Abstract
Atherosclerosis is the major cause of cardiovascular-disease-related death worldwide, resulting from the subendothelial accumulation of lipoprotein-derived cholesterol, ultimately leading to chronic inflammation and the formation of clinically significant atherosclerotic plaques. Oligosaccharides have been widely used in biomedical research and therapy, including tissue engineering, wound healing, and drug delivery. Moreover, oligosaccharides have been consumed by humans for centuries, and are cheap, and available in large amounts. Given the constantly increasing number of obesity, diabetes, and hyperlipidaemia cases, there is an urgent need for novel therapeutics that can economically and effectively slow the progression of atherosclerosis. In this review, we address the current state of knowledge in oligosaccharides research, and provide an update of the recent in vitro and in vivo experiments that precede clinical studies. The application of oligosaccharides could help to eliminate the residual risk after the application of other cholesterol-lowering medicines, and provide new therapeutic opportunities to reduce the associated burden of premature deaths because of atherosclerosis.
1. Introduction
Atherosclerosis is the major cause of various atherosclerotic cardiovascular diseases, such as peripheral artery disease, ischaemic stroke, and coronary artery disease. As a chronic inflammatory disease, atherosclerosis is characterised by a pathological lipid accumulation, and remodelling of the arterial wall. The outcome of atherosclerosis depends on the interplay between many genetic and environmental risk factors, with the degree of inflammation and hyperlipidaemia playing the main role in its pathogenesis [1]. In total, advanced atherogenesis and its complications cause 17.9 million deaths or 32% of all deaths per year worldwide [2].
Atherosclerosis results from an influx of cholesterol-rich low-density lipoprotein (LDL) particles, coated with apolipoprotein B (ApoB) protein, in the intimal layer of the arterial wall in atherosclerosis-susceptible regions of the arterial tree, such as in arterial bifurcations [3]. In the intimal layer, the LDL particles are subjected to various proteolytic and lipolytic enzymes, and oxidative agents, thus forming multiple modified low-density lipoprotein (mmLDL) particles. An MmLDL accumulation in the intima of blood vessels causes the dysfunction of endothelial cells (ECs), which is characterised by an increased expression of inflammatory mediators (such as interleukins (IL-1β, IL-6), TNF-α, and interferon-gamma (IFN-γ)) and adhesion molecules (such as ICAM (Intercellular Adhesion Molecule-1), VCAM (Vascular Cell Adhesion Molecule-1), and P- and E-selectins), and thus promotes inflammation and an increasing leukocyte adhesiveness. Further, macrophages uptake mmLDL and form foam cells, which facilitate inflammation, and stimulate humoral and adaptive immunity [4,5].
Oxidative stress is another essential player in the initiation and progression of endothelial dysfunction and atherosclerosis [6]. An increase in ROS (reactive oxygen species), and insufficient antioxidant production, facilitate lipid deposition, and promote inflammation and endothelial injury [7]. There are several other processes associated with the disease’s progression. The transition of epithelial cells to mesenchymal cells, and the differentiation of fibroblasts to myofibroblasts under the influence of inflammatory cytokines and pro-apoptotic regulators is known to reinforce endothelial dysfunction and plaque formation. Furthermore, the plaque at advanced stages could necrotise macrophages and smooth muscle cells, thus resulting in the formation of a plaque necrotic core, often covered by a fibrous cap, which is prone to rupture and cause thrombosis and vessel occlusion [8]. Additionally, the dysregulation of Ca2+ homeostasis in the plaque initiates a mineralisation process similar to bone formation, which leads to plaque calcification [9]. Extended calcifications are known to stabilise the plaque, while spotty calcifications increase the plaque’s vulnerability and chance of rupturing [10]. Therefore, an efficient anti-atherosclerotic treatment is usually based on lipid-lowering, antioxidant, and/or anti-inflammatory properties [11].
Oligosaccharides are formed by a small number (usually 3–10) of monosaccharide monomers, and are associated with various biological functions (cell adhesion and recognition, carbohydrate storage and transport, prebiotic effects). Oligosaccharides are normally linked to lipids or to amino acids via N- or O-glycosidic bonds, thus creating glycolipids or glycoproteins, respectively [12]. Other forms of oligosaccharides (not linked to lipids or amino acids) may be presented, for example, by the plant raffinose series, which perform the storage function, or maltodextrins and cellodextrins, which result from the microbial breakdown of larger polysaccharides, such as starch or cellulose [13]. In addition to the wide application of natural oligosaccharides (such as fructo-oligosaccharides (FOSs), galacto-oligosaccharides (GOSs), human milk oligosaccharides (HMOs), Mannan oligosaccharides (MOSs) and others) in human diets and animal feed [14], different synthetic analogues of natural oligosaccharides, created through a combination of modern chemical synthesis, computational, and analytical methods, have demonstrated high potential for applications in biology, nanotechnology, material science, and medicine [15].
Further on in this review, we focus on the recent studies investigating the molecular mechanisms of oligosaccharides and their derivatives used to treat atherosclerosis. The application of polysaccharides as therapeutic agents to treat atherosclerosis was recently reviewed [16,17] and, therefore, will be excluded from our review. Additionally, the oligosaccharide-gut microbiota axis has been extensively reviewed in several recent papers, towards which we wish to redirect interested readers [18,19,20].
2. Alginate Oligosaccharide
Alginate is a polysaccharide, the major component of the brown algae cell wall (approximately 40% of the dry weight), formed by the monomeric units of β-D-mannuronate and α-L-guluronate, linked through a β-1,4-glycosidic bond. Alginate oligosaccharide (AOS) is delivered via the de-polymerisation (for example, via acid hydrolysis, oxidation, and enzymatic degradation) of alginate, and is widely used in a variety of fields, including drug delivery and tissue engineering applications [21]. The unique advantageous properties (such as being water-soluble, biodegradable, non-toxic, and non-immunogenic) and activities (anti-inflammatory, anti-proliferative, antioxidative and anti-apoptotic) of AOS explain the growing interest it is receiving from the scientific community [22].
Recent in silico and in vitro investigations showed the high affinity of both polysaccharide forms (sulfated (fucoidan) and non-sulfated (alginate)) to L- and E-selectin, monocyte chemoattractant protein 1 (MCP-1), and ICAM-1, with no signs of cytotoxicity for THP-1 macrophages, even at high concentrations. The interaction with these inflammatory markers was manifested in vitro by an inhibited migration and reduced expression of MCP-1 and ICAM-1 in IFN-γ-induced THP-1 monocytes [23]. Experiments in vivo suggested that AOS protected mice against the cardiotoxic effects of the chemotherapeutic drug doxorubicin (DOX) (Table 1) [24], which is mediated through oxidative stress and endoplasmic reticulum (ER) apoptosis [25]. AOS application in mice greatly increased their survival rate, improved their DOX-induced cardiac dysfunction, and attenuated myocardial apoptosis. On the molecular level, AOS treatment decreased the expression of pro-apoptotic proteins (C/EBP homologous protein (CHOP) and apoptosis regulator Bcl-2-associated X protein (Bax)) and increased the expression of the anti-apoptotic protein B-cell lymphoma-2 (Bcl-2) [24]. A similar mechanism was also involved in AOS-mediated protection against myocardial ischaemic/reperfusion injury in mice. AOS treatment effectively decreased the generation of superoxide, 4-hydroxynonenal, and 3-nitrotyrosine, and down-regulated NADPH oxidase 2 and inducible nitric oxide synthase, thus attenuating oxidative stress. The decrease of myocardial apoptosis was reflected by the up-regulation of the anti-apoptotic Blc-2 protein, and the down-regulation of pro-apoptotic factors (CHOP, caspase-12, Bax, and glucose-regulated protein 78). Finally, AOS application greatly decreased the infarct size, and ameliorated cardiac dysfunction, after I/R injury in mice [26].

Table 1.
The anti-atherosclerotic effects of the discussed oligosaccharides.
Recent research on human umbilical vein endothelial cells (HUVECs) subjected to oxidative stress revealed that AOS regulated the expression of P21, focal adhesion kinase (FAK), cyclin-dependent kinase 2 (CDK2), integrin-α, phosphatase and tensin homolog (PTEN), and phosphoinositide 3-kinase alpha (PI3K), suggesting the involvement of the integrin-α/FAK/PI3K pathway. Additionally, AOS treatment protected HUVECs against oxidative stress-induced apoptosis by increasing the expression of Bcl-2, and decreasing the levels of caspase 3 and Bax [27]. As has been established, the integrin-α/FAK/PI3K pathway is involved in the regulation of various cellular processes, such as the cell cycle, signal transduction, cell proliferation, adhesion, and others, thus making it an important therapeutic target [54,55,56]. In another study, AOS effectively protected HUVECs against H2O2-induced oxidative stress and apoptosis, by up-regulating antioxidant enzymes (superoxide dismutase (SOD) and catalase (CAT)) and glutathione, and reducing the activities of caspase-3 and caspase-9 [28].
Furthermore, AOS has been suggested as a therapeutic agent to alleviate cardiac ageing. In a mouse model of D-galactose-induced ageing, AOS administration preserved the ejection fraction and fractional shortening. On the molecular level, OAS inhibited the D-galactose-induced up-regulation of ageing markers p21 and p53, and the cardiac peptide hormones brain natriuretic peptide (BNP) and natriuretic peptide A (ANP). AOS treatment also increased the mtDNA copy number and autophagy rate, sustained the mitochondrial membrane potential, and up-regulated the expression of peroxisome proliferator-activated receptor gamma coactivator 1-alpha (PGC-1α) and sirtuin-3 (SIRT3), thus improving mitochondrial biogenesis, turn-over, and integrity [29].
Together, these finding indicate that alginate is a potent compound which can protect endothelial cells against oxidative stress, endoplasmic reticulum stress-mediated apoptosis, and inflammation, thus providing novel alternative strategies to prevent atherosclerosis in the future. However, the efficiency of AOS has been demonstrated in a limited number of in vitro experiments and in vivo models. Therefore, future studies should focus on elucidating the exact molecular mechanisms regulating these beneficial effects, and the pharmacokinetics of AOS.
3. Chitosan Oligosaccharide
Chitosan oligosaccharide (COS) is known as an effective lipid-lowering natural material. COS’ unique biological activities and physico-chemical properties allow it to normalise levels of very-low-density lipoprotein (VLDL), LDL, and high-density lipoprotein (HDL), which is especially valuable in the treatment of diabetes mellitus, obesity, hyperlipidaemia, hyperglycaemia, metabolic syndrome, and associated diseases and morbidities [57,58].
COS administration decreased the lesion and plaque areas in the aortic root, increased plaque stability, and reduced plasma triglyceride, cholesterol, apoB100, and apoB48 levels in ApoE−/− mice fed on high-fat diet (Table 1). Moreover, in in vitro experiments, COS treatment increased the expression of scavenger receptor BI (SR-BI) and ATP binding cassette transporter A1 (ABCA1) in macrophages, and SR-BI and low-density lipoprotein receptor (LDL-R) in the liver (Figure 1). Interestingly, the plasma lipid level was not affected in LDL-R-deficient mice, thus suggesting that the COS lipid-lowering effect is mostly based on the LDL-R pathway [30]. Later, these in vivo results were confirmed in another study, which also provided further elucidation of the underlying molecular mechanism in the HepG2 cells treated with COS. Interestingly, while the LDL-R and HMG-CoA reductase (HMGCR) protein levels were not affected, the mRNA levels of proprotein convertase subtilisin/kexin type 9 (PCSK9) were down-regulated by COS treatment. Further differences were observed between the cell lysates, with an increased expression of Sterol-responsive element-binding protein 2 (SREBP-2) and hepatocyte nuclear factor-1α (HNF-1α) in the total cell lysates, while the levels of active nuclear subunit (nSREBP-2) were decreased, and the levels of forkhead box O3 (FOXO3a) were increased, in nuclear lysates after COS treatment. The combined in vivo and in vitro results suggested that COS acted on two transcription factors regulating PCSK9 synthesis at the gene transcription level, SREBPs and HNF-1α. Subsequently, PCSK9 (a known LDL-R inhibitor) modulated the hepatic LDL-R abundance and activity, thus improving the lipid intake by the liver, and decreasing the lipid concentration in the serum (Figure 1) [31].
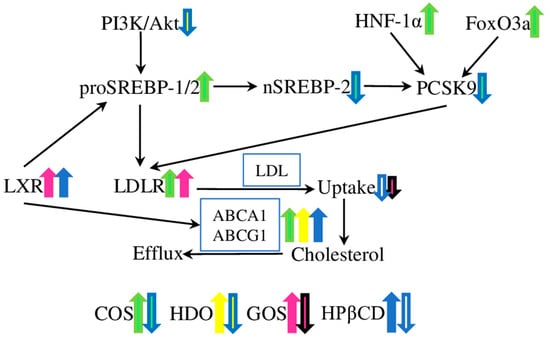
Figure 1.
The effects of oligosaccharides on cholesterol and lipid metabolism. The low-density lipoprotein receptor (LDL-R), ATP-binding cassette transporters A1 and G1 (ABCA1 and ABCG1), and liver X receptor (LXR) are the key targets of the COS and GOS, COS, HDO and HPβCD, GOS and HpβCD, respectively. Chitosan oligosaccharide (COS) treatment increased the lipid intake by the liver through the regulation of three transcription factors; forkhead box O3 (FOXO3a), sterol-responsive element-binding protein 2 (SREBPs), and hepatocyte nuclear factor-1α (HNF-1α); which decreased the level of the LDL-R inhibitor proprotein convertase subtilisin/kexin type 9 (PCSK9). Heparin-derived oligosaccharides (HDO) down-regulated the protein kinase B/phosphoinositide 3-kinase (Akt/PI3K); one of the major regulators of cellular metabolism, growth, and proliferation; and up-regulated the ABCA1 transporter. Galacto-oligosaccharides (GOSs) and 2-hydroxypropyl-β-cyclodextrin (HPβCD) acted through LRX and ABCA1 and ABCG1 in an LDL-R-dependent way, decreasing the levels of triglycerides, LDL, and very-low-density lipoprotein (VLDL) cholesterol.
Recently, lipid-lowering properties have also been demonstrated for cationically modified chitosan (via the covalent attachment of glycidyltrimethylammonium chloride), which inhibited atherosclerotic plaque development and modulated the expression of lipid metabolism genes. In particular, treatment with modified COS decreased the plasma LDL cholesterol level by 32%, and reduced the area of the atherosclerotic plaque by 33% in ApoE−/− mice. Additionally, the expression level of HMGCR was reduced in HepG2 cells after incubation with modified chitosan [32]. Similarly, interesting results were demonstrated for chitosan–fucoidan nanoparticles (CFNs), which were able to bind with P-selectin, and exhibited antioxidant and anti-inflammatory activities. Furthermore, CFNs intravenously injected into atherosclerotic ApoE−/− mice accumulated in atherosclerotic plaques, subsequently decreasing the average plaque area (by 36.5%), and the necrotic core area, and enhancing the fibrous cap thickness around the plaques, thus stabilising the atherosclerotic plaques. At the same time, long-term CFN treatment did not cause any significant adverse effects on the heart, liver, lungs, spleen, or kidneys, thus confirming its safety [33]. Together, the in vitro and in vivo results suggested that COS effectively reduced lipid levels, and suppressed local oxidative stress and inflammation, by targeting P-selectin in atheromatous plaques, and modulating the lipid metabolism, thereby preventing the progression of atherosclerosis.
4. Heparin-Derived Oligosaccharides
Heparin-derived oligosaccharides (HDOs) are delivered from heparin via enzymatic or chemical degradation, and have an ultra-low molecular weight and a narrow range of molecular weight distribution. HDOs have been proven to exhibit anti-proliferative and anti-inflammatory effects, which could be beneficial in the treatment of atherosclerosis [59,60]. As has been shown in rabbits fed a high-fat diet with vascular intimal hyperplasia in a balloon-injured carotid artery, HDOs could ameliorate the intimal hyperplasia, and inhibit the histopathology and restenosis induced by the balloon injury (Table 1). The HDO treatment decreased the expression of SR-BI, MCP-1, VCAM-1, vascular endothelial growth factor (VEGF), and basic fibroblast growth factor (bFGF) in the arterial wall. Additionally, the serum levels of lipids (total cholesterol, HDL, LDL, and triglycerides) were decreased. Interestingly, the expression level of ABCA1 was increased (Figure 1) [34]. ABCA1 is the major regulator of cellular cholesterol homeostasis, and is closely associated with atherosclerosis development [61,62].
The proliferation and migration of the vascular smooth muscle cells (VSMCs) play a crucial role in atherosclerosis development. Many growth factors (such as VEGF, bFGF, and platelet-derived growth factor (PDGF) can act through protein kinase C (PKC), the protein kinase B/phosphoinositide 3-kinase (Akt/PI3K) pathway, or mitogen-activated protein kinase (MAPK), to promote VSMC proliferation [63]. HDOs have been demonstrated to inhibit the expression of VEGF receptors 1 and 2, and interrupt normal binding between VEGF and VEGF receptors, thus acting as a VEGF antagonist. Moreover, the expression of MAPK, PI3K/Akt, and PKC was inhibited, thus suggesting a high potential for the application of HDOs for atherosclerosis prevention and cure [35]. Therefore, HDOs affect a wide variety of metabolic pathways, which could be potentially used to treat atherosclerosis. However, the underlying molecular pathways require a detailed investigation in future experiments.
5. Galacto-Oligosaccharides
Galacto-oligosaccharides (GOSs) are β(1,4)- or β(1,6)-linked oligosaccharides made up of galactose, with glucose/galactose present at the terminal end. Despite their direct effect on the gut microbiota and host immunity, a number of studies suggest various other biological activities [64]. The effect of GOSs on the lipid profile is one of the best-studied aspects in rat models of different diseases. For example, GOS administration effectively decreased the levels of total cholesterol, LDL, and VLDL cholesterol, and triglycerides in hypercholesterolaemic rats fed on a high-fat diet (Table 1) [36]. Similarly, GOS supplementation ameliorated the metabolic alterations induced by a high-fat Western-type diet in Ldlr−/− mice. In particular, lower levels of circulating macrophages and neutrophils (by 30 and 60%, respectively), suggested a reduced systemic inflammation. Moreover, GOS administration was associated with a reduced atherosclerotic lesion area, improved glucose tolerance, and reduced plasma lipopolysaccharide (LPS) concentrations [37]. Therefore, these results suggest that GOSs could be beneficial in the prevention and treatment of atherosclerosis and associated cardiovascular diseases.
Recently, GOSs were effectively used to improve the levels of HDL cholesterol, and reduce the levels of total cholesterol, triglycerides, and LDL cholesterol in the obesity rat model. GOSs also increased the expression levels of fat-browning genes (such as PR domain containing 16 (PRDM16), peroxisome proliferator-activated receptor-γ (PPARγ), peroxisome proliferator-activated receptor-γ coactivator 1α (PGC1α), and uncoupling protein 1 (UCP1)), thus promoting the browning of white fat cells, and the thermogenesis of brown fat cells. Furthermore, GOSs increased the levels of low-density lipoprotein receptor, peroxisome proliferation-activated receptor-α (PPARα), cholesterol 7α-hydroxylase proteins, and LXRα proteins in the liver of obese rats (Figure 1). Therefore, GOS inhibits obesity in rats, by facilitating white-fat browning and thermogenesis, and promoting cholesterol catabolism [38]. However, other, contradicting results were demonstrated in the rat model of streptozotocin-induced diabetes. GOS supplementation reduced the levels of MDA, total cholesterol, and CRP in diabetic rats, but had no effect on the concentrations of triglycerides and IL-6. Thus, GOSs showed a beneficial antioxidant property, while the effect on the inflammatory and lipid profiles was minor [39], suggesting that further studies are required, to establish and compare the detailed effects of GOSs in models of different diseases.
6. Cyclodextrins
Cyclodextrins (CDs) are cyclic oligosaccharides produced from various starch sources (such as corn or potatoes) by cyclodextrin glycosyltransferase, which can split and rearrange the polymer chain of starch to α-1,4-D-glucopyranoside oligomers. Common cyclodextrins are α-, β-, and γ-cyclodextrins, which are composed of six, seven, and eight glucose moieties, respectively [65]. The ability of cyclodextrins (especially β-cyclodextrin) to entrap cholesterol into their hydrophobic cavity and form stable inclusion complexes has been intensively exploited to extract excess cholesterol from atherosclerotic lesions in experimental animals [66,67]. Additionally, various physical and chemical methods have been used to modify cyclodextrins with the purpose of improving some of their characteristics (such as to improve solubility in water, decrease toxicity, and increase the ability to form complexes). At the same time, modifications take place in the hydroxyl groups (directed outwards) of cyclodextrins, thus the hydrophobicity and cavity diameter are not affected [68]. Later in this section, we focus on recent achievements in the application of various cyclodextrins in relation to atherosclerosis treatment. The application of CDs in pharmaceuticals as drug carriers has been covered in other reviews, and will be omitted here. We wish to redirect readers interested in this and related topics to the papers [69,70,71,72].
6.1. Effects on Lesion and Plaque Size and Cholesterol Transport
Experiments with ApoE−/− mice fed on a high-fat diet and supplemented with α-CD and β-CD demonstrated a 65% decrease in aortic atherosclerotic lesions in the α-CD group, and a decrease in the intestinal lipid absorption and the plasma level of free fatty acids in the β-CD group, while the major plasma lipids and body weight parameters were not affected in either experimental group (Table 1) [40]. Further in vitro experiments on aortic bovine ECs and SMCs showed that methylated β-CD decreased cholesterol release and the expression levels of ABCA1 and ABCG1 transporters, which are crucial players in regulating the cellular cholesterol pools (also known as reverse cholesterol transport, or RCT) [41]. Later, these results were confirmed on rabbits fed a high-fat diet and supplemented with 2-hydroxypropyl-β-cyclodextrin (HPβCD), which reduced inflammatory cytokine and adhesion protein (TNF-a, IL-6, IL-8, MCP-1, VCAM-1, and ICAM-1) and plasma triglyceride levels, while increasing the plasma HDL cholesterol levels. Additionally, the atherosclerosis lesion area, and collagen and macrophage content in the lesions were reduced. Furthermore, the expression levels of the ABCA1 and ABCG1 transporters in the liver and aortic plaque were increased [43]. Another β-CD modification, cyclodextrin polymer (CDP), inhibited plaque growth in a mouse model of atherosclerosis more effectively, compared to HPβCD. Moreover, CDP demonstrated better pharmacokinetics and reduced ototoxicity at high doses, thus overcoming the known limitations of CD therapy for the treatment of atherosclerosis [42].
Furthermore, HPβCD treatment reduced the atherosclerosis plaque size and cholesterol crystal load, and facilitated plaque regression in ApoE−/− mice on a continued cholesterol-rich diet. On a molecular level, HPβCD acted by targeting the liver X receptor (LXR) genes, thus reprogramming the cholesterol efflux (activating ABCA1 and ABCG1), and providing an anti-inflammatory effect (modulating the expression of the inflammasome sensor NLRP3 (NLR family pyrin domain containing 3), and the pro-inflammatory cytokines IL-1β and IL-18). The in vivo application of HPβCD on patients led to an increase in cholesterol excretion into the urine, suggesting that HPβCD extracts and transports cholesterol directly, or promotes RCT from macrophages in an LXR-dependent way (Figure 1) [44]. Similarly, experiments on macrophages showed that HPβCD can extract free cholesterol from aggregated LDL, and reduce the cholesterol uptake and cholesteryl ester accumulation from aggregated LDL by macrophages [45].
However, recent research has not confirmed the lesion-regression effect of HPβCD in atherosclerotic ApoE−/− mice fed on a high-fat diet. At the same time, experiments with macrophages have verified the ability of HPβCD to induce the cholesterol efflux from macrophages, thus supporting previous findings [73]. Therefore, despite the demonstrated safety, and beneficial effects on the key mechanisms of atherogenesis both in vitro and in vitro, further confirmatory studies are required on the regression-inducing potential of some cyclodextrin modification variants. The observed discrepancies in in vivo experiments suggest that the molecular mechanism of the HPβCD action still requires further investigation in order to be completely understood, and finely controlled, at the cellular level.
6.2. LDL Oxidation
Oxidised LDL (oxLDL) is one of the mmLDL forms implicated in atherosclerosis pathogenesis, and an important risk marker for human cardiovascular diseases [74]. It has been shown that α-CD, HPβCD, MβCD, and γ-CD can cause a significant separation between the lipid and protein components of native LDL. Furthermore, all four tested CDs (with the most pronounced results shown for HPβCD and MβCD) can reduce the LDL susceptibility to copper-induced oxidation in a concentration-dependent manner (Table 1) [46]. Similarly, MβCD and HPβCD have been shown to inhibit lipoxygenase-induced LDL oxidation in a concentration-dependent way, while increasing the lipoxygenase-induced oxidation of the lipids depleted from LDL [47]. However, it is necessary to note that the CDs were used at a relatively high concentration, and the results were demonstrated only in vitro. Therefore, further research is required to confirm these promising results in vivo.
6.3. Adhesion to Endothelial Cells
The adhesion of leukocytes to ECs promotes inflammation and endothelial dysfunction, and thus plays an important role in atherosclerosis pathogenesis. Multiple research studies have confirmed that cyclodextrins (mostly βCD and its modifications) can reduce leukocyte adhesion to ECs. As has been demonstrated, MβCD can impair the adhesion of THP-1 monocytes to HUVECs by decreasing the expression of ICAM-1 and modulating membrane–cytoskeleton coupling. In particular, MβCD reduced the average length of the individual actin filaments, and reduced the expression of the caveolin-1 and Phosphatidylinositol 4,5-bisphosphate (PIP2) levels, two major components of caveolae, and thus impaired the adhesion of circulating leukocytes to HUVECs, which could potentially help to inhibit inflammation and the initiation/progression of atherosclerosis [48].
Recently, several molecular mechanisms of the βCD have been suggested. Firstly, βCD was found to regulate NO production in bovine aortic endothelial cells by inactivating the protein kinase Cε (PKCε), which is known to dephosphorylate T495 in endothelial nitric oxide synthetase (eNOS) (Figure 2). Furthermore, the elevated NO level inhibited IκB degradation, and the subsequent TNFα-induced nuclear factor kappa B (NFκB) activation, thus blocking ICAM-1 and VCAM-1 expression, and thereby inhibiting monocyte adhesion to ECs (Table 1) [49]. Secondly, another mechanism has been identified on oxLDL and LPS treated HUVECs, whereby MβCD reduced by 2–3 times the expression of ICAM-1 and VCAM-1, acting through the IKK-NFκB (for LPS) or Akt-NFκB (for oxLDL) pathway [50]. Finally, luminol-conjugated β-cyclodextrin nanoparticles (LCD NP) inhibited neutrophil and macrophage infiltration, and subsequent pro-inflammatory events, in both in vitro and in vivo experiments. Accordingly, neutrophils treated with LCD NP reduced the phorbol 12-myristate 13-acetate (PMA)-induced production of TNFα, IL-1β, and ROS. The treatment of RAW264.7 macrophages with LCD NP inhibited their MCP-1-stimulated migration. Furthermore, long-term LCD NP administration in ApoE−/− mice fed on a high-fat diet showed an accumulation of nanoparticles in atherosclerotic plaques, which was associated with reduced plaque formation, a reduced neutrophil and macrophage number, and an increased collagen content around plaques [51]. Together, these results proposed potential mechanisms for the anti-atherosclerotic effects of CDs from the angle of monocyte–endothelial adhesion and plaque stabilisation.
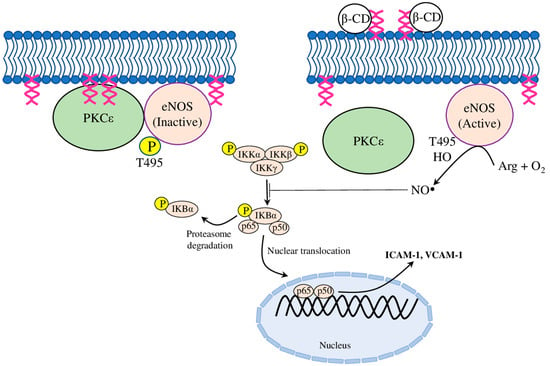
Figure 2.
The proposed mechanism of β-cyclodextrin action. β-CD binds and forms a β-CD-DAG complex with diacylglycerol (DAG), which prevents the recruitment of PKCε (protein kinase Cε) to the plasma membrane by DAG, and thus inactivates PKCε. Furthermore, the blocked PKCε-catalysed phosphorylation of eNOS (endothelial nitric oxide synthetase) at the active site T-495 activated eNOS, which produced NO (nitric oxide), and inhibited the activation of the nuclear factor kappa B (NFκB) pathway. Therefore, β-CD-mediated NO production blocked the expression of intercellular adhesion molecule-1 (ICAM-1) and vascular cell adhesion molecule-1 (VCAM-1), thereby inhibiting monocytic adhesion to endothelial cells.
6.4. Complement Activation
The complement system is a crucial part of innate immunity, which is known to contribute to the pathology of various inflammatory diseases. There are three known pathways to activate the complement system: the classical pathway, the lectin pathway, and the alternative pathway [75]. The classical pathway is activated by the direct binding of the pattern-recognition molecule C1q to various structures and ligands on pathogens or apoptotic cells, or indirectly via binding to other molecules, such as immunoglobulins or C-reactive protein. The lectin pathway is initiated through soluble mannose-binding lectin (MBL) and ficolins, which can recognise a wide variety of microorganisms, altered self-structures, and acetylated compounds, respectively [76]. The alternative pathway is activated via the spontaneous hydrolysis of the internal C3 thioester in the fluid phase, or directly on foreign surfaces [77]. Additionally, the alternative pathway functions to substantially amplify the activation induced by the classical and lectin pathways. All three pathways rely on the central C3 molecule, leading to the cleavage of C3 into its active fragments C3a and C3b, and the subsequent cleavage of C5 to C5a and C5b, and the further assembly of the terminal complement complex. Moreover, C3a and C5a are potent anaphylatoxins that can induce both sterile and nonsterile inflammatory processes [78].
It is known that cholesterol crystals have employed the complement system to activate cytokine production. Acting through both the classical and alternative complement pathways, cholesterol crystals induced the release of cytokines (IL1-β and TNFα) and ROS production, and up-regulated the production of active caspase-1 and complement receptor 3 (or CD18), with the latter leading to the phagocytosis of cholesterol crystals [79]. Further investigation showed that cholesterol crystals and complement initiated plaque inflammation, by activating the NLRP3 inflammasome. Accordingly, the complement C1q and C5b-9 complex accumulated around cholesterol crystal clefts, and the complement receptors C5aR1, C5aR2, and C3aR1 were higher in the carotid plaques of acute coronary syndrome patients. Furthermore, the priming of human carotid plaques with C5a, and incubation with cholesterol crystals increase the release of IL-1α, IL-1β, and IL-18; and priming with C5a and TNFα, and incubation with cholesterol crystals up-regulated the plaque expression of NLRP3 inflammasome components [80]. Similarly, cholesterol crystals activated the lectin pathway by binding with MBL and ficolin-2 (in a calcium-dependent and calcium-independent way, respectively). Furthermore, IgM was bound to cholesterol crystal, and facilitated its binding with C1q in vitro [81].
Recent research demonstrated that CDs were effective inhibitors of the cholesterol crystal-mediated activation of the complement system. It was shown that HPβCD inhibited C3c binding to cholesterol crystals, and reduced the terminal complement complex (TCC) generation and deposition on the surface of cholesterol crystals in human plasma, by reducing C1q and ficolin-2, and Igs (IgM and IgA) deposition on the surface of cholesterol crystals. Interestingly, HPβCD application reduced the expression of CR1 and CR3 receptors in monocytes, but not in granulocytes, where the phagocytosis of cholesterol crystals was increased (Table 1). However, the cholesterol crystal-stimulated ROS production was decreased after HPβCD treatment in both monocytes and granulocytes. Furthermore, the expression of key cytokines (such as TNFα, IL-1α, IL-1β, IL-6, and IL-8), NLRP3-related genes, and macrophage inflammatory protein-1α (MIP-1α) was decreased after exposure to HPβCD [52]. Later investigation has defined similar properties for αCD, which also inhibited cholesterol crystal-induced complement activation (by preventing the binding of C1q and ficolin-2), reduced ROS production in both monocytes and granulocytes, and reduced phagocytosis. Interestingly, unlike HPβCD, αCD could dissolve cholesterol crystals [53]. Together, these results suggested CDs as a potential treatment to prevent cholesterol crystal-induced complement activation and inflammation in atherosclerosis.
7. Oligosaccharides Which Stimulate Atherosclerosis
The association between the galactose-α-1,3-galactose (α-Gal) oligosaccharide and atherosclerosis was recently demonstrated. The novel α-Gal syndrome was described as an IgE-mediated immediate-type allergy to α-Gal [82]. The total IgE and α-Gal specific IgE were tested in the sera of 118 patients who had been subjected to medically warranted cardiac catheterisation and who had undergone an intravascular ultrasound. It was shown that α-Gal-specific IgE to α-Gal was detected in 26% of cases, and the atheroma burden and unstable features of the plaques were higher in sensitised subjects. The most pronounced association between the IgE to α-Gal and the atherosclerosis burden was defined in elder subjects (≤65 years of age) [83]. A further cross-sectional study among patients with, or at risk of, coronary artery disease (CAD) identified an association between α-Gal sensitisation and CAD burden and ST-segment-elevation myocardial infarction (STEMI). Independently of other risk factors, α-Gal sensitisation was associated with the presence of non-calcified plaque and obstructive CAD. Furthermore, the α-Gal sensitisation rate was higher in patients with STEMI than in the patients with CAD [84], thus proposing IgE sensitisation to α-Gal as a novel risk factor for coronary atherosclerosis.
Interestingly, a common disaccharide sucrose was recently identified as the main driver of the metabolic inflammation promoting atherosclerosis in LDLr−/− ApoB100/100 hyperlipidaemic mice. Mice fed on a high-fat/low-sucrose diet were characterised by pronounced dyslipidaemia, insulin resistance, and obesity, compared to mice fed on a low-fat/high-sucrose diet. On the other hand, a high-sucrose diet was associated with a reduced macrophage cholesterol efflux capacity, higher adipose tissue inflammation, liver inflammation and fibrosis, ~80% enhanced aortic atherosclerosis area, and left ventricular enlargement [85]. However, further studies, especially in human patients, are required, to define the exact impact of these results, because they could provide new insights into the pathogenesis of atherosclerosis, other cardiovascular diseases, and liver diseases.
8. Conclusions
Oligosaccharides derived from natural sources have many practical aspects that make them appealing therapeutics for the chronic processes associated with atherosclerosis. Despite a centuries-long history of consumption by domestic animals and humans supporting the safety of oligosaccharides in a diet, the exact molecular mechanisms of their action are mostly unknown, and require further research. The discussed oligosaccharides have demonstrated encouraging results in terms of increased antioxidant defence, reduced atherosclerotic plaque size, inhibited cholesterol uptake and foam cell formation, and arrested cholesterol crystal-induced complement activation and inflammation progress (Figure 3). Furthermore, modern methods of enzymatic degradation, polymerisation, chemical modification, and nanomodification can help to improve the pharmacokinetics and bio-distribution of oligosaccharides, thus improving anti-atherosclerotic activities while reducing side effects. Overall, there is great therapeutic potential in oligosaccharides, with the added benefits of a low cost, low toxicity, and abundant supply, enabling their chronic dietary administration to prevent and treat atherosclerosis, diabetes, obesity, and related diseases.
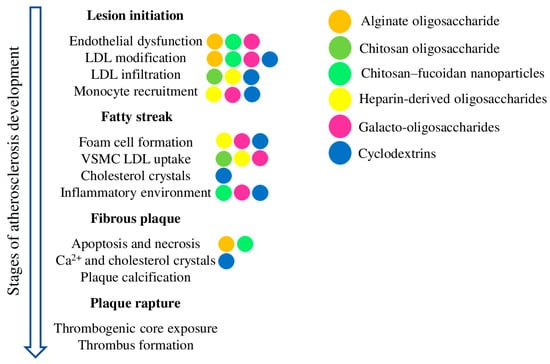
Figure 3.
Schematic representation of atherosclerosis development and the most important processes contributing to each stage. The coloured dots depict the effects of the corresponding oligosaccharides on the given process.
Author Contributions
S.A.D. and A.N.O. conceptualised the manuscript; S.A.D. wrote the manuscript text; V.N.S., A.A.M., V.A.K. and A.N.O. reviewed the text; V.N.S. and A.A.M. composed the methodology; A.A.M. and V.A.K. carried out the formal analysis; and V.N.S. and A.N.O. obtained the funding and supervised the study. All authors have read and agreed to the published version of the manuscript.
Funding
The work was supported by the Ministry of Science and Higher Education of the Russian Federation (Project # FGFU-2022-00008).
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Conflicts of Interest
The authors declare no conflict of interest. The funder played no role in the design of the study; in the collection, analysis, or interpretation of the data; in the writing of the manuscript, or in the decision to publish the results.
References
- Fan, J.; Watanabe, T. Atherosclerosis: Known and Unknown. Pathol. Int. 2022, 72, 151–160. [Google Scholar] [CrossRef]
- WHO. CVDs Fact Sheets Cardiovascular Diseases (CVDs). Available online: https://www.who.int/health-topics/cardiovascular-diseases#tab=tab_1 (accessed on 11 June 2023).
- Borén, J.; Chapman, M.J.; Krauss, R.M.; Packard, C.J.; Bentzon, J.F.; Binder, C.J.; Daemen, M.J.; Demer, L.L.; Hegele, R.A.; Nicholls, S.J.; et al. Low-Density Lipoproteins Cause Atherosclerotic Cardiovascular Disease: Pathophysiological, Genetic, and Therapeutic Insights: A Consensus Statement from the European Atherosclerosis Society Consensus Panel. Eur. Heart J. 2020, 41, 2313–2330. [Google Scholar] [CrossRef] [PubMed]
- Mezentsev, A.; Bezsonov, E.; Kashirskikh, D.; Baig, M.S.; Eid, A.H.; Orekhov, A. Proatherogenic Sialidases and Desialylated Lipoproteins: 35 Years of Research and Current State from Bench to Bedside. Biomedicines 2021, 9, 600. [Google Scholar] [CrossRef] [PubMed]
- Gisterå, A.; Hansson, G.K. The Immunology of Atherosclerosis. Nat. Rev. Nephrol. 2017, 13, 368–380. [Google Scholar] [CrossRef] [PubMed]
- Batty, M.; Bennett, M.R.; Yu, E. The Role of Oxidative Stress in Atherosclerosis. Cells 2022, 11, 3843. [Google Scholar] [CrossRef]
- Petrucci, G.; Rizzi, A.; Hatem, D.; Tosti, G.; Rocca, B.; Pitocco, D. Role of Oxidative Stress in the Pathogenesis of Atherothrombotic Diseases. Antioxidants 2022, 11, 1408. [Google Scholar] [CrossRef] [PubMed]
- Yurdagul, A.; Doran, A.C.; Cai, B.; Fredman, G.; Tabas, I.A. Mechanisms and Consequences of Defective Efferocytosis in Atherosclerosis. Front. Cardiovasc. Med. 2018, 4, 86. [Google Scholar] [CrossRef]
- Leszczynska, A.; O’Doherty, A.; Farrell, E.; Pindjakova, J.; O’Brien, F.J.; O’Brien, T.; Barry, F.; Murphy, M. Differentiation of Vascular Stem Cells Contributes to Ectopic Calcification of Atherosclerotic Plaque. Stem Cells 2016, 34, 913–923. [Google Scholar] [CrossRef]
- Hutcheson, J.D.; Goettsch, C.; Bertazzo, S.; Maldonado, N.; Ruiz, J.L.; Goh, W.; Yabusaki, K.; Faits, T.; Bouten, C.; Franck, G.; et al. Genesis and Growth of Extracellular-Vesicle-Derived Microcalcification in Atherosclerotic Plaques. Nat. Mater. 2016, 15, 335–343. [Google Scholar] [CrossRef]
- Libby, P. The Changing Landscape of Atherosclerosis. Nature 2021, 592, 524–533. [Google Scholar] [CrossRef]
- Yu, Y.; Delbianco, M. Conformational Studies of Oligosaccharides. Chem. Eur. J. 2020, 26, 9814–9825. [Google Scholar] [CrossRef] [PubMed]
- Hofman, D.L.; Van Buul, V.J.; Brouns, F.J.P.H. Nutrition, Health, and Regulatory Aspects of Digestible Maltodextrins. Crit. Rev. Food Sci. Nutr. 2016, 56, 2091–2100. [Google Scholar] [CrossRef]
- Lordan, C.; Thapa, D.; Ross, R.P.; Cotter, P.D. Potential for Enriching Next-Generation Health-Promoting Gut Bacteria through Prebiotics and Other Dietary Components. Gut Microbes 2020, 11, 1–20. [Google Scholar] [CrossRef] [PubMed]
- Tyrikos-Ergas, T.; Fittolani, G.; Seeberger, P.H.; Delbianco, M. Structural Studies Using Unnatural Oligosaccharides: Toward Sugar Foldamers. Biomacromolecules 2020, 21, 18–29. [Google Scholar] [CrossRef] [PubMed]
- Patil, N.P.; Le, V.; Sligar, A.D.; Mei, L.; Chavarria, D.; Yang, E.Y.; Baker, A.B. Algal Polysaccharides as Therapeutic Agents for Atherosclerosis. Front. Cardiovasc. Med. 2018, 5, 153. [Google Scholar] [CrossRef] [PubMed]
- Yang, Z.; Wang, H.; Liu, N.; Zhao, K.; Sheng, Y.; Pang, H.; Shao, K.; Zhang, M.; Li, S.; He, N. Algal Polysaccharides and Derivatives as Potential Therapeutics for Obesity and Related Metabolic Diseases. Food Funct. 2022, 13, 11387–11409. [Google Scholar] [CrossRef]
- Vourakis, M.; Mayer, G.; Rousseau, G. The Role of Gut Microbiota on Cholesterol Metabolism in Atherosclerosis. Int. J. Mol. Sci. 2021, 22, 8074. [Google Scholar] [CrossRef]
- Shen, X.; Li, L.; Sun, Z.; Zang, G.; Zhang, L.; Shao, C.; Wang, Z. Gut Microbiota and Atherosclerosis—Focusing on the Plaque Stability. Front. Cardiovasc. Med. 2021, 8, 668532. [Google Scholar] [CrossRef]
- Zhao, X.; Oduro, P.K.; Tong, W.; Wang, Y.; Gao, X.; Wang, Q. Therapeutic Potential of Natural Products against Atherosclerosis: Targeting on Gut Microbiota. Pharmacol. Res. 2021, 163, 105362. [Google Scholar] [CrossRef]
- Xing, M.; Cao, Q.; Wang, Y.; Xiao, H.; Zhao, J.; Zhang, Q.; Ji, A.; Song, S. Advances in Research on the Bioactivity of Alginate Oligosaccharides. Mar. Drugs 2020, 18, 144. [Google Scholar] [CrossRef]
- Liu, J.; Yang, S.; Li, X.; Yan, Q.; Reaney, M.J.T.; Jiang, Z. Alginate Oligosaccharides: Production, Biological Activities, and Potential Applications. Compr. Rev. Food Sci. Food Saf. 2019, 18, 1859–1881. [Google Scholar] [CrossRef] [PubMed]
- Huwait, E.; Al-Saedi, D.A.; Mirza, Z. Anti-Inflammatory Potential of Fucoidan for Atherosclerosis: In Silico and in vitro Studies in THP-1 Cells. Molecules 2022, 27, 3197. [Google Scholar] [CrossRef] [PubMed]
- Guo, J.-J.; Ma, L.-L.; Shi, H.-T.; Zhu, J.-B.; Wu, J.; Ding, Z.-W.; An, Y.; Zou, Y.-Z.; Ge, J.-B. Alginate Oligosaccharide Prevents Acute Doxorubicin Cardiotoxicity by Suppressing Oxidative Stress and Endoplasmic Reticulum-Mediated Apoptosis. Mar. Drugs 2016, 14, 231. [Google Scholar] [CrossRef]
- Fu, H.Y.; Sanada, S.; Matsuzaki, T.; Liao, Y.; Okuda, K.; Yamato, M.; Tsuchida, S.; Araki, R.; Asano, Y.; Asanuma, H.; et al. Chemical Endoplasmic Reticulum Chaperone Alleviates Doxorubicin-Induced Cardiac Dysfunction. Circ. Res. 2016, 118, 798–809. [Google Scholar] [CrossRef]
- Guo, J.-J.; Xu, F.-Q.; Li, Y.-H.; Li, J.; Liu, X.; Wang, X.-F.; Hu, L.-G.; An, Y. Alginate Oligosaccharide Alleviates Myocardial Reperfusion Injury by Inhibiting Nitrative and Oxidative Stress and Endoplasmic Reticulum Stress-Mediated Apoptosis. Drug Des. Dev. Ther. 2017, 11, 2387–2397. [Google Scholar] [CrossRef]
- Zhao, J.; Han, Y.; Wang, Z.; Zhang, R.; Wang, G.; Mao, Y. Alginate Oligosaccharide Protects Endothelial Cells against Oxidative Stress Injury via Integrin-α/FAK/PI3K Signaling. Biotechnol. Lett. 2020, 42, 2749–2758. [Google Scholar] [CrossRef]
- Jiang, Z.; Zhang, X.; Wu, L.; Li, H.; Chen, Y.; Li, L.; Ni, H.; Li, Q.; Zhu, Y. Exolytic Products of Alginate by the Immobilized Alginate Lyase Confer Antioxidant and Antiapoptotic Bioactivities in Human Umbilical Vein Endothelial Cells. Carbohydr. Polym. 2021, 251, 116976. [Google Scholar] [CrossRef]
- Feng, W.; Liu, J.; Wang, S.; Hu, Y.; Pan, H.; Hu, T.; Guan, H.; Zhang, D.; Mao, Y. Alginate Oligosaccharide Alleviates D-galactose-induced Cardiac Ageing via Regulating Myocardial Mitochondria Function and Integrity in Mice. J. Cell Mol. Med. 2021, 25, 7157–7168. [Google Scholar] [CrossRef] [PubMed]
- Yu, Y.; Luo, T.; Liu, S.; Song, G.; Han, J.; Wang, Y.; Yao, S.; Feng, L.; Qin, S. Chitosan Oligosaccharides Attenuate Atherosclerosis and Decrease Non-HDL in ApoE−/− Mice. J. Atheroscler. Thromb. 2015, 22, 926–941. [Google Scholar] [CrossRef] [PubMed]
- Yang, X.; Zhang, J.; Chen, L.; Wu, Q.; Yu, C. Chitosan Oligosaccharides Enhance Lipid Droplets via Down-Regulation of PCSK9 Gene Expression in HepG2 Cells. Exp. Cell Res. 2018, 366, 152–160. [Google Scholar] [CrossRef]
- Stefan, J.; Kus, K.; Wisniewska, A.; Lorkowska-Zawicka, B.; Kaminski, K.; Szczubialka, K.; Nowakowska, M.; Korbut, R. The Antiatherogenic Effect of New Biocompatible Cationically Modified Polysaccharides: Chitosan and Pullulan-the Comparison Study. J. Physiol. Pharmacol. 2018, 69, 26402. [Google Scholar] [CrossRef]
- Liu, M.; Zhang, Y.; Ma, X.; Zhang, B.; Huang, Y.; Zhao, J.; Wang, S.; Li, Y.; Zhu, Y.; Xiong, J.; et al. Synthesis and Characterization of Fucoidan-Chitosan Nanoparticles Targeting P-Selectin for Effective Atherosclerosis Therapy. Oxidative Med. Cell. Longev. 2022, 2022, 8006642. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.-R.; Wu, J.; Yu, X.-C.; Qian, X.; Xiong, R.; Wang, H.-F.; Yu, D.-F.; Liu, F.-F.; He, S.-Y. Heparin-Derived Oligosaccharide Inhibits Vascular Intimal Hyperplasia in Balloon-Injured Carotid Artery. Chin. J. Nat. Med. 2017, 15, 442–450. [Google Scholar] [CrossRef]
- Liu, J.; Wang, H.; Yu, D.; Chen, X.; He, S. Modulation of Binding to Vascular Endothelial Growth Factor and Receptor by Heparin Derived Oligosaccharide. Carbohydr. Polym. 2017, 174, 558–564. [Google Scholar] [CrossRef]
- Hashmi, A.; Naeem, N.; Farooq, Z.; Masood, S.; Iqbal, S.; Naseer, R. Effect of Prebiotic Galacto-Oligosaccharides on Serum Lipid Profile of Hypercholesterolemics. Probiotics Antimicro. Prot. 2016, 8, 19–30. [Google Scholar] [CrossRef]
- Ghosh, S.S.; Wang, J.; Yannie, P.J.; Sandhu, Y.K.; Korzun, W.J.; Ghosh, S. Dietary Supplementation with Galactooligosaccharides Attenuates High-Fat, High-Cholesterol Diet-Induced Glucose Intolerance and Disruption of Colonic Mucin Layer in C57BL/6 Mice and Reduces Atherosclerosis in Ldlr−/− Mice. J. Nutr. 2020, 150, 285–293. [Google Scholar] [CrossRef] [PubMed]
- Kong, S.; Huang, X.; Cao, H.; Bai, Y.; Che, Q.; Nie, H.; Su, Z. Anti-Obesity Effects of Galacto-Oligosaccharides in Obese Rats. Eur. J. Pharmacol. 2022, 917, 174728. [Google Scholar] [CrossRef]
- Boyanov, K.O.; Choneva, M.A.; Dimov, I.; Dimitrov, I.V.; Vlaykova, T.I.; Gerginska, F.D.; Delchev, S.D.; Hrischev, P.I.; Georgieva, K.N.; Bivolarska, A.V. Effect of Oligosaccharides on the Antioxidant, Lipid and Inflammatory Profiles of Rats with Streptozotocin-Induced Diabetes Mellitus. Z. Naturforschung C 2022, 77, 379–386. [Google Scholar] [CrossRef]
- Sakurai, T.; Sakurai, A.; Chen, Y.; Vaisman, B.L.; Amar, M.J.; Pryor, M.; Thacker, S.G.; Zhang, X.; Wang, X.; Zhang, Y.; et al. Dietary α-Cyclodextrin Reduces Atherosclerosis and Modifies Gut Flora in Apolipoprotein E-Deficient Mice. Mol. Nutr. Food Res. 2017, 61, 1600804. [Google Scholar] [CrossRef]
- Coisne, C.; Hallier-Vanuxeem, D.; Boucau, M.-C.; Hachani, J.; Tilloy, S.; Bricout, H.; Monflier, E.; Wils, D.; Serpelloni, M.; Parissaux, X.; et al. β-Cyclodextrins Decrease Cholesterol Release and ABC-Associated Transporter Expression in Smooth Muscle Cells and Aortic Endothelial Cells. Front. Physiol. 2016, 7, 185. [Google Scholar] [CrossRef]
- Kim, H.; Han, J.; Park, J.-H. Cyclodextrin Polymer Improves Atherosclerosis Therapy and Reduces Ototoxicity. J. Control. Release 2020, 319, 77–86. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Zhang, X.; Yu, B.; Peng, X.; Liu, Y.; Wang, A.; Zhao, D.; Pang, D.; OuYang, H.; Tang, X. Cyclodextrin Ameliorates the Progression of Atherosclerosis via Increasing High-Density Lipoprotein Cholesterol Plasma Levels and Anti-Inflammatory Effects in Rabbits. J. Cardiovasc. Pharmacol. 2019, 73, 334–342. [Google Scholar] [CrossRef] [PubMed]
- Zimmer, S.; Grebe, A.; Bakke, S.S.; Bode, N.; Halvorsen, B.; Ulas, T.; Skjelland, M.; De Nardo, D.; Labzin, L.I.; Kerksiek, A.; et al. Cyclodextrin Promotes Atherosclerosis Regression via Macrophage Reprogramming. Sci. Transl. Med. 2016, 8, 333ra50. [Google Scholar] [CrossRef]
- Singh, R.K.; Lund, F.W.; Haka, A.S.; Maxfield, F.R. High-Density Lipoprotein or Cyclodextrin Extraction of Cholesterol from Aggregated LDL Reduces Foam Cell Formation. J. Cell Sci. 2019, 132, jcs.237271. [Google Scholar] [CrossRef]
- Ao, M.; Gan, C.; Shao, W.; Zhou, X.; Chen, Y. Effects of Cyclodextrins on the Structure of LDL and Its Susceptibility to Copper-Induced Oxidation. Eur. J. Pharm. Sci. 2016, 91, 183–189. [Google Scholar] [CrossRef]
- Ao, M.; Chen, Y. Effects of MβCD on Lipoxygenase-Induced LDL Oxidation. Chem. Pharm. Bull. 2017, 65, 200–203. [Google Scholar] [CrossRef]
- Ao, M.; Wu, L.; Zhou, X.; Chen, Y. Methyl-β-Cyclodextrin Impairs the Monocyte-Adhering Ability of Endothelial Cells by Down-Regulating Adhesion Molecules and Caveolae and Reorganizing the Actin Cytoskeleton. Biol. Pharm. Bull. 2016, 39, 1029–1034. [Google Scholar] [CrossRef]
- Jang, S.; Lee, S.; Park, H. β-Cyclodextrin Inhibits Monocytic Adhesion to Endothelial Cells through Nitric Oxide-Mediated Depletion of Cell Adhesion Molecules. Molecules 2020, 25, 3575. [Google Scholar] [CrossRef]
- Chen, G.; Zhou, Y.; Zhang, W.; Qin, Y.; Wei, B.; Sun, Y.; Chen, Y. Methyl-β-Cyclodextrin Suppresses the Monocyte-Endothelial Adhesion Triggered by Lipopolysaccharide (LPS) or Oxidized Low-Density Lipoprotein (OxLDL). Pharm. Biol. 2021, 59, 1034–1042. [Google Scholar] [CrossRef]
- Guo, J.; Li, D.; Tao, H.; Li, G.; Liu, R.; Dou, Y.; Jin, T.; Li, L.; Huang, J.; Hu, H.; et al. Cyclodextrin-Derived Intrinsically Bioactive Nanoparticles for Treatment of Acute and Chronic Inflammatory Diseases. Adv. Mater. 2019, 31, 1904607. [Google Scholar] [CrossRef] [PubMed]
- Bakke, S.S.; Aune, M.H.; Niyonzima, N.; Pilely, K.; Ryan, L.; Skjelland, M.; Garred, P.; Aukrust, P.; Halvorsen, B.; Latz, E.; et al. Cyclodextrin Reduces Cholesterol Crystal–Induced Inflammation by Modulating Complement Activation. J. Immunol. 2017, 199, 2910–2920. [Google Scholar] [CrossRef] [PubMed]
- Pilely, K.; Bakke, S.S.; Palarasah, Y.; Skjoedt, M.-O.; Bartels, E.D.; Espevik, T.; Garred, P. Alpha-Cyclodextrin Inhibits Cholesterol Crystal-Induced Complement-Mediated Inflammation: A Potential New Compound for Treatment of Atherosclerosis. Atherosclerosis 2019, 283, 35–42. [Google Scholar] [CrossRef]
- Matsuoka, T.; Yashiro, M.; Nishioka, N.; Hirakawa, K.; Olden, K.; Roberts, J.D. PI3K/Akt Signalling Is Required for the Attachment and Spreading, and Growth in vivo of Metastatic Scirrhous Gastric Carcinoma. Br. J. Cancer 2012, 106, 1535–1542. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.-R.; Zhao, J.-T.; Xie, Z.-Z. Integrin-Mediated Cancer Progression as a Specific Target in Clinical Therapy. Biomed. Pharmacother. 2022, 155, 113745. [Google Scholar] [CrossRef] [PubMed]
- Wu, C.; You, J.; Fu, J.; Wang, X.; Zhang, Y. Phosphatidylinositol 3-Kinase/Akt Mediates Integrin Signaling to Control RNA Polymerase I Transcriptional Activity. Mol. Cell. Biol. 2016, 36, 1555–1568. [Google Scholar] [CrossRef]
- Van Der Gronde, T.; Hartog, A.; Van Hees, C.; Pellikaan, H.; Pieters, T. Systematic Review of the Mechanisms and Evidence behind the Hypocholesterolaemic Effects of HPMC, Pectin and Chitosan in Animal Trials. Food Chem. 2016, 199, 746–759. [Google Scholar] [CrossRef]
- Tao, W.; Wang, G.; Wei, J. The Role of Chitosan Oligosaccharide in Metabolic Syndrome: A Review of Possible Mechanisms. Mar. Drugs 2021, 19, 501. [Google Scholar] [CrossRef]
- Li, L.; Li, W.; Ren, Z.; He, S.; Xu, G.; Yang, L. Effect of Heparin-Derived Oligosaccharide on Vascular Smooth Muscle Cell Proliferation. Vasc. Endovasc. Surg. 2012, 46, 393–400. [Google Scholar] [CrossRef]
- Yu, D.; Rui, X.; He, S. Effect of Heparin-Derived Oligosaccharide on BFGFR1 and BFGFR2 in Vascular Smooth Muscle Cells. Vasc. Endovasc. Surg. 2014, 48, 289–296. [Google Scholar] [CrossRef]
- Abdel-Razek, O.; Sadananda, S.N.; Li, X.; Cermakova, L.; Frohlich, J.; Brunham, L.R. Increased Prevalence of Clinical and Subclinical Atherosclerosis in Patients with Damaging Mutations in ABCA1 or APOA1. J. Clin. Lipidol. 2018, 12, 116–121. [Google Scholar] [CrossRef]
- Ogura, M. HDL, Cholesterol Efflux, and ABCA1: Free from Good and Evil Dualism. J. Pharmacol. Sci. 2022, 150, 81–89. [Google Scholar] [CrossRef]
- Hellesøy, M.; Lorens, J.B. Cellular Context–Mediated Akt Dynamics Regulates MAP Kinase Signaling Thresholds during Angiogenesis. Mol. Biol. Cell 2015, 26, 2698–2711. [Google Scholar] [CrossRef] [PubMed]
- Mei, Z.; Yuan, J.; Li, D. Biological Activity of Galacto-Oligosaccharides: A Review. Front. Microbiol. 2022, 13, 993052. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Li, M.; Mao, H. Cross-linked Enzyme Aggregates of Recombinant Cyclodextrin Glycosyltransferase for High-purity Β-cyclodextrin Production. J. Chem. Technol. Biotechnol. 2019, 94, 1528–1533. [Google Scholar] [CrossRef]
- Fang, F.; Huang, X.-J.; Guo, Y.-Z.; Hong, X.; Wu, H.-M.; Liu, R.; Chen, D.-J. Selective and Regenerable Surface Based on β-Cyclodextrin for Low-Density Lipoprotein Adsorption. Langmuir 2018, 34, 8163–8169. [Google Scholar] [CrossRef] [PubMed]
- Fenyvesi, É.; Puskás, I.; Szente, L. Applications of Steroid Drugs Entrapped in Cyclodextrins. Environ. Chem. Lett. 2019, 17, 375–391. [Google Scholar] [CrossRef]
- Saokham, P.; Muankaew, C.; Jansook, P.; Loftsson, T. Solubility of Cyclodextrins and Drug/Cyclodextrin Complexes. Molecules 2018, 23, 1161. [Google Scholar] [CrossRef]
- Komiyama, M. Cyclodextrins as Eminent Constituents in Nanoarchitectonics for Drug Delivery Systems. Beilstein J. Nanotechnol. 2023, 14, 218–232. [Google Scholar] [CrossRef]
- Liu, J.; Ding, X.; Fu, Y.; Xiang, C.; Yuan, Y.; Zhang, Y.; Yu, P. Cyclodextrins Based Delivery Systems for Macro Biomolecules. Eur. J. Med. Chem. 2021, 212, 113105. [Google Scholar] [CrossRef]
- Topuz, F.; Uyar, T. Advances in the Development of Cyclodextrin-Based Nanogels/Microgels for Biomedical Applications: Drug Delivery and Beyond. Carbohydr. Polym. 2022, 297, 120033. [Google Scholar] [CrossRef]
- Zhang, L. Cyclodextrin Related Drug Delivery System to Promote Atherosclerosis Regression. Pharmazie 2020, 75, 619–625. [Google Scholar]
- Snip, O.S.C.; Hoekstra, M.; Zhang, Y.; Geerling, J.J.; Van Eck, M. 2-Hydroxypropyl-Beta-Cyclodextrin Treatment Does Not Induce Atherosclerotic Lesion Regression in Western-Type Diet-Fed Apolipoprotein E Knockout Mice. Biomolecules 2022, 12, 1205. [Google Scholar] [CrossRef]
- Jiang, H.; Zhou, Y.; Nabavi, S.M.; Sahebkar, A.; Little, P.J.; Xu, S.; Weng, J.; Ge, J. Mechanisms of Oxidized LDL-Mediated Endothelial Dysfunction and Its Consequences for the Development of Atherosclerosis. Front. Cardiovasc. Med. 2022, 9, 925923. [Google Scholar] [CrossRef]
- Pouw, R.B.; Ricklin, D. Tipping the Balance: Intricate Roles of the Complement System in Disease and Therapy. Semin. Immunopathol. 2021, 43, 757–771. [Google Scholar] [CrossRef] [PubMed]
- Wang, P.; Wu, Q.; Shuai, Z.-W. Emerging Role of Ficolins in Autoimmune Diseases. Pharmacol. Res. 2021, 163, 105266. [Google Scholar] [CrossRef]
- Dobó, J.; Kocsis, A.; Dani, R.; Gál, P. Proprotein Convertases and the Complement System. Front. Immunol. 2022, 13, 958121. [Google Scholar] [CrossRef] [PubMed]
- Barratt-Due, A.; Pischke, S.E.; Nilsson, P.H.; Espevik, T.; Mollnes, T.E. Dual Inhibition of Complement and Toll-like Receptors as a Novel Approach to Treat Inflammatory Diseases—C3 or C5 Emerge Together with CD14 as Promising Targets. J. Leukoc. Biol. 2017, 101, 193–204. [Google Scholar] [CrossRef]
- Samstad, E.O.; Niyonzima, N.; Nymo, S.; Aune, M.H.; Ryan, L.; Bakke, S.S.; Lappegård, K.T.; Brekke, O.-L.; Lambris, J.D.; Damås, J.K.; et al. Cholesterol Crystals Induce Complement-Dependent Inflammasome Activation and Cytokine Release. J. Immunol. 2014, 192, 2837–2845. [Google Scholar] [CrossRef]
- Niyonzima, N.; Bakke, S.S.; Gregersen, I.; Holm, S.; Sandanger, Ø.; Orrem, H.L.; Sporsheim, B.; Ryan, L.; Kong, X.Y.; Dahl, T.B.; et al. Cholesterol Crystals Use Complement to Increase NLRP3 Signaling Pathways in Coronary and Carotid Atherosclerosis. EBioMedicine 2020, 60, 102985. [Google Scholar] [CrossRef] [PubMed]
- Pilely, K.; Rosbjerg, A.; Genster, N.; Gal, P.; Pál, G.; Halvorsen, B.; Holm, S.; Aukrust, P.; Bakke, S.S.; Sporsheim, B.; et al. Cholesterol Crystals Activate the Lectin Complement Pathway via Ficolin-2 and Mannose-Binding Lectin: Implications for the Progression of Atherosclerosis. J. Immunol. 2016, 196, 5064–5074. [Google Scholar] [CrossRef]
- Fischer, J.; Yazdi, A.S.; Biedermann, T. Clinical Spectrum of α-Gal Syndrome: From Immediate-Type to Delayed Immediate-Type Reactions to Mammalian Innards and Meat. Allergo J. Int. 2016, 25, 55–62. [Google Scholar] [CrossRef] [PubMed]
- Wilson, J.M.; Nguyen, A.T.; Schuyler, A.J.; Commins, S.P.; Taylor, A.M.; Platts-Mills, T.A.E.; McNamara, C.A. IgE to the Mammalian Oligosaccharide Galactose-α-1,3-Galactose Is Associated with Increased Atheroma Volume and Plaques with Unstable Characteristics—Brief Report. Arterioscler. Thromb. Vasc. Biol. 2018, 38, 1665–1669. [Google Scholar] [CrossRef] [PubMed]
- Vernon, S.T.; Kott, K.A.; Hansen, T.; Finemore, M.; Baumgart, K.W.; Bhindi, R.; Yang, J.; Hansen, P.S.; Nicholls, S.J.; Celermajer, D.S.; et al. Immunoglobulin E Sensitization to Mammalian Oligosaccharide Galactose-α-1,3 (α-Gal) Is Associated with Noncalcified Plaque, Obstructive Coronary Artery Disease, and ST-Segment–Elevated Myocardial Infarction. Arterioscler. Thromb. Vasc. Biol. 2022, 42, 352–361. [Google Scholar] [CrossRef]
- Perazza, L.R.; Mitchell, P.L.; Jensen, B.A.H.; Daniel, N.; Boyer, M.; Varin, T.V.; Bouchareb, R.; Nachbar, R.T.; Bouchard, M.; Blais, M.; et al. Dietary Sucrose Induces Metabolic Inflammation and Atherosclerotic Cardiovascular Diseases More than Dietary Fat in LDLr ApoB100/100 Mice. Atherosclerosis 2020, 304, 9–21. [Google Scholar] [CrossRef] [PubMed]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).