
The Effect of Piperidine Nitroxides on the Properties of Metalloproteins in Human Red Blood Cells


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
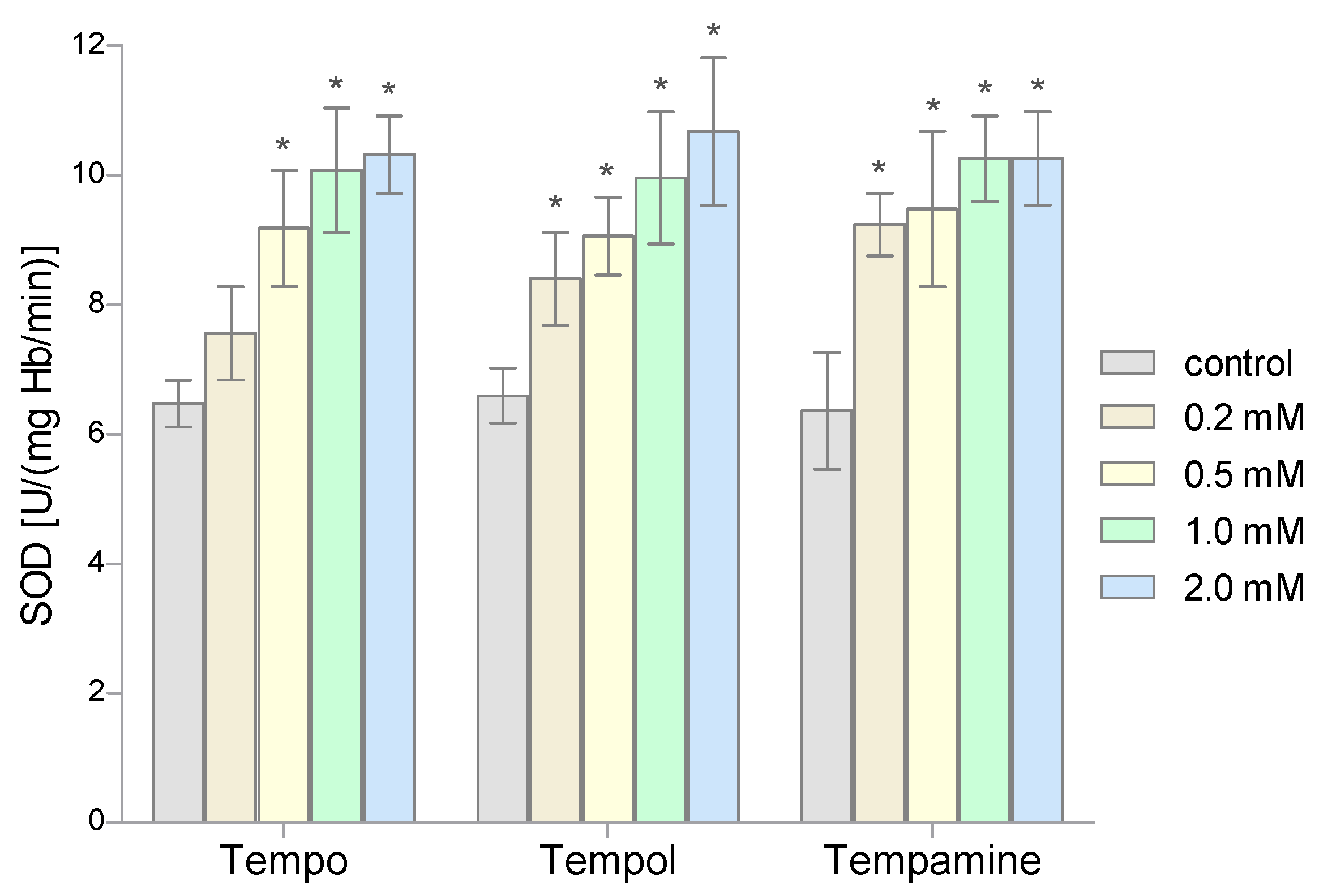
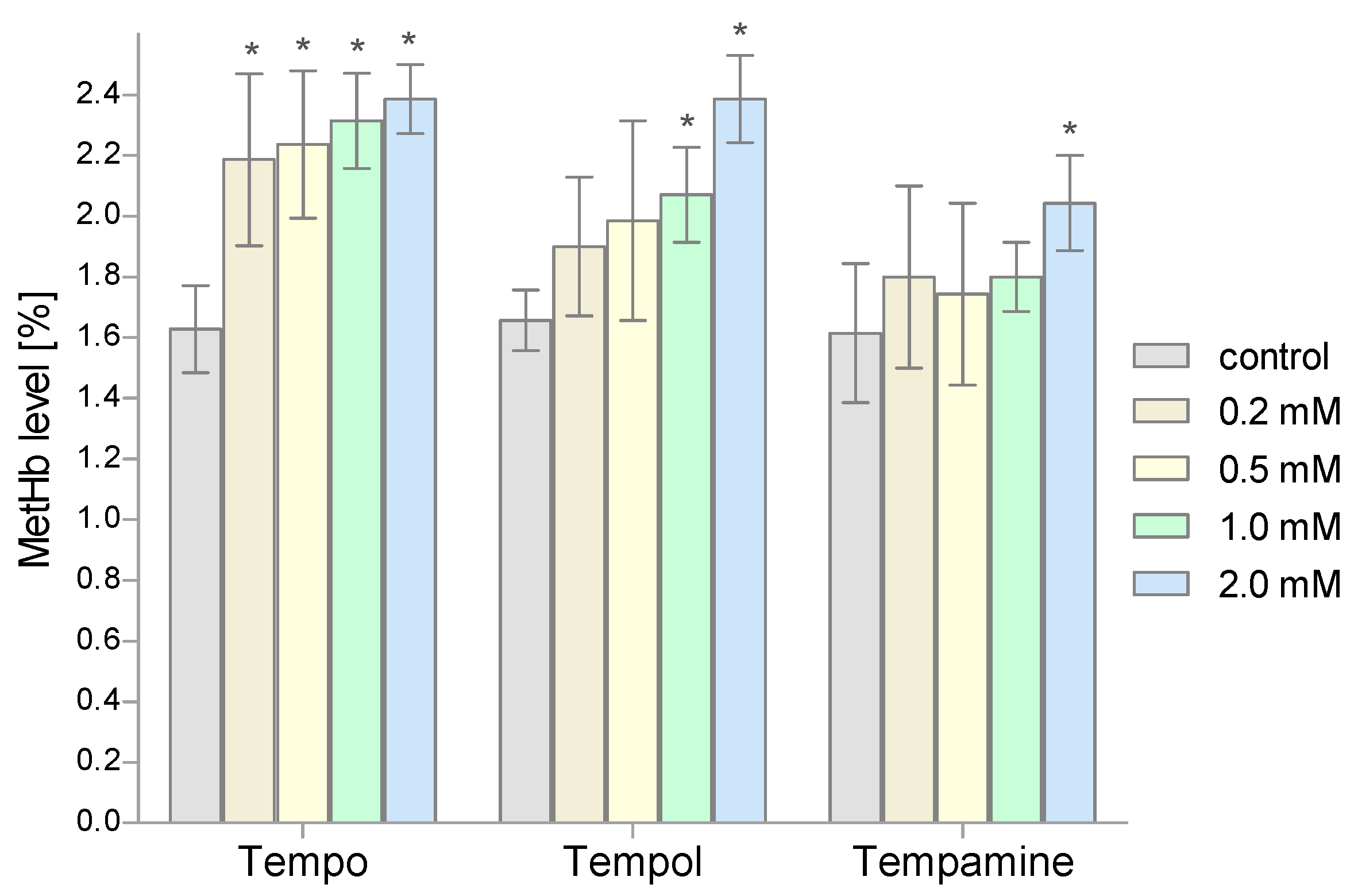
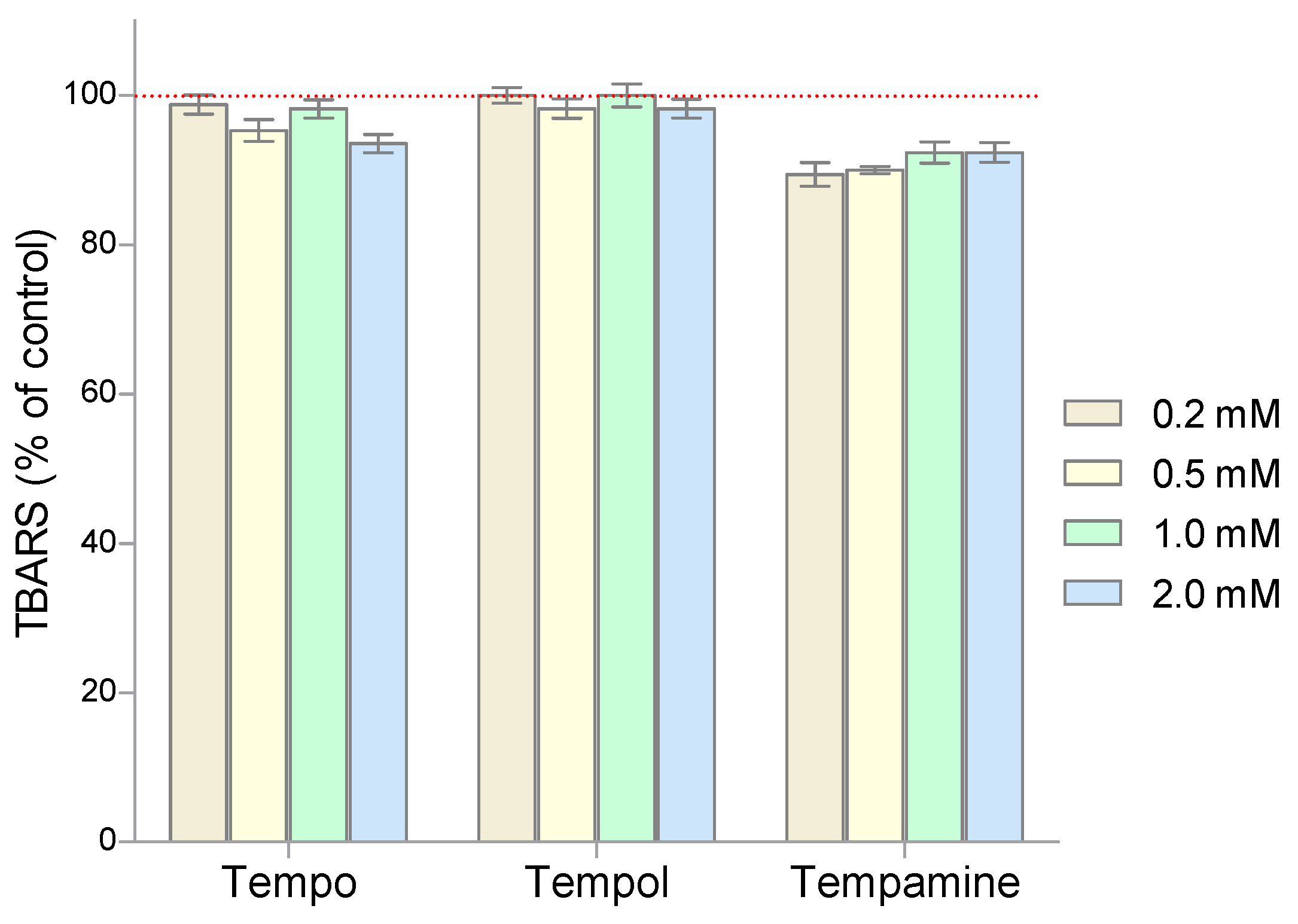
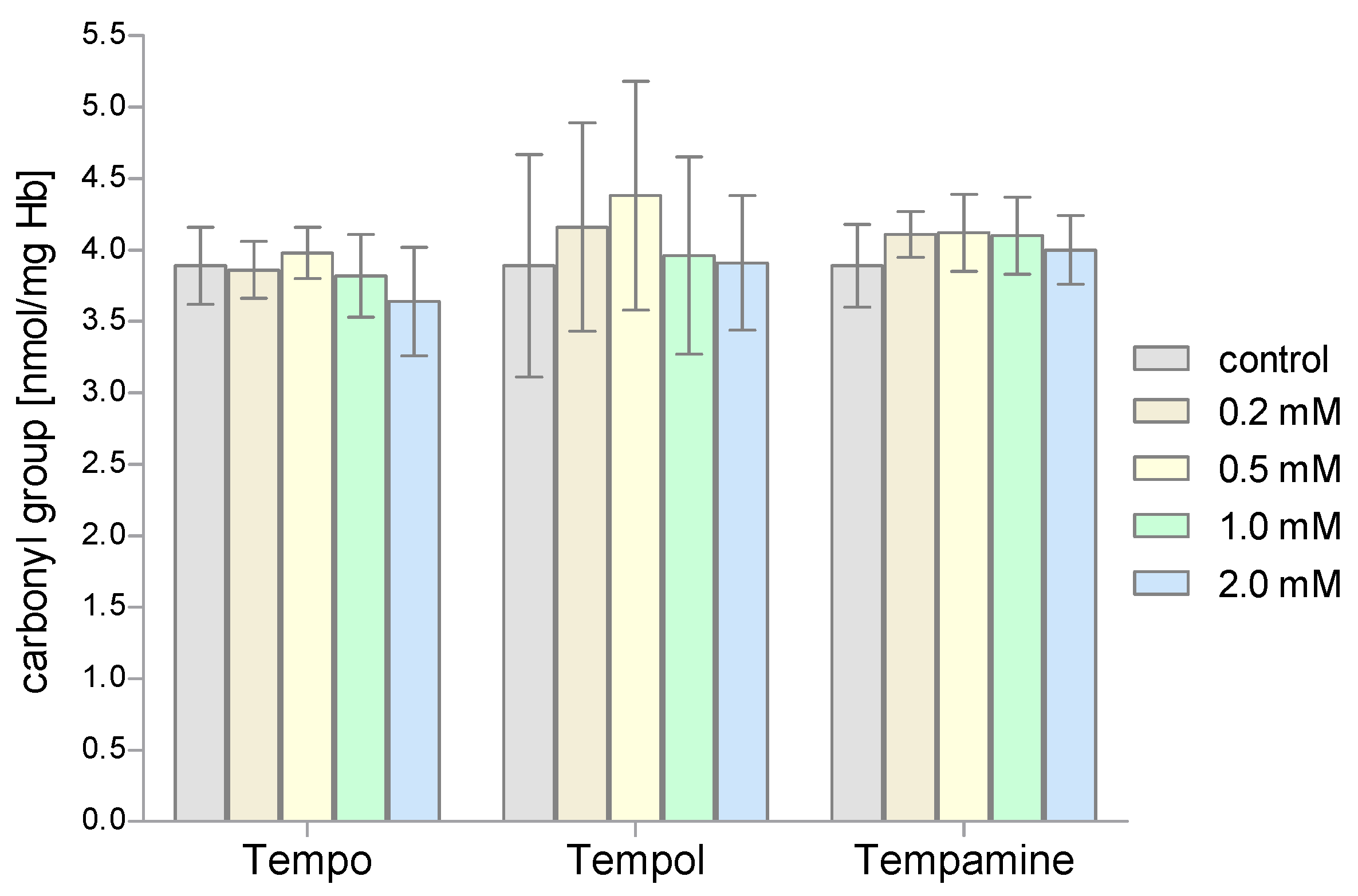
2. Results
3. Discussion
4. Material and Methods
4.1. Chemicals
4.2. Experiment Protocol
4.3. Determination of Ascorbic Acid Level
4.4. Superoxide Dismutase (SOD) Activity Assay
4.5. Determination of Catalase (CAT) Activity
4.6. Determination of Lactate Dehydrogenase (LDH) Activity
4.7. Determination of Thiobarbituric Acid Reactive Substances (TBARS) Level
4.8. Evaluation of Carbonyl Group Concentration
4.9. Determination of the Degree of Hb Autooxidation (% MetHb)
4.10. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Sample Availability
Abbreviations
| RBCs | red blood cells (erythrocytes) |
| Hb | hemoglobin |
| Mb | myoglobin |
| MetHb | methemoglobin |
| TBARS | thiobarbituric acid reactive substances |
| LDH | lactate dehydrogenase |
| SOD | superoxide dismutase |
| CAT | catalase |
References
- Soule, B.P.; Hyodo, F.; Matsumoto, K.-I.; Simone, N.L.; Cook, J.A.; Krishna, M.C.; Mitchell, J.B. The chemistry and biology of nitroxide compounds. Free Radic. Biol. Med. 2007, 42, 1632–1650. [Google Scholar] [CrossRef] [PubMed]
- Haidasz, E.A.; Meng, D.; Amorati, R.; Baschieri, A.; Ingold, K.U.; Valgimigli, L.; Pratt, D.A. Acid Is Key to the Radical-Trapping Antioxidant Activity of Nitroxides. J. Am. Chem. Soc. 2016, 138, 5290–5298. [Google Scholar] [CrossRef] [PubMed]
- Cimato, A.N.; Piehl, L.L.; Facorro, G.B.; Torti, H.B.; Hager, A.A. Antioxidant effects of water- and lipid-soluble nitroxide radicals in liposomes. Free Radic. Biol. Med. 2004, 37, 2042–2051. [Google Scholar] [CrossRef]
- Offer, T.; Samuni, A. Nitroxides inhibit peroxyl radical-mediated DNA scission and enzyme inactivation. Free Radic. Biol. Med. 2002, 32, 872–881. [Google Scholar] [CrossRef] [PubMed]
- Fedeli, D.; Damiani, E.; Greci, L.; Littarru, G.P.; Falcioni, G. Nitroxide radicals protect against DNA damage in rat epithelial cells induced by nitric oxide, nitroxyl anion and peroxynitrite. Mutat. Res. 2003, 535, 117–125. [Google Scholar] [CrossRef] [PubMed]
- Damiani, E.; Castagna, R.; Greci, L. The effects of derivatives of the nitroxide tempol on UVA-mediated in vitro lipid and protein oxidation. Free Radic. Biol. Med. 2002, 33, 128–136. [Google Scholar] [CrossRef] [PubMed]
- Hahn, S.M.; Sullivan, F.J.; DeLuca, A.M.; Krishna, C.M.; Wersto, N.; Venzon, D.; Russo, A.; Mitchell, J.B. Evaluation of tempol radioprotection in a murine tumor model. Free Radic. Biol. Med. 1997, 22, 1211–1216. [Google Scholar] [CrossRef] [PubMed]
- Hahn, S.M.; Tochner, Z.; Krishna, C.M.; Glass, J.; Wilson, L.; Samuni, A.; Sprague, M.; Venzon, D.; Glatstein, E.; Mitchell, J.B. Tempol, a stable free radical, is a novel murine radiation protector. Cancer Res. 1992, 52, 1750–1753. [Google Scholar]
- Lewandowski, M.; Gwoździński, K. Photoprotective and radioprotective properties of nitroxides and their application in magnetic resonance imaging. Postep. Hig. Med. Dosw. Online 2016, 70, 1101–1111. [Google Scholar] [CrossRef]
- Gariboldi, M.B.; Ravizza, R.; Petterino, C.; Castagnaro, M.; Finocchiaro, G.; Monti, E. Study of in vitro and in vivo effects of the piperidine nitroxide Tempol--a potential new therapeutic agent for gliomas. Eur. J. Cancer 2003, 39, 829–837. [Google Scholar] [CrossRef]
- Samuni, A.M.; DeGraff, W.; Krishna, M.C.; Mitchell, J.B. Nitroxides as antioxidants: Tempol protects against EO9 cytotoxicity. Mol. Cell. Biochem. 2002, 234–235, 327–333. [Google Scholar] [CrossRef]
- Gelvan, D.; Saltman, P.; Powell, S.R. Cardiac reperfusion damage prevented by a nitroxide free radical. Proc. Natl. Acad. Sci. USA 1991, 88, 4680–4684. [Google Scholar] [CrossRef]
- Guo, R.; Gao, X.-Y.; Wang, W.; Wang, H.-J.; Zhang, F.; Zhang, Y.; Zhu, G.-Q. Tempol reduces reperfusion-induced arrhythmias in anaesthetized rats. Pharmacol. Res. 2005, 52, 192–198. [Google Scholar] [CrossRef] [PubMed]
- Czepas, J.; Koceva-Chyła, A.; Gwoździński, K.; Jóźwiak, Z. Different effectiveness of piperidine nitroxides against oxidative stress induced by doxorubicin and hydrogen peroxide. Cell Biol. Toxicol. 2008, 24, 101–112. [Google Scholar] [CrossRef] [PubMed]
- Monti, E.; Cova, D.; Guido, E.; Morelli, R.; Oliva, C. Protective effect of the nitroxide tempol against the cardiotoxicity of adriamycin. Free Radic. Biol. Med. 1996, 21, 463–470. [Google Scholar] [CrossRef] [PubMed]
- Zhang, R.; Pinson, A.; Samuni, A. Both hydroxylamine and nitroxide protect cardiomyocytes from oxidative stress. Free Radic. Biol. Med. 1998, 24, 66–75. [Google Scholar] [CrossRef]
- Suy, S.; Mitchell, J.B.; Samuni, A.; Mueller, S.; Kasid, U. Nitroxide tempo, a small molecule, induces apoptosis in prostate carcinoma cells and suppresses tumor growth in athymic mice. Cancer 2005, 103, 1302–1313. [Google Scholar] [CrossRef]
- Koceva-Chyła, A.; Kochman, A.; Głebska, J.; Gwoździnski, K.; Jóźwiak, Z.; Metodiewa, D. Tempicol-3, a novel piperidine-N-oxide stable radical and antioxidant, with low toxicity acts as apoptosis inducer and cell proliferation modifier of Yoshida Sarcoma cells in vivo. Anticancer Res. 2000, 20, 4611–4618. [Google Scholar]
- Głebska, J.; Skolimowski, J.; Kudzin, Z.; Gwoździński, K.; Grzelak, A.; Bartosz, G. Pro-oxidative activity of nitroxides in their reactions with glutathione. Free Radic. Biol. Med. 2003, 35, 310–316. [Google Scholar] [CrossRef]
- Offer, T.; Russo, A.; Samuni, A. The pro-oxidative activity of SOD and nitroxide SOD mimics. FASEB J. 2000, 14, 1215–1223. [Google Scholar] [CrossRef]
- Mohanty, J.G.; Nagababu, E.; Rifkind, J.M. Red blood cell oxidative stress impairs oxygen delivery and induces red blood cell aging. Front. Physiol. 2014, 5, 84. [Google Scholar] [CrossRef] [PubMed]
- Mal, A.; Chatterjee, I.B. Mechanism of autoxidation of oxyhaemoglobin. J. Biosci. 1991, 16, 55–70. [Google Scholar] [CrossRef]
- Misra, H.P.; Fridovich, I. The generation of superoxide radical during the autoxidation of hemoglobin. J. Biol. Chem. 1972, 247, 6960–6962. [Google Scholar] [CrossRef] [PubMed]
- Ogura, Y. Catalase activity at high concentration of hydrogen peroxide. Arch. Biochem. Biophys. 1955, 57, 288–300. [Google Scholar] [CrossRef] [PubMed]
- Ladi, R.N.; Hollaar, L.; Souverijn, J.H.; van der Laarse, A. Quantitation of cumulative release of lactate dehydrogenase isoenzyme-1 in plasma of patients with acute myocardial infarction using a commercially available test. Clin. Physiol. Biochem. 1990, 8, 250–255. [Google Scholar] [PubMed]
- Heidari Beigvand, H.; Heidari, K.; Hashemi, B.; Saberinia, A. The Value of Lactate Dehydrogenase in Predicting Rhabdomyolysis-Induced Acute Renal Failure; a Narrative Review. Arch. Acad. Emerg. Med. 2021, 9, e24. [Google Scholar] [CrossRef] [PubMed]
- Lim, S.C.; Kim, Y.I. The role of the lactate dehydrogenase and the effect of prone position during ventilator-induced lung injury. J. Korean Med. Sci. 2004, 19, 223–228. [Google Scholar] [CrossRef] [PubMed]
- Price, J.R.; Hagrass, H.; Filip, A.B.; McGill, M.R. LDH and the MELD-LDH in Severe Acute Liver Injury and Acute Liver Failure: Preliminary Confirmation of a Novel Prognostic Score for Risk Stratification. J. Appl. Lab. Med. 2023, 8, 504–513. [Google Scholar] [CrossRef]
- Eriksson, U.G.; Tozer, T.N.; Sosnovsky, G.; Lukszo, J.; Brasch, R.C. Human erythrocyte membrane permeability and nitroxyl spin-label reduction. J. Pharm. Sci. 1986, 75, 334–337. [Google Scholar] [CrossRef]
- Gwozdzinski, K.; Bartosz, G. Nitroxide reduction in human red blood cells. Curr. Top. Biophys. 1996, 20, 60–65. [Google Scholar]
- Bujak, S.; Gwozdzinski, K. Nitroxides lead to reduced level of glutathione in red blood cells. In Free Radicals and Oxidative Stress: Chemistry, Biochemistry and Pathological Implications; Galaris, G., Ed.; Medimond International Proceedings: Bologna, Italy, 2003; pp. 105–108. [Google Scholar]
- Głębska, J.; Gwozdzinski, K. Oxygen–dependent reduction of nitroxides by ascorbic acid and glutathione. An EPR study. Curr. Top. Biophys. 1998, 22 (Suppl. B), 75–82. [Google Scholar]
- Balcerczyk, A.; Łuczak, K.; Soszyński, M.; Bartosz, G. Prooxidative effects of TEMPO on human erythrocytes. Cell Biol. Int. 2004, 28, 585–591. [Google Scholar] [CrossRef] [PubMed]
- Bujak, S.; Gwozdzinski, K. The effect of stable nitroxide radicals on the activity of glutathione-dependent enzymes: Reductase, peroxidase and transferase in human erythrocytes. Medimond Int. Proc. 2006, 71–77. [Google Scholar]
- Zhang, Y.; Fung, L.W. The roles of ascorbic acid and other antioxidants in the erythrocyte in reducing membrane nitroxide radicals. Free Radic. Biol. Med. 1994, 16, 215–222. [Google Scholar] [CrossRef] [PubMed]
- Krishna, M.C.; Grahame, D.A.; Samuni, A.; Mitchell, J.B.; Russo, A. Oxoammonium cation intermediate in the nitroxide-catalyzed dismutation of superoxide. Proc. Natl. Acad. Sci. USA 1992, 89, 5537–5541. [Google Scholar] [CrossRef] [PubMed]
- Samuni, A.; Krishna, C.M.; Mitchell, J.B.; Collins, C.R.; Russo, A. Superoxide reaction with nitroxides. Free Radic. Res. Commun. 1990, 9, 241–249. [Google Scholar] [CrossRef] [PubMed]
- Goldstein, S.; Samuni, A.; Hideg, K.; Merenyi, G. Structure-activity relationship of cyclic nitroxides as SOD mimics and scavengers of nitrogen dioxide and carbonate radicals. J. Phys. Chem. A 2006, 110, 3679–3685. [Google Scholar] [CrossRef] [PubMed]
- Golestani, A.; Rastegar, R.; Shariftabrizi, A.; Khaghani, S.; Payabvash, S.M.; Salmasi, A.H.; Dehpour, A.R.; Pasalar, P. Paradoxical dose- and time-dependent regulation of superoxide dismutase and antioxidant capacity by vitamin E in rat. Clin. Chim. Acta 2006, 365, 153–159. [Google Scholar] [CrossRef]
- Seth, V.; Banerjee, B.D.; Chakravorty, A.K. Lipid Peroxidation, Free Radical Scavenging Enzymes, and Glutathione Redox System in Blood of Rats Exposed to Propoxur. Pestic. Biochem. Physiol. 2001, 71, 133–139. [Google Scholar] [CrossRef]
- Láng, I.; Deák, G.; Müzes, G.; Prónai, L.; Fehér, J. Effect of the natural bioflavonoid antioxidant silymarin on superoxide dismutase (SOD) activity and expression in vitro. Biotechnol. Ther. 1993, 4, 263–270. [Google Scholar]
- Yan, S.; Hong, X.; Hu, Y.; Liao, K. Tempol, one of nitroxides, is a novel ultraviolet-A1 radiation protector for human dermal fibroblasts. J. Dermatol. Sci. 2005, 37, 137–143. [Google Scholar] [CrossRef] [PubMed]
- Hahn, S.M.; Krishna, M.C.; DeLuca, A.M.; Coffin, D.; Mitchell, J.B. Evaluation of the hydroxylamine Tempol-H as an in vivo radioprotector. Free Radic. Biol. Med. 2000, 28, 953–958. [Google Scholar] [CrossRef] [PubMed]
- DeJulius, C.R.; Dollinger, B.R.; Kavanaugh, T.E.; Dailing, E.; Yu, F.; Gulati, S.; Miskalis, A.; Zhang, C.; Uddin, J.; Dikalov, S.; et al. Optimizing an Antioxidant TEMPO Copolymer for Reactive Oxygen Species Scavenging and Anti-Inflammatory Effects in Vivo. Bioconjugate Chem. 2021, 32, 928–941. [Google Scholar] [CrossRef]
- Samuni, A.M.; Lipman, A.; Barenholz, Y. Damage to liposomal lipids: Protection by antioxidants and cholesterol-mediated dehydration. Chem. Phys. Lipids 2000, 105, 121–134. [Google Scholar] [CrossRef] [PubMed]
- Samuni, A.M.; Barenholz, Y. Stable nitroxide radicals protect lipid acyl chains from radiation damage. Free Radic. Biol. Med. 1997, 22, 1165–1174. [Google Scholar] [CrossRef] [PubMed]
- Young, A.J.; Lowe, G.M. Antioxidant and prooxidant properties of carotenoids. Arch. Biochem. Biophys. 2001, 385, 20–27. [Google Scholar] [CrossRef] [PubMed]
- Samuni, A.; Goldstein, S. Hydroxylamines inhibit tyrosine oxidation and nitration: The role of their respective nitroxide radicals. Free Radic. Biol. Med. 2020, 160, 837–844. [Google Scholar] [CrossRef] [PubMed]
- Zareba, M.; Widomska, J.; Burke, J.M.; Subczynski, W.K. Nitroxide free radicals protect macular carotenoids against chemical destruction (bleaching) during lipid peroxidation. Free Radic. Biol. Med. 2016, 101, 446–454. [Google Scholar] [CrossRef]
- Krishna, M.C.; Samuni, A.; Taira, J.; Goldstein, S.; Mitchell, J.B.; Russo, A. Stimulation by nitroxides of catalase-like activity of hemeproteins. Kinetics and mechanism. J. Biol. Chem. 1996, 271, 26018–26025. [Google Scholar] [CrossRef]
- Mehlhorn, R.J.; Swanson, C.E. Nitroxide-stimulated H2O2 decomposition by peroxidases and pseudoperoxidases. Free Radic. Res. Commun. 1992, 17, 157–175. [Google Scholar] [CrossRef]
- Krishna, M.C.; Halevy, R.F.; Zhang, R.; Gutierrez, P.L.; Samuni, A. Modulation of streptonigrin cytotoxicity by nitroxide SOD mimics. Free Radic. Biol. Med. 1994, 17, 379–388. [Google Scholar] [CrossRef] [PubMed]
- Falcioni, G.; Gabbianelli, R.; Damiani, E.; Santroni, A.M.; Fedeli, D.; Wozniak, M.; Greci, L. The effect of indolinic and quinolinic nitroxide radicals on trout erythrocytes exposed to oxidative stress. Free Radic. Res. 1998, 28, 507–516. [Google Scholar] [CrossRef] [PubMed]
- Polnaszek, C.F.; Schreier, S.; Butler, K.W.; Smith, I.C.P. Analysis of the factors determining the EPR spectra of spin probes that partition between aqueous and lipid phases. J. Am. Chem. Soc. 1978, 100, 8223–8232. [Google Scholar] [CrossRef]
- Liong, E.C.; Dou, Y.; Scott, E.E.; Olson, J.S.; Phillips, G.N. Waterproofing the heme pocket. Role of proximal amino acid side chains in preventing hemin loss from myoglobin. J. Biol. Chem. 2001, 276, 9093–9100. [Google Scholar] [CrossRef] [PubMed]
- Yamaguchi, T.; Nakano, T.; Kimoto, E. Oxidation of nitroxide radicals by the reaction of hemoglobin with hydrogen peroxide. Biochem. Biophys. Res. Commun. 1984, 120, 534–539. [Google Scholar] [CrossRef] [PubMed]
- Kendig, D.M.; Tarloff, J.B. Inactivation of lactate dehydrogenase by several chemicals: Implications for in vitro toxicology studies. Toxicol. Vitr. 2007, 21, 125–132. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Faivre, B.; Menu, P.; Labrude, P.; Vigneron, C. Hemoglobin autooxidation/oxidation mechanisms and methemoglobin prevention or reduction processes in the bloodstream. Literature review and outline of autooxidation reaction. Artif. Cells Blood Substit. Immobil. Biotechnol. 1998, 26, 17–26. [Google Scholar] [CrossRef]
- Kopperschläger, G.; Kirchberger, J. Methods for the separation of lactate dehydrogenases and clinical significance of the enzyme. J. Chromatogr. B Biomed. Appl. 1996, 684, 25–49. [Google Scholar] [CrossRef]
- Balcerczyk, A.; Grzelak, A.; Soszynski, M.; Bartosz, G. Pro-oxidative effects of Tempo in systems containing oxidants. Redox Rep. 2004, 9, 153–159. [Google Scholar] [CrossRef]
- Mustafa, A.G.; Bani-Ahmad, M.A.; Jaradat, A.Q.; Allouh, M.Z. Tempol protects blood proteins and lipids against peroxynitrite-mediated oxidative damage. Exp. Biol. Med. 2015, 240, 109–112. [Google Scholar] [CrossRef]
- Drabkin, D.L. Spectrophotometric studies; the crystallographic and optical properties of the hemoglobin of man in comparison with those of other species. J. Biol. Chem. 1946, 164, 703–723. [Google Scholar] [CrossRef] [PubMed]
- Omaye, S.T.; Turnbull, J.D.; Sauberlich, H.E. Selected methods for the determination of ascorbic acid in animal cells, tissues, and fluids. Methods Enzymol. 1979, 62, 3–11. [Google Scholar] [CrossRef] [PubMed]
- Aebi, H. Catalase in vitro. Methods Enzymol. 1984, 105, 121–126. [Google Scholar] [CrossRef] [PubMed]
- Wroblewski, F.; Ladue, J.S. Lactic dehydrogenase activity in blood. Proc. Soc. Exp. Biol. Med. 1955, 90, 210–213. [Google Scholar] [CrossRef] [PubMed]
- Stocks, J.; Dormandy, T.L. The autoxidation of human red cell lipids induced by hydrogen peroxide. Br. J. Haematol. 1971, 20, 95–111. [Google Scholar] [CrossRef] [PubMed]
- Levine, R.L.; Garland, D.; Oliver, C.N.; Amici, A.; Climent, I.; Lenz, A.G.; Ahn, B.W.; Shaltiel, S.; Stadtman, E.R. Determination of carbonyl content in oxidatively modified proteins. Methods Enzymol. 1990, 186, 464–478. [Google Scholar] [CrossRef]








Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bujak-Pietrek, S.; Pieniazek, A.; Gwozdzinski, K.; Gwozdzinski, L. The Effect of Piperidine Nitroxides on the Properties of Metalloproteins in Human Red Blood Cells. Molecules 2023, 28, 6174. https://doi.org/10.3390/molecules28166174
Bujak-Pietrek S, Pieniazek A, Gwozdzinski K, Gwozdzinski L. The Effect of Piperidine Nitroxides on the Properties of Metalloproteins in Human Red Blood Cells. Molecules. 2023; 28(16):6174. https://doi.org/10.3390/molecules28166174
Chicago/Turabian StyleBujak-Pietrek, Stella, Anna Pieniazek, Krzysztof Gwozdzinski, and Lukasz Gwozdzinski. 2023. "The Effect of Piperidine Nitroxides on the Properties of Metalloproteins in Human Red Blood Cells" Molecules 28, no. 16: 6174. https://doi.org/10.3390/molecules28166174
APA StyleBujak-Pietrek, S., Pieniazek, A., Gwozdzinski, K., & Gwozdzinski, L. (2023). The Effect of Piperidine Nitroxides on the Properties of Metalloproteins in Human Red Blood Cells. Molecules, 28(16), 6174. https://doi.org/10.3390/molecules28166174