


Ring-Modified Histidine-Containing Cationic Short Peptides Exhibit Anticryptococcal Activity by Cellular Disruption
 ,
,
Abstract
1. Introduction
2. Results and Discussion
2.1. Synthesis
2.2. Antimicrobial Susceptibility Testing
2.3. Cytotoxicity Assay
2.4. Hemolytic Assay
2.5. Time Kill Kinetics
2.6. Drug Combination Study
2.7. Mechanistic Investigations
2.7.1. Cell Death Pattern Analysis Using Flow Cytometry
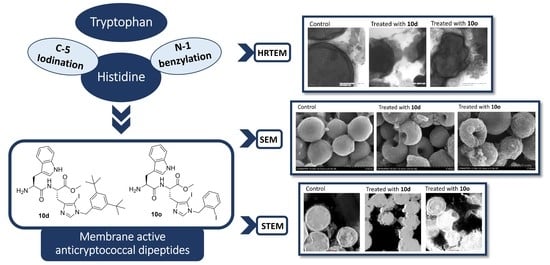
2.7.2. Surface Morphology Analysis Using Scanning Electron Microscopy (SEM)
2.7.3. Internal Cell Morphology Analysis Using Transmission Electron Microscopy (TEM)
2.7.4. Confocal Laser Scanning Microscopy (CLSM)
- (a)
- Membrane permeabilization and localization of FITC-labeled peptide
- (b)
- Membrane disruption and DNA interaction analysis by propidium iodide (PI) uptake
- (c)
- Detection of nuclear fragmentation
2.7.5. Possible Mechanism
3. Conclusions and Summary
4. Material and Methods
4.1. Broth Microdilution Assay
4.2. Cytotoxicity Assay
4.3. Hemolytic Assay
4.4. Time Kill Assay
4.5. Drug Combination Study
4.6. Flow Cytometry
4.7. SEM
4.8. TEM
4.9. CLSM
- (a)
- FITC-labeled peptide uptake assay
- (b)
- PI uptake assay
- (c)
- DAPI uptake assay
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Rajasingham, R.; Govender, N.P.; Jordan, A.; Loyse, A.; Shroufi, A.; Denning, D.W.; Meya, D.B.; Chiller, T.M.; Boulware, D.R. The global burden of HIV-associated cryptococcal infection in adults in 2020: A modelling analysis. Lancet Infect. Dis. 2022, 22, 1748–1755. [Google Scholar] [CrossRef] [PubMed]
- Ngan, N.T.T.; Flower, B.; Day, J.N. Treatment of Cryptococcal Meningitis: How Have We Got Here and Where are We Going? Drugs 2022, 82, 1237–1249. [Google Scholar] [CrossRef] [PubMed]
- Jarvis, J.N.; Lawrence, D.S.; Meya, D.B.; Kagimu, E.; Kasibante, J.; Mpoza, E.; Rutakingirwa, M.K.; Ssebambulidde, K.; Tugume, L.; Rhein, J. Single-Dose Liposomal Amphotericin B Treatment for Cryptococcal Meningitis. N. Engl. J. Med. 2022, 386, 1109–1120. [Google Scholar] [CrossRef] [PubMed]
- Muhaj, F.F.; George, S.J.; Nguyen, C.D.; Tyring, S.K. Antimicrobials and resistance part II: Antifungals, antivirals, and antiparasitics. J. Am. Acad. Dermatol. 2022, 86, 1207–1226. [Google Scholar] [CrossRef] [PubMed]
- Magana, M.; Pushpanathan, M.; Santos, A.L.; Leanse, L.; Fernandez, M.; Ioannidis, A.; Giulianotti, M.A.; Apidianakis, Y.; Bradfute, S.; Ferguson, A.L.; et al. The value of antimicrobial peptides in the age of resistance. Lancet Infect. Dis. 2020, 20, e216–e230. [Google Scholar] [CrossRef] [PubMed]
- Mahindra, A.; Sharma, K.K.; Rathore, D.; Khan, S.I.; Jacob, M.R.; Jain, R. Synthesis and antimicrobial activities of His(2-aryl)-Arg and Trp-His(2-aryl) classes of dipeptidomimetics. MedChemComm 2014, 5, 671–676. [Google Scholar] [CrossRef] [PubMed]
- Iyer, K.R.; Revie, N.M.; Fu, C.; Robbins, N.; Cowen, L.E. Treatment strategies for cryptococcal infection: Challenges, advances and future outlook. Nat. Rev. Microbiol. 2021, 19, 454–466. [Google Scholar] [CrossRef]
- Struyfs, C.; Cammue, B.P.A.; Thevissen, K. Membrane-interacting antifungal peptides. Front. Cell Dev. Biol. 2021, 9, 649875. [Google Scholar] [CrossRef]
- Sharma, K.K.; Maurya, I.K.; Khan, S.I.; Jacob, M.R.; Kumar, V.; Tikoo, K.; Jain, R. Discovery of a membrane-active, ring-modified histidine containing ultrashort amphiphilic peptide that exhibits potent inhibition of Cryptococcus neoformans. J. Med. Chem. 2017, 60, 6607–6621. [Google Scholar] [CrossRef]
- Sharma, K.K.; Ravi, R.; Maurya, I.K.; Kapadia, A.; Khan, S.I.; Kumar, V.; Tikoo, K.; Jain, R. Modified histidine containing amphipathic ultrashort antifungal peptide, His-[2-p-(n-butyl) phenyl]-Trp-Arg-OMe exhibits potent anticryptococcal activity. Eur. J. Med. Chem. 2021, 223, 113635. [Google Scholar] [CrossRef]
- Mahindra, A.; Bagra, N.; Wangoo, N.; Jain, R.; Khan, S.I.; Jacob, M.R.; Jain, R. Synthetically modified L-histidine-rich peptidomimetics exhibit potent activity against Cryptococcus neoformans. Bioorg. Med. Chem. Lett. 2014, 24, 3150–3154. [Google Scholar] [CrossRef]
- Mahindra, A.; Bagra, N.; Wangoo, N.; Khan, S.I.; Jacob, M.R.; Jain, R. Discovery of short peptides exhibiting high potency against Cryptococcus neoformans. ACS Med. Chem. Lett. 2014, 5, 315–320. [Google Scholar] [CrossRef] [PubMed]
- Sharma, K.K.; Sharma, K.; Kudwal, A.; Khan, S.I.; Jain, R. Peptide-Heterocycle Conjugates as Antifungals against Cryptococcosis. Asian J. Org. Chem. 2022, 11, e202200196. [Google Scholar] [CrossRef]
- Greco, I.; Molchanova, N.; Holmedal, E.; Jenssen, H.; Hummel, B.D.; Watts, J.L.; Håkansson, J.; Hansen, P.R.; Svenson, J. Correlation between hemolytic activity, cytotoxicity and systemic in vivo toxicity of synthetic antimicrobial peptides. Sci. Rep. 2020, 10, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Sharma, K.; Sharma, K.K.; Sharma, A.; Jain, R. Peptide-based drug discovery: Current status and recent advances. Drug Discov. Today 2022, 103464. [Google Scholar] [CrossRef] [PubMed]
- Mahindra, A.; Bagra, N.; Jain, R. Palladium-catalyzed regioselective C-5 arylation of protected L-histidine: Microwave-assisted C–H activation adjacent to donor arm. J. Org. Chem. 2013, 78, 10954–10959. [Google Scholar] [CrossRef] [PubMed]
- Sharma, K.K.; Mandloi, M.; Jain, R. Regioselective copper-catalyzed N(1)-(hetero)arylation of protected histidine. Org. Biomol. Chem. 2016, 14, 8937–8941. [Google Scholar] [CrossRef]
- Sharma, K.K.; Mandloi, M.; Jain, R. Regioselective access to 1,2-diarylhistidines through the copper-catalyzed n1-arylation of 2-arylhistidines. Eur. J. Org. Chem. 2017, 2017, 984–988. [Google Scholar] [CrossRef]
- Song, L.; Ojeda-Carralero, G.M.; Parmar, D.; González-Martínez, D.A.; Van Meervelt, L.; Van der Eycken, J.; Goeman, J.; Rivera, D.G.; Van der Eycken, E.V. Chemoselective Peptide Backbone Diversification and Bioorthogonal Ligation by Ruthenium-Catalyzed C−H Activation/Annulation. Adv. Synth. Catal. 2021, 363, 3297–3304. [Google Scholar] [CrossRef]
- O’Brien, E.A.; Sharma, K.K.; Byerly-Duke, J.; Camacho, L.A., III; VanVeller, B. A General Strategy to Install Amidine Functional Groups Along the Peptide Backbone. J. Am. Chem. Soc. 2022, 144, 22397–22402. [Google Scholar] [CrossRef]
- Jia, F.; Zhang, Y.; Wang, J.; Peng, J.; Zhao, P.; Zhang, L.; Yao, H.; Ni, J.; Wang, K. The effect of halogenation on the antimicrobial activity, antibiofilm activity, cytotoxicity and proteolytic stability of the antimicrobial peptide Jelleine-I. Peptides 2019, 112, 56–66. [Google Scholar] [CrossRef] [PubMed]
- Arias, M.; Hoffarth, E.R.; Ishida, H.; Aramini, J.M.; Vogel, H.J. Recombinant expression, antimicrobial activity and mechanism of action of tritrpticin analogs containing fluoro-tryptophan residues. Biochim. Biophys. Acta Biomembr. 2016, 1858, 1012–1023. [Google Scholar] [CrossRef] [PubMed]
- Cruz, J.o.C.S.; Iorio, M.; Monciardini, P.; Simone, M.; Brunati, C.; Gaspari, E.; Maffioli, S.I.; Wellington, E.; Sosio, M.; Donadio, S. Brominated variant of the lantibiotic NAI-107 with enhanced antibacterial potency. J. Nat. Prod. 2015, 78, 2642–2647. [Google Scholar] [CrossRef] [PubMed]
- Molchanova, N.; Nielsen, J.E.; Sørensen, K.B.; Prabhala, B.K.; Hansen, P.R.; Lund, R.; Barron, A.E.; Jenssen, H. Halogenation as a tool to tune antimicrobial activity of peptoids. Sci. Rep. 2020, 10, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Gottler, L.M.; de la Salud Bea, R.; Shelburne, C.E.; Ramamoorthy, A.; Marsh, E.N.G. Using Fluorous Amino Acids to Probe the Effects of Changing Hydrophobicity on the Physical and Biological Properties of the β-Hairpin Antimicrobial Peptide Protegrin-1. Biochemistry 2008, 47, 9243–9250. [Google Scholar] [CrossRef]
- Sharma, K.; Aaghaz, S.; Maurya, I.K.; Rudramurthy, S.M.; Singh, S.; Kumar, V.; Tikoo, K.; Jain, R. Antifungal evaluation and mechanistic investigations of membrane active short synthetic peptides-based amphiphiles. Bioorg. Chem. 2022, 127, 106002. [Google Scholar] [CrossRef]
- Sharma, K.; Aaghaz, S.; Maurya, I.K.; Sharma, K.K.; Singh, S.; Rudramurthy, S.M.; Kumar, V.; Tikoo, K.; Jain, R. Synthetic amino acids-derived peptides target Cryptococcus neoformans by inducing cell membrane disruption. Bioorg. Chem. 2023, 130, 106252. [Google Scholar] [CrossRef]
- Jain, R.; Avramovitch, B.; Cohen, L.A. Synthesis of ring-halogenated histidines and histamines. Tetrahedron 1998, 54, 3235–3242. [Google Scholar] [CrossRef]
- Maurya, I.K.; Thota, C.K.; Sharma, J.; Tupe, S.G.; Chaudhary, P.; Singh, M.K.; Thakur, I.S.; Deshpande, M.; Prasad, R.; Chauhan, V.S. Mechanism of action of novel synthetic dodecapeptides against Candida albicans. Biochim. Biophys. Acta 2013, 1830, 5193–5203. [Google Scholar] [CrossRef]
- Vermes, I.; Haanen, C.; Steffens-Nakken, H.; Reutelingsperger, C. Flow cytometric detection of phosphatidylserine expression on early apoptotic cells using fluorescein labelled Annexin V. J. Immunol. Methods 1995, 184, 39–51. [Google Scholar] [CrossRef]
- Adan, A.; Alizada, G.; Kiraz, Y.; Baran, Y.; Nalbant, A. Flow cytometry: Basic principles and applications. Crit. Rev. Biotechnol. 2017, 37, 163–176. [Google Scholar] [CrossRef] [PubMed]
- Liu, T.; Zhu, W.; Yang, X.; Chen, L.; Yang, R.; Hua, Z.; Li, G. Detection of apoptosis based on the interaction between annexin V and phosphatidylserine. Anal. Chem. 2009, 81, 2410–2413. [Google Scholar] [CrossRef]
- Henry, C.M.; Hollville, E.; Martin, S.J. Measuring apoptosis by microscopy and flow cytometry. Methods 2013, 61, 90–97. [Google Scholar] [CrossRef] [PubMed]
- CLSI. Reference Method for Broth Dilution Antifungal Susceptibility Testing of Yeasts, CLSI Supplement M27; Clinical and Laboratory Standards Institute: Wayne, PA, USA, 2017. [Google Scholar]
- CLSI. Reference Method for Broth Dilution Antifungal Susceptibility Testing of Filamentous Fungi, CLSI Supplement M38; Clinical and Laboratory Standards Institute: Wayne, PA, USA, 2017. [Google Scholar]
- CLSI. Methods for Dilution Antimicrobial Susceptibility Tests for Bacteria That Grow Aerobically, CLSI Supplement M07; Clinical and Laboratory Standards Institute: Wayne, PA, USA, 2018. [Google Scholar]
- Sharma, A.; Singh, S.; Tewari, R.; Bhatt, V.P.; Sharma, J.; Maurya, I.K. Phytochemical analysis and mode of action against Candida glabrata of Paeonia emodi extracts. J. Mycol. Med. 2018, 28, 443–451. [Google Scholar] [CrossRef] [PubMed]
- Green, R.M.; Bicker, K.L. Evaluation of peptoid mimics of short, lipophilic peptide antimicrobials. Int. J. Antimicrob. Agents 2020, 56, 106048. [Google Scholar] [CrossRef]
- Mares, D. Electron microscopy of Microsporum cookei after ’in vitro’ treatment with protoanemonin: A combined SEM and TEM study. Mycopathologia 1989, 108, 37–46. [Google Scholar] [CrossRef] [PubMed]
- Park, C.; Lee, D.G. Melittin induces apoptotic features in Candida albicans. Biochem. Biophys. Res. Commun. 2010, 394, 170–172. [Google Scholar] [CrossRef]
- Abdo, M.-R.; Joseph, P.; Boigegrain, R.-A.; Liautard, J.-P.; Montero, J.-L.; Köhler, S.; Winum, J.-Y. Brucella suis histidinol dehydrogenase: Synthesis and inhibition studies of a series of substituted benzylic ketones derived from histidine. Bioorg. Med. Chem. 2007, 15, 4427–4433. [Google Scholar] [CrossRef]
- Meena, C.L.; Thakur, A.; Nandekar, P.P.; Sangamwar, A.T.; Sharma, S.S.; Jain, R. Synthesis of CNS active thyrotropin-releasing hormone (TRH)-like peptides: Biological evaluation and effect on cognitive impairment induced by cerebral ischemia in mice. Bioorg. Med. Chem. 2015, 23, 5641–5653. [Google Scholar] [CrossRef]












{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
 | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| No. | R | C. neoformans | C. albicans | C. glabrata | C. parapsilosis | C. krusei | |||||
| IC50 a | MIC b | IC50 | MIC | IC50 | MIC | IC50 | MIC | IC50 | MIC | ||
| 9a | H | 17.78 | 32.34 | >35.2 | >64.0 | >35.2 | >64.0 | >35.2 | >64.0 | >35.2 | >64.0 |
| 9b | 4-tert-butyl | >35.2 | >64.0 | >35.2 | >64.0 | >35.2 | >64.0 | >35.2 | >64.0 | >35.2 | >64.0 |
| 9c | 4-iso-propyl | 20.39 | 37.08 | >35.2 | >64.0 | >35.2 | >64.0 | >35.2 | >64.0 | >35.2 | >64.0 |
| 9d | 3,5-di-tert-butyl | >35.2 | >64.0 | >35.2 | >64.0 | >35.2 | >64.0 | >35.2 | >64.0 | >35.2 | >64.0 |
| 9e | 3-trifluoromethyl | 20.71 | 37.65 | >35.2 | >64.0 | >35.2 | >64.0 | >35.2 | >64.0 | >35.2 | >64.0 |
| 9f | 2-trifluoromethyl | 30.56 | 55.57 | >35.2 | >64.0 | >35.2 | >64.0 | >35.2 | >64.0 | >35.2 | >64.0 |
| 9g | 4-trifluoromethyl | >35.2 | >64.0 | >35.2 | >64.0 | >35.2 | >64.0 | >35.2 | >64.0 | >35.2 | >64.0 |
| 9h | 4-nitro | >35.2 | >64.0 | >35.2 | >64.0 | >35.2 | >64.0 | >35.2 | >64.0 | >35.2 | >64.0 |
| 9i | 3,4-difluoro | 17.62 | 32.05 | >35.2 | >64.0 | >35.2 | >64.0 | >35.2 | >64.0 | >35.2 | >64.0 |
| 9j | 3,5-difluoro | 17.73 | 32.25 | >35.2 | >64.0 | >35.2 | >64.0 | >35.2 | >64.0 | >35.2 | >64.0 |
| 9k | 3-fluoro | 18.16 | 32.02 | >35.2 | >64.0 | >35.2 | >64.0 | >35.2 | >64.0 | >35.2 | >64.0 |
| 9l | 3-chloro | 18.59 | 33.80 | >35.2 | >64.0 | >35.2 | >64.0 | >35.2 | >64.0 | >35.2 | >64.0 |
| 9m | 3-bromo | 18.63 | 33.88 | >35.2 | >64.0 | >35.2 | >64.0 | >35.2 | >64.0 | >35.2 | >64.0 |
| 9n | 3-iodo | 23.58 | 42.87 | >35.2 | >64.0 | >35.2 | >64.0 | >35.2 | >64.0 | >35.2 | >64.0 |
| 9o | 2-iodo | >35.2 | >64.0 | >35.2 | >64.0 | >35.2 | >64.0 | >35.2 | >64.0 | >35.2 | >64.0 |
| Amphotericin B | 0.50 | 1.00 | 0.36 | 0.72 | 0.18 | 0.36 | 0.36 | 0.72 | 0.36 | 0.72 | |
 | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| No. | R1 | C. neoformans | C. albicans | C. glabrata | C. parapsilosis | C. krusei | |||||
| IC50 a | MIC b | IC50 | MIC | IC50 | MIC | IC50 | MIC | IC50 | MIC | ||
| 10a | H | 17.68 | 32.15 | >35.2 | >64.0 | >35.2 | >64.0 | >35.2 | >64.0 | >35.2 | >64.0 |
| 10b | 4-tert-butyl | 5.15 | 9.358 | >35.2 | >64.0 | >35.2 | >64.0 | >35.2 | >64.0 | >35.2 | >64.0 |
| 10c | 4-iso-propyl | 8.81 | 16.02 | >35.2 | >64.0 | >35.2 | >64.0 | >35.2 | >64.0 | >35.2 | >64.0 |
| 10d | 3,5-di-tert-butyl | 2.20 | 4.01 | >35.2 | >64.0 | >35.2 | >64.0 | >35.2 | >64.0 | >35.2 | >64.0 |
| 10e | 3-trifluoromethyl | 4.87 | 8.86 | >35.2 | >64.0 | >35.2 | >64.0 | >35.2 | >64.0 | >35.2 | >64.0 |
| 10f | 2-trifluoromethyl | 17.70 | 32.18 | >35.2 | >64.0 | >35.2 | >64.0 | >35.2 | >64.0 | >35.2 | >64.0 |
| 10g | 4-trifluoromethyl | 13.53 | 24.60 | >35.2 | >64.0 | >35.2 | >64.0 | >35.2 | >64.0 | >35.2 | >64.0 |
| 10h | 4-nitro | >35.2 | >64.0 | >35.2 | >64.0 | >35.2 | >64.0 | >35.2 | >64.0 | >35.2 | >64.0 |
| 10i | 3,4-difluoro | 10.28 | 18.70 | >35.2 | >64.0 | >35.2 | >64.0 | >35.2 | >64.0 | >35.2 | >64.0 |
| 10j | 3,5-difluoro | 6.09 | 11.08 | >35.2 | >64.0 | >35.2 | >64.0 | >35.2 | >64.0 | >35.2 | >64.0 |
| 10k | 3-fluoro | 2.78 | 4.64 | >35.2 | >64.0 | >35.2 | >64.0 | >35.2 | >64.0 | >35.2 | >64.0 |
| 10l | 3-chloro | 17.66 | 32.11 | >35.2 | >64.0 | >35.2 | >64.0 | >35.2 | >64.0 | >35.2 | >64.0 |
| 10m | 3-bromo | 9.23 | 16.79 | >35.2 | >64.0 | >35.2 | >64.0 | >35.2 | >64.0 | >35.2 | >64.0 |
| 10n | 3-iodo | 17.62 | 32.05 | >35.2 | >64.0 | >35.2 | >64.0 | >35.2 | >64.0 | >35.2 | >64.0 |
| 10o | 2-iodo | 2.59 | 4.59 | >35.2 | >64.0 | >35.2 | >64.0 | >35.2 | >64.0 | >35.2 | >64.0 |
| Amphotericin B | 0.50 | 1.00 | 0.36 | 0.72 | 0.18 | 0.36 | 0.36 | 0.72 | 0.36 | 0.72 | |
 | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| No. | R1 | R2 | E. faecalis | S. aureus | S. pyogens | E. coli | P. aeruginosa | |||||
| IC50 a | MIC b | IC50 | MIC | IC50 | MIC | IC50 | MIC | IC50 | MIC | |||
| 10b | 4-tert-butylbenzyl | H | >35.2 | >64.0 | 35.2 | 64.0 | >35.2 | >64.0 | 35.2 | 64.0 | >35.2 | >64.0 |
| 10c | 4-n-butylbenzyl | H | >35.2 | >64.0 | >35.2 | >64.0 | >35.2 | >64.0 | >35.2 | >64.0 | >35.2 | >64.0 |
| 10d | 3,5-di-tert-butylbenzyl | H | 17.6 | 32.0 | 17.6 | 32.0 | >35.2 | >64.0 | 17.6 | 32.0 | >35.2 | >64.0 |
| 10e | 3-trifluoromethyl | H | >35.2 | >64.0 | >35.2 | >64.0 | >35.2 | >64.0 | >35.2 | >64.0 | >35.2 | >64.0 |
| 10k | 3,5-difluoro | H | >35.2 | >64.0 | >35.2 | >64.0 | >35.2 | >64.0 | >35.2 | >64.0 | >35.2 | >64.0 |
| 10l | 3-fluoro | H | >35.2 | >64.0 | >35.2 | >64.0 | >35.2 | >64.0 | >35.2 | >64.0 | >35.2 | >64.0 |
| 10n | 3-bromo | H | >35.2 | >64.0 | >35.2 | >64.0 | >35.2 | >64.0 | >35.2 | >64.0 | >35.2 | >64.0 |
| 10o | 2-iodo | H | 17.6 | 32.0 | >35.2 | >64.0 | >35.2 | >64.0 | >35.2 | >64.0 | >35.2 | >64.0 |
| Rifampicin | 1.00 | 2.00 | 2.00 | 4.00 | 1.00 | 2.00 | 1.00 | 2.00 | 16.00 | 32.00 | ||
| Peptides | MIC (μg/mL) | Cytotoxicity (TC50) a (μg/mL) | % Hemolysis (μg/mL) | ||
|---|---|---|---|---|---|
| HEK 293 | HeLa | HC10 b | HC50 c | ||
| 10d | 4.01 | >100 | >100 | 50.53 | >100 |
| 10o | 4.59 | >100 | >100 | 52.17 | >100 |
| Treatment | MIC (μg/mL) | Fold Decrease in MIC | FICI | Effect | |
|---|---|---|---|---|---|
| Alone | In Combination | ||||
| 10d | 4.01 | 0.250 | 16 | 0.18 | Synergistic |
| Amp B | 1.00 | 0.125 | 8 | ||
| 10d | 4.01 | 0.501 | 8 | 0.19 | Synergistic |
| Flu | 10.0 | 0.625 | 16 | ||
| 10o | 4.59 | 0.287 | 16 | 0.31 | Synergistic |
| Amp B | 1.00 | 0.250 | 4 | ||
| 10o | 4.59 | 2.999 | 1.5 | 1.00 | Additive |
| Flu | 10.0 | 5.0 | 2 | ||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sharma, K.; Aaghaz, S.; Maurya, I.K.; Singh, S.; Rudramurthy, S.M.; Kumar, V.; Tikoo, K.; Jain, R. Ring-Modified Histidine-Containing Cationic Short Peptides Exhibit Anticryptococcal Activity by Cellular Disruption. Molecules 2023, 28, 87. https://doi.org/10.3390/molecules28010087
Sharma K, Aaghaz S, Maurya IK, Singh S, Rudramurthy SM, Kumar V, Tikoo K, Jain R. Ring-Modified Histidine-Containing Cationic Short Peptides Exhibit Anticryptococcal Activity by Cellular Disruption. Molecules. 2023; 28(1):87. https://doi.org/10.3390/molecules28010087
Chicago/Turabian StyleSharma, Komal, Shams Aaghaz, Indresh Kumar Maurya, Shreya Singh, Shivaprakash M. Rudramurthy, Vinod Kumar, Kulbhushan Tikoo, and Rahul Jain. 2023. "Ring-Modified Histidine-Containing Cationic Short Peptides Exhibit Anticryptococcal Activity by Cellular Disruption" Molecules 28, no. 1: 87. https://doi.org/10.3390/molecules28010087
APA StyleSharma, K., Aaghaz, S., Maurya, I. K., Singh, S., Rudramurthy, S. M., Kumar, V., Tikoo, K., & Jain, R. (2023). Ring-Modified Histidine-Containing Cationic Short Peptides Exhibit Anticryptococcal Activity by Cellular Disruption. Molecules, 28(1), 87. https://doi.org/10.3390/molecules28010087