

GC Analysis, Anticancer, and Antibacterial Activities of Secondary Bioactive Compounds from Endosymbiotic Bacteria of Pomegranate Aphid and Its Predator and Protector
 , , , and
, , , and
Abstract
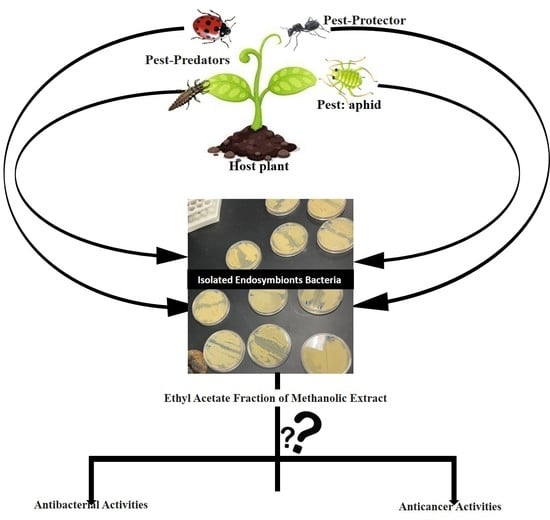
1. Introduction
2. Results
2.1. Collection of Insects and Isolation of Insects Endosymbiotic Bacteria
2.2. Antibacterial Activities of Endosymbiotic Bacteria
2.3. Antiproliferative Activities of Endosymbiotic Bacteria
2.4. Chemical Analyses for Endosymbiotic Bacterial EA-ME
3. Discussion
4. Materials and Methods
4.1. Aphid Sampling and Identification
4.2. Isolation of Insect Bacteria
4.3. DNA Extraction, Amplification and 16S rRNA Gene Sequencing
4.4. Preparation of Ethyl Acetate Fraction of Methanolic Extract (EA-ME) from Endosymbiotic Bacteria
4.5. Antibacterial Activities of EA-ME against Pathogenic Bacteria
4.6. Resazurin-Based 96-Well Plate Microdilution Assay for the Determination of MICs
4.7. Antiproliferative Activity of the Endosymbiont EA m Extracts
4.8. Gas Chromatography–Mass Spectroscopy Analysis (GC-MS)
4.9. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Sample Availability
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| No | Species | Sequence | Accession Number |
|---|---|---|---|
| Aphid and aphid predators and ants sequences of Cytochrome oxidase subunit I | |||
| 1 | Aphis illinoisensis | GGATCTTCACTTAGAATTTTAATTCGATTAGAATTAAGTCAAATTAATTCAATTATTAACAATAATCAATTATATAATGTAATCGTAACAATTCATGCTTTTATTATAATTTTTTTTATAACTATACCTATTGTAATTGGAGGATTTGGAAATTGGTTAATTCCTATAATAATAGGATGTCCAGATATATCTTTCCCACGACTAAATAATATTAGATTTTGGTTACTACCACCATCATTAATAATAATAATTTGTAGATTTATAATTAATAATGGAACAGGAACAGGATGGGCTATTTATCCACCTTTATCAAATAATATTGCTCACAATAATATTTCAGTTGATCTAACTATTTTTTCTCTTCATATAGCAGGAATTTCATCAATTTTAGGAGCAATTAATTTTATTTGTCAATTTTAAACATAATACCACATAATATAAAACTAAATCAAATTCCTTTATTCCCATGGTCAATCTTAATTACAGCCATATTATTAATTTTATCTTTACCAGTTTTAGCTGGTGCTATTACAATATTATTAACTGATCGAAATTTAAATACATCATTTTTTGATCCAGCAGGAGGAGGAGACCCTATTCTTTATCAACATTTATTCTGGTTTTTT | MZ091377 |
| 2 | Aphis punicae | GGTTCTTCTCTTAGAATTTTAATCCGATTAGAATTAAGTCAAATTAATTCAATTATTAATAATAATCAACTATATAATGTAATTGTTACAATTCATGCTTTTATTATAATTTTTTTTATAACTATACCAATCGTTATTGGAGGTTTTGGAAATTGGTTAATTCCTATAATAATAGGATGCCCAGATATATCTTTCCCACGACTAAATAATATTAGATTCTGGTTATTACCACCCTCATTAATAATAATAATTTGCAGATTTATAATTAATAATGGAACAGGAACAGGATGGACTATTTATCCACCTTTATCAAATAACATTGCTCATAATAATATTTCAGTAGACTTAACTATTTTTTCTTTACATTTAGCAGGTATTTCATCAATTTTAGGAGCAATTAATTTCATCTGCACTATCTTAAATATAATACCCAATAATATAAAATTAAATCAAATTCCTTTATTTCCATGGTCAATTTTAATTACAGCTATATTATTAATTTTATCCTTACCCGTATTAGCTGGTGCTATTACTATATTATTAACAGATCGAAATTTAAATACATCATTTTTTGATCCAGCAGGTGGTGGAGACCCTATTCTTTATCAACATTTATTTTGGTTTTTT | MZ091379 |
| 3 | Macrosiphum rosae | ACTCTTAGAATTTTAATTCGATTAGAATTAAGACAAATTAATTCTATTATTAATAATAATCAATTATATAATGTAATTGTTACAATTCATGCTTTTATTATAATTTTTTTTATAACTATACCAATTGTAATTGGAGGATTTGGAAATTGGTTAATTCCTATAATAATAGGATGCCCTGATATATCATTTCCACGTTTAAATAATATTAGATTTTGGTTATTACCTCCATCATTAATAATAATAATTTGTAGATTTTTAATTAATAACGGTACAGGAACAGGATGGACAATTTATCCACCTTTATCAAACAATATTGCACACAATAATATTTCAGTTGATTTAACTATTTTTTCTCTGCATTTAGCAGGAATTTCATCAATCTTAGGAGCAATTAACTTTATTTGTACAATTCTTAATATAATACCAAATAATTTAAAACTTAATCAAATTCCTCTCTTTCCTTGGTCAATTTTAATTACAGCTATTTTACTAATTTTATCTTTACCAGTTTTAGCCGGTGCTATTACAATATTACTAACTGATCGTAATTTAAATACATCATTTTTTGATCCAGCAGGAGGAGGAGACCCTATTTTATATCAACATTTATTTTGGTTTTTG | OL823183 |
| 4 | Chrysoperla carnea | GGATCATCTCTAAGTTTATTGATTCGAGCTGAATTAGGTCAACCAGGTTCTTTAATTGGTGATGATCAAATTTATAATGTAATTGTTACAGCACATGCTTTTATTATAATTTTTTTTATAGTAATACCTATTGTAATTGGAGGTTTTGGTAATTGGTTAGTTCCTTTAATATTAGCTGCTCCTGATATAGCTTTTCCACGAATAAATAATATAAGTTTCTGGATATTACCTCCTTCATTAACTTTATTACTTGCTTCTTCTATAGTAGAAAGAGGAGCTGGAACTGGTTGGACAGTTTACCCTCCTTTATCATCAAGAATTGCTCATGCTGGAGCTTCTGTTGATTTAGCTATTTTTAGTTTACATCTTGCCGGTATTTCATCAATTTTAGGAGCAGTAAATTTTATTACAACAGTAATTAATATACGATTAAGTTATATAACATTAGATCGTATACCATTATTTGTATGGTCAGTAGTAATTACAGCTTTATTATTATTACTTTCATTACCTGTATTAGCTGGTGCTATTACTATATTATTAACTGATCGTAATTTAAATACTTCATTTTTTGA | ON149796 |
| 5 | Coccinella undecimpunctata | GGATCATCTCTAAGAATCTTAATTCGGCTAGAACTTGGTACTACAAACAGATTAATTGGAAATGACCAAATTTATAATGTTATTGTAACAGCTCATGCATTTATTATAATTTTTTTCATAGTAATACCAATTATAATTGGAGGATTTGGAAATTGGTTAGTTCCCCTAATAATTGGGGCACCTGATATAGCTTTCCCACGTTTAAATAATATAAGATTCTGGTTATTACCTCCTGCATTAACTCTCTTAATCATTAGAAGATTAGTAGAAATAGGGGCAGGAACAGGTTGGACTGTTTACCCACCTTTATCTTCTAATTTAGCTCATAATGGGCCTTCTGTAGATTTAGTAATTTTTAGATTACACTTAGCAGGAATTTCTTCAATTCTTGGAGCTGTAAATTTCATCTCTACAATTATAAATATACGCCCCTTTGGAATAAATTTGGATAAAACTCCTTTATTTGTTTGGTCAGTACTTATTACTGCTATTTTATTACTTCTTTCATTACCAGTTTTAGCTGGGGCTATTACAATACTATTAACTGACCGTAATATTAATACATCTTTTTTTGA | ON149797 |
| 6 | Tapinoma magnum | GGATCATCTCTAAGAATAATTATCCGTATTGAATTAGGAACATGTGGAGCATTAATTAATAATGATCAAATTTATAATTCAATTGTTACAGGACATGCTTTTATTATAATTTTTTTTATAGTTATACCTTTTATAATTGGTGGATTTGGAAATTTTTTAGTCCCATTAATATTAGGTGCACCAGATATGGCTTATCCTCGAATAAATAATATAAGATTTTGGTTATTACCCCCATCAATTTTATTATTAACTATTAGAAATTTTATCAGATCAGGGGTAGGTACTGGTTGGACAGTATACCCACCCTTAGCATCTAATATTTATCATAACGGACCTTCAGTAGATTTAGCTATTTTTTCTTTACATATTGCAGGAATATCATCAATCTTAGGCGCAATTAATTTTATTTCTACAATTATTAATATACATCATAAAAATTTTTCTATTGATAAAATTCCTTTATTAGTATGGTCAATTTTAATTACTGCAATTTTATTACTTTTATCTCTTCCAGTTTTAGCAGGAGCAATTACTATGTTATTAACTGATCGAAATTTAAATACATCATTTTTTGA | ON149799 |
| 16s rRNA sequences of Bacterial isolates | |||
| 7 | Enterobacter sp. | ATTGACGTTACCCGCAGAAGAAGCACCGGCTAACTCCGTGCCAGCAGCCGCGGTAATACGGAGGGTGCAAGCGTTAATCGGAATTACTGGGCGTAAAGCGCACGCAGGCGGTCTGTCAAGTCGGATGTGAAATCCCCGGGCTCAACCTGGGAACTGCATTCGAAACTGGCAGGCTAGAGTCTTGTAGAGGGGGGTAGAATTCCAGGTGTAGCGGTGAAATGCGTAGAGATCTGGAGGAATACCGGTGGCGAAGGCGGCCCCCTGGACAAAGACTGACGCTCAGGTGCGAAAGCGTGGGGAGCAAACAGGATTAGATACCCCGGTAGTCA | OP320676 |
| 8 | Klebsiella aerogenes | ATTGACGTTACCCGCAGAAGAAGCACCGGCTAACTCCGTGCCAGCAGCCGCGGTAATACGGAGGGTGCAAGCGTTAATCGGAATTACTGGGCGTAAAGCGCACGCAGGCGGTCTGTCAAGTCGGATGTGAAATCCCCGGGCTCAACCTGGGAACTGCATTCGAAACTGGCAGGCTAGAGTCTTGTAGAGGGGGGTAGAATTCCAGGTGTAGCGGTGAAATGCGTAGAGATCTGGAGGAATACCGGTGGCGAAGGCGGCCCCCTGGACAAAGACTGACGCTCAGGTGCGAAAGCGTGGGGAGCAAACAGGATTAGATACCCCGGTAGTCAA | OP320677 |
| 9 | Pantoea agglomerans | ATTGACGTTACCCGCAGAAGAAGCACCGGCTAACTCCGTGCCAGCAGCCGCGGTAATACGGAGGGTGCAAGCGTTAATCGGAATTACTGGGCGTAAAGCGCACGCAGGCGGTTTGTTAAGTCTGATGTGAAATCCCCGGGCTCAACCGGGGAACTGCATTGGAAACTGGGAGGCTTGAGTCTTGTAGAGGGGGGTAGAATTCCAGGTGTAGCGGTGAAATGCGTAGAGATCTGGAGGAATACCGGTGGCGAAGGCGGCCCCCTGGACAGAGACTGACGCTCAGGTGCGAAAGCGTGGGGAGCAAACAGGATTAGATACCCCCGGTAGTCAA | OP320678 |
| 10 | Planococcus sp. | CTTGACGGTACCTCACCAGAAAGCCACGGCTAACTACGTGCCAGCAGCCGCGGTAATACGTAGGTGGCAAGCGTTGTCCGGAATTATTGGGCGTAAAGCGCGCGCAGGCGGTTCCTTAAGTCTGATGTGAAAGCCCACGGCTCAACCGTGGAGGGTCATTGGAAACTGGGGAACTTGAGTGCAGAAGAGGAAAGTGGAATTCCACGTGTAGCGGTGAAATGCGTAGAGATGTGGAGGAACACCAGTGGCGAAGGCGACTTTCTGGTCTGTAACTGACGCTGAGGCGCGAAAGCGTGGGGAGCAAACAGGATTAGATACCCCGGTAGTCA | OP320679 |
| 11 | Providencia stuartii | ATTGACGTTACCGACAGAAGAAGCACCGGCTAACTCCGTGCCAGCAGCCGCGGTAATACGGAGGGTGCAAGCGTTAATCGGAATTACTGGGCGTAAAGCGCACGCAGGCGGTTGATTAAGTTAGATGTGAAATCCCCGGGCTTAACCTGGGAATGGCATCTAAAACTGGTCAGCTAGAGTCTTGTAGAGGGGGGTAGAATTCCATGTGTAGCGGTGAAATGCGTAGAGATGTGGAGGAATACCGGTGGCGAAGGCGGCCCCCTGGACAAAGACTGACGCTCAGGTGCGAAAGCGTGGGGAGCAAACAGGATTAGATACCCCGGTAGTCAA | OP320680 |
| 12 | Serratia fonticola | ATTGACGTTACTCGCAGAAGAAGCACCGGCTAACTCCGTGCCAGCAGCCGCGGTAATACGGAGGGTGCAAGCGTTAATCGGAATTACTGGGCGTAAAGCGCACGCAGGCGGTTTGTTAAGTCAGATGTGAAATCCCCGAGCTTAACTTGGGAACTGCATTTGAAACTGGCAAGCTAGAGTCTTGTAGAGGGGGGTAGAATTCCAGGTGTAGCGGTGAAATGCGTAGAGATCTGGAGGAATACCGGTGGCGAAGGCGGCCCCCTGGACAAAGACTGACGCTCAGGTGCGAAAGCGTGGGGAGCAAACAGGATTAGATACCCCGGTAGTCA | OP320681 |
| 13 | Serratia odorifera | ATTGACGTTACTCGCAGAAGAAGCACCGGCTAACTCCGTGCCAGCAGCCGCGGTAATACGGAGGGTGCAAGCGTTAATCGGAATTACTGGGCGTAAAGCGCACGCAGGCGGTTTGTTAAGTCAGATGTGAAATCCCCGCGCTTAACGTGGGAACTGCATTTGAAACTGGCAAGCTAGAGTCTCGTAGAGGGGGGTAGAATTCCAGGTGTAGCGGTGAAATGCGTAGAGATCTGGAGGAATACCGGTGGCGAAGGCGGCCCCCTGGACGAAGACTGACGCTCAGGTGCGAAAGCGTGGGGAGCAAACAGGATTAGATACCCCGGTAGTCAAGCG | OP320682 |
| 14 | Bacillus safensis | CTTGACGGTACCTAACCAGAAAGCCACGGCTAACTACGTGCCAGCAGCCGCGGTAATACGTAGGTGGCAAGCGTTGTCCGGAATTATTGGGCGTAAAGGGCTCGCAGGCGGTTTCTTAAGTCTGATGTGAAAGCCCCCGGCTCAACCGGGGAGGGTCATTGGAAACTGGGAAACTTGAGTGCAGAATAGGAGAGTGGAATTCCACGTGTAGCGGTGAAATGCGTAGAGATGTGGAGGAACACCAGTGGCGAAGGCGACTCTCTGGTCTGTAACTGACGCTGAGGAGCGAAAGCGTGGGGAGCGAACAGGATTAGATACCCTGGTAGTCC | OQ351925 |
| 15 | Enterococcus avium | CTTGACGGTATCTAACCAGAAAGCCACGGCTAACTACGTGCCAGCAGCCGCGGTAATACGTAGGTGGCAAGCGTTGTCCGGATTTATTGGGCGTAAAGCGAGCGCAGGCGGTTTCTTAAGTCTGATGTGAAAGCCCCCGGCTCAACCGGGGAGGGTCATTGGAAACTGGGAAACTTGAGTGCAGAAGAGGAGAGTGGAATTCCATGTGTAGCGGTGAAATGCGTAGATATATGGAGGAACACCAGTGGCGAAGGCGGCTCTCTGGTCTGTAACTGACGCTGAGGCTCGAAAGCGTGGGAGCAAACAGGATTAGATACCCTGGTAGTCC | OQ351926 |
| 16 | Bacillus megaterium | CTTGACGGTACCTAACCAGAAAGCCACGGCTAACTACGTGCCAGCAGCCGCGGTAATACGTAGGTGGCAAGCGTTATCCGGAATTATTGGGCGTAAAGCGCGCGCAGGCGGTTTCTTAAGTCTGATGTGAAAGCCCACGGCTCAACCGTGGAGGGTCATTGGAAACTGGGGAACTTGAGTGCAGAAGAGAAAAGCGGAATTCCACGTGTAGCGGTGAAATGCGTAGAGATGTGGAGGAACACCAGTGGCGAAGGCGGCTTTTTGGTCTGTAACTGACGCTGAGGCGCGAAAGCGTGGGGAGCAAACAGGATTAGATACCCTGGTAGTCC | OQ351927 |
References
- Dube, P.N.; Sakle, N.S.; Dhawale, S.A.; More, S.A.; Mokale, S.N. Synthesis, Biological Investigation and Docking Study of Novel Chromen Derivatives as Anti-Cancer Agents. Anticancer Agents Med. Chem. 2019, 19, 1150–1160. [Google Scholar] [CrossRef]
- Siegel, R.L.; Miller, K.D.; Jemal, A. Cancer statistics, 2020. CA Cancer J. Clin. 2020, 70, 7–30. [Google Scholar] [CrossRef] [PubMed]
- Nurgali, K.; Jagoe, R.T.; Abalo, R. Editorial: Adverse Effects of Cancer Chemotherapy: Anything New to Improve Tolerance and Reduce Sequelae? Front. Pharmacol. 2018, 9, 245. [Google Scholar] [CrossRef] [PubMed]
- Pucci, C.; Martinelli, C.; Ciofani, G. Innovative approaches for cancer treatment: Current perspectives and new challenges. Ecancermedicalscience 2019, 13, 961. [Google Scholar] [CrossRef] [PubMed]
- Sebola, T.E.; Uche-Okereafor, N.C.; Mekuto, L.; Makatini, M.M.; Green, E.; Mavumengwana, V. Antibacterial and Anticancer Activity and Untargeted Secondary Metabolite Profiling of Crude Bacterial Endophyte Extracts from Crinum macowanii Baker Leaves. Int. J. Microbiol. 2020, 2020, 8839490. [Google Scholar] [CrossRef] [PubMed]
- Sebola, T.E.; Uche-Okereafor, N.C.; Tapfuma, K.I.; Mekuto, L.; Green, E.; Mavumengwana, V. Evaluating antibacterial and anticancer activity of crude extracts of bacterial endophytes from Crinum macowanii Baker bulbs. Microbiologyopen 2019, 8, e914. [Google Scholar] [CrossRef] [PubMed]
- Brader, G.; Compant, S.; Mitter, B.; Trognitz, F.; Sessitsch, A. Metabolic potential of endophytic bacteria. Curr. Opin. Biotechnol. 2014, 27, 30–37. [Google Scholar] [CrossRef]
- Abdelghani, Z.; Hourani, N.; Zaidan, Z.; Dbaibo, G.; Mrad, M.; Hage-Sleiman, R. Therapeutic applications and biological activities of bacterial bioactive extracts. Arch. Microbiol. 2021, 203, 4755–4776. [Google Scholar] [CrossRef]
- Bérdy, J. Bioactive microbial metabolites. J. Antibiot. 2005, 58, 1–26. [Google Scholar] [CrossRef]
- Khalifa, S.A.M.; Elias, N.; Farag, M.A.; Chen, L.; Saeed, A.; Hegazy, M.F.; Moustafa, M.S.; Abd El-Wahed, A.; Al-Mousawi, S.M.; Musharraf, S.G.; et al. Marine Natural Products: A Source of Novel Anticancer Drugs. Mar. Drugs 2019, 17, 491. [Google Scholar] [CrossRef]
- Clements, T.; Ndlovu, T.; Khan, W. Broad-spectrum antimicrobial activity of secondary metabolites produced by Serratia marcescens strains. Microbiol. Res. 2019, 229, 126329. [Google Scholar] [CrossRef] [PubMed]
- Karbalaei-Heidari, H.R.; Partovifar, M.; Memarpoor-Yazdi, M. Evaluation of the Bioactive Potential of Secondary Metabolites Produced by a New Marine Micrococcus Species Isolated from the Persian Gulf. Avicenna J. Med. Biotechnol. 2020, 12, 61–65. [Google Scholar] [PubMed]
- Sader, H.S.; Streit, J.M.; Fritsche, T.R.; Jones, R.N. Antimicrobial activity of daptomycin against multidrug-resistant Gram-positive strains collected worldwide. Diagn. Microbiol. Infect. Dis. 2004, 50, 201–204. [Google Scholar] [CrossRef]
- Singh, V.K.; Mishra, A.; Jha, B. 3-Benzyl-Hexahydro-Pyrrolo[1,2-a]Pyrazine-1,4-Dione Extracted From Exiguobacterium indicum Showed Anti-biofilm Activity Against Pseudomonas aeruginosa by Attenuating Quorum Sensing. Front. Microbiol. 2019, 10, 1269. [Google Scholar] [CrossRef] [PubMed]
- Krishnan, K.; Mani, A.; Jasmine, S. Cytotoxic Activity of Bioactive Compound 1, 2-Benzene Dicarboxylic Acid, Mono 2-Ethylhexyl Ester Extracted from a Marine Derived Streptomyces sp. VITSJK8. Int. J. Mol. Cell. Med. 2014, 3, 246–254. [Google Scholar] [PubMed]
- Demain, A.L.; Vaishnav, P. Natural products for cancer chemotherapy. Microb. Biotechnol. 2011, 4, 687–699. [Google Scholar] [CrossRef]
- Moran, N.A.; Degnan, P.H. Functional genomics of Buchnera and the ecology of aphid hosts. Mol. Ecol. 2006, 15, 1251–1261. [Google Scholar] [CrossRef]
- Oliver, K.M.; Degnan, P.H.; Burke, G.R.; Moran, N.A. Facultative symbionts in aphids and the horizontal transfer of ecologically important traits. Annu. Rev. Entomol. 2010, 55, 247–266. [Google Scholar] [CrossRef]
- Shigenobu, S.; Watanabe, H.; Hattori, M.; Sakaki, Y.; Ishikawa, H. Genome sequence of the endocellular bacterial symbiont of aphids Buchnera sp. APS. Nature 2000, 407, 81–86. [Google Scholar] [CrossRef]
- Koga, R.; Tsuchida, T.; Fukatsu, T. Changing partners in an obligate symbiosis: A facultative endosymbiont can compensate for loss of the essential endosymbiont Buchnera in an aphid. Proc. R. Soc. B-Biol. Sci 2003, 270, 2543–2550. [Google Scholar] [CrossRef]
- Koga, R.; Tsuchida, T.; Sakurai, M.; Fukatsu, T. Selective elimination of aphid endosymbionts: Effects of antibiotic dose and host genotype, and fitness consequences. FEMS Microbiol. Ecol. 2007, 60, 229–239. [Google Scholar] [CrossRef] [PubMed]
- Oliver, K.M.; Moran, N.A.; Hunter, M.S. Variation in resistance to parasitism in aphids is due to symbionts not host genotype. Proc. Natl. Acad. Sci. USA 2005, 102, 12795–12800. [Google Scholar] [CrossRef]
- Chen, D.Q.; Montllor, C.B.; Purcell, A.H. Fitness effects of two facultative endosymbiotic bacteria on the pea aphid, Acyrthosiphon pisum, and the blue alfalfa aphid, A-kondoi. Entomol. Exp. Appl. 2000, 95, 315–323. [Google Scholar] [CrossRef]
- Russell, J.A.; Moran, N.A. Costs and benefits of symbiont infection in aphids: Variation among symbionts and across temperatures. Proc. R. Soc. B-Biol. Sci. 2006, 273, 603–610. [Google Scholar] [CrossRef]
- Scarborough, C.L.; Ferrari, J.; Godfray, H.C.J. Aphid protected from pathogen by endosymbiont. Science 2005, 310, 1781. [Google Scholar] [CrossRef] [PubMed]
- Yoram, G.; Inbar, M. Distinct antimicrobial activities in aphid galls on Pistacia atlantica. Plant Signal. Behav. 2011, 6, 2008–2012. [Google Scholar] [CrossRef]
- Gerchman, Y. Antimicrobial activities in Pistacia atlantica—Aphids make a difference! Multidiscip. Digit. Publ. Inst. Proc. 2020, 66, 9. [Google Scholar]
- Uchi, N.; Fukudome, M.; Nozaki, N.; Suzuki, M.; Osuki, K.-i.; Shigenobu, S.; Uchiumi, T. Antimicrobial activities of cysteine-rich peptides specific to bacteriocytes of the pea aphid Acyrthosiphon pisum. Microbes Environ. 2019, 34, 155–160. [Google Scholar] [CrossRef]
- Krämer, J.; Lüddecke, T.; Marner, M.; Maiworm, E.; Eichberg, J.; Hardes, K.; Schäberle, T.F.; Vilcinskas, A.; Predel, R. Antimicrobial, insecticidal and cytotoxic activity of linear venom peptides from the pseudoscorpion Chelifer cancroides. Toxins 2022, 14, 58. [Google Scholar] [CrossRef]
- Rashwan, R.S.; Alghamdi, A.S.; Althagafi, S.S. Morphological Identification of Aphid Species Infesting Some Ornamental plants in Taif Governorate. Egypt. Acad. J. Biol. Sci. A Entomol. 2016, 9, 15–35. [Google Scholar] [CrossRef]
- Sayed, S.M.; Ali, E.F.; Al-Otaibi, S.S. Efficacy of indigenous entomopathogenic fungus, Beauveria bassiana (Balsamo) Vuillemin, isolates against the rose aphid, Macrosiphum rosae L. (Hemiptera: Aphididae) in rose production. Egypt. J. Biol. Pest Control 2019, 29, 19. [Google Scholar] [CrossRef]
- Baazeem, A.; Alotaibi, S.S.; Khalaf, L.K.; Kumar, U.; Zaynab, M.; Alharthi, S.; Darwish, H.; Alghamdi, A.; Jat, S.K.; Al-Barty, A.; et al. Identification and environment-friendly biocontrol potential of five different bacteria against Aphis punicae and Aphis illinoisensis (Hemiptera: Aphididae). Front. Microbiol. 2022, 13, 961349. [Google Scholar] [CrossRef] [PubMed]
- Vela, J.; Montiel, E.E.; Mora, P.; Lorite, P.; Palomeque, T. Aphids and Ants, Mutualistic Species, Share a Mariner Element with an Unusual Location on Aphid Chromosomes. Genes 2021, 12, 1966. [Google Scholar] [CrossRef] [PubMed]
- Riddick, E.W. Identification of Conditions for Successful Aphid Control by Ladybirds in Greenhouses. Insects 2017, 8, 38. [Google Scholar] [CrossRef]
- Koutsoula, G.; Stamkopoulou, A.; Pekas, A.; Wäckers, F.; Broufas, G.; Pappas, M.L. Predation efficiency of the green lacewings Chrysoperla agilis and C. mutata against aphids and mealybugs in sweet pepper. Bull. Entomol. Res. 2023, 113, 162–168. [Google Scholar] [CrossRef] [PubMed]
- Huang, L.; Zhu, X.; Zhou, S.; Cheng, Z.; Shi, K.; Zhang, C.; Shao, H. Phthalic Acid Esters: Natural Sources and Biological Activities. Toxins 2021, 13, 495. [Google Scholar] [CrossRef]
- Bhattacharyya, M.; Dhar, R.; Basu, S.; Das, A.; Reynolds, D.M.; Dutta, T.K. Molecular evaluation of the metabolism of estrogenic di(2-ethylhexyl) phthalate in Mycolicibacterium sp. Microb. Cell Factories 2023, 22, 82. [Google Scholar] [CrossRef]
- Farooq, A.; Ali, S.; Ullah, H.; Khan, A.; Jahan, N.; Agha, I.; Tareen, R.; Bakhsh, R. Evaluation of Antioxidant, Antimicrobial activity and GC-MS analysis of Phlomis stewartii. Pure Appl. Biol. 2019, 8, 2420–2433. [Google Scholar] [CrossRef]
- Sayik, A.; YusufoĞLu, A.; Açık, L.; Türker, G.; Aydin, B.; Arslan, L. DNA-Binding, Biological Activities and Chemical Composition of Wild Growing Epilobium angustifolium L. Extracts from Canakkale, Turkey. J. Turk. Chem. Soc. Sect. A Chem. 2017, 4, 811–840. [Google Scholar] [CrossRef]
- Jansen, J.J.; Allwood, J.W.; Marsden-Edwards, E.; van der Putten, W.H.; Goodacre, R.; van Dam, N.M. Metabolomic analysis of the interaction between plants and herbivores. Metabolomics 2009, 5, 150–161. [Google Scholar] [CrossRef]
- Rivas-Ubach, A.; Peñuelas, J.; Hódar, J.A.; Oravec, M.; Paša-Tolić, L.; Urban, O.; Sardans, J. We Are What We Eat: A Stoichiometric and Ecometabolomic Study of Caterpillars Feeding on Two Pine Subspecies of Pinus sylvestris. Int. J. Mol. Sci. 2018, 20, 59. [Google Scholar] [CrossRef] [PubMed]
- Newman, D.J.; Cragg, G.M. Natural Products as Sources of New Drugs from 1981 to 2014. J. Nat. Prod. 2016, 79, 629–661. [Google Scholar] [CrossRef] [PubMed]
- Salehi, B.; Gultekin-Ozguven, M.; Kirkin, C.; Ozcelik, B.; Morais-Braga, M.F.B.; Carneiro, J.N.P.; Bezerra, C.F.; da Silva, T.G.; Coutinho, H.D.M.; Amina, B.; et al. Antioxidant, Antimicrobial, and Anticancer Effects of Anacardium Plants: An Ethnopharmacological Perspective. Front. Endocrinol. 2020, 11, 295. [Google Scholar] [CrossRef] [PubMed]
- Tuama, A.A.; Mohammed, A.A. Phytochemical screening and in vitro antibacterial and anticancer activities of the aqueous extract of Cucumis sativus. Saudi J. Biol. Sci. 2019, 26, 600–604. [Google Scholar] [CrossRef]
- Van Vo, G.; Guest, P.C.; Nguyen, N.H. Evaluation of Antimicrobial and Anticancer Activities of Bouea macrophylla Ethanol Extract. Methods Mol. Biol. 2022, 2343, 215–228. [Google Scholar] [CrossRef]
- Christina, A.; Christapher, V.; Bhore, S.J. Endophytic bacteria as a source of novel antibiotics: An overview. Pharm. Rev. 2013, 7, 11–16. [Google Scholar] [CrossRef]
- Gouda, S.; Das, G.; Sen, S.K.; Shin, H.S.; Patra, J.K. Endophytes: A Treasure House of Bioactive Compounds of Medicinal Importance. Front. Microbiol. 2016, 7, 1538. [Google Scholar] [CrossRef]
- Husain, D.R.; Wardhani, R. Antibacterial activity of endosymbiotic bacterial compound from Pheretima sp. earthworms inhibit the growth of Salmonella Typhi and Staphylococcus aureus: In vitro and in silico approach. Iran. J. Microbiol. 2021, 13, 537–543. [Google Scholar] [CrossRef]
- Makuwa, S.C.; Serepa-Dlamini, M.H. The Antibacterial Activity of Crude Extracts of Secondary Metabolites from Bacterial Endophytes Associated with Dicoma anomala. Int. J. Microbiol. 2021, 2021, 8812043. [Google Scholar] [CrossRef]
- Pasrija, P.; Girdhar, M.; Kumar, M.; Arora, S.; Katyal, A. Endophytes: An unexplored treasure to combat Multidrug resistance. Phytomed. Plus 2022, 2, 100249. [Google Scholar] [CrossRef]
- Gebhardt, K.; Schimana, J.; Muller, J.; Fiedler, H.P.; Kallenborn, H.G.; Holzenkampfer, M.; Krastel, P.; Zeeck, A.; Vater, J.; Holtzel, A.; et al. Screening for biologically active metabolites with endosymbiotic bacilli isolated from arthropods. FEMS Microbiol. Lett. 2002, 217, 199–205. [Google Scholar] [CrossRef] [PubMed]
- Tomar, P.; Thakur, N.; Yadav, A.N. Endosymbiotic microbes from entomopathogenic nematode (EPNs) and their applications as biocontrol agents for agro-environmental sustainability. Egypt. J. Biol. Pest Control 2022, 32, 80. [Google Scholar] [CrossRef]
- Gopi, M.; Ajith Kumar, T.T.; Balagurunathan, R.; Vinoth, R.; Dhaneesh, K.V.; Rajasekaran, R.; Balasubramanian, T. Phylogenetic study of sponge associated bacteria from the Lakshadweep archipelago and the antimicrobial activities of their secondary metabolites. World J. Microbiol. Biotechnol. 2012, 28, 761–766. [Google Scholar] [CrossRef] [PubMed]
- Husain, D.R.; Wardhani, R.; Erviani, A.E. Antibacterial activity of bacteria isolated from earthworm (Pheretima sp.) gut against Salmonella typhi and Staphylococcus aureus: In vitro experiments supported by computational docking. Biodivers. J. Biol. Divers. 2022, 22, 1125–1131. [Google Scholar] [CrossRef]
- Uche-Okereafor, N.; Sebola, T.; Tapfuma, K.; Mekuto, L.; Green, E.; Mavumengwana, V. Antibacterial Activities of Crude Secondary Metabolite Extracts from Pantoea Species Obtained from the Stem of Solanum mauritianum and Their Effects on Two Cancer Cell Lines. Int. J. Environ. Res. Public Health 2019, 16, 602. [Google Scholar] [CrossRef]
- Sawhney, S.; Mishra, J.K. Bioactive potential of bacterial endosymbionts isolated from Lamellodysidea herbacea, marine sponge from the coast of South Andaman, India, against human bacterial pathogens. J. Appl. Pharm. Sci. 2019, 9, 1–8. [Google Scholar]
- Mukherjee, A.; Bhattacharjee, P.; Das, R.; Pal, A.; Paul, A.K. Endophytic bacteria with plant growth promoting abilities from Ophioglossum reticulatum L. AIMS Microbiol. 2017, 3, 596–612. [Google Scholar] [CrossRef]
- Sadrati, N.; Daoud, H.; Zerroug, A.; Dahamna, S.; Bouharati, S. Screening of antimicrobial and antioxidant secondary metabolites from endophytic fungi isolated from wheat (Triticum durum). J. Plant Prot. Res. 2013, 53, 128–136. [Google Scholar] [CrossRef]
- Simon, J.-C.; Boutin, S.; Tsuchida, T.; Koga, R.; Le Gallic, J.-F.; Frantz, A.; Outreman, Y.; Fukatsu, T. Facultative Symbiont Infections Affect Aphid Reproduction. PLoS ONE 2011, 6, e21831. [Google Scholar] [CrossRef]
- Brumin, M.; Levy, M.; Ghanim, M. Transovarial transmission of Rickettsia spp. and organ-specific infection of the whitefly Bemisia tabaci. Appl. Environ. Microbiol. 2012, 78, 5565–5574. [Google Scholar] [CrossRef]
- Lv, N.; Wang, L.; Sang, W.; Liu, C.Z.; Qiu, B.L. Effects of Endosymbiont Disruption on the Nutritional Dynamics of the Pea Aphid Acyrthosiphon pisum. Insects 2018, 9, 161. [Google Scholar] [CrossRef] [PubMed]
- Oliver, K.M.; Russell, J.A.; Moran, N.A.; Hunter, M.S. Facultative bacterial symbionts in aphids confer resistance to parasitic wasps. Proc. Natl. Acad. Sci. USA 2003, 100, 1803–1807. [Google Scholar] [CrossRef]
- Rothacher, L.; Ferrer-Suay, M.; Vorburger, C. Bacterial endosymbionts protect aphids in the field and alter parasitoid community composition. Ecology 2016, 97, 1712–1723. [Google Scholar] [CrossRef] [PubMed]
- Ferdous, U.T.; Shishir, M.; Khan, S.N.; Hoq, M. Bacillus spp.: Attractive Sources of Anti-cancer and Anti-proliferative Biomolecules. Microb. Bioact. 2018, 1, 33–45. [Google Scholar] [CrossRef]
- Maela, M.P.; van der Walt, H.; Serepa-Dlamini, M.H. The Antibacterial, Antitumor Activities, and Bioactive Constituents’ Identification of Alectra sessiliflora Bacterial Endophytes. Front. Microbiol. 2022, 13, 870821. [Google Scholar] [CrossRef]
- Aboul-Soud, M.A.M.; Al-Amri, M.Z.; Kumar, A.; Al-Sheikh, Y.A.; Ashour, A.E.; El-Kersh, T.A. Specific Cytotoxic Effects of Parasporal Crystal Proteins Isolated from Native Saudi Arabian Bacillus thuringiensis Strains against Cervical Cancer Cells. Molecules 2019, 24, 506. [Google Scholar] [CrossRef]
- Assaeedi, A.S.; Osman, G.H. Screening and Identification of Bacillus thuringiensis Strains Native to Saudi Arabia that Exhibit Demonstrable Anticancer Activity. J. Pure Appl. Microbiol. 2017, 11, 119–128. [Google Scholar] [CrossRef]
- Chen, Y.T.; Yuan, Q.; Shan, L.T.; Lin, M.A.; Cheng, D.Q.; Li, C.Y. Antitumor activity of bacterial exopolysaccharides from the endophyte Bacillus amyloliquefaciens sp. isolated from Ophiopogon japonicus. Oncol. Lett. 2013, 5, 1787–1792. [Google Scholar] [CrossRef]
- Dolati, M.; Tafvizi, F.; Salehipour, M.; Movahed, T.K.; Jafari, P. Inhibitory effects of probiotic Bacillus coagulans against MCF7 breast cancer cells. Iran. J. Microbiol. 2021, 13, 839–847. [Google Scholar] [CrossRef]
- Shao, Y.; Wang, X.Y.; Qiu, X.; Niu, L.L.; Ma, Z.L. Isolation and Purification of a New Bacillus Subtilis Strain from Deer Dung with Anti-microbial and Anti-cancer Activities. Curr. Med. Sci. 2021, 41, 832–840. [Google Scholar] [CrossRef]
- Mohammed, G.J.; Kadhim, M.J.; Hussein, H.M. Characterization of Bioactive Chemical Compounds from Aspergillus terreus and Evaluation of Antibacterial and Antifungal Activity. Int. J. Pharmacogn. Phytochem. Res. 2016, 8, 889–905. [Google Scholar]
- Peng, W.; Li, D.; Zhang, M.; Ge, S.; Mo, B.; Li, S.; Ohkoshi, M. Characteristics of antibacterial molecular activities in poplar wood extractives. Saudi J. Biol. Sci. 2017, 24, 399–404. [Google Scholar] [CrossRef] [PubMed]
- Quoc, T.; Pham, L. Physicochemical properties and antibacterial activity of essential Oil of Ageratum conyzoides L. Leaves. Agric. Conspec. Sci. 2020, 85, 139–144. [Google Scholar]
- Su, Z.; Huang, H.; Li, J.; Zhu, Y.; Huang, R.; Qiu, S.X. Chemical Composition and Cytotoxic Activities of Petroleum Ether Fruit Extract of Fruits of Brucea javanica (Simarubaceae). Trop. J. Pharm. Res. 2013, 12, 735–742. [Google Scholar] [CrossRef]
- Adetutu, A.; Olaniyi, T.D.; Owoade, O.A. GC-MS analysis and in silico assessment of constituents of Psidium guajava leaf extract against DNA gyrase of Salmonella enterica serovar Typhi. Inform. Med. Unlocked 2021, 26, 100722. [Google Scholar] [CrossRef]
- Kang, M.S.; Choi, E.K.; Choi, D.H.; Ryu, S.Y.; Lee, H.H.; Kang, H.C.; Koh, J.T.; Kim, O.S.; Hwang, Y.C.; Yoon, S.J.; et al. Antibacterial activity of pyrrolidine dithiocarbamate. FEMS Microbiol. Lett. 2008, 280, 250–254. [Google Scholar] [CrossRef]
- Marinescu, M.; Popa, C.V. Pyridine Compounds with Antimicrobial and Antiviral Activities. Int. J. Mol. Sci. 2022, 23, 5659. [Google Scholar] [CrossRef]
- Bonomo, M.G.; Giura, T.; Salzano, G.; Longo, P.; Mariconda, A.; Catalano, A.; Iacopetta, D.; Ceramella, J.; Sinicropi, M.S.; Saturnino, C. Bis-Thiourea Quaternary Ammonium Salts as Potential Agents against Bacterial Strains from Food and Environmental Matrices. Antibiotics 2021, 10, 1466. [Google Scholar] [CrossRef]
- Karrouchi, K.; Radi, S.; Ramli, Y.; Taoufik, J.; Mabkhot, Y.N.; Al-Aizari, F.A.; Ansar, M. Synthesis and Pharmacological Activities of Pyrazole Derivatives: A Review. Molecules 2018, 23, 134. [Google Scholar] [CrossRef]
- Khloya, P.; Kumar, P.; Mittal, A.; Aggarwal, N.K.; Sharma, P.K. Synthesis of some novel 4-arylidene pyrazoles as potential antimicrobial agents. Org. Med. Chem. Lett. 2013, 3, 9. [Google Scholar] [CrossRef]
- Matiadis, D.; Sagnou, M. Pyrazoline Hybrids as Promising Anticancer Agents: An Up-to-Date Overview. Int. J. Mol. Sci. 2020, 21, 5507. [Google Scholar] [CrossRef] [PubMed]
- Carrillo, C.; Cavia Mdel, M.; Alonso-Torre, S.R. Antitumor effect of oleic acid; mechanisms of action: A review. Nutr. Hosp. 2012, 27, 1860–1865. [Google Scholar] [CrossRef] [PubMed]
- Giulitti, F.; Petrungaro, S.; Mandatori, S.; Tomaipitinca, L.; de Franchis, V.; D’Amore, A.; Filippini, A.; Gaudio, E.; Ziparo, E.; Giampietri, C. Anti-tumor Effect of Oleic Acid in Hepatocellular Carcinoma Cell Lines via Autophagy Reduction. Front. Cell Dev. Biol. 2021, 9, 629182. [Google Scholar] [CrossRef]
- Dias, C.J.F.; Raposo, A.; de Sousa, C.D.T.; de Araújo-Neto, J.B.; Tintino, S.R.; Oliveira-Tintino, C.D.d.M.; Araújo, I.M.; Coutinho, H.D.M.; Costa, M.G.M.; Lima, C.G.; et al. Modification of Antibiotic Activity by Fixed Oil of the Artocarpus heterophyllus Almond against Standard and Multidrug-Resistant Bacteria Strains. Biology 2022, 11, 1835. [Google Scholar] [CrossRef] [PubMed]
- Jumina, J.; Lavendi, W.; Singgih, T.; Triono, S.; Steven Kurniawan, Y.; Koketsu, M. Preparation of Monoacylglycerol Derivatives from Indonesian Edible Oil and Their Antimicrobial Assay against Staphylococcus aureus and Escherichia coli. Sci. Rep. 2019, 9, 10941. [Google Scholar] [CrossRef]
- Casillas-Vargas, G.; Ocasio-Malave, C.; Medina, S.; Morales-Guzman, C.; Del Valle, R.G.; Carballeira, N.M.; Sanabria-Rios, D.J. Antibacterial fatty acids: An update of possible mechanisms of action and implications in the development of the next-generation of antibacterial agents. Prog. Lipid Res. 2021, 82, 101093. [Google Scholar] [CrossRef]
- Youssef, A.M.M.; Maaty, D.A.M.; Al-Saraireh, Y.M. Phytochemical Analysis and Profiling of Antitumor Compounds of Leaves and Stems of Calystegia silvatica (Kit.) Griseb. Molecules 2023, 28, 630. [Google Scholar] [CrossRef]
- Lotfy, M.M.; Hassan, H.M.; Hetta, M.H.; El-Gendy, A.O.; Mohammed, R. Di-(2-ethylhexyl) Phthalate, a major bioactive metabolite with antimicrobial and cytotoxic activity isolated from River Nile derived fungus Aspergillus awamori. Beni-Suef Univ. J. Basic Appl. Sci. 2018, 7, 263–269. [Google Scholar] [CrossRef]
- To, N.B.; Nguyen, Y.T.; Moon, J.Y.; Ediriweera, M.K.; Cho, S.K. Pentadecanoic Acid, an Odd-Chain Fatty Acid, Suppresses the Stemness of MCF-7/SC Human Breast Cancer Stem-Like Cells through JAK2/STAT3 Signaling. Nutrients 2020, 12, 1663. [Google Scholar] [CrossRef]
- Torres-Castillo, J.A.; Olazarán-Santibáñez, F.E. Insects as source of phenolic and antioxidant entomochemicals in the food industry. Front. Nutr. 2023, 10, 1133342. [Google Scholar] [CrossRef]
- Musundire, R.; Zvidzai, C.; Chidewe, C.; Ngadze, R.; Macheka, L.; Manditsera, F.; Mubaiwa, J.; Masheka, A. Nutritional and bioactive compounds composition of Eulepida mashona, an edible beetle in Zimbabwe. J. Insects Food Feed 2016, 2, 179–187. [Google Scholar] [CrossRef]
- Alajmi, R.A.; Metwally, D.M.; El-Khadragy, M.F.; Yehia, H.M.; El-Ashram, S.; Almusawi, Z.; Amjad Bashir, M.; Alotaibi, N.J.; Abdel-Gaber, R. Molecular identification of Campanulotes bidentatus Scopoli, 1763 (Phthiraptera, Philopteridae) infecting the domestic pigeon Columba livia from Saudi Arabia. Saudi J. Biol. Sci. 2021, 28, 2613–2617. [Google Scholar] [CrossRef] [PubMed]
- Folmer, O.; Black, M.; Hoeh, W.; Lutz, R.; Vrijenhoek, R. DNA primers for amplification of mitochondrial cytochrome c oxidase subunit I from diverse metazoan invertebrates. Mol. Mar. Biol. Biotechnol. 1994, 3, 294–299. [Google Scholar] [PubMed]
- Syngai, G.G.; Barman, P.; Bharali, R.; Dey, S. BLAST: An introductory tool for students to Bioinformatics Applications. Keanean J. Sci. 2013, 2, 67–76. [Google Scholar]
- Gallo-Franco, J.J.; Duque-Gamboa, D.N.; Toro-Perea, N. Bacterial communities of Aphis gossypii and Myzus persicae (Hemiptera: Aphididae) from pepper crops (Capsicum sp.). Sci. Rep. 2019, 9, 5766. [Google Scholar] [CrossRef] [PubMed]
- Pandy, V.; Narasingam, M.; Vijeepallam, K.; Mohan, S.; Mani, V.; Mohamed, Z. The ethyl acetate fraction of a methanolic extract of unripe noni (Morinda citrifolia Linn.) fruit exhibits a biphasic effect on the dopaminergic system in mice. Exp. Anim. 2017, 66, 283–291. [Google Scholar] [CrossRef]
- Ahmed, D.; Saeed, R.; Shakeel, N.; Fatima, K.; Arshad, A. Antimicrobial activities of methanolic extract of Carissa opaca roots and its fractions and compounds isolated from the most active ethyl acetate fraction. Asian Pac. J. Trop. Biomed. 2015, 5, 541–545. [Google Scholar] [CrossRef]
- Tagne, R.S.; Telefo, B.P.; Nyemb, J.N.; Yemele, D.M.; Njina, S.N.; Goka, S.M.; Lienou, L.L.; Nwabo Kamdje, A.H.; Moundipa, P.F.; Farooq, A.D. Anticancer and antioxidant activities of methanol extracts and fractions of some Cameroonian medicinal plants. Asian Pac. J. Trop. Med. 2014, 7, S442–S447. [Google Scholar] [CrossRef]
- Morsi, A.E.; Ahmed, O.H.; Abdel-Hady, H.; El-Sayed, M.; Shemis, A.M. GC-analysis, and Antioxidant, Anti-inflammatory, and Anticancer Activities of Some Extracts and Fractions of Linum usitatissimum. Curr. Bioact. Compd. 2020, 16, 1306–1318. [Google Scholar] [CrossRef]
- Kirby, W.M.; Yoshihara, G.M.; Sundsted, K.S.; Warren, J.H. Clinical usefulness of a single disc method for antibiotic sensitivity testing. Antibiot. Annu. 1956, 892–897. Available online: https://pubmed.ncbi.nlm.nih.gov/13425478/ (accessed on 24 March 2023).
- Jenie, R.I.; Handayani, S.; Susidarti, R.A.; Udin, L.Z.; Meiyanto, E. The Cytotoxic and Antimigratory Activity of Brazilin-Doxorubicin on MCF-7/HER2 Cells. Adv. Pharm. Bull. 2018, 8, 507–516. [Google Scholar] [CrossRef]
- Thakur, A.N.; Thakur, N.L.; Indap, M.M.; Pandit, R.A.; Datar, V.V.; Müller, W.E. Antiangiogenic, antimicrobial, and cytotoxic potential of sponge-associated bacteria. Mar. Biotechnol. 2005, 7, 245–252. [Google Scholar] [CrossRef] [PubMed]
- Abd El-Kareem, M.; Rabbih, M.; Selim, E.; Elsherbiny, E.A.; El-Khateeb, A. Application of GC/EIMS in Combination with Semi-Empirical Calculations for Identification and Investigation of Some Volatile Components in Basil Essential Oil. Int. J. Anal. Mass Spectrom. Chromatogr. 2016, 4, 14–25. [Google Scholar] [CrossRef]
- Alsufyani, T.; Weiss, A.; Wichard, T. Time Course Exo-Metabolomic Profiling in the Green Marine Macroalga Ulva (Chlorophyta) for Identification of Growth Phase-Dependent Biomarkers. Mar. Drugs 2017, 15, 14. [Google Scholar] [CrossRef] [PubMed]
- Rajamanikyam, M.; Vadlapudi, V.; Parvathaneni, S.P.; Koude, D.; Sripadi, P.; Misra, S.; Amanchy, R.; Upadhyayula, S.M. Isolation and characterization of phthalates from Brevibacterium mcbrellneri that cause cytotoxicity and cell cycle arrest. EXCLI J. 2017, 16, 375–387. [Google Scholar]
- Habib, M.R.; Karim, M.R. Antimicrobial and Cytotoxic Activity of Di-(2-ethylhexyl) Phthalate and Anhydrosophoradiol-3-acetate Isolated from Calotropis gigantea (Linn.) Flower. Mycobiology 2009, 37, 31–36. [Google Scholar] [CrossRef]
- Koch, H.M.; Preuss, R.; Angerer, J.D. Di(2-ethylhexyl)phthalate (DEHP): Human metabolism and internal exposure—An update and latest results1. Int. J. Androl. 2006, 29, 155–165. [Google Scholar] [CrossRef] [PubMed]
- Lan, K.C.; Weng, T.I.; Chiang, W.C.; Chiu, C.Y.; Chan, D.C.; Yang, R.S.; Liu, S.H. Plasticizer Di-(2-ethylhexyl) Phthalate and Its Metabolite Mono(2-ethylhexyl) Phthalate Inhibit Myogenesis in Differentiating Mouse and Human Skeletal Muscle Cell Models. Appl. Sci. 2022, 12, 9195. [Google Scholar] [CrossRef]
- Jananie, R.K.; Priya, V.; Vijayalakshmi, K. Secondary metabolites of Cynodon dactylon as an antagonist to angiotensin II type1 receptor: Novel in silico drug targeting approach for diabetic retinopathy. J. Pharmacol. Pharmacother. 2012, 3, 20–25. [Google Scholar] [CrossRef]
- Abdussalam-Mohammed, W.; Mohamed, L.; Abraheem, M.S.; Mansour, M.M.; Sherif, A.M. Biofabrication of Silver Nanoparticles Using Teucrium Apollinis Extract: Characterization, Stability, and Their Antibacterial Activities. Chemistry 2023, 5, 54–64. [Google Scholar] [CrossRef]
- Abdel-Raheem, S.A.; El-Dean AM, K.; Hassanien, R.; El-Sayed, M.E.; Abd-Ella, A.A. Synthesis and biological activity of 2-((3-Cyano-4,6-distyrylpyridin-2- yl) thio) acetamide and its cyclized form. Alger. J. Biosceinces 2020, 1, 46–50. [Google Scholar] [CrossRef]




| EA-ME | Pathogenic Bacteria | |||||
|---|---|---|---|---|---|---|
| S. aureus | S. epidermidis | E. cloacae | K. pneumoniae | E. coli | ||
| Planococcus sp. | Inhibition zone | 17.00 ± 0.75 | 16.00 ± 0.91 | - | 11.00 ± 0.35 | - |
| MIC | 0.3125 | 0.3125 | 0.625 | |||
| MBC | 0.3125 | 0.3125 | 1.25 | |||
| Enterobacter sp. | - | - | - | - | - | |
| S. odorifera | - | - | - | - | - | |
| K. aerogenes | Inhibition zone | 15.00 ± 0.41 | 16.00 ± 0.29 | - | 20.00 ± 1.11 | 18.00 ± 0.65 |
| MIC | 0.625 | 0.3125 | 0.156 | 0.156 | ||
| MBC | 0.625 | 0.3125 | 0.625 | 0.3125 | ||
| S. fonticola | - | - | - | - | - | |
| P. stuartii | Inhibition zone | 09.00 ± 0.22 | - | - | - | - |
| MIC | 1.25 | |||||
| MBC | 2.5 | |||||
| E. avium | Inhibition zone | 10.00 ± 0.13 | 17.00 ± 0.37 | 11.00 ± 0.39 | - | - |
| MIC | 1.25 | 0.3125 | 0.625 | |||
| MBC | 2.5 | 0.625 | 1.25 | |||
| P. agglomerans | - | - | - | - | - | |
| B. megaterium | - | - | - | - | - | |
| B. safensis | Inhibition zone | 10.00 ± 0.45 | - | - | - | - |
| MIC | 1.25 | |||||
| MBC | 2.5 | |||||
| Ampicillin | 19.00 ± 0.52 0.3125 | 16.00 ± 0.95 0.625 | 15.00 ± 0.37 0.625 | - | 11.00 ± 0.33 0.625 | |
| Tetracycline | 35.00 ± 0.54 0.039 | 33.00 ± 0.36 0.039 | 25.00 ± 0.22 0.078 | 29.00 ± 0.63 0.078 | 30.00 ± 0.54 0.039 | |
| Compound Name | RT | Molecular Formula | Molecular Weight |
|---|---|---|---|
| 2-(2,5-Hexadiynyloxy) tetrahydro-2H-pyran ?? | 4.10 | C11H14O2 | 178 |
| Dimethyl diphenyl tethylidyl pyrrolidine ?? | 4.97 | C20H23N | 277 |
| 2-Aminoethanethiol hydrogen sulfate ester ?? | 6.27 | C2H7NO3S2 | 157 |
| 3-Methyl-4-nitro-5-(1-pyrazolyl) pyrazole ?? | 6.48 | C7H7N5O2 | 193 |
| 4a,5,8,8a-Tetrahydro-4,4a-dimethyl-2(1H)-naphthalenone ?? | 8.00 | C12H16O | 176 |
| Folinic acid ?? | 8.15 | C20H23N7O7 | 473 |
| 2,3-Dihydro-2-thioxo-3-diallylaminom ethyl benzoxazol ?? | 9.14 | C14H16N2OS | 260 |
| 1-(á-d-Arabinofuranosyl)-4-O-difluoro methyl uracil ??? | 14.79 | C10H12F2N2O6 | 294 |
| 4-(1,1-Dimethylethoxy) benzoic acid ?? | 15.05 | C11H14O3 | 194 |
| Methyl 13,16-octadecadiynate ? | 19.09 | C19H30O2 | 290 |
| (2-Phenyl-1,3-dioxolan-4-yl)methyl 9-octadecenoate ??? | 20.96 | C28H44O4 | 444 |
| Leukotriene D4 methyl ester ?? | 22.65 | C26H42N2O6S | 510 |
| Z-8-Methyl-9-tetradecenoic acid ?? | 22.72 | C15H28O2 | 240 |
| 9,10-Dihydroxy methyl octadecanoat?? | 23.38 | C19H38O4 | 330 |
| 2,3-Bis(acetyloxy)propyl dodecanoic acid,?? | 23.90 | C19H34O6 | 358 |
| Estra-1,3,5(10)-trien-17á-ol ?? | 24.80 | C18H24O | 256 |
| D-Fructose, diethyl mercaptal, pentaacetate ?? | 25.17 | C20H32O10S2 | 496 |
| 7,8,15,16-Tetramethyl-1,9-dio xacyclohexadeca-4,13-diene-2,10-dione ??? | 25.27 | C18H28O4 | 308 |
| D-Fructose, diethyl mercaptal, pentaacetate | 25.58 | C20H32O10S2 | 496 |
| (z,z,z)-6,9,12-Phenylmethyl octadecatrienoate, ester,? | 39.05 | C25H36O2 | 368 |
| Cholest-5-en-3-ol (3á) ? | 39.57 | C27H46O | 386 |
| Compound Name | RT | Molecular Formula | Molecular Weight |
|---|---|---|---|
| 5.Alpha pregnane-3.alpha.,11.beta.,20.beta.,21-tetrol, cyclic 20,21-(2-methyl-2-propaneboronate) ?? | 4.09 | C25H43BO4 | 418 |
| 2-Octyl methyl cyclopropanedodecanoiate ?? | 14.77 | C24H46O2 | 366 |
| Methyl-16-hydroxy-hexadecanoate ?? | 19.06 | C17H34O3 | 286 |
| Methyl hexandecanoiate ? | 22.11 | C17H34O2 | 270 |
| N-(3-Chlorobenzylidene)-10-undecenoic acid hydrazide ??? | 23.40 | C18H25ClN2O | 320 |
| Pyrrolo[1,2-a]pyrazine-1,4-dione, hexahydro-3-(2-methylpropyl) ?? | 24.85 | C11H18N2O2 | 210 |
| Pentadecanoic acid ?? | 24.95 | C15H30O2 | 242 |
| 13-Methyl oxacyclotetradecan-2-one ?? | 26.00 | C14H26O2 | 226 |
| Methyl-8,11-octadecadiynoic acid ?? | 27.14 | C19H30O2 | 290 |
| 1,2-Benzene dicarboxylic acid ? | 33.57 | C24H38O4 | 390 |
| Mono(2-ethylhexyl) phthalate ?? | 36.42 | C16H22O4 | 278 |
| Arachidonic acid ?? | 39.41 | C20H32O2 | 304 |
| Cholest-5-en-3-yl (9z)-9-octadecenoate ? | 39.57 | C45H78O2 | 650 |
| Compound Name | RT | Molecular Formula | Molecular Weight |
|---|---|---|---|
| 2-(2,5-Hexadiynyloxy) tetrahydro-2H-pyran ?? | 4.09 | C11H14O2 | 178 |
| N-(p-methoxybenzyl) acetamide ?? | 4.62 | C10H13NO2 | 179 |
| 1-(1-Cyclopenten-1-yl)- pyrrolidine ? | 14.74 | C12H16S | 192 |
| Methyl 13,16-octadecadiynoiate? | 19.07 | C19H30O2 | 290 |
| 2,4-Dimethyl hexanedioic acid ?? | 23.42 | C8H14O4 | 174 |
| Oleic acid ? | 24.87 | C18H34O2 | 282 |
| (Z)-9-Methyl octadecenoate? | 26.30 | C19H36O2 | 296 |
| Methyl cyclopentane undecanoiate ? | 26.81 | C17H32O2 | 268 |
| Didodecyl phthalate ? | 33.58 | C32H54O4 | 502 |
| 2-Aminoethanethiol hydrogen sulfate ? | 39.57 | C2H7NO3S2 | 157 |
| Insects | Insects’ Species | Collection Date | Endosymbiotic Bacteria | Accession Numbers |
|---|---|---|---|---|
| Pomegranate aphid | A. punicae | 27 June 2021 | Planococcus sp. | OP320679 |
| Bacillus megaterium | OQ351927 | |||
| Taif rose aphid | M. rosae | 11 September 2021 | Pantoea agglomerans | OP320678 |
| Bacillus safensis | OQ351925 | |||
| Grape aphid | A. illinoisensis | 21 May 2021 | Bacillus sp. | - |
| Lacewing | C. carnea | 27 June 2021 | Klebsiella aerogene | OP320677 |
| Serratia fonticola | OP320681 | |||
| Providencia stuartii | OP320680 | |||
| Eleven-spot ladybird | C. undecimpunctata | 27 June 2021 | Enterobacter sp. | OP320676 |
| Bacillus sp. | - | |||
| Serratia odorifera | OP320682 | |||
| Ant | T. magnum | 27 June 2021 | Bacillus sp. | - |
| Serratia odorifera | OP320682 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Alsufyani, T.; Al-Otaibi, N.; Alotaibi, N.J.; M’sakni, N.H.; Alghamdi, E.M. GC Analysis, Anticancer, and Antibacterial Activities of Secondary Bioactive Compounds from Endosymbiotic Bacteria of Pomegranate Aphid and Its Predator and Protector. Molecules 2023, 28, 4255. https://doi.org/10.3390/molecules28104255
Alsufyani T, Al-Otaibi N, Alotaibi NJ, M’sakni NH, Alghamdi EM. GC Analysis, Anticancer, and Antibacterial Activities of Secondary Bioactive Compounds from Endosymbiotic Bacteria of Pomegranate Aphid and Its Predator and Protector. Molecules. 2023; 28(10):4255. https://doi.org/10.3390/molecules28104255
Chicago/Turabian StyleAlsufyani, Taghreed, Najwa Al-Otaibi, Noura J. Alotaibi, Nour Houda M’sakni, and Eman M. Alghamdi. 2023. "GC Analysis, Anticancer, and Antibacterial Activities of Secondary Bioactive Compounds from Endosymbiotic Bacteria of Pomegranate Aphid and Its Predator and Protector" Molecules 28, no. 10: 4255. https://doi.org/10.3390/molecules28104255
APA StyleAlsufyani, T., Al-Otaibi, N., Alotaibi, N. J., M’sakni, N. H., & Alghamdi, E. M. (2023). GC Analysis, Anticancer, and Antibacterial Activities of Secondary Bioactive Compounds from Endosymbiotic Bacteria of Pomegranate Aphid and Its Predator and Protector. Molecules, 28(10), 4255. https://doi.org/10.3390/molecules28104255