Abstract
Energy transfer to ground state triplet molecular oxygen results in the generation of singlet molecular oxygen (1O2), which has potent oxidizing ability. Irradiation of light, notably ultraviolet A, to a photosensitizing molecule results in the generation of 1O2, which is thought to play a role in causing skin damage and aging. It should also be noted that 1O2 is a dominant tumoricidal component that is generated during the photodynamic therapy (PDT). While type II photodynamic action generates not only 1O2 but also other reactive species, endoperoxides release pure 1O2 upon mild exposure to heat and, hence, are considered to be beneficial compounds for research purposes. Concerning target molecules, 1O2 preferentially reacts with unsaturated fatty acids to produce lipid peroxidation. Enzymes that contain a reactive cysteine group at the catalytic center are vulnerable to 1O2 exposure. Guanine base in nucleic acids is also susceptible to oxidative modification, and cells carrying DNA with oxidized guanine units may experience mutations. Since 1O2 is produced in various physiological reactions in addition to photodynamic reactions, overcoming technical challenges related to its detection and methods used for its generation would allow its potential functions in biological systems to be better understood.
1. Introduction
Interactions of oxygen molecules with electrons leaked from enzymatic and non-enzymatic processes produces reactive oxygen species (ROS), such as superoxide (O2−•) and hydrogen peroxides (H2O2) [1,2,3]. Hydroxyl radicals (HO•) are the most reactive type of ROS and are likely produced by the reaction of H2O2 and ferrous iron, via the so called Fenton reaction [4], although there is some debate as to which ROS are the primary products in this reaction [5,6]. Singlet molecular oxygen (1O2), is a high-energy oxygen species but possesses unique properties that are different from other ROS [7,8,9]. While most ROS are produced by electron transfer reactions, 1O2 is generated by the transfer of energy to the ground state, triplet molecular oxygen (3O2), the most abundant oxygen molecule in atmosphere. The type II photodynamic reaction promoted by the presence of photosensitizing molecules is widely employed to generate 1O2 in biological systems [10,11]. However, the problem with the photodynamic action is that other ROS are also generated as byproducts [12,13]. This methodological issue appears to have hindered progress in research on the biological effects of 1O2, despite its importance.
1O2 possesses high energy and is considered to be a major cause for skin damage induced by ultraviolet (UV) irradiation [14,15]. Meanwhile, due to its strong cytotoxicity, 1O2 is the molecule that is responsible for killing tumor cells during photodynamic therapy (PDT) [10,11]. 1O2 as well as HO• preferentially reacts with conjugated double bonds, and hence polyunsaturated fatty acids (PUFA), which are dominantly present in the form of phospholipids in the cell membrane, are likely targets [9]. It is known that, upon mitotic stimuli, a small amount of ROS, notably H2O2, is produced, and this species modulates phosphorylation-mediated signaling pathways [16,17]. While signal modulation by H2O2 involves the transient oxidation of cysteine (Cys), reactions with 1O2 tend to result in irreversible oxidation. In most cases, exposure to 1O2 impairs cellular function but also occasionally stimulates tumorigenic cell growth [18,19]. Concerning cell death, results reported in many studies indicate that the apoptotic pathway is activated by 1O2 [20,21,22]. However, recent studies suggest that ferroptosis, an iron-dependent necrotic cell death [23,24], is also involved in 1O2-promoted cell death [25,26].
In this review article, we outline the characteristics and biological actions of 1O2 and then discuss cell injury and death induced by 1O2. Because details of the chemical reactions of 1O2 and its clinical applications to PDT are not the primary focus of this article, readers are referred to corresponding reviews on these issues [8,9,27].
2. Fundamental Knowledge of the Biochemical Properties of 1O2
Because 1O2 is a major oxidant that is responsible for photoaging and for tumor killing during PDT, numerous studies have been conducted from these perspectives in in vitro systems. It is important to understand its generation and reactions with biological molecules in order to achieve a correct evaluation of the significant roles of 1O2 in vivo.
2.1. Properties of 1O2 as a Potent Oxidant
Oxidative stress is induced by either the production of large amounts of ROS or an insufficient amount of antioxidants which include enzymes that eliminate ROS or small antioxidant compounds, such as glutathione (GSH), carotenoids and tocopherols [2]. Electrons that are leaked from enzymatic and non-enzymatic reactions initiate the generation of ROS, as represented by O2−•, H2O2 and HO•, and hence, the radical electron plays pivotal roles in the development of oxidative stress in many situations [1,3]. However, 1O2 is generated when the oxygen molecule in the ground triplet state 3O2 is excited by receiving energy without the transfer of an electron. The 1O2-generating system involves enzymatic reactions, such as myeloperoxidase, lipoxygenase and cyclooxygenase as well as chemical reactions, such as O2−•-mediated GSH oxidation and the interaction of peroxides with hypochlorite or peroxynitrite [13,28,29,30]. Photoaging and PDT are the subjects that have been most extensively investigated in terms of 1O2–mediated reactions that are associated with human physiology and the pathogenesis of related diseases. In spite of the high oxidizing power, reactions of 1O2 are thought to exert only limited effects compared to those of HO• in biological systems.
HO• is considered to be the most reactive ROS and appears to be responsible for a variety of pathological conditions. However, the half-life of HO• is quite short (10 nsec), so it only reacts with molecules that are in close proximity to the site where it is generated. On the other hand, the half-life of 1O2 is approximately 4 µsec in aqueous solution, which allows it to diffuse 150–220 nm [31,32]. Thus, 1O2 may react at various locations beyond where it is generated and, therefore, can affect surrounding molecules and organelles more widely compared to HO•. Nevertheless, this distance is insufficient for extracellularly produced 1O2 to move to the interior of a cell. Therefore, 1O2 that is generated inside the cell has the ability to damage various cellular components including DNA and organelles.
2.2. Chemical Probes for Detecting 1O2
Analyses employing a cell biological approach are essential for answering basic questions as to which part of the cell produces 1O2 in photoaging and during PDT and how cellular responses proceed in such situations. For that purpose, the use of a fluorescent chemical probe is the most convenient approach. 1O2 sensor green (SOSG) is a prototype that is popularly used in studies for detecting 1O2, although it has some disadvantages such as lack in membrane permeability [33]. Other compounds have been designed to overcome the disadvantage of SOSG. For example, Aarhus Sensor Green, which is a tetrafluoro-substituted fluorescein derivative that is covalently linked to a 9,10-diphenyl anthracene moiety [34] and the classic indocyanine green probe may also be applicable for this objective in certain experiments [35]. To increase the cellular delivery of SOSG, biocompatible nanosensors, with SOSG encapsulated within their hydrophobic core, have been developed, and these modifications appear to improve its delivery [36,37,38].
The cell membrane permeable far-red fluorescence probe Si-DMA, which is composed of silicon-containing rhodamine and anthracene moieties as a chromophore, has also been developed [39]. Upon reaction with 1O2, Si-DMA is converted into an endoperoxide at the anthracene moiety that emits strongly. The use of Si-DMA reportedly enables the visualization of 1O2 generated in a single mitochondrial tubule during PDT. After the treatment of cells with the endoperoxide, dose-dependent increases in fluorescence of Si-DMA were observed [40]. Thus, these results suggest that chemical probes may be applicable for studies concerning the cellular effects of 1O2. This compound is now commercially available. Another compound, a rhodamine 6G-aminomethylanthracene-linked donor–acceptor molecule (RA), was reported to exhibit unique properties [41]. RA acts as a fluorogenic 1O2 sensor molecule and also acts as a photosensitizer to generate 1O2 upon exposure to green light. Other fluorescent reagents, such as one based on an aminocoumarin-methylanthracene-based electron donor–acceptor molecule [42], are also being developed. Since information on the use of these newly developed probes is currently limited, it is necessary to carefully choose which compounds are suitable for the intended research.
2.3. Photodynamic Reaction as a 1O2 -Generating System
PDT mainly contributes to 1O2 generation in biological systems through type II mechanisms that involve energy transfer from triplet excited molecules to triplet oxygen. Photosensitizers may also act according to competitive type I photosensitization mechanism that mostly involves charge transfer between suitable targets and a photosensitizer in its triplet excited state [43,44]. In order to protect cells against the deteriorating action of UV light, the effects of 1O2 on skin tissues have been extensively investigated. In the meantime, 1O2 is considered to be the main molecule that promotes cytotoxic processes during PDT, and hence, multiple studies are currently underway with the aim of understanding the mechanism responsible for 1O2-mediated cell death and developing efficient photosensitizers [8,10]. UV radiation causes skin photoaging and oxidatively generated damage to dermal cells and is especially troublesome in cases of sunburn which occurs by exposure to excessive UV for long periods of time [14,45]. UVB (280–315 nm) comprises approximately 5% of the solar UV and causes the direct photodamage to many molecules including DNA and proteins in skin tissues through its high energy photochemical reactions. Genetic damage caused by the oxidative modification of DNA and other molecules emerges in a short time after exposure to UVB. In the case of UVA (315–400 nm) that accounts for approximately 95% of the solar UV, cellular damage occurs through the activation of chromophores that act as photosensitizers to generate 1O2 and other ROS, and hence, the oxidative reaction proceeds indirectly via the ROS that are generated. It is rather difficult to determine if changes in cells that have been exposed to UVA are the consequence of the generation of either 1O2 or other ROS because they are produced simultaneously by the photodynamic reactions and result in essentially the same end products [46].
In order to elucidate the reactions caused by 1O2, reliable methods for generating 1O are required [12]. The most common method for this purpose is irradiation of the photosensitizer with UV or visible light because it is simple and easy to control its production [27,47]. Figure 1 represents the “Jablonski diagram” that depicts conceptual images of the generation of 1O2 by the irradiation of a photosensitizing molecule (S) with light followed by transferring energy to 3O2 [48]. When a photosensitizer is exposed to light, most likely the UVA in natural light, photon energy converts the photosensitizer in the ground state (0S) to that in the excited state (1S). On returning to the ground state 0S, a part of the released photoenergy can be transferred to 3O2, which results in the electron spin state being altered and the generation of 1O2. Under this situation, photodynamic action generates not only 1O2 but also other ROS such as O2−• and HO• [8].
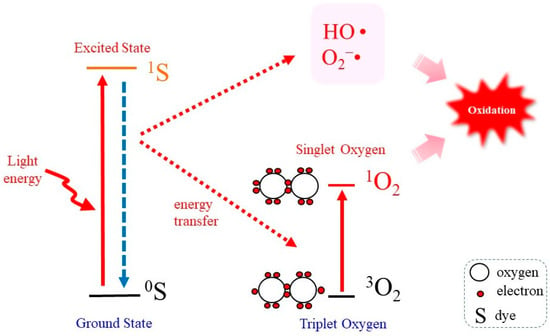
Figure 1.
Photodynamic 1O2 generation. Photoirradiation of a photosensitizing molecule in the ground state (0S) leads the production of the excited form (1S). On returning to the ground state, energy is transferred to 3O2, which becomes excited to 1O2. In the meantime, however, other ROS such as O2−• and HO• may also be produced.
To observe cellular responses to 1O2, cell-permeable and non-cytotoxic compounds need to be used as the photosensitizer. For example, Rose Bengal and methylene blue meet the conditions and, hence, are popularly used for the purpose of examining biological action of 1O2 [47]. Since PDT is a useful therapeutic for eliminating tumors, many attempts have been made to improve the treatment by developing convenient photosensitizers [11,27,49]. This issue, i.e., the applications of photosensitizers, is not discussed further because it is beyond the scope of this review, which is focused on the underlying mechanisms concerning the biological action of 1O2.
2.4. Endoperoxides as Donor Compounds for Generating Pure 1O2
Endoperoxides that release 1O2 without other ROS have been developed to evaluate its unique reaction [13,50]. Naphthalene endoperoxide-based 1O2 donor compounds were first developed, and the structure–function relationships of the compounds have been described in detail in a review article [13]. Consequently, several naphthalene endoperoxides have been established and utilized for the in vitro evaluation of the effects of 1O2, as representative structures in Figure 2A. Upon mild heating at 37 °C, the endoperoxides spontaneously release pure 1O2, which then directly reacts with surrounding compounds and organelles. Heat-labile endoperoxides are considered clean sources of 1O2 for highly specific oxidation of cellular biomolecules and have been applied for 1O2-mediated oxidation of the DNA guanine base in cells [51,52].
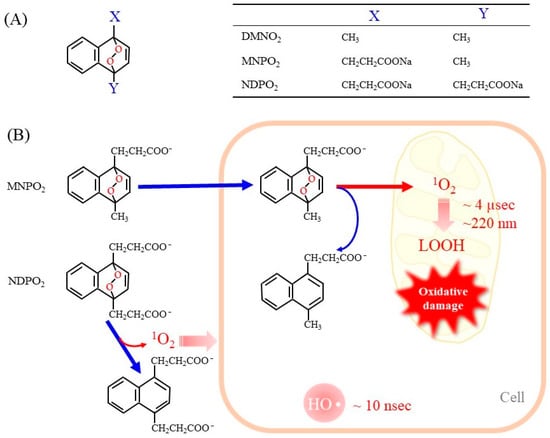
Figure 2.
Representative endoperoxides and the release of 1O2. (A) The general structure of naphthalene endoperoxides is shown on the left. The table on the right shows the chemical groups attached to the naphthalene ring. DMNO2, 1,4-dimethylnaphthalene endoperoxide; MNPO2, 1-methylnaphthalene-4-propanonate endoperoxide; NDPO2, 1,4-naphthalenedipropanoate endoperoxide. (B) Endoperoxides represented by MNPE and NDPE generate 1O2 spontaneously at physiologic temperature 37 °C. MNPE, which is relatively hydrophobic, can enter the cell, but NDPE cannot cross the cell membrane. The short life of 1O2 (~4 µsec) makes it diffuse only 150–220 nm in aqueous solution. As a result, 1O2 released from NDPE is present outside the cell only, while 1O2 from MNPE can act inside the cell. For reference, hydroxy radicals are also shown.
Here we discuss the advantages and disadvantages of 1O2 donor compounds in comparison with the photodynamic action. The benefits include the following: (1) Endoperoxides produce pure 1O2. (2) The concentrations of the released 1O2 are easily controlled. (3) Heating at physiological temperature, generally under cell culture conditions, can promote the release of 1O2 from endoperoxides. (4) It may be possible to design endoperoxides that are localized to a specific organelle by appropriate chemical modification of the compounds. Limitations include the following: (1) A high concentration of endoperoxides is required to generate sufficient levels of 1O2. (2) It is essential to consider effects of the raw material after the release of 1O2 because they are sometimes toxic to cells. (3) Endoperoxides may not be evenly distributed inside cells due to their chemical nature. (4) The amount of 1O2 released is initially maximal but then gradually decreases with increasing endoperoxide consumption.
When endoperoxides are applied to a cell culture system, it is necessary to use compounds that are able to pass through the cell membrane. In fact, the side chains of naphthalene endoperoxides determine whether they enter cells or remain outside of cells (Figure 2B) [53]. 1-Methylnaphthalene-4-propanonate endoperoxide (MNPO2) is cell membrane-permeable and generates 1O2 within cells. However, 1,4-naphthalenedipropanoate endoperoxide (NDPO2) cannot enter cells. Accordingly, while MNPO2 induces cell damage, NDPO2 at the same concentration has no effects, although both compounds trigger the release of cyt c from isolated mitochondria to a similar extent [53]. We have not examined 1,4-dimethylnaphthalene endoperoxide (DMNO2), but it can also enter cells due to the hydrophobic property of its side chains.
While the generation of 1O2 by a light-irradiated photosensitizer is frequently used, the use of 1O2 donor compounds has been limited because they are complex molecules that are difficult to synthesize. Some naphthalene endoperoxides are now commercially available. New compounds other than naphthalene-based endoperoxides are also being developed. For the efficient delivery of a 1O2 donor to cancer cells, a porphyrin-based covalent organic framework that contains a naphthalene endoperoxide has also been developed [54]. Trials to develop new types of endoperoxides, which are based on 2-pyridone and anthracene, are also underway [50]. Two 1O2-producing systems, photodynamic reactions and naphthalene endoperoxides, have provided rather consistent results so far [13], implying that the contribution of other byproducts may be negligible. Development of these systems will hopefully accelerate our understanding of the nature of 1O2.
2.5. Natural or Synthetic 1O2-Scavenging Compounds
The body is protected from oxidatively generated damage by a variety of natural and synthetic compounds that scavenge 1O2. The quantitative evaluation of the 1O2-scavenging ability of a compound provides useful information not only for basic research but also for developing functional foods and medicines concerning antioxidation [55]. Many nutritional compounds, such as tocopherols, carotenoids and flavonoids, possess antioxidant capacity and protect susceptible molecules from 1O2. The oxygen radical absorption capacity (ORAC) assay is a representative method for the detection of 1O2-scavenging ability of food ingredients [56]. Thereafter, a simple method called a singlet oxygen absorption capacity (SOAC) assay has been established for the evaluation of 1O2-scavenging ability [56,57]. These methods are useful in exploring popularly used 1O2-scavenging compounds, especially in the field of food chemistry.
While carotenoids react with 1O2 more rapidly than α-tocopherol, in a nearly diffusion-limited manner (~1010 M−1s−1) [58,59], lycopene, which is found in fruits and vegetables such as tomato, is one of the strongest natural carotenoids [60]. By employing the SOAC assay method, carotenoids have been found to quench 1O2 approximately 30–100 times faster than α-tocopherol [57]. After transferring excitation energy to carotenoids, 1O2 returns to the ground state. The excited carotenoids spontaneously release thermal energy and then return to the ground state. Hence, carotenoids are spontaneously recycled and have the advantage of quenching 1O2 without affecting other molecules. This chemical property of 1O2 is a major difference from other radicals that are generated by electron transfer reactions that require another radical to quench.
Based on in vitro data on the action of anti-oxidants, the biological benefit of carotenoids has also been examined by some studies on humans. The administration of a representative carotenoid β-carotene to humans failed to alleviate sunburn reactions under the conditions used [61]. However, a later study reported that carotenoids, β-carotene and lycopene effectively protect erythema formation induced using a solar light simulator [62,63]. Lycopene has also attracted attention as a nutrient with anticancer effects [64]. For another example, lutein is a xanthophyll carotenoid that is found in foods such as dark green leafy vegetables and exhibits strong antioxidant activity via its ability to scavenge ROS including 1O2 and lipid peroxyl radicals [65]. Lutein appears to exert anti-inflammatory actions against some diseases, including neurodegenerative disorders, eye diseases, cardiovascular diseases and skin diseases.
Many synthetic antioxidant compounds that scavenge ROS including 1O2 have been developed as medicines. Edaravone (3-methyl-1-phenyl-2-pyrazolin-5-one) is a compound that eliminates a variety of radical species and was the first approved compound for use as a medicine for the treatment of acute brain infarctions. Edaravone can scavenge 1O2 that is generated by activated human neutrophils [66] and by photoactivated Rose Bengal [67]. The plasma lipid peroxidation caused by 1O2, however, cannot be suppressed by edaravone and other clinical drugs with antioxidant ability, which include roglitazone, probucol, carvedilol, pentoxifylline and ebselen, although they exhibit suppressive effects on lipid peroxidation caused by free radicals, peroxynitrite, hypochlorite, and lipoxygenase reactions [68]. Because blood plasma contains high concentrations of proteins and many other compounds that could potentially interfere with the scavenging reaction by these chemicals, such biological compounds may have influenced the results.
3. Oxidative Modification of Biological Molecules and Damage to Organelles by 1O2
1O2 preferentially reacts with double bonds but can also react with sulfhydryl groups of Cys in proteins. In fact, 1O2 oxidizes important biological molecules including lipids, proteins and nucleic acids, while oxidation end products by 1O2 are largely similar to those produced by other ROS.
3.1. Lipid Peroxidation by 1O2
Both 1O2 and HO• effectively oxidize lipids, leading to the formation of lipid hydroperoxides (LOOH), although the pathways and efficiency of their production differ between these oxidants [2,8]. Because biological membranes contain large quantities of phospholipids and cholesterol, their oxidative modification may cause the destruction of the cell membrane. Mono-unsaturated fatty acids (MUFA) and poly-unsaturated fatty acids (PUFA) are both targets of 1O2, but PUFA are among the preferred target that is present in the cell membrane as well as lipid droplets in the form of triacylglycerol molecules. 1O2 also oxidizes cholesterol, which results in the formation of several oxidized products that are collectively referred to as oxysterols [69,70,71]. Accordingly, cholesterol hydroperoxides are specific biomarkers of type I and type II photosensitization reactions [72,73]. Phospholipids generally contain unsaturated fatty acids at the sn-2 position, which may undergo oxidation by 1O2 (Figure 3A). In the presence of metal ions or HOCl, P-LOOH may also generate 1O2 based on a Russell mechanism [74]. Because the resultant oxidation products are structurally the same irrespective of the causal oxidant, 1O2 and HO•, it is difficult to determine which reactive species actually caused the lipid peroxidation.
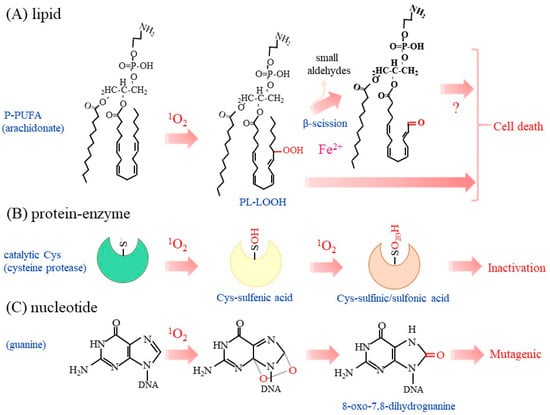
Figure 3.
Oxidation of biological molecules by 1O2. Typical oxidation reaction of a phospholipid, enzymes, and nucleic acids with 1O2 are depicted. (A) Lipid peroxidation preferentially occurs in PUFA in phospholipids, as typified by arachidonic acid. The presence of metal ions, as exemplified by iron, may initiate radical chain reactions and cause the β-scission of LOOH. Cell death may be induced by the accumulated P-LOOH, although it is not clear whether or not β-scission is essential. (B) The sulfhydryl group in the catalytic Cys residue of enzymes is most sensitive to 1O2–induced oxidation in proteins. The Cys-OH may be hyperoxidized to Cys-O2H and then Cys-O3H, which are collectively presented as Cys-O2/3H. (C) Purine bases, notably guanine, are prone to oxidation by 1O2. 8-oxo-7,8-dihydroguanine is consequently produced and may induce mutation in DNA.
Following the peroxidation of lipids, the β-scission of LOOH may release small aldehyde molecules, as represented by 4-hydroxyl nonenal, which impairs cellular function but may also act as a signaling molecule [46,75,76]. The accumulation of P-LOOH along with iron may consequently induce cell rupture [77], which is typically observed in ferroptosis [23,24]. Although the accumulation of lipid-derived aldehydes, such as 4-hydroxyl nonenal and malondialdehyde, is considered to be a hallmark of ferroptosis [78,79], it is not clear at this moment if the β-scission of P-LOOH is essential or not for executing cell death. LOOH produced by 1O2 during PDT along with iron may produce lipid radicals LOO• and then initiate a chain reaction, which leads to cell death [80]. Nitric oxide reacts with LOO• to form LOONO, which terminates the chain reaction and hence suppresses cell death [81]. Anti-cancer chemotherapy generally induces cell death by means of the production of ROS, while some malignant tumors overexpress nitric oxide synthase (NOS). These collective observations suggest that the inhibition of nitric oxide synthesis would increase the efficacy of killing the NOS-expressing tumors during PDT [82].
3.2. Oxidation of Amino Acids and Proteins by 1O2
Proteins are present in high concentrations in the cytosol and organelle membranes. Side chains of amino acids in proteins contain reactive groups which reportedly consume approximately 68% of the 1O2 that is generated inside cells. 1O2 selectively reacts with only a few amino acids in proteins, which include Cys, methionine (Met), histidine (His), tyrosine (Tyr) and tryptophan (Trp) [8]. When cyt c is employed as a model protein, Met, His and Trp were reported to be preferentially oxidized by the 1O2 generated by the photoexcitation of phthalocyanine Pc 4 [83]. Interestingly, the same amino acids in lysozyme are oxidized by the 1O2 released from endoperoxides [84]. The oxidation of the same amino acids, despite the use of different 1O2 sources and different model proteins, suggests that these amino acids are also susceptible targets of 1O2 in other proteins as well.
While sulfhydryl groups in isolated proteins are largely present in the form of disulfide bonds, this makes them resistant to further oxygenation. Intracellular proteins mainly contain Cys in the reduced form (Cys-SH) due to the reducing environment inside cells. Oxidation converts Cys and GSH to the oxidized Cys dimer (cystine) and oxidized glutathione dimer (GSSG), respectively. The oxidation of sulfhydryl groups in proteins yields three additional forms of oxidation products, depending on the redox microenvironment [85]. In the absence of nearby Cys or GSH, Cys-SH is oxidized to cysteine sulfenic acid (Cys-SOH), cysteine sulfinic acid (Cys-SO2H) or cysteine sulfonic acid (Cys-SO3). While Cys-SOH can be reduced back to Cys-SH by means of reductants such as GSH, Cys-SO2H and Cys-SO3H cannot be reduced to Cys-SH in mammalian cells. Accordingly, enzymes with reactive Cys residues in the catalytic center are permanently inactive after hyperoxidation to Cys-O2/3H.
Lysosomes are organelles that hydrolyze polymeric compounds that are incorporated via endocytosis and those encapsulated within autolysosomes during autophagy. Cathepsins are a series of lysosomal acidic proteases and are classified into three groups, based on their catalytic amino acids: serine proteases, cysteine proteases and aspartate proteases. Cathepsins B and L/S belong to the cysteine protease group and are abundantly present in many mammalian tissues [86], while cathepsins D/E are representatives of the aspartate protease group [87]. The release of 1O2 from a naphthalene endoperoxide selectively inactivates isolated cysteine proteases cathepsins B and L/S but not aspartate proteases cathepsins D/E in vitro [88]. Serine proteases, trypsin and chymotrypsin, as well as other cysteine proteases, such as papain and calpain II, have also been reported to be inactivated by 1O2 [89]. The hydroxyl group in the side chain of serine (Ser) is generally less reactive compared to Cys-SH, but the tertiary structure of the catalytic center of a serine protease, named the serine-histidine-aspartate catalytic triad, makes it highly reactive, which enables electrophilic attack at the peptide bond of the substrate protein. As a result of these properties, the hydroxyl group of the catalytic Ser is highly susceptible to oxidative insult, as has also been found for the catalytic Cys in cysteine proteases. This mechanism may explain the high susceptibility of trypsin and chymotrypsin to 1O2 and results in a high sensitivity comparable to that for cysteine proteases [89].
Phosphotyrosine phosphatases (PTP) are another enzyme family that possesses reactive Cys for catalysis, as found in cysteine proteases. The high sensitivity to oxidants of the catalytic Cys in PTP allows modulation of the phosphorylation signal by hydrogen peroxide [16,17]. 1O2 from the endoperoxide oxidatively inactivates protein tyrosine phosphatase-1B (PTP1B), which plays a pivotal role in cellular signal transduction and undergoes redox regulation [90]. A mass-spectrometric analysis identified Cys-OH and Cys-O2H as oxidized amino acids in 1O2-treated PTP1B [91]. Thus, these Cys-centered enzymes appear to be common targets of 1O2 (Figure 3B). This issue is discussed further below in explaining the biological action of 1O2.
3.3. Oxidation of Nucleotides by 1O2
UVB can lead to the development of skin cancer. The incidence of skin cancer is greatly increased in xeroderma pigmentosum (XP) patients who have defects in the nucleotide excision repair system [92]. However UVB itself is considered to be a very poor DNA oxidizing agent in most living systems. 1O2 causes the oxidative modification of nucleotides [93,94]. 2′-Deoxyguanosine is the most reactive nucleoside compared to other nucleosides and produces 8-oxo-7,8-dihydro-2′-deoxyguanosine (8-oxodG) (Figure 3C). The rate of reaction of 1O2 with 2′-deoxyguanosine is approximately two orders of magnitude faster than that with other nucleosides [8]. This trend is similar to that for the oxidative modification of 2′-deoxyguanosine by HO•, which also produces 8-oxodG [95]. The presence of an 8-oxodG residue in DNA frequently causes a G to T transversion, which is associated with mutations of genes and may lead to the development of tumors. UVB radiation dominantly causes dimeric pyrimidine photoproducts, while 8-oxoG represents at best 1% of the overall amount of dimeric pyrimidine photoproducts generated by exposure to UVB radiation [96,97]. Accordingly, UVB-induced skin cancer is essentially due to the photo-induced formation of cytosine-containing bipyrimidine photoproducts but not 8-oxoG formation. Meantime UVA generates 1O2 with respect to HO•-mediated oxidation as well as cyclobutane pyrimidine dimers as the main photoproducts. Thus, UVA is considered to be a better DNA-oxidizing agent [98]. These collective findings imply that the excessive exposure to UV, not only to UVB but also to UVA, may increase the incidence of skin cancer [99]. 1O2 also oxidizes NAD(P)H to NAD(P)+, which appears to be associated with dysfunctional mitochondrial metabolism [100].
While intact DNA is not antigenic in the general sense, that treated with 1O2 exhibits a high immunogenicity and may become an epitope for autoantibodies in autoimmune diseases such as systemic lupus erythematosus (SLE) [101]. Consistent with this mechanism, a neoepitope is, in fact, generated by 1O2 exposure and can be recognized by immunoglobulin G [92,102]. Exposure to UVA in the presence of excessive endogenous photosensitizing molecules, such as porphyrin, increases the generation of 1O2, which may also increase the incidence of skin cancer [94]. Porphyrin-related compounds accumulate in porphyria that are divided into eight distinct groups by the defect in the genes involved in the synthesis of heme. Accumulated porphyrin-related compounds act as photosensitizers upon exposure to sunlight. Generated ROS including 1O2 are considered to be largely involved in the hypersensitivity of skin in porphyria, while protein aggregation induced by direct interactions with porphyrin-related compounds appears to play roles in pathogenic damage in internal organs such as the liver [103].
3.4. Organelle Damage by 1O2
Concerning oxidatively generated damage in cells, some organelles are also sensitive targets of 1O2, and mitochondria and lysosomes are the primary target organelles of 1O2 generated by PDT [104]. A sublethal dose of UVA irradiation causes the common deletion in mitochondrial DNA in human fibroblasts, which can be attributable to the generation of 1O2 within mitochondria [105]. Mitochondrial permeability transition is either activated or inactivated by 1O2, depending on the site where the 1O2 is produced [106], whereas Bcl-2 may also be a sensitive target of 1O2 and responsible for apoptosis [107]. Irradiation with a 1267 nm laser produces 1O2 without other ROS and induces apoptosis by opening mitochondrial permeability transition in B16 melanoma cells but has no effects on fibroblasts [22]. In the case of B16 melanomas, melanin and its precursor molecules, which act as both photoprotectors and photosensitizers [108], are abundantly produced and, upon laser irradiation, may exert an influence on the sensitivity of the cells.
1O2 largely oxidizes mitochondrial membranes, leading to the stimulation of autophagic degradation of damaged mitochondria, a process that is referred to as mitophagy [109,110,111]. Moreover, lysosomes are also one of sensitive targets of PDT [112]. The destruction of lysosomes by oxidatively generated damage during PDT releases a variety of hydrolytic enzymes that may cleave cytosolic components including organelle proteins and lipids. Because lysosomes store relatively large amounts of iron, which is originated from transferrin through endosomes [113], the stored iron is also released during PDT. Because iron is involved in the production of oxygen radical species including HO•, its release accelerates cell dysfunction and death [6]. Thus, the destruction of lysosomes along with mitochondrial damage coordinately stimulates cytotoxic effects and enhances the therapeutic efficiency of PDT [114,115]. Based on this mechanism, photosensitizers that target mitochondria or lysosomes are considered to be more effective in tumoricidal action [116,117].
4. Cellular Responses to 1O2
The high reactivity of 1O2 is not only a tumoricidal component of PDT but is also applicable for sterilizing microorganisms [118,119,120]. The generation of excess levels of 1O2 causes cell damage and death [112,121], whereas physiological levels of 1O2 likely act as modulators of cellular signaling, as typically seen in the action of hydrogen peroxide [16].
4.1. O2 in Protection against Microbial Infection
ROS and reactive nitric oxide species (RNOS) are recruited by the immune system to combat microbial infections. Neutrophil extracellular traps (NET) play pivotal roles in innate host defense by enveloping invading microbes. 1O2 is produced upon the stimulation of neutrophils with phorbol myristate acetate and is essential for NET formation, independent of ROS production by activated NADPH-oxidase [122]. Meanwhile, 1O2 generated by either the enzymatic system or from naphthalene endoperoxides directly kills bacteria [118,123]. Accordingly, the antibacterial ability of 1O2 generated by the photodynamic action has attracted much attention for the purpose of killing multidrug-resistant pathogenic bacteria [10].
Anti-viral action is another beneficial reaction of 1O2. 1O2 inhibits infections without damaging viral DNA under the conditions used [100] and may be useful in exterminating viruses in blood components [124]. Efforts are being made to develop photosensitizers that exhibit antiviral actions more effectively by targeting the envelope lipids of viruses [120]. For the purpose of anti-microbial infections, photosensitizers that are activatable by near-infrared light, which penetrates deep into tissue, are being developed and have become another approach to the use of 1O2 [125].
4.2. Redox Modulation of Mitotic Signaling by Oxidants
The role of ROS in mitotic signal transduction, notably in mitotic exit by means of phosphatases, has been elucidated over the past three decades [16,17,126] as shown in Figure 4. Regarding ROS signaling in growth stimuli, a pioneering study on the regulation of PTP1B was performed on cells that had been stimulated with EGF growth factor [127]. In order to understand the potential mechanism for signal regulation by 1O2, we first described the signal transduction system for growth stimulation followed by its modulation by H2O2, a major oxidant with moderate oxidizing properties.
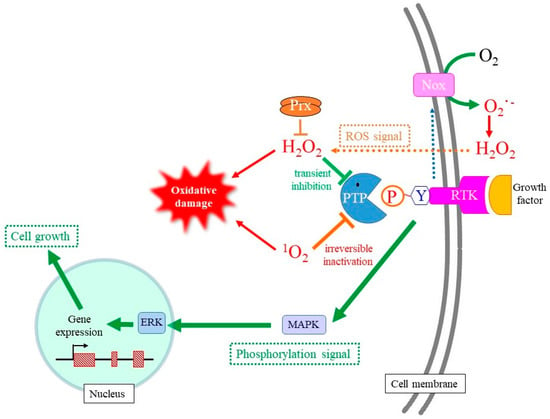
Figure 4.
Stimulation of cell growth by hydrogen peroxide and 1O2. Activation of RTK stimulates the production of O2−• by NADPH-oxidation and the subsequent formation of H2O2 via the spontaneous dismutation or an SOD-catalyzed reaction. Because hydrogen peroxide transiently inhibits PTP via the oxidation of catalytic Cys-SH to Cys-SOH, mitotic signaling mediated by Tyr phosphorylation is transmitted to a following signaling molecule. As result, the transient production of hydrogen peroxide upon mitotic stimuli promotes cell growth. 1O2 also oxidatively modifies the catalytic Cys-SH in PTP, but overoxidation converts it to Cys-O2/3H, which cannot be reduced back to Cys-SH. Thus, 1O2 likely induces the permanent inactivation of PTP, which makes mitotic signaling uncontrollable and leads to tumorigenic cell growth.
Growth factors bind to the corresponding plasma membrane receptor for tyrosine kinase (RTK) at the cell surface and transmit mitotic signals to genes in the nucleus through the phosphorylation of signaling molecules [17,128]. Two types of phosphorylation can occur in amino acid side chains of proteins: Ser/threonine (Thr) and Tyr, whereas the phosphatidylinositide 3-kinase (PI3K)-Akt pathway also involves the phosphorylation of phosphatidylinositol. The binding of the growth factor at extracellular space initiates a signal cascade that proceeds via the autophosphorylation of specific Tyr residues in the cytosolic domain of RTK [128]. The binding of adaptor proteins to phosphotyrosine converts GDP-bound Ras to GTP-bound Ras, allowing the activation of Raf, the most upstream protein kinase in the signaling pathway in mitotic signaling. In the meantime, PI3K is also activated, which results in the stimulation of the protein kinase B/Akt pathway in the case of certain mitogens [11]. The expressions of a series of genes are ultimately stimulated and promote the cell cycle for growth.
The coordinated action of phosphorylation and dephosphorylation reactions of signaling molecules functions to control cell growth properly. While activation of the signaling cascade is corroborated by phosphorylation, dephosphorylation catalyzed by a phosphatase is needed to terminate the signaling to avoid excessive proliferation [129]. Most protein phosphatases are either specific to phospho-Ser/-Thr, designated as PPT, or phospho-Tyr, designated as PTP, while phosphatase and tensin homologs (PTEN) are specific to phosphatidylinositides [90,126,130]. The reaction mechanisms for PPT and PTP during dephosphorylation are completely different, while there is a similarity between PTP and PTEN in that they both employ Cys as the catalytic amino acid.
Upon the activation of RTK, NADPH-oxidase embedded in plasma membranes is activated and produces O2−•/H2O2 via a one- or two-electron donation to molecular oxygen on the cell surface [17]. Although O2−• cannot permeate the plasma membrane due to its negative charge, the presence of the Cl− channel-3 (CIC-3) may allow it in some types of cells [131]. H2O2 is generally formed either directly by NADPH-oxidase or by superoxide dismutation outside cells after which it then enters the cells [16]. H2O2 reversibly oxidizes catalytic Cys-SH to Cys-OH, which enables the transient suppression of enzymatic activities of reactive PTP and PTEN upon growth factor stimuli [132,133]. PTP families that include PTP1B, Cdc25, SHP1, and SHP2 have been found to be sensitive targets for controlling this type of ROS signal. While the pKa value of the sulfhydryl of free Cys-SH or the conventional Cys-SH in proteins is approximately 8.5, the pKa value of Cys-SH in the catalytic center of PTP is generally maintained at less than 5.0 due to the microenvironment of the catalytic center. The resulting cysteine thiolate anion (Cys–S−) is highly susceptible to oxidation, and even H2O2 can oxidize Cys–S− to Cys–SOH and inactivate PTP. When H2O2 is eliminated by the action of peroxidases, such as peroxireodxins, Cys–SOH is reduced back to Cys–S− by redox molecules such as GSH. PTEN also has a catalytic Cys-SH and appears to be regulated by a similar mechanism [117,127]. However, there is concern that extracellularly produced H2O2 could diffuse into the extracellular space, and that not all of the H2O2 moves inside the cell. The disadvantage of extracellularly produced O2−•/H2O2 may be overcome by forming redoxosomes, which are endosomes that are constructed during RTK internalization [134]. The generation of hydrogen peroxide in the spherical redoxosome allows more hydrogen peroxide to reach the target PTP at the cytoplasmic side.
Concerning the action of 1O2, both the stimulation [18,19] and the inhibition [135] of cell growth have been reported. Mitochondrial respiration and ATP production in brain cells is stimulated by the 1O2 generated by a 1267 nm laser pulse [136]. 1O2 activates Akt via the PI3K pathway, which contributes to a survival response after UVA irradiation [18]. UVA also reportedly activates p38 MAPK in dermal fibroblasts through the activation of ligand-receptor signaling pathways or a ribotoxic stress mechanism [19]. This mitotic stimulation may be associated with the tumor growth activity of 1O2. In the meantime, however, 1O2 rapidly disrupts EGF receptor (EGFR)-mediated signaling by decreasing both the protein level and the level of phosphorylation of EGFR [137]. Because the inhibition of protein phosphatases with okadaic acid blocked the dephosphorylation of EGFR, ERK1/2 and Akt, responsible phosphatases appear to be activated by 1O2 under this condition. Thus, the action of 1O2 regarding this issue is complicated, and further studies will clearly be needed to elucidate the inconsistent observations concerning the action of 1O2 on mitotic signaling.
4.3. Apoptotic Cell Death Induced by 1O2
Although several pathways are involved in cell death, in this review, we mainly discuss this issue as it is related to apoptosis and ferroptosis. While apoptosis and necrosis are major classifications of cell death, they are now divided into several subclasses based on morphology and pathways [138]. Apoptosis is the most common type of cell death and is thought to account for more than 90% of physiologic cell death [139]. Apoptotic pathways can be roughly divided into two categories: an extrinsic pathway activated by death receptors and intrinsic pathways involving mitochondria. The extrinsic pathway involves death receptors, notably the Fas and TNFα receptors, and, upon stimulation, procaspase 8 is converted into active caspase 8, resulting in the activation of caspase 3 [140]. In the case of the intrinsic pathway, mitochondria are the first target of apoptotic stimuli, and cyt c is usually then released from mitochondria to the cytosol. While cyt c is a component in the electron transport system that is localized at the mitochondrial inner membrane and transfers an electron from ETC-III to ETC-IV (cytochrome c oxidase), it is released to the cytosol and also plays a key role in promoting apoptosis. The oxidized form of cyt c triggers the assembly of apoptosome that contains Apaf-1 and procaspase 9 as protein components [141,142] and leads to the activation of caspase 9 by autolysis. Procaspase 3 undergoes proteolytic activation by cascade 9, and the resulting active caspase 3 cleaves a variety of proteins, including an inhibitor of caspase-dependent DNase (ICAD). Thus, hydrolytic degradation occurs in proteins and DNA, but membrane structures consisting of phospholipid bilayer remain intact. Accordingly, cellular and nuclear body compaction occurs, but cellular components do not leak out of the apoptotic body in typical apoptosis. Under these conditions, the immune system virtually does not respond to apoptotic cells.
Studies employing photodynamic reactions and endoperoxides imply that apoptotic cell death is induced by 1O2, as judged by hallmarks including caspase activation and DNA cleavage [143,144,145]. While mitogen-activated protein kinases (MAPK) are originally named based on their roles in mediating mitotic signals [146], 1O2 and UV irradiation reportedly activates MAPK, p38 MAPK, c-Jun N-terminal kinase (JNK) and extracellular-signal regulated kinase (ERK) members, which eventually leads to the execution of apoptosis [13,147,148]. Because catalytic Cys in PTP is susceptible to 1O2, the inhibition of PTP may allow the phosphorylated Tyr in the upstream protein kinase to be sustained, leading to the preservation of the active state of downstream MAP kinases [21]. Although it is rather difficult to actually identify the ROS that are involved, one report suggested the differential involvement of 1O2 and O2−• in apoptosis pathways by means of cyclosporine A sensitivity to UVA1 (340–400 nm) irradiation [149]. In addition, 1O2 as well as H2O2 trigger the cleavage of Bid, which then releases cyt c from mitochondria [150]. This appears to be another mechanism of 1O2-induced apoptosis, which is independent from the action of the phosphatase/kinase system.
4.4. Ferroptosis Can Be an Alternate Cell Death Pathway by 1O2
About a decade ago, ferroptosis, which is exclusively caused by lipid peroxidation products, was characterized as an iron-dependent necrotic cell death [23,24]. Lipid radicals generated by the presence of ferrous iron are thought to enhance lipid peroxidation reactions and lead to membrane destruction [151]. Ferrous iron donates an electron to H2O2 which results in the formation of HO• by the Fenton reaction. The resulting HO• then reacts with polyunsaturated fatty acids to produce lipid peroxyl radicals, which initiate radical chain reaction and amplify the production of lipid peroxides [80]. Glutathione peroxidase (GPX4), a major suppressor of ferroptosis, reduces lipid peroxides to alcohols by transferring two electrons from GSH [152]. Different from apoptosis, ferroptosis stimulates the release of intracellular biological molecules, which may be responsible for aggravating inflammation or autoimmune responses under conditions of 1O2 production [153]. Inflammatory responses to necrotic cell death appear to be consistent with pathological conditions and damage caused by UV irradiation.
Despite the fact that many studies have appeared regarding apoptosis, our study in which MNPO2 was used as the 1O2 donor showed neither the activation of caspases nor DNA ladder formation during the cell death [53]. On the contrary, we found that 1O2 suppresses etoposide-induced apoptosis, although cell damage is further aggravated. 1O2 induces the release of cyt c, but it neither causes caspase activation nor the formation of apoptotic bodies. Cell death is alleviated in cells that overexpress GPX4 in mitochondria but not in cytoplasm. Treatment of the oxidized cyt c with MNPO2, in fact, completely inhibited its ability to induce caspase activation in an in vitro reconstitution assay. Because treatment with 1O2 resulted in the oxidation of Met, His and Trp in isolated cyt c [83], it had no effect on the oxidative state of heme [142]. These collective data imply that the oxidation of these amino acids in cyt c by 1O2 may counteract the induction of apoptosis through disabling interactions with Apaf-1.
Hela cells that had been treated with MNPO2 showed elevated hallmarks of ferroptosis, e.g., elevation in lipid peroxidation and free ferrous iron [25]. Ferrostatin-1, which specifically inhibits lipid peroxidation, partly suppresses cell death. The MNPO2 treatment had no effect on the status of Cys-GSH [25]. However, glutathione peroxidase and thioredoxin reductase, which are both selenocysteine-containing enzymes, have been reported to be prone to oxidation by 1O2 generated by photodynamic activation [154,155]. Several studies also have indicated that 1O2-induced lipid peroxidation is the cause of cell death, implying that ferroptosis is an alternate form of cell death by 1O2 [26,156].
Accordingly, it would be reasonable to assume that a mixture of apoptosis and ferroptosis is induced by 1O2 generated during PDT, as was implied [157]. The occurrence of this mixed type of cell death may be attributed to the properties of 1O2 as schematically depicted in Figure 5. Cyt c is anchored on the inner mitochondrial membrane via an interaction with cardiolipin, which contains four fatty acids, most of which are PUFA. Upon apoptotic stimuli, cyt c is released from mitochondria to the cytosol upon the preferable oxidation of cardiolipin [158]. The oxidized cyt c then assembles with other components to form apoptosomes, which activate caspases, thus resulting in the execution of apoptosis. It therefore appears that cells exposed to moderate levels of 1O2 may undergo apoptosis (Figure 5A).
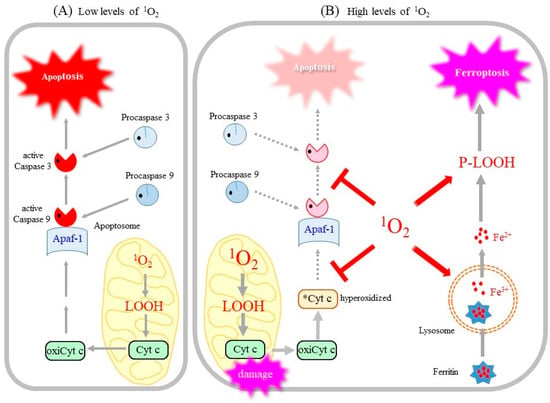
Figure 5.
1O2 dose differentially acts on cells and induces either apoptosis or ferroptosis. (A) A low dose of 1O2 peroxidizes PUFA, notably cardiolipin, results in the release of cyt c from mitochondria. Oxidized cyt c initiates the assembly of apoptosome and results in the activation of the caspase cascade, which leads to the eventual execution of apoptosis. (B) A high dose of 1O2 inactivates cyt c and caspase and aborts the apoptotic pathway. In the meantime, 1O2 causes lipid peroxidation, which destroys the lysosomal membrane and releases iron. Free iron along with P-LOOH accelerates membrane destruction and leads to the execution of ferroptosis. Mitochondrial damage and the accumulation of P-LOOH are also hallmarks of ferroptosis. oxiCyt c, oxidized, apoptosis-competent cyt c; *Cyt c, oxidized, apoptosis-incompetent cyt c.
However, when exposed to excess 1O2 (Figure 5B), cyt c and caspase, as cysteine proteases, undergo oxidative inactivation by 1O2, which aborts the apoptotic pathway [53,142]. Instead of apoptosis, the resulting LOOH and iron released from lysosomes may collectively activate the ferroptotic pathway. Thus, it is conceivable that the dose of 1O2 may determine if cells die by apoptosis or ferroptosis [22]. This hypothetical mechanism concerning 1O2-triggered cell death needs confirmation by further experimentation.
5. Concluding Remarks
1O2 is largely produced by photodynamic reactions and, hence, is a major factor in the photoaging of skin, while 1O2 generated during PDT is major component that exerts tumoricidal activity. Target molecules of 1O2 are somewhat limited compared to those of HO•, but oxidatively generated damage to multiple molecules and organelles can still occur. Although 1O2 is produced by many enzymatic and non-enzymatic reactions, the functional significance of generated 1O2 is poorly understood compared to other ROS. The application of donor compounds endoperoxides as well as newly developed sensor probes could contribute to our understanding of the action of 1O2 on cellular functions. Since there are contradictory observations on the action of 1O2 regarding cell growth and death, it will be necessary to clarify these issues.
Author Contributions
J.F. mainly wrote this manuscript, and Y.S. and Y.M. contributed to specific sections based on their specialty. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by the JSPS KAKENHI Grant-in-Aid for Scientific Research (C) (21K06850) to J.F. and the YU-COE program (S6) to J.F. from Yamagata University.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Fridovich, I. Superoxide radical and superoxide dismutases. Annu. Rev. Biochem. 1995, 64, 97–112. [Google Scholar] [CrossRef]
- Sies, H.; Berndt, C.; Jones, D.P. Oxidative Stress. Annu. Rev. Biochem. 2017, 86, 715–748. [Google Scholar] [CrossRef] [PubMed]
- Fujii, J.; Homma, T.; Osaki, T. Superoxide Radicals in the Execution of Cell Death. Antioxidants 2022, 11, 501. [Google Scholar] [CrossRef] [PubMed]
- Tang, Z.; Zhao, P.; Wang, H.; Liu, Y.; Bu, W. Biomedicine Meets Fenton Chemistry. Chem Rev. 2021, 121, 1981–2019. [Google Scholar] [CrossRef]
- Enami, S.; Sakamoto, Y.; Colussi, A.J. Fenton chemistry at aqueous interfaces. Proc. Natl. Acad. Sci. USA 2014, 111, 623–628. [Google Scholar] [CrossRef] [PubMed]
- Koppenol, W.H.; Hider, R.H. Iron and redox cycling. Do’s and don’ts. Free Radic. Biol. Med. 2019, 133, 3–10. [Google Scholar] [CrossRef]
- Petrou, A.L.; Terzidaki, A. A meta-analysis and review examining a possible role for oxidative stress and singlet oxygen in diverse diseases. Biochem. J. 2017, 474, 2713–2731. [Google Scholar] [CrossRef]
- Di Mascio, P.; Martinez, G.R.; Miyamoto, S.; Ronsein, G.E.; Medeiros, M.H.G.; Cadet, J. Singlet Molecular Oxygen Reactions with Nucleic Acids, Lipids, and Proteins. Chem. Rev. 2019, 119, 2043–2086. [Google Scholar] [CrossRef]
- Murotomi, K.; Umeno, A.; Shichiri, M.; Tanito, M.; Yoshida, Y. Significance of Singlet Oxygen Molecule in Pathologies. Int. J. Mol. Sci. 2023, 24, 2739. [Google Scholar] [CrossRef]
- Ahmad, N.; Mukhtar, H. Mechanism of photodynamic therapy-induced cell death. Methods Enzymol. 2000, 319, 342–358. [Google Scholar] [CrossRef]
- Chilakamarthi, U.; Giribabu, L. Photodynamic Therapy: Past, Present and Future. Chem. Rec. 2017, 17, 775–802. [Google Scholar] [CrossRef]
- Garcia-Diaz, M.; Huang, Y.Y.; Hamblin, M.R. Use of fluorescent probes for ROS to tease apart Type I and Type II photochemical pathways in photodynamic therapy. Methods 2016, 109, 158–166. [Google Scholar] [CrossRef] [PubMed]
- Pierlot, C.; Aubry, J.M.; Briviba, K.; Sies, H.; Di Mascio, P. Naphthalene endoperoxides as generators of singlet oxygen in biological media. Methods Enzymol. 2000, 319, 3–20. [Google Scholar] [CrossRef] [PubMed]
- Krutmann, J. Ultraviolet A radiation-induced biological effects in human skin: Relevance for photoaging and photodermatosis. J. Dermatol. Sci. 2000, 23 (Suppl. S1), S22–S26. [Google Scholar] [CrossRef]
- Tyrrell, R.M. Solar ultraviolet A radiation: An oxidizing skin carcinogen that activates heme oxygenase-1. Antioxid. Redox Signal. 2004, 6, 835–840. [Google Scholar] [CrossRef] [PubMed]
- Rhee, S.G. Cell signaling. H2O2, a necessary evil for cell signaling. Science 2006, 312, 1882–1883. [Google Scholar] [CrossRef]
- Finkel, T. Signal transduction by reactive oxygen species. J. Cell Biol. 2011, 194, 7–15. [Google Scholar] [CrossRef]
- Zhuang, S.; Kochevar, I.E. Singlet oxygen-induced activation of Akt/protein kinase B is independent of growth factor receptors. Photochem. Photobiol. 2003, 78, 361–371. [Google Scholar] [CrossRef]
- Le Panse, R.; Dubertret, L.; Coulomb, B. p38 mitogen-activated protein kinase activation by ultraviolet A radiation in human dermal fibroblasts. Photochem. Photobiol. 2003, 78, 168–174. [Google Scholar] [CrossRef]
- Morita, A.; Werfel, T.; Stege, H.; Ahrens, C.; Karmann, K.; Grewe, M.; Grether-Beck, S.; Ruzicka, T.; Kapp, A.; Klotz, L.O.; et al. Evidence that singlet oxygen-induced human T helper cell apoptosis is the basic mechanism of ultraviolet-A radiation phototherapy. J. Exp. Med. 1997, 186, 1763–1768. [Google Scholar] [CrossRef]
- Klotz, L.O.; Kröncke, K.D.; Sies, H. Singlet oxygen-induced signaling effects in mammalian cells. Photochem. Photobiol. Sci. 2003, 2, 88–94. [Google Scholar] [CrossRef]
- Novikova, I.N.; Potapova, E.V.; Dremin, V.V.; Dunaev, A.V.; Abramov, A.Y. Laser-induced singlet oxygen selectively triggers oscillatory mitochondrial permeability transition and apoptosis in melanoma cell lines. Life Sci. 2022, 304, 120720. [Google Scholar] [CrossRef]
- Dixon, S.J.; Lemberg, K.M.; Lamprecht, M.R.; Skouta, R.; Zaitsev, E.M.; Gleason, C.E.; Patel, D.N.; Bauer, A.J.; Cantley, A.M.; Yang, W.S.; et al. Ferroptosis: An iron-dependent form of nonapoptotic cell death. Cell 2012, 149, 1060–1072. [Google Scholar] [CrossRef] [PubMed]
- Stockwell, B.R. Ferroptosis turns 10: Emerging mechanisms, physiological functions, and therapeutic applications. Cell 2022, 185, 2401–2421. [Google Scholar] [CrossRef] [PubMed]
- Homma, T.; Kobayashi, S.; Fujii, J. Induction of ferroptosis by singlet oxygen generated from naphthalene endoperoxide. Biochem. Biophys. Res. Commun. 2019, 518, 519–525. [Google Scholar] [CrossRef] [PubMed]
- Meng, X.; Deng, J.; Liu, F.; Guo, T.; Liu, M.; Dai, P.; Fan, A.; Wang, Z.; Zhao, Y. Triggered All-Active Metal Organic Framework: Ferroptosis Machinery Contributes to the Apoptotic Photodynamic Antitumor Therapy. Nano Lett. 2019, 19, 7866–7876. [Google Scholar] [CrossRef] [PubMed]
- Maharjan, P.S.; Bhattarai, H.K. Singlet Oxygen, Photodynamic Therapy, and Mechanisms of Cancer Cell Death. J. Oncol. 2022, 2022, 7211485. [Google Scholar] [CrossRef]
- Wefers, H.; Sies, H. Oxidation of glutathione by the superoxide radical to the disulfide and the sulfonate yielding singlet oxygen. Eur. J. Biochem. 1983, 137, 29–36. [Google Scholar] [CrossRef]
- Di Mascio, P.; Briviba, K.; Sasaki, S.T.; Catalani, L.H.; Medeiros, M.H.; Bechara, E.J.; Sies, H. The reaction of peroxynitrite with tert-butyl hydroperoxide produces singlet molecular oxygen. Biol. Chem. 1997, 378, 1071–1074. [Google Scholar]
- Miyamoto, S.; Martinez, G.R.; Medeiros, M.H.; Di Mascio, P. Singlet molecular oxygen generated by biological hydroperoxides. J. Photochem. Photobiol. B. 2014, 139, 24–33. [Google Scholar] [CrossRef]
- Redmond, R.W.; Kochevar, I.E. Spatially resolved cellular responses to singlet oxygen. Photochem. Photobiol. 2006, 82, 1178–1186. [Google Scholar] [CrossRef] [PubMed]
- Jiménez-Banzo, A.; Sagristà, M.L.; Mora, M.; Nonell, S. Kinetics of singlet oxygen photosensitization in human skin fibroblasts. Free Radic. Biol. Med. 2008, 44, 1926–1934. [Google Scholar] [CrossRef] [PubMed]
- Driever, S.M.; Fryer, M.J.; Mullineaux, P.M.; Baker, N.R. Imaging of reactive oxygen species in vivo. Methods Mol. Biol. 2009, 479, 109–116. [Google Scholar] [CrossRef]
- Pedersen, S.K.; Holmehave, J.; Blaikie, F.H.; Gollmer, A.; Breitenbach, T.; Jensen, H.H.; Ogilby, P.R. Aarhus sensor green: A fluorescent probe for singlet oxygen. J. Org. Chem. 2014, 79, 3079–3087. [Google Scholar] [CrossRef]
- Tang, C.Y.; Wu, F.Y.; Yang, M.K.; Guo, Y.M.; Lu, G.H.; Yang, Y.H. A Classic Near-Infrared Probe Indocyanine Green for Detecting Singlet Oxygen. Int. J. Mol. Sci. 2016, 17, 219. [Google Scholar] [CrossRef]
- Ruiz-González, R.; Bresolí-Obach, R.; Gulías, Ò.; Agut, M.; Savoie, H.; Boyle, R.W.; Nonell, S.; Giuntini, F. NanoSOSG: A Nanostructured Fluorescent Probe for the Detection of Intracellular Singlet Oxygen. Angew Chem. Int. Ed. Engl. 2017, 56, 2885–2888. [Google Scholar] [CrossRef] [PubMed]
- Chen, B.; Yang, Y.; Wang, Y.; Yan, Y.; Wang, Z.; Yin, Q.; Zhang, Q.; Wang, Y. ACS Precise Monitoring of Singlet Oxygen in Specific Endocytic Organelles by Super-pH-Resolved Nanosensors. Appl. Mater. Interfaces 2021, 13, 18533–18544. [Google Scholar] [CrossRef]
- Nath, P.; Hamadna, S.S.; Karamchand, L.; Foster, J.; Kopelman, R.; Amar, J.G.; Ray, A. Intracellular detection of singlet oxygen using fluorescent nanosensors. Analyst 2021, 146, 3933–3941. [Google Scholar] [CrossRef]
- Kim, S.; Tachikawa, T.; Fujitsuka, M.; Majima, T. Far-red fluorescence probe for monitoring singlet oxygen during photodynamic therapy. J. Am. Chem. Soc. 2014, 136, 11707–11715. [Google Scholar] [CrossRef] [PubMed]
- Murotomi, K.; Umeno, A.; Sugino, S.; Yoshida, Y. Quantitative kinetics of intracellular singlet oxygen generation using a fluorescence probe. Sci. Rep. 2020, 10, 10616. [Google Scholar] [CrossRef]
- Zhao, H.; Takano, Y.; Sasikumar, D.; Miyatake, Y.; Biju, V. Excitation-Wavelength-Dependent Functionalities of Temporally Controlled Sensing and Generation of Singlet Oxygen by a Photoexcited State Engineered Rhodamine 6G-Anthracene Conjugate. Chemistry 2022, 28, e202202014. [Google Scholar] [CrossRef]
- Sasikumar, D.; Takano, Y.; Zhao, H.; Kohara, R.; Hamada, M.; Kobori, Y.; Biju, V. Caging and photo-triggered uncaging of singlet oxygen by excited state engineering of electron donor-acceptor-linked molecular sensors. Sci. Rep. 2022, 12, 11371. [Google Scholar] [CrossRef]
- Baptista, M.S.; Cadet, J.; Di Mascio, P.; Ghogare, A.A.; Greer, A.; Hamblin, M.R.; Lorente, C.; Nunez, S.C.; Ribeiro, M.S.; Thomas, A.H.; et al. Type I and Type II Photosensitized Oxidation Reactions: Guidelines and Mechanistic Pathways. Photochem. Photobiol. 2017, 93, 912–919. [Google Scholar] [CrossRef]
- Baptistal, M.S.; Cadet, J.; Greer, A.; Thomas, A.H. Photosensitization Reactions of Biomolecules: Definition, Targets and Mechanisms. Photochem. Photobiol. 2021, 97, 1456–1483. [Google Scholar] [CrossRef] [PubMed]
- Girotti, A.W.; Kriska, T. Role of lipid hydroperoxides in photo-oxidative stress signaling. Antioxid. Redox Signal. 2004, 6, 301–310. [Google Scholar] [CrossRef] [PubMed]
- Reis, A.; Spickett, C.M. Chemistry of phospholipid oxidation. Biochim. Biophys. Acta 2012, 1818, 2374–2387. [Google Scholar] [CrossRef] [PubMed]
- Kochevar, I.E.; Redmond, R.W. Photosensitized production of singlet oxygen. Methods Enzymol. 2000, 319, 20–28. [Google Scholar] [CrossRef]
- Jablonski, A. Efficiency of anti-stokes fluorescence in dyes. Nature 1933, 131, 839–840. [Google Scholar] [CrossRef]
- Aziz, B.; Aziz, I.; Khurshid, A.; Raoufi, E.; Esfahani, F.N.; Jalilian, Z.; Mozafari, M.R.; Taghavi, E.; Ikram, M. An Overview of Potential Natural Photosensitizers in Cancer Photodynamic Therapy. Biomedicines 2023, 11, 224. [Google Scholar] [CrossRef]
- Wei, L.; Zhang, Z.; Kumar, A.; Banerjee, S.; Huang, H. Endoperoxides Compounds for Highly Efficient Cancer Treatment under Hypoxia. Chemistry 2022, 28, e202202233. [Google Scholar] [CrossRef] [PubMed]
- Ravanat, J.L.; Di Mascio, P.; Martinez, G.R.; Medeiros, M.H.; Cadet, J. Singlet oxygen induces oxidation of cellular DNA. J. Biol. Chem. 2000, 275, 40601–40604. [Google Scholar] [CrossRef]
- Ravanat, J.L.; Douki, T.; Duez, P.; Gremaud, E.; Herbert, K.; Hofer, T.; Lasserre, L.; Saint-Pierre, C.; Favier, A.; Cadet, J. Cellular background level of 8-oxo-7,8-dihydro-2’-deoxyguanosine: An isotope based method to evaluate artefactual oxidation of DNA during its extraction and subsequent work-up. Carcinogenesis 2002, 23, 1911–1918. [Google Scholar] [CrossRef]
- Otsu, K.; Sato, K.; Ikeda, Y.; Imai, H.; Nakagawa, Y.; Ohba, Y.; Fujii, J. An abortive apoptotic pathway induced by singlet oxygen is due to the suppression of caspase activation. Biochem. J. 2005, 389, 197–206. [Google Scholar] [CrossRef] [PubMed]
- Lu, F.; Pan, L.; Wu, T.; Pan, W.; Gao, W.; Li, N.; Tang, B. An endoperoxide-containing covalent organic framework as a singlet oxygen reservoir for cancer therapy. Chem. Commun. 2022, 58, 11013–11016. [Google Scholar] [CrossRef]
- Gülçin, İ. Antioxidant activity of food constituents: An overview. Arch. Toxicol. 2012, 86, 345–391. [Google Scholar] [CrossRef]
- Ou, B.; Hampsch-Woodill, M.; Prior, R.L. Development and validation of an improved oxygen radical absorbance capacity assay using fluorescein as the fluorescent probe. J. Agric. Food Chem. 2001, 49, 4619–4626. [Google Scholar] [CrossRef] [PubMed]
- Ouchi, A.; Aizawa, K.; Iwasaki, Y.; Inakuma, T.; Terao, J.; Nagaoka, S.; Mukai, K. Kinetic study of the quenching reaction of singlet oxygen by carotenoids and food extracts in solution. Development of a singlet oxygen absorption capacity (SOAC) assay method. J. Agric. Food Chem. 2010, 58, 9967–9978. [Google Scholar] [CrossRef] [PubMed]
- Foote, C.S.; Chang, Y.C.; Denny, R.W. Chemistry of singlet oxygen. X. Carotenoid quenching parallels biological protection. J. Am. Chem. Soc. 1970, 92, 5216–5218. [Google Scholar] [CrossRef]
- Edge, R.; Truscott, T.G. Singlet Oxygen and Free Radical Reactions of Retinoids and Carotenoids-A Review. Antioxidants 2018, 7, 5. [Google Scholar] [CrossRef]
- Di Mascio, P.; Kaiser, S.; Sies, H. Lycopene as the most efficient biological carotenoid singlet oxygen quencher. Arch. Biochem. Biophys. 1989, 274, 532–538. [Google Scholar] [CrossRef]
- Garmyn, M.; Ribaya-Mercado, J.D.; Russel, R.M.; Bhawan, J.; Gilchrest, B.A. Effect of beta-carotene supplementation on the human sunburn reaction. Exp. Dermatol. 1995, 4, 104–111. [Google Scholar] [CrossRef] [PubMed]
- Stahl, W.; Sies, H. Carotenoids and protection against solar UV radiation. Skin Pharmacol. Appl. Skin Physiol. 2002, 15, 291–296. [Google Scholar] [CrossRef] [PubMed]
- Sies, H.; Stahl, W. Carotenoids and UV protection. Photochem. Photobiol. Sci. 2004, 3, 749–752. [Google Scholar] [CrossRef]
- Ozkan, G.; Günal-Köroğlu, D.; Karadag, A.; Capanoglu, E.; Cardoso, S.M.; Al-Omari, B.; Calina, D.; Sharifi-Rad, J.; Cho, W.C. A mechanistic updated overview on lycopene as potential anticancer agent. Biomed. Pharmacother. 2023, 161, 114428. [Google Scholar] [CrossRef] [PubMed]
- Ahn, Y.J.; Kim, H. Lutein as a Modulator of Oxidative Stress-Mediated Inflammatory Diseases. Antioxidants 2021, 10, 1448. [Google Scholar] [CrossRef]
- Sommani, P.; Arai, T.; Yamashita, K.; Miyoshi, T.; Mori, H.; Sasada, M.; Makino, K. Effects of edaravone on singlet oxygen released from activated human neutrophils. J. Pharmacol. Sci. 2007, 103, 117–120. [Google Scholar] [CrossRef] [PubMed]
- Nishinaka, Y.; Mori, H.; Endo, N.; Miyoshi, T.; Yamashita, K.; Adachi, S.; Arai, T. Edaravone directly reacts with singlet oxygen and protects cells from attack. Life Sci. 2010, 86, 808–813. [Google Scholar] [CrossRef]
- Morita, M.; Naito, Y.; Yoshikawa, T.; Niki, E. Inhibition of plasma lipid oxidation induced by peroxyl radicals, peroxynitrite, hypochlorite, 15-lipoxygenase, and singlet oxygen by clinical drugs. Bioorg. Med. Chem. Lett. 2016, 26, 5411–5417. [Google Scholar] [CrossRef]
- Girotti, A.W.; Korytowski, W. Cholesterol Hydroperoxide Generation, Translocation, and Reductive Turnover in Biological Systems. Cell Biochem. Biophys. 2017, 75, 413–419. [Google Scholar] [CrossRef]
- Zerbinati, C.; Iuliano, L. Cholesterol and related sterols autoxidation. Free Radic. Biol. Med. 2017, 111, 151–155. [Google Scholar] [CrossRef] [PubMed]
- Miyamoto, S.; Lima, R.S.; Inague, A.; Viviani, L.G. Electrophilic oxysterols: Generation, measurement and protein modification. Free Radic. Res. 2021, 55, 416–440. [Google Scholar] [CrossRef]
- Korytowski, W.; Girotti, A.W. Singlet oxygen adducts of cholesterol: Photogeneration and reductive turnover in membrane systems. Photochem. Photobiol. 1999, 70, 484–489. [Google Scholar] [CrossRef]
- Girotti, A.W.; Korytowski, W. Cholesterol Peroxidation as a Special Type of Lipid Oxidation in Photodynamic Systems. Photochem. Photobiol. 2019, 95, 73–82. [Google Scholar] [CrossRef] [PubMed]
- Miyamoto, S.; Martinez, G.R.; Rettori, D.; Augusto, O.; Medeiros, M.H.; Di Mascio, P. Linoleic acid hydroperoxide reacts with hypochlorous acid, generating peroxyl radical intermediates and singlet molecular oxygen. Proc. Natl. Acad. Sci. USA 2006, 103, 293–298. [Google Scholar] [CrossRef] [PubMed]
- Uchida, K. 4-Hydroxy-2-nonenal: A product and mediator of oxidative stress. Prog. Lipid Res. 2003, 42, 318–343. [Google Scholar] [CrossRef] [PubMed]
- Jaganjac, M.; Milkovic, L.; Gegotek, A.; Cindric, M.; Zarkovic, K.; Skrzydlewska, E.; Zarkovic, N. The relevance of pathophysiological alterations in redox signaling of 4-hydroxynonenal for pharmacological therapies of major stress-associated diseases. Free Radic. Biol. Med. 2020, 157, 128–153. [Google Scholar] [CrossRef]
- Tsubone, T.M.; Baptista, M.S.; Itri, R. Understanding membrane remodelling initiated by photosensitized lipid oxidation. Biophys. Chem. 2019, 254, 106263. [Google Scholar] [CrossRef] [PubMed]
- Petrillo, S.; Pietrafusa, N.; Trivisano, M.; Calabrese, C.; Saura, F.; Gallo, M.G.; Bertini, E.S.; Vigevano, F.; Specchio, N.; Piemonte, F. Imbalance of Systemic Redox Biomarkers in Children with Epilepsy: Role of Ferroptosis. Antioxidants 2021, 10, 1267. [Google Scholar] [CrossRef]
- Zhang, Q.; Li, N.; Deng, L.; Jiang, X.; Zhang, Y.; Lee, L.T.O.; Zhang, H. ACSL1-induced ferroptosis and platinum resistance in ovarian cancer by increasing FSP1 N-myristylation and stability. Cell Death Discov. 2023, 9, 83. [Google Scholar] [CrossRef]
- Niki, E. Lipid peroxidation: Physiological levels and dual biological effects. Free Radic. Biol. Med. 2009, 47, 469–484. [Google Scholar] [CrossRef]
- Girotti, A.W.; Korytowski, W. Nitric Oxide Inhibition of Chain Lipid Peroxidation Initiated by Photodynamic Action in Membrane Environments. Cell Biochem. Biophys. 2020, 78, 149–156. [Google Scholar] [CrossRef]
- Girotti, A.W.; Fahey, J.M.; Korbelik, M. Photodynamic Therapy as an Oxidative Anti-Tumor Modality: Negative Effects of Nitric Oxide on Treatment Efficacy. Pharmaceutics 2021, 13, 593. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.; Rodriguez, M.E.; Guo, M.; Kenney, M.E.; Oleinick, N.L.; Anderson, V.E. Oxidative modification of cytochrome c by singlet oxygen. Free Radic. Biol. Med. 2008, 44, 1700–1711. [Google Scholar] [CrossRef] [PubMed]
- Marques, E.F.; Medeiros, M.H.G.; Di Mascio, P. Lysozyme oxidation by singlet molecular oxygen: Peptide characterization using [18O]-labeling oxygen and nLC-MS/MS. J. Mass Spectrom. 2017, 52, 739–751. [Google Scholar] [CrossRef]
- Fujii, J.; Ito, J.I.; Zhang, X.; Kurahashi, T. Unveiling the roles of the glutathione redox system in vivo by analyzing genetically modified mice. J. Clin. Biochem. Nutr. 2011, 49, 70–78. [Google Scholar] [CrossRef]
- Turk, B.; Turk, V.; Turk, D. Structural and functional aspects of papain-like cysteine proteinases and their protein inhibitors. Biol. Chem. 1997, 378, 141–150. [Google Scholar]
- Yamamoto, K. Cathepsin E and cathepsin D: Biosynthesis, processing and subcellular location. Adv. Exp. Med. Biol. 1995, 362, 223–229. [Google Scholar] [CrossRef] [PubMed]
- Nagaoka, Y.; Otsu, K.; Okada, F.; Sato, K.; Ohba, Y.; Kotani, N.; Fujii, J. Specific inactivation of cysteine protease-type cathepsin by singlet oxygen generated from naphthalene endoperoxides. Biochem. Biophys. Res. Commun. 2005, 331, 215–223. [Google Scholar] [CrossRef]
- Suto, D.; Iuchi, Y.; Ikeda, Y.; Sato, K.; Ohba, Y.; Fujii, J. Inactivation of cysteine and serine proteases by singlet oxygen. Arch. Biochem. Biophys. 2007, 461, 151–158. [Google Scholar] [CrossRef]
- Netto, L.E.S.; Machado, L.E.S.F. Preferential redox regulation of cysteine-based protein tyrosine phosphatases: Structural and biochemical diversity. FEBS J. 2022, 289, 5480–5504. [Google Scholar] [CrossRef]
- von Montfort, C.; Sharov, V.S.; Metzger, S.; Schöneich, C.; Sies, H.; Klotz, L.O. Singlet oxygen inactivates protein tyrosine phosphatase-1B by oxidation of the active site cysteine. Biol. Chem. 2006, 387, 1399–1404. [Google Scholar] [CrossRef] [PubMed]
- Brambullo, T.; Colonna, M.R.; Vindigni, V.; Piaserico, S.; Masciopinto, G.; Galeano, M.; Costa, A.L.; Bassetto, F. Xeroderma Pigmentosum: A Genetic Condition Skin Cancer Correlated-A Systematic Review. Biomed. Res. Int. 2022, 2022, 8549532. [Google Scholar] [CrossRef] [PubMed]
- Brem, R.; Guven, M.; Karran, P. Oxidatively-generated damage to DNA and proteins mediated by photosensitized UVA. Free Radic. Biol. Med. 2017, 107, 101–109. [Google Scholar] [CrossRef]
- Mullenders, L.H.F. Solar UV damage to cellular DNA: From mechanisms to biological effects. Photochem. Photobiol. Sci. 2018, 17, 1842–1852. [Google Scholar] [CrossRef] [PubMed]
- Halliwell, B. Oxidative stress and cancer: Have we moved forward? Biochem. J. 2007, 401, 1–11. [Google Scholar] [CrossRef]
- Cadet, J.; Douki, T.; Ravanat, J.L. Oxidatively generated damage to cellular DNA by UVB and UVA radiation. Photochem. Photobiol. 2015, 91, 140–155. [Google Scholar] [CrossRef] [PubMed]
- Cadet, J.; Douki, T. Formation of UV-induced DNA damage contributing to skin cancer development. Photochem. Photobiol. Sci. 2018, 17, 1816–1841. [Google Scholar] [CrossRef]
- Pouget, J.P.; Douki, T.; Richard, M.J.; Cadet, J. DNA damage induced in cells by gamma and UVA radiation as measured by HPLC/GC-MS and HPLC-EC and Comet assay. Chem. Res. Toxicol. 2000, 13, 541–549. [Google Scholar] [CrossRef]
- Piette, J. Biological consequences associated with DNA oxidation mediated by singlet oxygen. J. Photochem. Photobiol. B 1991, 11, 241–260. [Google Scholar] [CrossRef]
- Petrat, F.; Pindiur, S.; Kirsch, M.; de Groot, H. NAD(P)H, a primary target of 1O2 in mitochondria of intact cells. J. Biol. Chem. 2003, 278, 3298–3307. [Google Scholar] [CrossRef]
- Ahmad, R.; Ahsan, H. Singlet oxygen species and systemic lupus erythematosus: A brief review. J. Immunoass. Immunochem. 2019, 40, 343–349. [Google Scholar] [CrossRef] [PubMed]
- Khan, F.; Khan, F.; Ali, R. Immunogenicity of DNA modified by singlet oxygen: Implications in systemic lupus erythematosus and cancer. Biotechnol. Appl. Biochem. 2007, 46, 97–103. [Google Scholar] [CrossRef] [PubMed]
- Maitra, D.; Bragazzi Cunha, J.; Elenbaas, J.S.; Bonkovsky, H.L.; Shavit, J.A.; Omary, M.B. Porphyrin-Induced Protein Oxidation and Aggregation as a Mechanism of Porphyria-Associated Cell Injury. Cell Mol. Gastroenterol. Hepatol. 2019, 8, 535–548. [Google Scholar] [CrossRef]
- da Veiga Moreira, J.; Schwartz, L.; Jolicoeur, M. Targeting Mitochondrial Singlet Oxygen Dynamics Offers New Perspectives for Effective Metabolic Therapies of Cancer. Front. Oncol. 2020, 10, 573399. [Google Scholar] [CrossRef] [PubMed]
- Berneburg, M.; Grether-Beck, S.; Kürten, V.; Ruzicka, T.; Briviba, K.; Sies, H.; Krutmann, J. Singlet oxygen mediates the UVA-induced generation of the photoaging-associated mitochondrial common deletion. J. Biol. Chem. 1999, 274, 15345–15349. [Google Scholar] [CrossRef]
- Moreno, G.; Poussin, K.; Ricchelli, F.; Salet, C. The effects of singlet oxygen produced by photodynamic action on the mitochondrial permeability transition differ in accordance with the localization of the sensitizer. Arch. Biochem. Biophys. 2001, 386, 243–250. [Google Scholar] [CrossRef]
- Xue, L.Y.; Chiu, S.M.; Oleinick, N.L. Photochemical destruction of the Bcl-2 oncoprotein during photodynamic therapy with the phthalocyanine photosensitizer Pc 4. Oncogene 2001, 20, 3420–3427. [Google Scholar] [CrossRef]
- Hill, H.Z.; Li, W.; Xin, P.; Mitchell, D.L. Melanin: A two edged sword? Pigment Cell Res. 1997, 10, 158–161. [Google Scholar] [CrossRef]
- Selbo, P.K.; Høgset, A.; Prasmickaite, L.; Berg, K. Photochemical internalisation: A novel drug delivery system. Tumour Biol. 2002, 23, 103–112. [Google Scholar] [CrossRef]
- Reiners, J.J., Jr.; Agostinis, P.; Berg, K.; Oleinick, N.L.; Kessel, D. Assessing autophagy in the context of photodynamic therapy. Autophagy 2010, 6, 7–18. [Google Scholar] [CrossRef]
- Sreedhar, A.; Aguilera-Aguirre, L.; Singh, K.K. Mitochondria in skin health, aging, and disease. Cell Death Dis. 2020, 11, 444. [Google Scholar] [CrossRef]
- Kessel, D.; Reiners, J.J. Photodynamic therapy: Autophagy and mitophagy, apoptosis and paraptosis. Autophagy 2020, 16, 2098–2101. [Google Scholar] [CrossRef]
- Rizzollo, F.; More, S.; Vangheluwe, P.; Agostinis, P. The lysosome as a master regulator of iron metabolism. Trends Biochem. Sci. 2021, 46, 960–975. [Google Scholar] [CrossRef]
- Martins, W.K.; Santos, N.F.; Rocha, C.S.; Bacellar, I.O.L.; Tsubone, T.M.; Viotto, A.C.; Matsukuma, A.Y.; Abrantes, A.B.P.; Siani, P.; Dias, L.G.; et al. Parallel damage in mitochondria and lysosomes is an efficient way to photoinduce cell death. Autophagy 2019, 15, 259–279. [Google Scholar] [CrossRef]
- Tonolli, P.N.; Martins, W.K.; Junqueira, H.C.; Silva, M.N.; Severino, D.; Santacruz-Perez, C.; Watanabe, I.; Baptista, M.S. Lipofuscin in keratinocytes: Production, properties, and consequences of the photosensitization with visible light. Free Radic. Biol. Med. 2020, 160, 277–292. [Google Scholar] [CrossRef]
- Zhang, Y.; Zhou, Q.; Bu, Y.; Xu, T.; Zhu, X.; Zhang, J.; Yu, Z.; Wang, L.; Zhong, F.; Zhou, H. Real-time imaging mitochondrial viscosity dynamic during mitophagy mediated by photodynamic therapy. Anal. Chim. Acta 2021, 1178, 338847. [Google Scholar] [CrossRef] [PubMed]
- Ding, S.; Yang, M.; Lv, J.; Li, H.; Wei, G.; Gao, J.; Yuan, Z. Novel Lysosome-Targeting Fluorescence off-on Photosensitizer for Near-Infrared Hypoxia Imaging and Photodynamic Therapy in vitro and in vivo. Molecules 2022, 27, 3457. [Google Scholar] [CrossRef] [PubMed]
- Pellieux, C.; Dewilde, A.; Pierlot, C.; Aubry, J.M. Bactericidal and virucidal activities of singlet oxygen generated by thermolysis of naphthalene endoperoxides. Methods Enzymol. 2000, 319, 197–207. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Qin, R.; Zaat, S.A.J.; Breukink, E.; Heger, M. Antibacterial photodynamic therapy: Overview of a promising approach to fight antibiotic-resistant bacterial infections. J. Clin. Transl. Res. 2015, 1, 140–167. [Google Scholar] [PubMed]
- Mariewskaya, K.A.; Tyurin, A.P.; Chistov, A.A.; Korshun, V.A.; Alferova, V.A.; Ustinov, A.V. Photosensitizing Antivirals. Molecules 2021, 26, 3971. [Google Scholar] [CrossRef]
- Mishchenko, T.; Balalaeva, I.; Gorokhova, A.; Vedunova, M.; Krysko, D.V. Which cell death modality wins the contest for photodynamic therapy of cancer? Cell Death Dis. 2022, 13, 455. [Google Scholar] [CrossRef]
- Nishinaka, Y.; Arai, T.; Adachi, S.; Takaori-Kondo, A.; Yamashita, K. Singlet oxygen is essential for neutrophil extracellular trap formation. Biochem. Biophys. Res. Commun. 2011, 413, 75–79. [Google Scholar] [CrossRef] [PubMed]
- Tatsuzawa, H.; Maruyama, T.; Misawa, N.; Fujimori, K.; Hori, K.; Sano, Y.; Kambayashi, Y.; Nakano, M. Inactivation of bacterial respiratory chain enzymes by singlet oxygen. FEBS Lett. 1998, 439, 329–333. [Google Scholar] [CrossRef] [PubMed]
- Wagner, S.J. Virus inactivation in blood components by photoactive phenothiazine dyes. Transfus. Med. Rev. 2002, 16, 61–66. [Google Scholar] [CrossRef]
- Mariewskaya, K.A.; Krasilnikov, M.S.; Korshun, V.A.; Ustinov, A.V.; Alferova, V.A. Near-Infrared Dyes: Towards Broad-Spectrum Antivirals. Int. J. Mol. Sci. 2022, 24, 188. [Google Scholar] [CrossRef]
- Wurzenberger, C.; Gerlich, D.W. Phosphatases: Providing safe passage through mitotic exit. Nat. Rev. Mol. Cell Biol. 2011, 12, 469–482. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.R.; Kwon, K.S.; Kim, S.R.; Rhee, S.G. Reversible inactivation of protein-tyrosine phosphatase 1B in A431 cells stimulated with epidermal growth factor. J. Biol. Chem. 1998, 273, 15366–15372. [Google Scholar] [CrossRef] [PubMed]
- Sebolt-Leopold, J.S.; Herrera, R. Targeting the mitogen-activated protein kinase cascade to treat cancer. Nat. Rev. Cancer 2004, 4, 937–947. [Google Scholar] [CrossRef] [PubMed]
- Maehama, T.; Taylor, G.S.; Dixon, J.E. PTEN and myotubularin: Novel phosphoinositide phosphatases. Annu. Rev. Biochem. 2001, 70, 247–279. [Google Scholar] [CrossRef] [PubMed]
- Shillingford, S.R.; Bennett, A.M. Mitogen-Activated Protein Kinase Phosphatases: No Longer Undruggable? Annu. Rev. Pharmacol. Toxicol. 2023, 63, 617–636. [Google Scholar] [CrossRef]
- Fisher, A.B. Redox signaling across cell membranes. Antioxid. Redox Signal. 2009, 11, 1349–1356. [Google Scholar] [CrossRef] [PubMed]
- Cho, S.H.; Lee, C.H.; Ahn, Y.; Kim, H.; Kim, H.; Ahn, C.Y.; Yang, K.S.; Lee, S.R. Redox regulation of PTEN and protein tyrosine phosphatases in H2O2 mediated cell signaling. FEBS Lett. 2004, 560, 7–13. [Google Scholar] [CrossRef]
- Böhmer, F.; Szedlacsek, S.; Tabernero, L.; Ostman, A.; den Hertog, J. Protein tyrosine phosphatase structure-function relationships in regulation and pathogenesis. FEBS J. 2013, 280, 413–431. [Google Scholar] [CrossRef] [PubMed]
- Tsutsumi, R.; Harizanova, J.; Stockert, R.; Schröder, K.; Bastiaens, P.I.H.; Neel, B.G. Assay to visualize specific protein oxidation reveals spatio-temporal regulation of SHP2. Nat. Commun. 2017, 8, 466. [Google Scholar] [CrossRef]
- Schieke, S.M.; von Montfort, C.; Buchczyk, D.P.; Timmer, A.; Grether-Beck, S.; Krutmann, J.; Holbrook, N.J.; Klotz, L.O. Singlet oxygen-induced attenuation of growth factor signaling: Possible role of ceramides. Free Radic. Res. 2004, 38, 729–737. [Google Scholar] [CrossRef]
- Sokolovski, S.G.; Rafailov, E.U.; Abramov, A.Y.; Angelova, P.R. Singlet oxygen stimulates mitochondrial bioenergetics in brain cells. Free Radic. Biol. Med. 2021, 163, 306–313. [Google Scholar] [CrossRef]
- Zhuang, S.; Ouedraogo, G.D.; Kochevar, I.E. Downregulation of epidermal growth factor receptor signaling by singlet oxygen through activation of caspase-3 and protein phosphatases. Oncogene 2003, 22, 4413–4424. [Google Scholar] [CrossRef]
- Tang, D.; Kang, R.; Berghe, T.V.; Vandenabeele, P.; Kroemer, G. The molecular machinery of regulated cell death. Cell Res. 2019, 29, 347–364. [Google Scholar] [CrossRef] [PubMed]
- Fuchs, Y.; Steller, H. Programmed cell death in animal development and disease. Cell 2011, 147, 742–758. [Google Scholar] [CrossRef] [PubMed]
- Nagata, S. Apoptosis and Clearance of Apoptotic Cells. Annu. Rev. Immunol. 2018, 36, 489–517. [Google Scholar] [CrossRef]
- Hampton, M.B.; Zhivotovsky, B.; Slater, A.F.; Burgess, D.H.; Orrenius, S. Importance of the redox state of cytochrome c during caspase activation in cytosolic extracts. Biochem. J. 1998, 329, 95–99. [Google Scholar] [CrossRef]
- Suto, D.; Sato, K.; Ohba, Y.; Yoshimura, T.; Fujii, J. Suppression of the pro-apoptotic function of cytochrome c by singlet oxygen via a haem redox state-independent mechanism. Biochem. J. 2005, 392, 399–406. [Google Scholar] [CrossRef] [PubMed]
- Granville, D.J.; Carthy, C.M.; Jiang, H.; Shore, G.C.; McManus, B.M.; Hunt, D.W. Rapid cytochrome c release, activation of caspases 3, 6, 7 and 8 followed by Bap31 cleavage in HeLa cells treated with photodynamic therapy. FEBS Lett. 1998, 437, 5–10. [Google Scholar] [CrossRef]
- Kochevar, I.E.; Lynch, M.C.; Zhuang, S.; Lambert, C.R. Singlet oxygen, but not oxidizing radicals, induces apoptosis in HL-60 cells. Photochem. Photobiol. 2000, 72, 548–553. [Google Scholar] [CrossRef] [PubMed]
- Valencia, A.; Morán, J. Reactive oxygen species induce different cell death mechanisms in cultured neurons. Free Radic. Biol. Med. 2004, 36, 1112–1125. [Google Scholar] [CrossRef] [PubMed]
- Gallo, K.A.; Johnson, G.L. Mixed-lineage kinase control of JNK and p38 MAPK pathways. Nat. Rev. Mol. Cell Biol. 2002, 3, 663–672. [Google Scholar] [CrossRef] [PubMed]
- Klotz, L.O.; Briviba, K.; Sies, H. Singlet oxygen mediates the activation of JNK by UVA radiation in human skin fibroblasts. FEBS Lett. 1997, 408, 289–291. [Google Scholar] [CrossRef]
- Klotz, L.O.; Pellieux, C.; Briviba, K.; Pierlot, C.; Aubry, J.M.; Sies, H. Mitogen-activated protein kinase (p38-, JNK-, ERK-) activation pattern induced by extracellular and intracellular singlet oxygen and UVA. Eur. J. Biochem. 1999, 260, 917–922. [Google Scholar] [CrossRef]
- Godar, D.E. UVA1 radiation triggers two different final apoptotic pathways. J. Investig. Dermatol. 1999, 112, 3–12. [Google Scholar] [CrossRef] [PubMed]
- Zhuang, S.; Demirs, J.T.; Kochevar, I.E. p38 mitogen-activated protein kinase mediates bid cleavage, mitochondrial dysfunction, and caspase-3 activation during apoptosis induced by singlet oxygen but not by hydrogen peroxide. J. Biol. Chem. 2000, 275, 25939–25948. [Google Scholar] [CrossRef]
- Stoyanovsky, D.A.; Tyurina, Y.Y.; Shrivastava, I.; Bahar, I.; Tyurin, V.A.; Protchenko, O.; Jadhav, S.; Bolevich, S.B.; Kozlov, A.V.; Vladimirov, Y.A.; et al. Iron catalysis of lipid peroxidation in ferroptosis: Regulated enzymatic or random free radical reaction? Free Radic. Biol. Med. 2019, 133, 153–161. [Google Scholar] [CrossRef]
- Ursini, F.; Maiorino, M. Lipid peroxidation and ferroptosis: The role of GSH and GPx4. Free Radic. Biol. Med. 2020, 152, 175–185. [Google Scholar] [CrossRef] [PubMed]
- Jiang, X.; Stockwell, B.R.; Conrad, M. Ferroptosis: Mechanisms, biology and role in disease. Nat. Rev. Mol. Cell Biol. 2021, 22, 266–282. [Google Scholar] [CrossRef]
- Tabatabaie, T.; Floyd, R.A. Susceptibility of glutathione peroxidase and glutathione reductase to oxidative damage and the protective effect of spin trapping agents. Arch. Biochem. Biophys. 1994, 314, 112–119. [Google Scholar] [CrossRef] [PubMed]
- Suryo Rahmanto, A.; Pattison, D.I.; Davies, M.J. Photo-oxidation-induced inactivation of the selenium-containing protective enzymes thioredoxin reductase and glutathione peroxidase. Free Radic. Biol. Med. 2012, 53, 1308–1316. [Google Scholar] [CrossRef]
- Chin, Y.C.; Yang, L.X.; Hsu, F.T.; Hsu, C.W.; Chang, T.W.; Chen, H.Y.; Chen, L.Y.; Chia, Z.C.; Hung, C.H.; Su, W.C.; et al. Iron oxide@chlorophyll clustered nanoparticles eliminate bladder cancer by photodynamic immunotherapy-initiated ferroptosis and immunostimulation. J. Nanobiot. 2022, 20, 373. [Google Scholar] [CrossRef]
- Ke, L.; Wei, F.; Xie, L.; Karges, J.; Chen, Y.; Ji, L.; Chao, H. A Biodegradable Iridium(III) Coordination Polymer for Enhanced Two-Photon Photodynamic Therapy Using an Apoptosis-Ferroptosis Hybrid Pathway. Angew Chem. Int. Ed. Engl. 2022, 61, e202205429. [Google Scholar] [CrossRef]
- Nomura, K.; Imai, H.; Koumura, T.; Kobayashi, T.; Nakagawa, Y. Mitochondrial phospholipid suppressing the peroxidation of cardiolipin in hypoglycaemia-induced apoptosis. Biochem. J. 2000, 351, 183–193. [Google Scholar] [CrossRef] [PubMed]





Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).