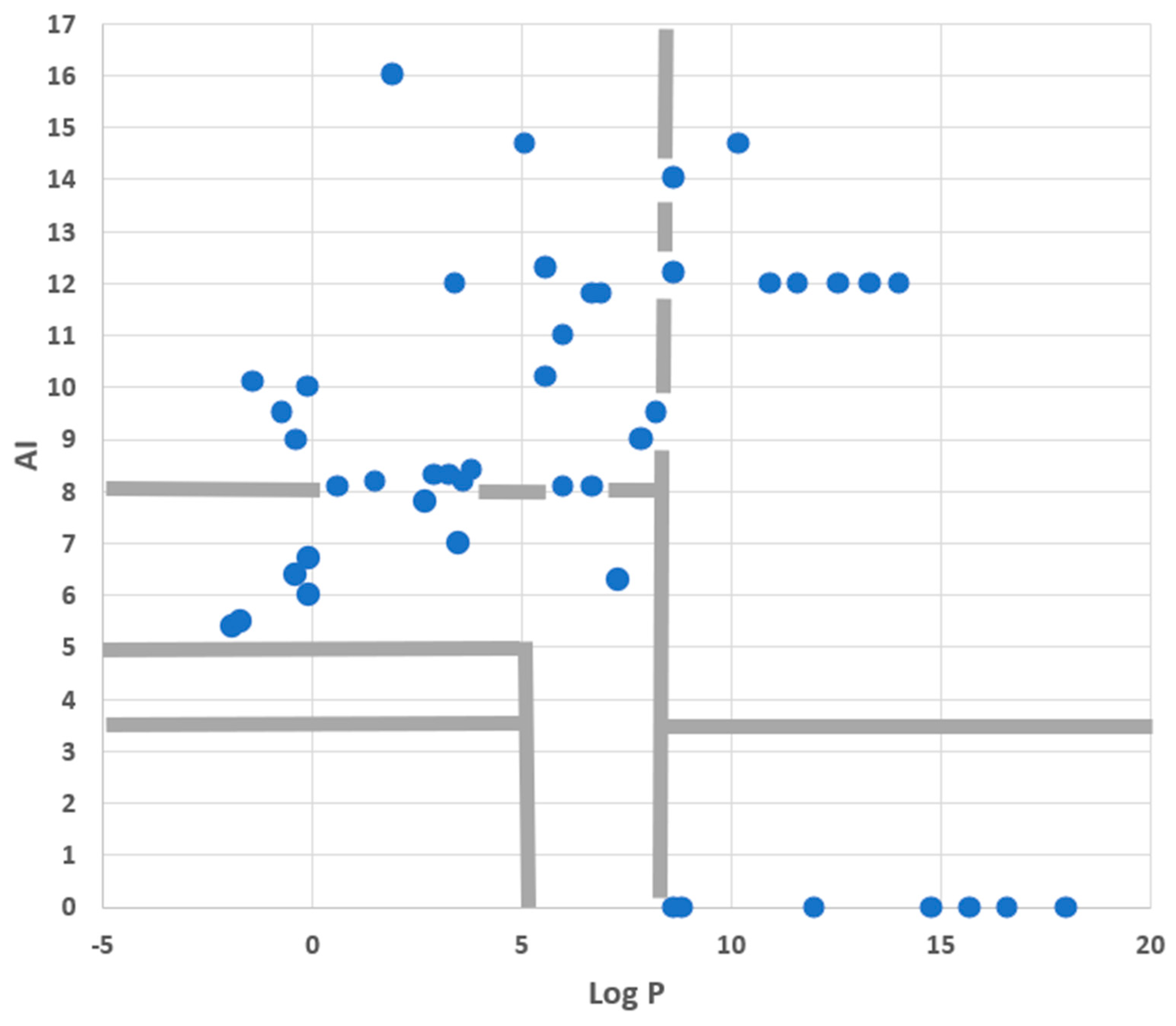
3.4. Assessing the QSAR PM Staining Model and Its Cut-Off Values
3.5. Possible Limitations and Extensions to the QSAR Model
4. Materials and Methods
- Wide coverage of application areas was achieved by assembling sets of commercial surfactants, surface active drugs, saponins, fluorescent PM probes, and fluorescent PM probes of the styryl class.
- For each set of compounds, a single source document was sought that described at least a dozen compounds. Document types were varied and included a book chapter and a monograph, as well as review articles.
- Documents that contained either structural formulae of the compounds or that used nomenclature enabled easy retrieval of the structures from readily accessible sources.
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Liu, C.; Gao, X.; Yuan, J.; Zhang, R. Advances in the development of fluorescent probes for cell plasma membrane imaging. Trends Anal. Chem. 2020, 133, 116092. [Google Scholar] [CrossRef]
- Klymchenko, A.S. Fluorescent probes for lipid membranes: From the cell surface to organelles. Acc. Chem. Res. 2022, 56, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Yang, L.; Chen, Q.; Wang, Z.; Zhang, H.; Sun, H. Small-molecule fluorescent probes for plasma membrane staining: Design, mechanisms and biological applications. Coord. Chem. Rev. 2023, 474, 214862. [Google Scholar] [CrossRef]
- Hansch, C.; Leo, A. The FRAGMENT method of calculating partition coefficients. In Substitution Constants for Correlation Analysis in Chemistry and Biology; Wiley-Interscience: New York, NY, USA, 1979; pp. 18–43. [Google Scholar]
- Amézqueta, S.; Subirats, X.; Fuguet, E.; Rosés, M.; Ràfols, C. Octanol-water partition constant. In Liquid-Phase Extraction; Handbooks in Separation Science; Poole, C.F., Ed.; Elsevier: Amsterdam, The Netherlands, 2020; pp. 183–208. [Google Scholar] [CrossRef]
- Giaginis, C.; Tsantili-Kakoulidou, A. Alternative measures of lipophilicity: From octanol–water partitioning to IAM retention. J. Pharm. Sci. 2008, 97, 2984–3004. [Google Scholar] [CrossRef] [PubMed]
- El Tayar, N.; Testa, B.; Carrupt, P.-A. Polar intermolecular interactions encoded in partition coefficients: An indirect estimation of hydrogen-bond parameters of polyfunctional solutes. J. Phys. Chem. 1992, 96, 1455–1459. [Google Scholar] [CrossRef]
- Goñi, F.M. The basic structure and dynamics of cell membranes: An update of the Singer–Nicolson model. Biochim. Biophys. Acta (BBA)-Biomembr. 2014, 1838, 1467–1476. [Google Scholar] [CrossRef] [PubMed]
- Rosen, M.J.; Kunjappu, J.T. Surfactants and molecular modelling. In Surfactants and Interfacial Phenomena, 4th ed.; Wiley: Hoboken, NJ, USA, 2012; pp. 531–568. [Google Scholar]
- Allhusen, J.S.; Conboy, J.C. The ins and outs of lipid flip-flop. Acc. Chem. Res. 2017, 50, 58–65. [Google Scholar] [CrossRef] [PubMed]
- Horobin, R.W.; Stockert, J.C. Using an integrated QSAR model to check whether small-molecule xenobiotics will accumulate in biomembranes, with particular reference to fluorescent imaging probes. In Quantitative Analysis of Cellular Drug Transport, Disposition and Delivery, Methods in Pharmacology and Toxicology; Rosania, G.R., Thurber, G.M., Eds.; Springer Science & Business Media: New York, NY, USA, 2021; pp. 163–177. [Google Scholar] [CrossRef]
- Blawut, B.; Wolfe, B.; Premanandan, C.; Schuenenmann, G.; Ludsin, S.A.; Liu, S.-L.; Veeramachaneni, D.N.R.; da Silva, M.A.C. Effects of activation and assisted reproduction techniques on the composition, structure, and properties of the sauger (Sander Canadensis) spermatozoa plasma membrane. Theriogenology 2023, 198, 87–99. [Google Scholar] [CrossRef] [PubMed]
- Niikura, K.; Nambara, K.; Okajima, T.; Matsuo, Y.; Ijiro, K. Influence of hydrophobic structures on the plasma membrane permeability of lipidlike molecules. Langmuir 2010, 26, 9170–9175. [Google Scholar] [CrossRef] [PubMed]
- Rosen, M.J. Characteristic features of surfactants. In Surfactants and Interfacial Phenomena, 3rd ed.; Wiley: Hoboken, NJ, USA, 2004; pp. 1–33. [Google Scholar]
- Schreier, S.; Malheiros, S.V.P.; de Paula, E. Surface active drugs: Self-association and interaction with membranes and surfactants. Physicochemical and biological aspects. Biochim. Biophys. Acta (BBA)-Biomembr. 2000, 1508, 210–234. [Google Scholar] [CrossRef]
- Abdelrahman, M.; Jogaigh, S. Bioactive Molecules in Plant Defence. Saponins; Springer: Cham, Switzerland, 2020. [Google Scholar] [CrossRef]
- Betz, W.J.; Mao, F.; Smith, C.B. Imaging exocytosis and endocytosis. Curr. Opin. Neurobiol. 1996, 6, 365–371. [Google Scholar] [CrossRef] [PubMed]
- Stockert, J.C.; Blázquez-Castro, A. Fluorescence Microscopy in Life Sciences; Bentham Science Publishers: Sharjah, United Arab Emirates, 2017. [Google Scholar] [CrossRef]
- Mason, S.F. Color and the Electronic States of Organic Molecules. Chemistry of the Synthetic Dyes; Wiley and Sons: New York, NY, USA, 1970; pp. 169–221. [Google Scholar]
- Lillie, R.D. Conn’s Biological Stains, 9th ed.; Williams & Wilkins: Baltimore, MD, USA, 1977. [Google Scholar]
- Rekker, R.F. The Hydrophobic Fragmental Constant: Its Derivation and Application. A Means of Characterizing Membrane Systems; Elsevier: Amsterdam, The Netherlands, 1977. [Google Scholar]
- Hansch, C.; Leo, A. Exploring QSAR: Fundamentals and Applications in Chemistry and Biology; American Chemical Society: Washington, DC, USA, 1995. [Google Scholar]
- Shahzadi, I.; Nazir, I.; Phan, T.N.Q.; Bernkop-Schnürch, A. About the impact of superassociation of hydrophobic ion pairs on membrane permeability. Eur. J. Pharm. Biopharm. 2020, 151, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Stockert, J.C.; Durantini, E.N.; Gonzalez Lopez, E.J.; Durantini, J.E.; Villanueva, A.; Horobin, R.W. Fluorescence labeling of mitochondria in living cells by the cationic photosensitizer ZnTM2,3PyPz, and the possible role of pseudobase formation and redox processes in facilitating dye uptake. Biotech. Histochem. 2022, 97, 473–479. [Google Scholar] [CrossRef] [PubMed]
- Schmid, S.L.; Sorkin, A. Endocytosis; Cold Spring Harbor Laboratory Press: Cold Spring Harbor, NY, USA, 2014. [Google Scholar]
- Kumari, S.; Swetha, M.G.; Mayor, S. Endocytosis unplugged: Multiple ways to enter the cell. Cell Res. 2010, 20, 256–275. [Google Scholar] [CrossRef] [PubMed]
- Rennick, J.J.; Johnston, A.P.R.; Parton, R.G. Key principles and methods for studying the endocytosis of biological and nanoparticle therapeutics. Nat. Nanotechnol. 2021, 16, 266–276. [Google Scholar] [CrossRef] [PubMed]
- Dragsten, P.R.; Webb, W.W. Mechanism of the membrane potential sensitivity of the fluorescent membrane probe merocyanine 540. Biochemistry 1978, 17, 5228–5240. [Google Scholar] [CrossRef] [PubMed]
- Canton, I.; Battaglia, G. Polymersomes-mediated delivery of fluorescent probes for targeted and long-term imaging in live cell microscopy. In Cellular and Subcellular Nanotechnology: Methods and Protocols; Weissig, V., Elbayoumi, T., Olsen, M., Eds.; Springer Science & Business Media: New York, NY, USA, 2013; pp. 343–351. [Google Scholar] [CrossRef]
- Gafurova, C.R.; Tsentsevitsky, A.N.; Petrov, A.M. Frequency-dependent engagement of synaptic vesicle pools in the mice motor nerve terminals. Cell. Mol. Neurobiol. 2023, 43, 729–739. [Google Scholar] [CrossRef] [PubMed]
- Jilinková, A.; Malínská, K.; Petrášek, J. Using FM dyes to study endomembranes and their dynamics in plants and cell suspensions. In Methods in Molecular Biology, Plant Cell Morphogenesis: Methods and Protocols; Humana: New York, NY, USA, 2019; Volume 1992, pp. 173–187. [Google Scholar] [CrossRef]
- Klymchenko, A.S.; Kreder, R. Fluorescent probes for lipid rafts: From model membranes to living cells. Chem. Biol. 2014, 21, 97–113. [Google Scholar] [CrossRef] [PubMed]
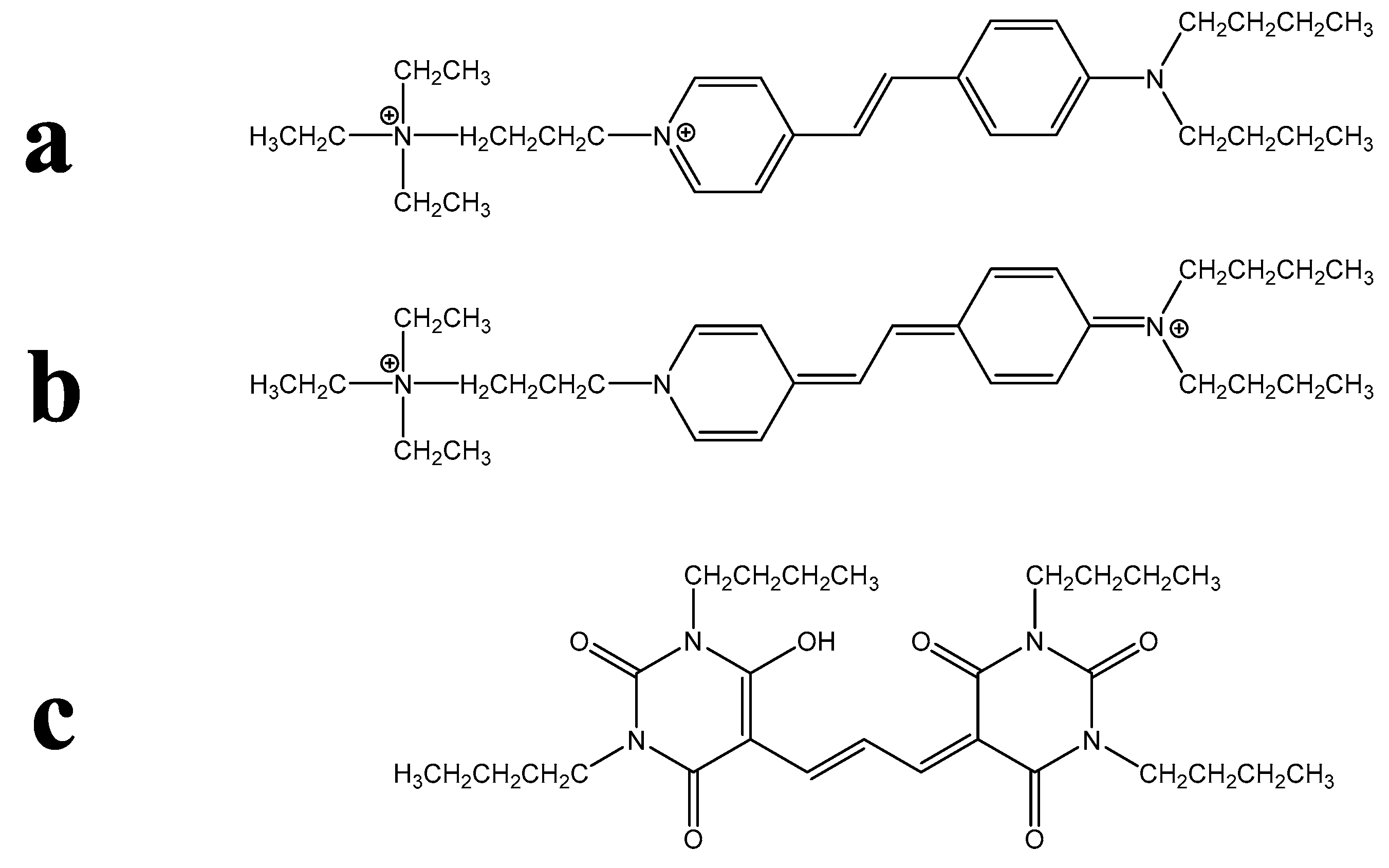

| Surfactant Class or Example of Class | Structure Parameters | ||
|---|---|---|---|
| AI (Typical Chain Length of R) | HGH | Z | |
| Ionised straight-chain fatty acids: R–CO2− M+ | 5.8–11.2 (C10–20) | −5.2 | 1− |
| Linear alkyl-benzene sulfonates: R-C6H4-SO3−M+ | 8.5 (C12) | −4.5 | 1− |
| Long chain amines and their salts: R–NH3+ X− | 5.5–8.7 (C12–18) | −2.2 | 0 |
| −3.4 | 1+ | ||
| Quaternary ammonium salts, e.g., R–N+(CH3)3 X− | 7.6 (C22, behenyl) | −4.1 | 1+ |
| Quaternary ammonium salts, e.g., R–N+(CH2C6H5)(CH3)2 Cl− | 5.2 (C16, cetyl) | −1.5 | 1+ |
| N-alkyl pyrrolidones | 4.7–8.5 (C8–12) | −1.0 | 0 |
| Alkyl polyglycosides | 5.8–7.9 (C10–14) | −3.4 | 0 |
| β-N-Alkylaminopropionic acids: R+H2CH2CH2CO2− | 5.5 (C12) | −3.6 | 0 |
| α-Sulfofatty acid methyl esters: R–CH(SO3−)CO2CH3 M+ | 6.2–9.4 (C12–18) | −2.2 | 1− |
| N-Acyl-L-glutamates: R–CONH–CH(CO2−)CH2CH2CO2− 2M+ | 6.2 and 10.1 (C12 and C18) | −7.2 | 2− |
| N-Lauryl sarcosinate: C11H23–CON(CH3)CH2CO2− M+ | 6.3 (C11) | −3.2 | 1− |
| Name (Type of Drug) | Structure Parameters | |||
|---|---|---|---|---|
| AI | HGH | Log P | Z | |
| Acetobutolol (antihypertensive) | 1.9 | −3.0 | −1.1 | 1+ |
| Adiphenine (anticholinergic) | 4.0 | −3.3 | 0.7 | 1+ |
| Amitriptyline (antidepressant) | 5.6 | −4.0 | 1.6 | 1+ |
| Bromodiphenylhydramine (antihistamine) | 5.2 | −3.9 | 1.3 | 1+ |
| Chlorpromazine (antipsychotic) | 4.2 | −2.6 | 1.6 | 1+ |
| Dextropropoxyphene (analgesic) | 4.9 | −3.5 | 1.4 | 1+ |
| Dibucaine (local anesthetic) | 3.9 | −3.5 | 0.4 | 1+ |
| Flupenthixol (tranquilizer) | 5.2 | −3.8 | 1.4 | 1+ |
| Nortriptyline (antidepressant) | 5.7 | −3.7 | 2.0 | 1+ |
| Penicillin G (antibiotic) | 3.6 | −4.2 | −0.6 | 1− |
| Piperidolate (anticholinergic) | 4.3 | −5.2 | −0.9 | 1 |
| Sodium fusidate (antibiotic) | 7.6 | −6.3 | 1.3 | 1− |
| Tetracaine (local anesthetic) | 3.0 | −3.0 | 0.0 | 1+ |
| Thiopental (general anesthetic) | 4.5 | −2.6 | 2.9 | 0 |
| Trifluopromazine (antipsychotic) | 4.7 | −3.4 | 1.3 | 1+ |
| Tripelennamine (antihistamine) | 3.7 | −2.8 | 0.9 | 1+ |
| Name | Structure Parameters | ||
|---|---|---|---|
| AI | HGH | Z | |
| Aliospiroside A | 8.6 | −3.5 | 0 |
| α-Chaconine | 8.0 | −3.5 | 0 |
| Echynocystic acid | 8.8 | −3.6 | 1− |
| Gypsogenin | 7.5 | −3.2 | 1− |
| Phytolaccinic acid | 7.8 | −5.1 | 1− |
| Phytolaccinic acid, 23-O-Ac | 8.7 | −5.1 | 1− |
| Quillaic acid | 6.1 | −3.6 | 1− |
| Quillaic acid, 22β-OH | 7.5 | −5.2 | 1− |
| Soyasaponin Aa | 8.3 | −9.8 | 0 |
| Soyasaponin βg | 10.8 | −6.5 | 0 |
| Steroidal alkaloid saponin | 9.3 | −11.4 | 0 |
| Steroidal saponin | 5.6 | −1.7 | 0 |
| Triterpenoid saponin | 9.7 | −5.9 | 0 |
| Probe Name | Structure Parameters | Cell Target of Probe | |||
|---|---|---|---|---|---|
| AI | HGH | Log P | Z | ||
| 1 | 9.0 | −1.1 | 7.9 | 0 | Thiols |
| 1P | 14.0 | −5.4 | 8.6 | 2− | Plasma membrane marker |
| 8-TBT-8 | 8.3 | −5.4 | 2.9 | 2+ | Plasma membrane marker |
| ACal | 6.3 | −4.4 | 1.9 | 4− | Calcium ions |
| B-2AZ | 16.0 | −8.7 | 7.3 | 0 Zw | Plasma membrane marker |
| C-1 | 6.3 | −3.7 | 3.5 | 1− | Membrane tension |
| C-2 | 7.0 | −3.9 | 4.8 | 1− | Membrane tension |
| Calcium green C18 | 9.5 | −10.2 | −0.7 | 6− | Calcium ions |
| CL | 8.1 | −1.4 | 6.7 | 1− | Membrane tension |
| CS | 9.5 | −1.4 | 8.2 | 1− | Membrane tension |
| C18-Fura-2 | 10.1 | −11.5 | −1.4 | 5− | Calcium ions |
| C-Laurdan | 8.1 | −3.9 | 6.0 | 1− | Membrane microdomain |
| CL-Laurdan | 8.1 | −3.9 | 1.5 | 1− | Membrane microdomain |
| Di-4-ANEPPDHQ | 8.0 | −7.4 | 0.6 | 1+ | Membrane microdomain |
| DSDMHDAB | 9.0 | −9.4 | −0.4 | 2− | Nitric oxide |
| FD-9 | NA | NA | 14.8 | 0 | Plasma membrane marker |
| Flu7 | 12.0 | −8.6 | 3.4 | 2+ | With Q12, enzyme activity |
| FC12SM | 11.8 | −5.0 | 6.8 | 0 Zw | Plasma membrane marker |
| F2N8 | 10.2 | −4.6 | 5.6 | 1+ | Membrane microdomain |
| F2N12S | 11.0 | −5.0 | 6.0 | 0 Zw | Plasma membrane marker |
| F2N12SM | 11.8 | −5.6 | 6.8 | 0 Zw | Plasma membrane marker |
| HGMem-3 | NA | NA | 16.6 | 0 | Hg2+ |
| HOCMem | NA | NA | 18.0 | 0 | HOCl |
| Laurdan | NA | NA | 8.7 | 0 | Plasma membrane marker |
| Mem-5 | NA | NA | 15.7 | 0 | Cu2+ |
| Mem-6 | NA | NA | 12.0 | 0 | Cu2+ |
| MemBright C3 | 14.7 | −4.5 | 10.2 | 1+ | Plasma membrane marker |
| MemBright C3.5 | 12.0 | −4.5 | 12.6 | 1+ | Plasma membrane marker |
| MemBright C5 | 12.0 | −4.5 | 10.9 | 1+ | Plasma membrane marker |
| MemBright C5.5 | 12.0 | −4.5 | 13.3 | 1+ | Plasma membrane marker |
| MemBright C7 | 12.0 | −4.5 | 11.6 | 1+ | Plasma membrane marker |
| MemBright C7.5 | 12.0 | −4.5 | 14.0 | 1+ | Plasma membrane marker |
| Mem-NO | 12.3 | −6.7 | 5.6 | 1+ | Nitric oxide |
| Mem-SQAC | 7.8 | −5.1 | 2.7 | 1+ | Plasma membrane marker |
| N4RA | 8.4 | −4.6 | 3.8 | 1− | Membrane microdomain |
| NR12S | 12.2 | −3.6 | 8.6 | 0 Zw | Membrane microdomain |
| PY3174 | 5.4 | −7.2 | −1.8 | 2+ | Membrane microdomain |
| PY3184 | 5.5 | −7.2 | −1.7 | 2+ | Membrane microdomain |
| PY3304 | 6.4 | −6.8 | −0.4 | 2+ | Membrane microdomain |
| Q12 | 8.3 | ca. −5 | 3.3 | 1− | With Flu-7, enzyme activity |
| SL2 | 5.4 | −1.8 | 3.6 | 1− | Membrane microdomain |
| Structure 1 | 6.0 | −6.1 | −0.1 | 4− | Thioredoxin |
| TPE-MEM | 14.7 | −9.6 | 5.1 | 2+ | Plasma membrane marker |
| TPE-Py-EEGTIGYG | 10.0 | −10.1 | −0.1 | 2− | Cu2+ |
| TTVP | 6.7 | −6.8 | −0.1 | 2+ | Plasma membrane marker |
| ZTRS-C12 | NA | NA | 8.6 | 0 | Zn2+ |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Horobin, R.W.; Stockert, J.C. How to Target Small-Molecule Fluorescent Imaging Probes to the Plasma Membrane—The Influence and QSAR Modelling of Amphiphilicity, Lipophilicity, and Flip-Flop. Molecules 2023, 28, 7589. https://doi.org/10.3390/molecules28227589
Horobin RW, Stockert JC. How to Target Small-Molecule Fluorescent Imaging Probes to the Plasma Membrane—The Influence and QSAR Modelling of Amphiphilicity, Lipophilicity, and Flip-Flop. Molecules. 2023; 28(22):7589. https://doi.org/10.3390/molecules28227589
Chicago/Turabian StyleHorobin, Richard W., and Juan C. Stockert. 2023. "How to Target Small-Molecule Fluorescent Imaging Probes to the Plasma Membrane—The Influence and QSAR Modelling of Amphiphilicity, Lipophilicity, and Flip-Flop" Molecules 28, no. 22: 7589. https://doi.org/10.3390/molecules28227589
APA StyleHorobin, R. W., & Stockert, J. C. (2023). How to Target Small-Molecule Fluorescent Imaging Probes to the Plasma Membrane—The Influence and QSAR Modelling of Amphiphilicity, Lipophilicity, and Flip-Flop. Molecules, 28(22), 7589. https://doi.org/10.3390/molecules28227589