

Ameliorative Effect of Citrus Lemon Peel Extract and Resveratrol on Premature Ovarian Failure Rat Model: Role of iNOS/Caspase-3 Pathway
 , , , , , , and
, , , , , , and
Abstract
1. Introduction
2. Results
2.1. Effect of LPE and RES on the Serum Hormonal Markers of Cyclophosphamide-Treated Rats
2.2. Effect of LPE and RES on the Serum Antioxidants and Oxidative Stress Markers of Cyclophosphamide-Treated Rats
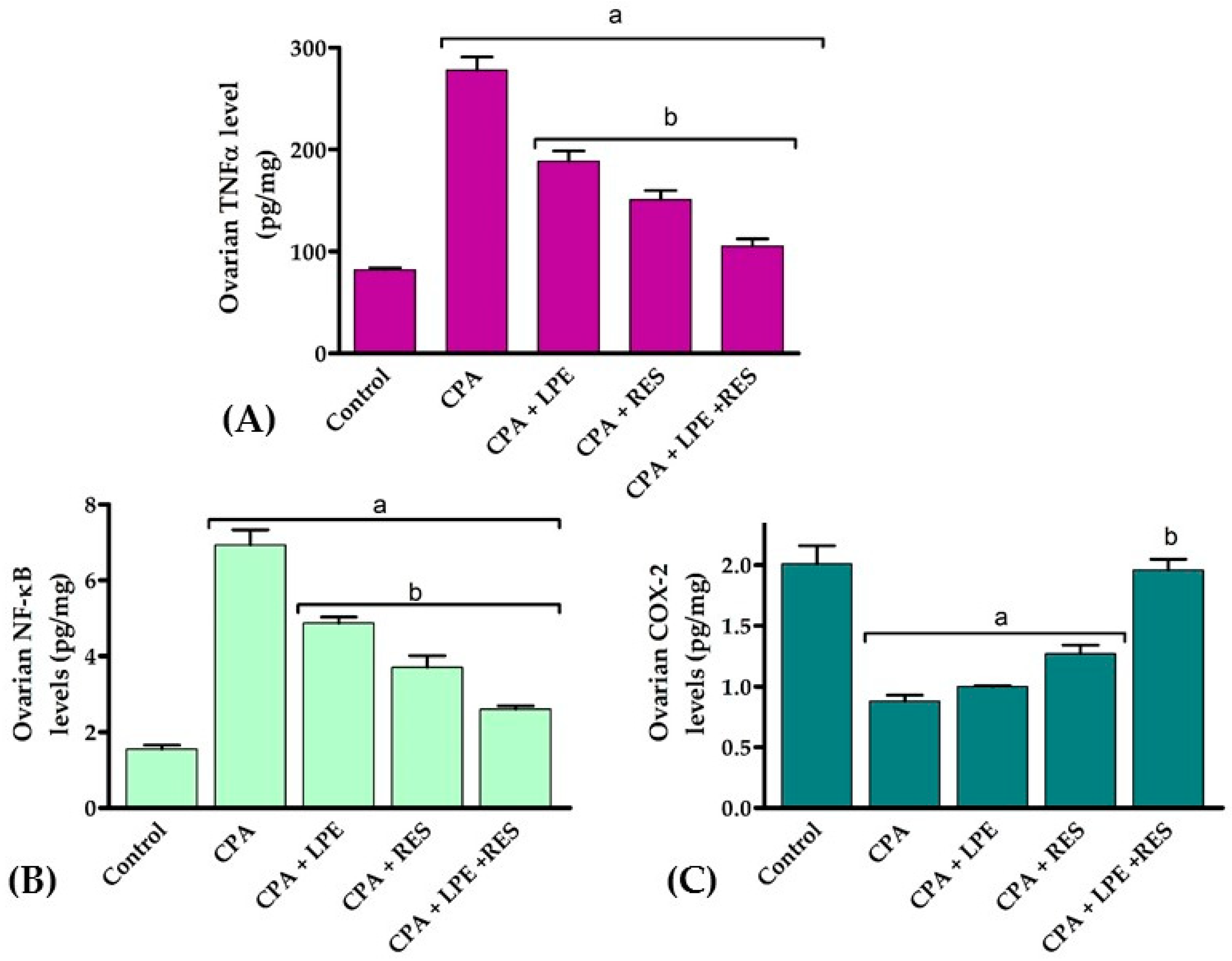
2.3. Effects of LPE and RES on Ovarian Tissue Inflammation Markers in Cyclophosphamide-Treated Rats
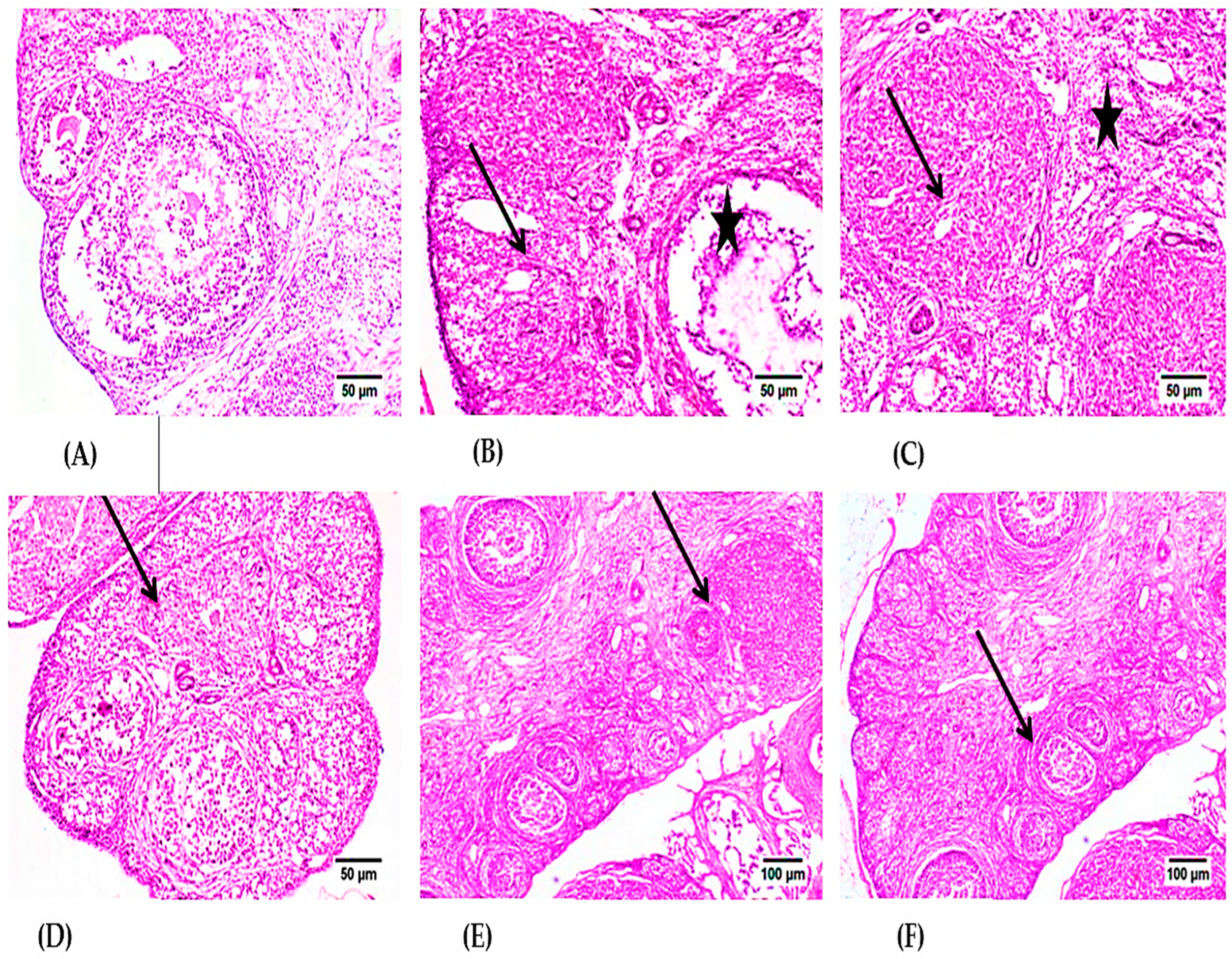
2.4. Histopathological Investigations
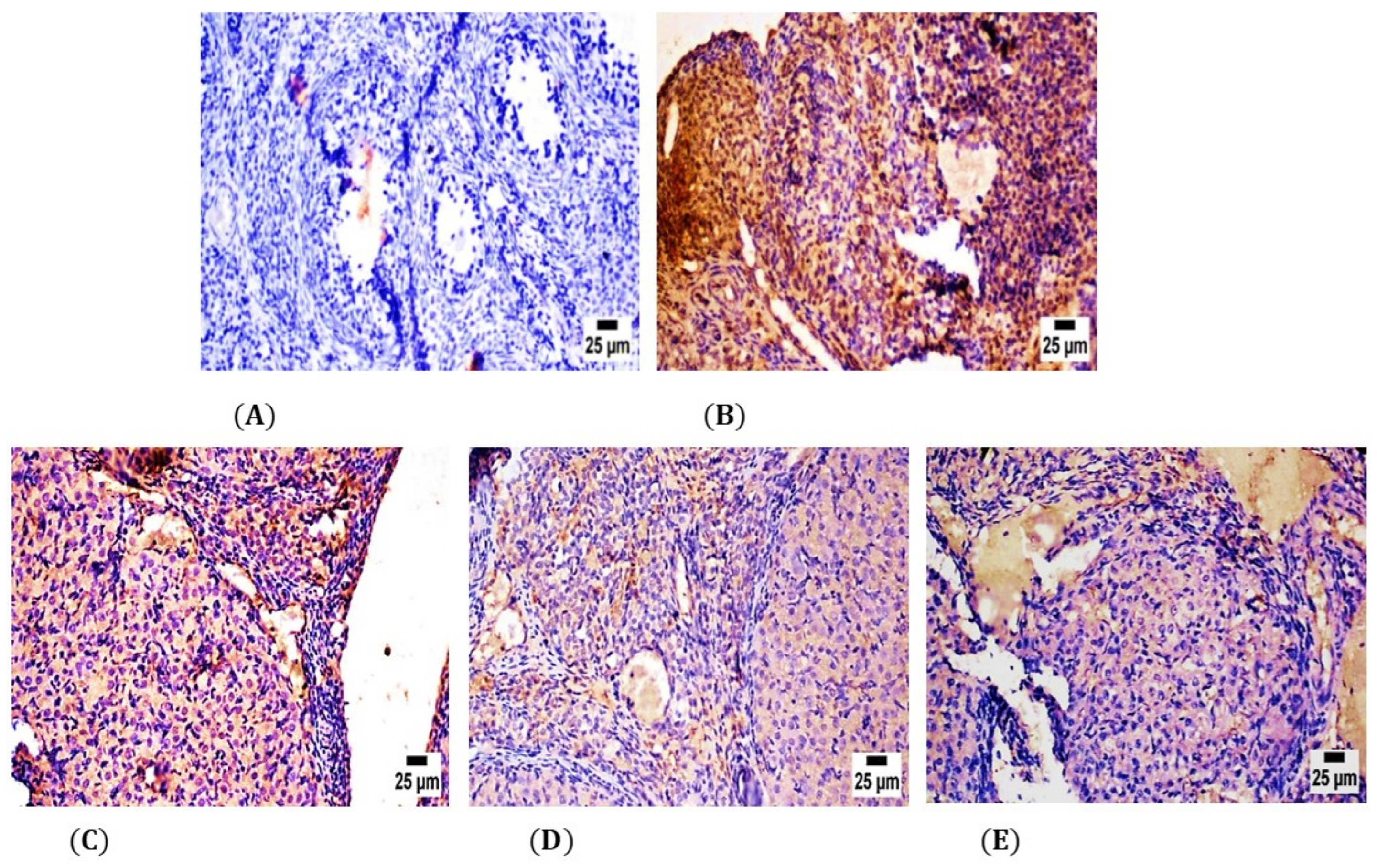
2.5. Immunohistochemical Analysis
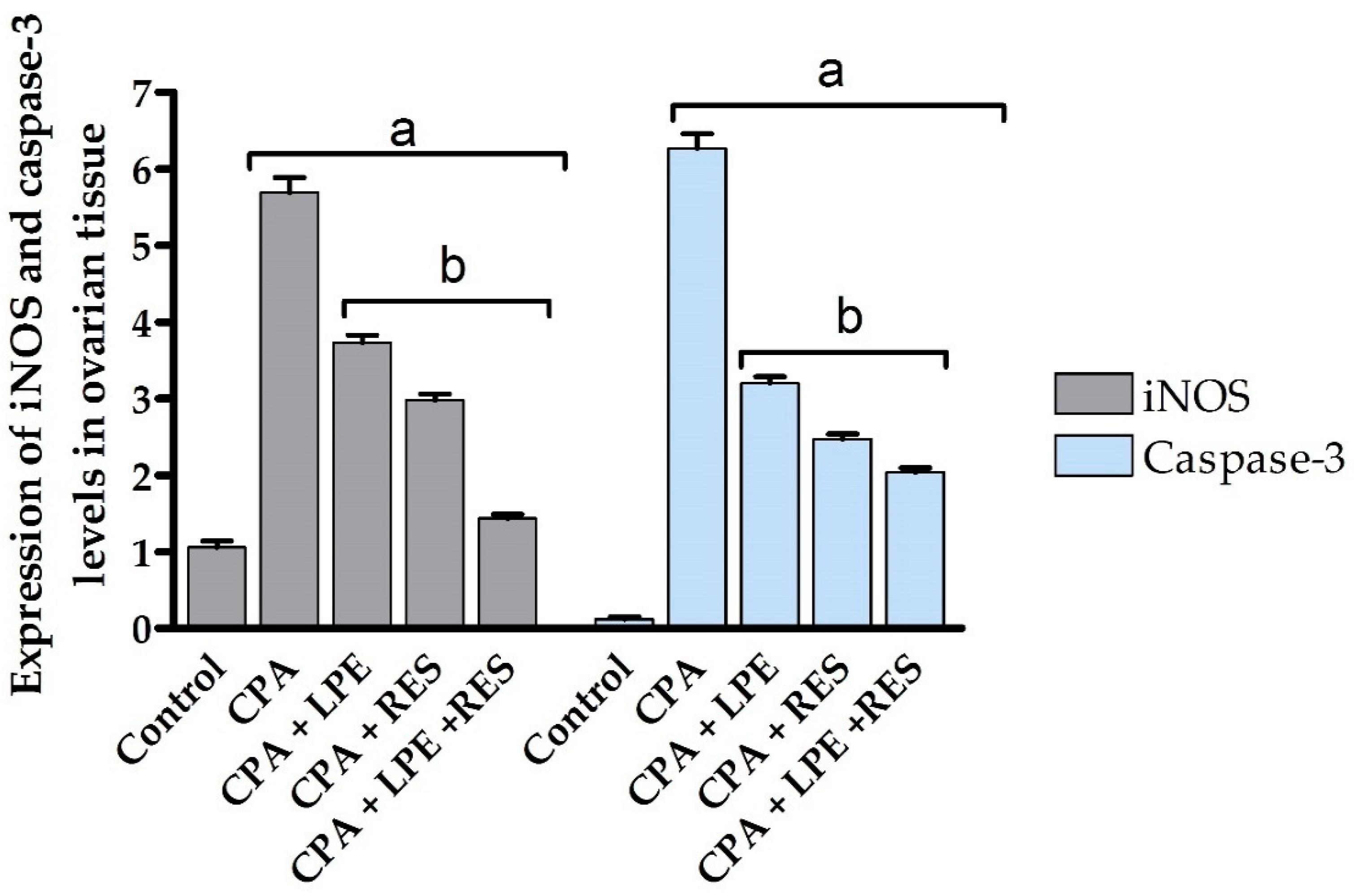
2.6. Gene Expression Analysis
2.6.1. iNOS Gene Expression Analysis
2.6.2. Casp3 Gene Expression Analysis
3. Discussion
4. Materials and Methods
4.1. Drugs and Chemicals
4.2. Lemon Peel Extract Preparation
4.3. Experimental Animals
4.3.1. Induction of POF
4.3.2. Study Design
4.3.3. Collection of Samples
4.4. Biochemical Analysis
4.4.1. Serum Fertility Hormones Measurements
4.4.2. Determination of Serum Oxidative Stress and Antioxidant Markers
4.4.3. Measurements of Ovarian Inflammatory Markers
4.5. Histopathological Evaluation
4.6. Immunohistochemical Evaluation
4.7. qRTPCR for Determination of iNOS and Casp3 Expression Levels
4.8. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Sample Availability
References
- Rahman, R.; Panay, N. Diagnosis and Management of Premature Ovarian Insufficiency. Best Pract. Res. Clin. Endocrinol. Metab. 2021, 35, 101600. [Google Scholar] [CrossRef] [PubMed]
- Cooper, A.R.; Baker, V.L.; Sterling, E.W.; Ryan, M.E.; Woodruff, T.K.; Nelson, L.M. The Time Is Now for a New Approach to Primary Ovarian Insufficiency. Fertil. Steril. 2011, 95, 1890–1897. [Google Scholar] [CrossRef] [PubMed]
- Bedoschi, G.; Navarro, P.A.; Oktay, K. Chemotherapy-Induced Damage to Ovary: Mechanisms and Clinical Impact. Future Oncol. 2016, 12, 2333–2344. [Google Scholar] [CrossRef] [PubMed]
- Yuksel, A.; Bildik, G.; Senbabaoglu, F.; Akin, N.; Arvas, M.; Unal, F.; Kilic, Y.; Karanfil, I.; Eryılmaz, B.; Yilmaz, P.; et al. The Magnitude of Gonadotoxicity of Chemotherapy Drugs on Ovarian Follicles and Granulosa Cells Varies Depending upon the Category of the Drugs and the Type of Granulosa Cells. Hum. Reprod. 2015, 30, 2926–2935. [Google Scholar] [CrossRef]
- Manger, K.; Wildt, L.; Kalden, J.R.; Manger, B. Prevention of Gonadal Toxicity and Preservation of Gonadal Function and Fertility in Young Women with Systemic Lupus Erythematosus Treated by Cyclophosphamide: The PREGO-Study. Autoimmun. Rev. 2006, 5, 269–272. [Google Scholar] [CrossRef]
- Luo, Q.; Yin, N.; Zhang, L.; Yuan, W.; Zhao, W.; Luan, X.; Zhang, H. Role of SDF-1/CXCR4 and Cytokines in the Development of Ovary Injury in Chemotherapy Drug Induced Premature Ovarian Failure Mice. Life Sci. 2017, 179, 103–109. [Google Scholar] [CrossRef]
- Ali, S. Lemon Juice Antioxidant Activity against Oxidative Stress. Baghdad Sci. J. 2020, 17, 207. [Google Scholar] [CrossRef]
- Saini, R.K.; Ranjit, A.; Sharma, K.; Prasad, P.; Shang, X.; Gowda, K.G.M.; Keum, Y.-S. Bioactive Compounds of Citrus Fruits: A Review of Composition and Health Benefits of Carotenoids, Flavonoids, Limonoids, and Terpenes. Antioxidants 2022, 11, 239. [Google Scholar] [CrossRef]
- Nawaz, R.; Safdar, N.; Ainee, A.; Jabbar, S. Development and Storage Stability Studies of Functional Fruit Drink Supplemented with Polyphenols Extracted from Lemon Peels. J. Food Process. Preserv. 2021, 45, e15268. [Google Scholar] [CrossRef]
- Jabeen, E.; Janjua, N.K.; Ahmed, S.; Ali, T.; Murtaza, I.; Ashraf, Z.; Masood, N.; Kalsoom, S. Antioxidant Activity and Hepatotoxicity of Flavonoids and Their Metal Complexes Through Co-Administration of β-Cyclodextrin. ChemistrySelect 2019, 4, 9420–9432. [Google Scholar] [CrossRef]
- Campêlo, L.M.L.; de Almeida, A.A.C.; de Freitas, R.L.M.; Cerqueira, G.S.; de Sousa, G.F.; Saldanha, G.B.; Feitosa, C.M.; de Freitas, R.M. Antioxidant and Antinociceptive Effects of Citrus Limon Essential Oil in Mice. J. Biomed. Biotechnol. 2011, 2011, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Zaidun, N.H.; Thent, Z.C.; Abd Latiff, A. Combating oxidative stress disorders with citrus flavonoid: Naringenin. Life Sci. 2018, 208, 111–122. [Google Scholar] [CrossRef] [PubMed]
- Zaychenko, G.; Stryga, O.; Sinitsyna, O.; Doroshenko, A.; Sulaieva, O.; Falalyeyeva, T.; Kobyliak, N. Resveratrol Effects on the Reproductive System in Ovariectomized Rats: Deciphering Possible Mechanisms. Molecules 2022, 27, 4916. [Google Scholar] [CrossRef] [PubMed]
- Hambrock, A.; de Oliveira Franz, C.B.; Hiller, S.; Grenz, A.; Ackermann, S.; Schulze, D.U.; Drews, G.; Osswald, H. Resveratrol Binds to the Sulfonylurea Receptor (SUR) and Induces Apoptosis in a SUR Subtype-Specific Manner. J. Biol. Chem. 2007, 282, 3347–3356. [Google Scholar] [CrossRef]
- Li, N.; Liu, L. Mechanism of Resveratrol in Improving Ovarian Function in a Rat Model of Premature Ovarian Insufficiency. J. Obstet. Gynaecol. Res. 2018, 44, 1431–1438. [Google Scholar] [CrossRef]
- Finkel, T.; Deng, C.-X.; Mostoslavsky, R. Recent Progress in the Biology and Physiology of Sirtuins. Nature 2009, 460, 587–591. [Google Scholar] [CrossRef]
- Morita, Y.; Wada-Hiraike, O.; Yano, T.; Shirane, A.; Hirano, M.; Hiraike, H.; Koyama, S.; Oishi, H.; Yoshino, O.; Miyamoto, Y.; et al. Resveratrol Promotes Expression of SIRT1 and StAR in Rat Ovarian Granulosa Cells: An Implicative Role of SIRT1 in the Ovary. Reprod. Biol. Endocrinol. 2012, 10, 1–10. [Google Scholar] [CrossRef]
- Pasquariello, R.; Verdile, N.; Brevini, T.A.; Gandolfi, F.; Boiti, C.; Zerani, M.; Maranesi, M. The role of resveratrol in mammalian reproduction. Molecules 2020, 25, 4554. [Google Scholar] [CrossRef]
- Jalalie, L.; Rezaee, M.A.; Rezaie, M.J.; Jalili, A.; Raoofi, A.; Rustamzade, A. Human Umbilical Cord Mesenchymal Stem Cells Improve Morphometric and Histopathologic Changes of Cyclophosphamide-Injured Ovarian Follicles in Mouse Model of Premature Ovarian Failure. Acta Histochem. 2021, 123, 151658. [Google Scholar] [CrossRef]
- Melekoglu, R.; Ciftci, O.; Eraslan, S.; Cetin, A.; Basak, N. Beneficial Effects of Curcumin and Capsaicin on Cyclophosphamide-Induced Premature Ovarian Failure in a Rat Model. J. Ovarian Res. 2018, 11, 33. [Google Scholar] [CrossRef]
- Afifi, N.M.; Reyad, O.N. Role of Mesenchymal Stem Cell Therapy in Restoring Ovarian Function in a Rat Model of Chemotherapy-Induced Ovarian Failure: A Histological and Immunohistochemical Study. Egypt. J. Histol. 2013, 36, 114–126. [Google Scholar] [CrossRef]
- Piasecka-Srader, J.; Blanco, F.F.; Delman, D.H.; Dixon, D.A.; Geiser, J.L.; Ciereszko, R.E.; Petroff, B.K. Tamoxifen Prevents Apoptosis and Follicle Loss from Cyclophosphamide in Cultured Rat Ovaries1. Biol. Reprod. 2015, 92, 1–8. [Google Scholar] [CrossRef]
- Kalich-Philosoph, L.; Roness, H.; Carmely, A.; Fishel-Bartal, M.; Ligumsky, H.; Paglin, S.; Wolf, I.; Kanety, H.; Sredni, B.; Meirow, D. Cyclophosphamide Triggers Follicle Activation and “Burnout”; AS101 Prevents Follicle Loss and Preserves Fertility. Sci. Transl. Med. 2013, 5, 185ra62. [Google Scholar] [CrossRef] [PubMed]
- Tang, D.; Feng, X.; Ling, L.; Zhang, W.; Luo, Y.; Wang, Y.; Xiong, Z. Experimental Study for the Establishment of a Chemotherapy-Induced Ovarian Insufficiency Model in Rats by Using Cyclophosphamide Combined with Busulfan. Regul. Toxicol. Pharmacol. 2021, 122, 104915. [Google Scholar] [CrossRef] [PubMed]
- Abogresha, N.M.; Mohammed, S.S.; Hosny, M.M.; Abdallah, H.Y.; Gadallah, A.M.; Greish, S.M. Diosmin Mitigates Cyclophosphamide Induced Premature Ovarian Insufficiency in Rat Model. Int. J. Mol. Sci. 2021, 22, 3044. [Google Scholar] [CrossRef]
- Du, K.; Tsichlis, P.N. Regulation of the Akt Kinase by Interacting Proteins. Oncogene 2005, 24, 7401–7409. [Google Scholar] [CrossRef]
- Otto, A.M.; Faderl, M.; Schönenberger, H. Dissociation of Estrogenic and Cytotoxic Properties of an Estrogen Receptor-Binding Platinum Complex in Human Breast Cancer Cell Lines. Cancer Res. 1991, 51, 3217–3223. [Google Scholar]
- Chinwe, G.S.; Azuka, O.I.; Adaeze, N.C. Resveratrol Supplementation Rescues Pool of Growing Follicles and Ovarian Stroma from Cisplatin-Induced Toxicity on the Ovary in Sprague-Dawley Rats: An Experimental Study. Int. J. Reprod. Biomed. 2018, 16, 19–30. [Google Scholar]
- Amin, A.; Hamza, A.A. Effects of Roselle and Ginger on Cisplatin-Induced Reproductive Toxicity in Rats. Asian J. Androl. 2006, 8, 607–612. [Google Scholar] [CrossRef]
- Fouad, A.A.; Albuali, W.H.; Zahran, A.; Gomaa, W. Protective Effect of Naringenin against Gentamicin-Induced Nephrotoxicity in Rats. Environ. Toxicol. Pharmacol. 2014, 38, 420–429. [Google Scholar] [CrossRef]
- Rahman, S.; Islam, R. Mammalian Sirt1: Insights on Its Biological Functions. Cell Commun. Signal 2011, 9, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Jang, M.; Cai, L.; Udeani, G.O.; Slowing, K.V.; Thomas, C.F.; Beecher, C.W.; Fong, H.H.; Farnsworth, N.R.; Kinghorn, A.D.; Mehta, R.G. Cancer Chemopreventive Activity of Resveratrol, a Natural Product Derived from Grapes. Science 1997, 275, 218–220. [Google Scholar] [CrossRef]
- Poudel, R.; Stanley, J.L.; Rueda-Clausen, C.F.; Andersson, I.J.; Sibley, C.P.; Davidge, S.T.; Baker, P.N. Effects of Resveratrol in Pregnancy Using Murine Models with Reduced Blood Supply to the Uterus. PLoS ONE 2013, 8, e64401. [Google Scholar] [CrossRef]
- Özcan, P.; Fıçıcıoğlu, C.; Yıldırım, Ö.K.; Özkan, F.; Akkaya, H.; Aslan, İ. Protective Effect of Resveratrol against Oxidative Damage to Ovarian Reserve in Female Sprague–Dawley Rats. Reprod. Biomed. Online 2015, 31, 404–410. [Google Scholar] [CrossRef] [PubMed]
- Ibrahim, M.A.; Albahlol, I.A.; Wani, F.A.; Abd-Eltawab Tammam, A.; Kelleni, M.T.; Sayeed, M.U.; Abd El-Fadeal, N.M.; Mohamed, A.A. Resveratrol Protects against Cisplatin-Induced Ovarian and Uterine Toxicity in Female Rats by Attenuating Oxidative Stress, Inflammation and Apoptosis. Chem.-Biol. Interact. 2021, 338, 109402. [Google Scholar] [CrossRef] [PubMed]
- Poad, B.L.J.; Maccarone, A.T.; Yu, H.; Mitchell, T.W.; Saied, E.M.; Arenz, C.; Hornemann, T.; Bull, J.N.; Bieske, E.J.; Blanksby, S.J. Differential-Mobility Spectrometry of 1-Deoxysphingosine Isomers: New Insights into the Gas Phase Structures of Ionized Lipids. Anal. Chem. 2018, 90, 5343–5351. [Google Scholar] [CrossRef] [PubMed]
- Meirow, D.; Assad, G.; Dor, J.; Rabinovici, J. The GnRH Antagonist Cetrorelix Reduces Cyclophosphamide-Induced Ovarian Follicular Destruction in Mice. Hum. Reprod. 2004, 19, 1294–1299. [Google Scholar] [CrossRef]
- Mobasher, M.A.; Valverde, Á.M. Signalling Pathways Involved in Paracetamol-Induced Hepatotoxicity: New Insights on the Role of Protein Tyrosine Phosphatase 1B. Arch. Physiol. Biochem. 2014, 120, 51–63. [Google Scholar] [CrossRef]
- Wang, S.; Sun, M.; Yu, L.; Wang, Y.; Yao, Y.; Wang, D. Niacin Inhibits Apoptosis and Rescues Premature Ovarian Failure. Cell. Physiol. Biochem. 2018, 50, 2060–2070. [Google Scholar] [CrossRef]
- Devine, P.J.; Perreault, S.D.; Luderer, U. Roles of Reactive Oxygen Species and Antioxidants in Ovarian Toxicity. Biol Reprod 2012, 86, 1–10. [Google Scholar] [CrossRef]
- Khirallah, S.M.; Ramadan, H.M.M.; Aladl, H.A.A.; Ayaz, N.O.; Kurdi, L.A.F.; Jaremko, M.; Alshawwa, S.Z.; Saied, E.M. Antidiabetic Potential of Novel 1,3,5-Trisubstituted-2-Thioxoimidazloidin-4-One Analogues: Insights into α-Glucosidase, α-Amylase, and Antioxidant Activities. Pharmaceuticals 2022, 15, 1576. [Google Scholar] [CrossRef]
- Peters, A.E.; Mihalas, B.P.; Bromfield, E.G.; Roman, S.D.; Nixon, B.; Sutherland, J.M. Autophagy in Female Fertility: A Role in Oxidative Stress and Aging. Antioxid. Redox Signal. 2020, 32, 550–568. [Google Scholar] [CrossRef]
- Sohel, M.M.H.; Akyuz, B.; Konca, Y.; Arslan, K.; Sariozkan, S.; Cinar, M.U. Oxidative Stress Modulates the Expression of Apoptosis-Associated MicroRNAs in Bovine Granulosa Cells in Vitro. Cell Tissue Res. 2019, 376, 295–308. [Google Scholar] [CrossRef]
- Maclaran, K.; Nikolaou, D. Early Ovarian Ageing. Obstet. Gynaecol. 2019, 21, 107–116. [Google Scholar] [CrossRef]
- Samaha, D.; Hamdo, H.H.; Cong, X.; Schumacher, F.; Banhart, S.; Aglar, Ö.; Möller, H.M.; Heuer, D.; Kleuser, B.; Saied, E.M.; et al. Liposomal FRET Assay Identifies Potent Drug-Like Inhibitors of the Ceramide Transport Protein (CERT). Chem. Eur. J. 2020, 26, 16616–16621. [Google Scholar] [CrossRef]
- Yang, H.S.; Han, D.K.; Kim, J.R.; Sim, J.C. Effects of α-Tocopherol on Cadmium-Induced Toxicity in Rat Testis and Spermatogenesis. J. Korean Med. Sci. 2006, 21, 445–451. [Google Scholar] [CrossRef]
- García Beltrán, J.M.; Espinosa, C.; Guardiola, F.A.; Esteban, M.Á. Dietary Dehydrated Lemon Peel Improves the Immune but Not the Antioxidant Status of Gilthead Seabream (Sparus aurata L.). Fish Shellfish. Immunol. 2017, 64, 426–436. [Google Scholar] [CrossRef]
- Zhuo, L.-C.; Yong, A.S.K.; Shapawi, R.; Lin, Y.-H. Effects of Fermented Lemon Peel Supplementation in Diet on Growth, Immune Responses, and Intestinal Morphology of Asian Sea Bass, Lates Calcarifer. Aquacult. Rep. 2021, 21, 100801. [Google Scholar] [CrossRef]
- Arwa, P.S.; Zeraik, M.L.; Ximenes, V.F.; da Fonseca, L.M.; da Silva Bolzani, V.; Silva, D.H.S. Redox-Active Biflavonoids from Garcinia Brasiliensis as Inhibitors of Neutrophil Oxidative Burst and Human Erythrocyte Membrane Damage. J. Ethnopharmacol. 2015, 174, 410–418. [Google Scholar] [CrossRef]
- Presentato, A.; Scurria, A.; Albanese, L.; Lino, C.; Sciortino, M.; Pagliaro, M.; Zabini, F.; Meneguzzo, F.; Alduina, R.; Nuzzo, D.; et al. Superior Antibacterial Activity of Integral Lemon Pectin Extracted via Hydrodynamic Cavitation. ChemistryOpen 2020, 9, 628–630. [Google Scholar] [CrossRef]
- Bradamante, S.; Barenghi, L.; Villa, A. Cardiovascular Protective Effects of Resveratrol. Cardiovasc. Drug Rev. 2004, 22, 169–188. [Google Scholar] [CrossRef]
- Aryal, S.; Baniya, M.K.; Danekhu, K.; Kunwar, P.; Gurung, R.; Koirala, N. Total Phenolic Content, Flavonoid Content and Antioxidant Potential of Wild Vegetables from Western Nepal. Plants 2019, 8, 96. [Google Scholar] [CrossRef] [PubMed]
- Xia, N.; Daiber, A.; Habermeier, A.; Closs, E.I.; Thum, T.; Spanier, G.; Lu, Q.; Oelze, M.; Torzewski, M.; Lackner, K.J. Resveratrol Reverses Endothelial Nitric-Oxide Synthase Uncoupling in Apolipoprotein E Knockout Mice. J. Pharmacol. Exp. Ther. 2010, 335, 149–154. [Google Scholar] [CrossRef] [PubMed]
- Kong, X.-X.; Fu, Y.-C.; Xu, J.-J.; Zhuang, X.-L.; Chen, Z.-G.; Luo, L.-L. Resveratrol, an Effective Regulator of Ovarian Development and Oocyte Apoptosis. J. Endocrinol. Investig. 2011, 34, e374–e381. [Google Scholar] [CrossRef]
- Ata, N.; Kulhan, N.; Kulhan, M.; Türkler, C.; Kiremitli, T.; Kiremitli, S.; Cimen, F.; Süleyman, H.; Toprak, V. The Effect of Resveratrol on Oxidative Ovary-Damage Induced by Methotrexate in Rats (Resveratrol Oxidative Ovary-Damage). Gen. Physiol. Biophys. 2019, 38, 519–524. [Google Scholar] [CrossRef]
- Ozatik, F.Y.; Ozatik, O.; Yigitaslan, S.; Kaygısız, B.; Erol, K. Do Resveratrol and Dehydroepiandrosterone Increase Diminished Ovarian Reserve? Eurasian J. Med. 2020, 52, 6–11. [Google Scholar] [CrossRef]
- Lai, J.; Ge, M.; Shen, S.; Yang, L.; Jin, T.; Cao, D.; Xu, H.; Zheng, X.; Qiu, S.; Wang, K.; et al. Activation of NFKB-JMJD3 Signaling Promotes Bladder Fibrosis via Boosting Bladder Smooth Muscle Cell Proliferation and Collagen Accumulation. Biochim. Biophys. Acta (BBA)-Mol. Basis Dis. 2019, 1865, 2403–2410. [Google Scholar] [CrossRef]
- Galal, S.M.; Mansour, H.H.; Elkhoely, A.A. Diallyl Sulfide Alleviates Cyclophosphamide-Induced Nephropathic Encephalopathy in Rats. Toxicol. Mech. Methods 2020, 30, 208–218. [Google Scholar] [CrossRef]
- He, K.; Gkioxari, G.; Dollár, P.; Girshick, R. Mask R-CNN. In Proceedings of the 2017 IEEE International Conference on Computer Vision (ICCV), Venice, Italy, 22–29 October 2017; pp. 2980–2988. [Google Scholar]
- Iqbal, S.; Hayman, E.G.; Hong, C.; Stokum, J.A.; Kurland, D.B.; Gerzanich, V.; Simard, J.M. Inducible Nitric Oxide Synthase (NOS-2) in Subarachnoid Hemorrhage: Regulatory Mechanisms and Therapeutic Implications. Brain Circ. 2016, 2, 8–19. [Google Scholar] [CrossRef]
- El-Naggar, S.A.; El-Said, K.S.; Mobasher, M.; Elbakry, M. Enhancing Antitumor Efficacy of Cisplatin Low Dose by EDTA in Ehrlich Ascetic Carcinoma Bearing Mice. Braz. Arch. Biol. Technol. 2019, 62, e19180716. [Google Scholar] [CrossRef]
- Bai, Y.; Jiang, Y.; Liu, T.; Li, F.; Zhang, J.; Luo, Y.; Zhang, L.; Yan, G.; Feng, Z.; Li, X.; et al. Xinjiang Herbal Tea Exerts Immunomodulatory Activity via TLR2/4-Mediated MAPK Signaling Pathways in RAW264.7 Cells and Prevents Cyclophosphamide-Induced Immunosuppression in Mice. J. Ethnopharmacol. 2019, 228, 179–187. [Google Scholar] [CrossRef] [PubMed]
- Kwon, D.-J.; Ju, S.M.; Youn, G.S.; Choi, S.Y.; Park, J. Suppression of INOS and COX-2 Expression by Flavokawain A via Blockade of NF-ΚB and AP-1 Activation in RAW 264.7 Macrophages. Food Chem. Toxicol. 2013, 58, 479–486. [Google Scholar] [CrossRef] [PubMed]
- Ding, H.; Chen, J.; Su, M.; Lin, Z.; Zhan, H.; Yang, F.; Li, W.; Xie, J.; Huang, Y.; Liu, X.; et al. BDNF Promotes Activation of Astrocytes and Microglia Contributing to Neuroinflammation and Mechanical Allodynia in Cyclophosphamide-Induced Cystitis. J. Neuroinflamm. 2020, 17, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Noh, E.-M.; Kim, J.-M.; Lee, H.Y.; Song, H.-K.; Joung, S.O.; Yang, H.J.; Kim, M.J.; Kim, K.S.; Lee, Y.-R. Immuno-Enhancement Effects of Platycodon Grandiflorum Extracts in Splenocytes and a Cyclophosphamide-Induced Immunosuppressed Rat Model. BMC Complement. Altern. Med. 2019, 19, 322. [Google Scholar] [CrossRef] [PubMed]
- Cengiz, M.; Sahinturk, V.; Yildiz, S.C.; Şahin, İ.K.; Bilici, N.; Yaman, S.O.; Altuner, Y.; Appak-Baskoy, S.; Ayhanci, A. Cyclophosphamide Induced Oxidative Stress, Lipid per Oxidation, Apoptosis and Histopathological Changes in Rats: Protective Role of Boron. J. Trace Elem. Med. Biol. 2020, 62, 126574. [Google Scholar] [CrossRef] [PubMed]
- Mobasher, M.A.; Germoush, M.O.; Galal El-Tantawi, H.; Samy El-Said, K. Metformin Improves Biochemical and Pathophysiological Changes in Hepatocellular Carcinoma with Pre-Existed Diabetes Mellitus Rats. Pathogens 2021, 10, 59. [Google Scholar] [CrossRef]
- Said, R.S.; Mantawy, E.M.; El-Demerdash, E. Mechanistic Perspective of Protective Effects of Resveratrol against Cisplatin-Induced Ovarian Injury in Rats: Emphasis on Anti-Inflammatory and Anti-Apoptotic Effects. Naunyn-Schmiedeberg’s Arch. Pharmacol. 2019, 392, 1225–1238. [Google Scholar] [CrossRef]
- Vyas, D.; Laput, G.; Vyas, A.K. Chemotherapy-Enhanced Inflammation May Lead to the Failure of Therapy and Metastasis. Onco Targets Ther. 2014, 7, 1015–1023. [Google Scholar] [CrossRef]
- Domitrovic, R. The Molecular Basis for the Pharmacological Activity of Anthocyans. CMC 2011, 18, 4454–4469. [Google Scholar] [CrossRef]
- El Azab, I.H.; Saied, E.M.; Osman, A.A.; Mehana, A.E.; Saad, H.A.; Elkanzi, N.A. Novel N-Bridged Pyrazole-1-Carbothioamides with Potential Antiproliferative Activity: Design, Synthesis, in Vitro and in Silico Studies. Future Med. Chem. 2021, 13, 1743–1766. [Google Scholar] [CrossRef]
- Khirallah, S.M.; Ramadan, H.M.M.; Shawky, A.; Qahl, S.H.; Baty, R.S.; Alqadri, N.; Alsuhaibani, A.M.; Jaremko, M.; Emwas, A.-H.; Saied, E.M. Development of Novel 1,3-Disubstituted-2-Thiohydantoin Analogues with Potent Anti-Inflammatory Activity; In Vitro and In Silico Assessments. Molecules 2022, 27, 6271. [Google Scholar] [CrossRef] [PubMed]
- Khan, M.K.; Zill-E-Huma; Dangles, O. A Comprehensive Review on Flavanones, the Major Citrus Polyphenols. J. Food Compos. Anal. 2014, 33, 85–104. [Google Scholar] [CrossRef]
- Sridharan, B.; Mehra, Y.; Ganesh, R.N.; Viswanathan, P. Regulation of Urinary Crystal Inhibiting Proteins and Inflammatory Genes by Lemon Peel Extract and Formulated Citrus Bioflavonoids on Ethylene Glycol Induced Urolithic Rats. Food Chem. Toxicol. 2016, 94, 75–84. [Google Scholar] [CrossRef] [PubMed]
- Pantsulaia, I.; Iobadze, M.; Pantsulaia, N.; Chikovani, T. The Effect of Citrus Peel Extracts on Cytokines Levels and T Regulatory Cells in Acute Liver Injury. BioMed Res. Int. 2014, 2014, 127879. [Google Scholar] [CrossRef] [PubMed]
- Bin-Jumah, M.; Abdel-Fattah, A.-F.M.; Saied, E.M.; El-Seedi, H.R.; Abdel-Daim, M.M. Acrylamide-Induced Peripheral Neuropathy: Manifestations, Mechanisms, and Potential Treatment Modalities. Environ. Sci. Pollut. Res. 2021, 28, 13031–13046. [Google Scholar] [CrossRef]
- Kim, H.-K.; Lee, G.-H.; Bhattarai, K.R.; Junjappa, R.P.; Lee, H.-Y.; Handigund, M.; Marahatta, A.; Bhandary, B.; Baek, I.-H.; Pyo, J.S.; et al. PI3Kδ Contributes to ER Stress-Associated Asthma through ER-Redox Disturbances: The Involvement of the RIDD–RIG-I–NF-ΚB Axis. Exp. Mol. Med. 2018, 50, e444. [Google Scholar] [CrossRef]
- McKinnon, B.; Bersinger, N.A.; Huber, A.W.; Kuhn, A.; Mueller, M.D. PPAR-γ Expression in Peritoneal Endometriotic Lesions Correlates with Pain Experienced by Patients. Fertil. Steril. 2010, 93, 293–296. [Google Scholar] [CrossRef]
- Rada, P.; Pardo, V.; Mobasher, M.A.; García-Martínez, I.; Ruiz, L.; Gonzalez-Rodriguez, A.; Sanchez-Ramos, C.; Muntané, J.; Alemany, S.; James, L.P.; et al. SIRT1 controls acetaminophen hepatotoxicity by modulating inflammation and oxidative stress. Antioxid. Redox Signal. 2018, 28, 1187–1208. [Google Scholar] [CrossRef]
- Salminen, A.; Kauppinen, A.; Suuronen, T.; Kaarniranta, K. SIRT1 longevity factor suppresses NF-κB -driven immune responses: Regulation of aging via NF-κB acetylation? BioEssays 2008, 30, 939–942. [Google Scholar] [CrossRef]
- Khodarahmian, M.; Amidi, F.; Moini, A.; Kashani, L.; Salahi, E.; Danaii-mehrabad, S.; Nashtaei, M.S.; Mojtahedi, M.F.; Esfandyari, S.; Sobhani, A. A Randomized Exploratory Trial to Assess the Effects of Resveratrol on VEGF and TNF-α 2 Expression in Endometriosis Women. J. Reprod. Immunol. 2021, 143, 103248. [Google Scholar] [CrossRef]
- Saied, E.M.; Arenz, C. Small Molecule Inhibitors of Ceramidases. CPB 2014, 34, 197–212. [Google Scholar] [CrossRef] [PubMed]
- Abdel-Wahab, B.A.; Abd El-Kareem, F.; Alzamami, H.; Fahmy, A.; Elesawy, B.; Mostafa Mahmoud, M.; Ghareeb, A.; El Askary, A.; Abo Nahas, H.; Attallah, N.; et al. Novel Exopolysaccharide from Marine Bacillus Subtilis with Broad Potential Biological Activities: Insights into Antioxidant, Anti-Inflammatory, Cytotoxicity, and Anti-Alzheimer Activity. Metabolites 2022, 12, 715. [Google Scholar] [CrossRef]
- Meng, T.; Xiao, D.; Muhammed, A.; Deng, J.; Chen, L.; He, J. Anti-Inflammatory Action and Mechanisms of Resveratrol. Molecules 2021, 26, 229. [Google Scholar] [CrossRef] [PubMed]
- Wiciński, M.; Socha, M.; Walczak, M.; Wódkiewicz, E.; Malinowski, B.; Rewerski, S.; Górski, K.; Pawlak-Osińska, K. Beneficial Effects of Resveratrol Administration—Focus on Potential Biochemical Mechanisms in Cardiovascular Conditions. Nutrients 2018, 10, 1813. [Google Scholar] [CrossRef] [PubMed]
- Ezoe, K.; Daikoku, T.; Yabuuchi, A.; Murata, N.; Kawano, H.; Abe, T.; Okuno, T.; Kobayashi, T.; Kato, K. Ovarian Stimulation Using Human Chorionic Gonadotrophin Impairs Blastocyst Implantation and Decidualization by Altering Ovarian Hormone Levels and Downstream Signaling in Mice. Mol. Hum. Reprod. 2014, 20, 1101–1116. [Google Scholar] [CrossRef]
- Sonigo, C.; Beau, I.; Grynberg, M.; Binart, N. AMH Prevents Primordial Ovarian Follicle Loss and Fertility Alteration in Cyclophosphamide-Treated Mice. FASEB J. 2019, 33, 1278–1287. [Google Scholar] [CrossRef]
- Çil, N.; Mete, G.A. The Effect of Adipose-Derived Mesenchymal Stem Cell Treatment on MTOR and p-MTOR Expression in Ovarian Damage Due to Cyclophosphomide. Reprod. Toxicol. 2021, 103, 71–78. [Google Scholar] [CrossRef]
- Nie, Z.; Zhang, L.; Chen, W.; Zhang, Y.; Hua, R.; Wang, W.; Zhang, T.; Wu, H. The Protective Effects of Pretreatment with Resveratrol in Cyclophosphamide-Induced Rat Ovarian Granulosa Cell Injury: In Vitro Study. Reprod. Toxicol. 2020, 95, 66–74. [Google Scholar] [CrossRef]
- Çolak, H.; Karaköse, E. Green Synthesis and Characterization of Nanostructured ZnO Thin Films Using Citrus Aurantifolia (Lemon) Peel Extract by Spin-Coating Method. J. Alloys Compd. 2017, 690, 658–662. [Google Scholar] [CrossRef]
- El-Sheikh, A.A.; Abdelzaher, W.Y.; Gad, A.A.; Abdel-Gaber, S.A. Purine versus Non-Purine Xanthine Oxidase Inhibitors against Cyclophosphamide-Induced Cardiac and Bone Marrow Toxicity in Rats. Hum. Exp. Toxicol. 2020, 39, 249–261. [Google Scholar] [CrossRef]
- Akunna, G.G.; Nwafor, J.; Egwu, O.A.; Ezemagu, U.K.; Obaje, G. Cisplatin-Induced Ovarian Cytotoxicity and the Modulating Role of Aqueous Zest Extract of Citrus Limonium (AZECL) in Rat Models. J. Tradit. Med. Clin. Nat. 2017, 6, 2. [Google Scholar] [CrossRef]
- Siiteri, P.K. Review of Studies on Estrogen Biosynthesis in the Human. Cancer Res. 1982, 42, 3269s–3273s. [Google Scholar] [PubMed]
- Ohkawa, H.; Ohishi, N.; Yagi, K. Assay for Lipid Peroxides in Animal Tissues by Thiobarbituric Acid Reaction. Anal. Biochem. 1979, 95, 351–358. [Google Scholar] [CrossRef] [PubMed]
- Downie, T. Theory and Practice of Histological Techniques Edited by J.D. Bancroft & A. Stevens, Churchill Livingstone, Edinburgh, 740 Pages, £55.00. Histopathology 1990, 17, 386. [Google Scholar] [CrossRef]
- Bloomer, S.A.; Moyer, E.D.; Brown, K.E.; Kregel, K.C. Aging Results in Accumulation of M1 and M2 Hepatic Macrophages and a Differential Response to Gadolinium Chloride. Histochem. Cell Biol. 2020, 153, 37–48. [Google Scholar] [CrossRef] [PubMed]
- Ebiya, R.A.; Montaser, M.M.; Darwish, S.M. Downregulated StAR Gene and Male Reproductive Dysfunction Caused by Nifedipine and Ethosuximide. J. Basic Appl. Zool. 2016, 76, 42–51. [Google Scholar] [CrossRef]
- Montaser, M.; Abiya, R.A.E.; Afifi, M.; Saddick, S.; Allogmani, A.S.; Almaghrabi, O.A. Effect of Natural and Synthetic Food Colorants on Spermatogenesis and the Expression of Its Controlling Genes. Vet. Med. Health Econ. 2018, 55, 187–199. [Google Scholar]
- Hassen, M.T.; Mohamed, H.K.; Montaser, M.M.; El-Sharnouby, M.E.; Awad, N.; Ebiya, R.A. Molecular, Immunomodulatory, and Histopathological Role of Mesenchymal Stem Cells and Beetroot Extract on Cisplatin Induced Testicular Damage in Albino Rats. Animals 2021, 11, 1142. [Google Scholar] [CrossRef]
- Mohamed, D.I.; Alaa El-Din Aly El-Waseef, D.; Nabih, E.S.; El-Kharashi, O.A.; Abd El-Kareem, H.F.; Abo Nahas, H.H.; Abdel-Wahab, B.A.; Helmy, Y.A.; Alshawwa, S.Z.; Saied, E.M. Acetylsalicylic Acid Suppresses Alcoholism-Induced Cognitive Impairment Associated with Atorvastatin Intake by Targeting Cerebral MiRNA155 and NLRP3: In Vivo, and In Silico Study. Pharmaceutics 2022, 14, 529. [Google Scholar] [CrossRef]
- Mohamed, D.I.; Abou-Bakr, D.A.; Ezzat, S.F.; El-Kareem, H.F.A.; Nahas, H.H.A.; Saad, H.A.; Mehana, A.E.; Saied, E.M. Vitamin D3 Prevents the Deleterious Effects of Testicular Torsion on Testis by Targeting MiRNA-145 and ADAM17: In Silico and In Vivo Study. Pharmaceuticals 2021, 14, 1222. [Google Scholar] [CrossRef]
- Mohamed, D.I.; Ezzat, S.F.; Elayat, W.M.; El-Kharashi, O.A.; El-Kareem, H.F.A.; Nahas, H.H.A.; Abdel-Wahab, B.A.; Alshawwa, S.Z.; Saleh, A.; Helmy, Y.A.; et al. Hepatoprotective Role of Carvedilol against Ischemic Hepatitis Associated with Acute Heart Failure via Targeting MiRNA-17 and Mitochondrial Dynamics-Related Proteins: An In Vivo and In Silico Study. Pharmaceuticals 2022, 15, 832. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene Name—Accession No. | Gene | 5-Primer Sequence-3 | Tm |
|---|---|---|---|
| Induced nitric oxide synthase (NM_000625) | iNOS | Fd GCTCTACACCTCCAATGTGACC Rv CTGCCGAGATTTGAGCCTCATG | 60 |
| Caspase-3 (NM_004346.4) | Casp3 | Fd AGGACTCTAGACGGCATCCA Rv CAGTGAGACTTGGTGCAGTGA | 60 |
| Glyceraldehyde-3-phosphatedehydrogenase (NM_002046) | GAPDH | Fd TGGATTTGGACGCATTGGTC Rv TTTGCACTGGTACGTGTTGAT | 56 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mobasher, M.A.; Hassen, M.T.; Ebiya, R.A.; Alturki, N.A.; Alzamami, A.; Mohamed, H.K.; Awad, N.S.; Khodeer, D.; Abd El-Motelp, B.A. Ameliorative Effect of Citrus Lemon Peel Extract and Resveratrol on Premature Ovarian Failure Rat Model: Role of iNOS/Caspase-3 Pathway. Molecules 2023, 28, 122. https://doi.org/10.3390/molecules28010122
Mobasher MA, Hassen MT, Ebiya RA, Alturki NA, Alzamami A, Mohamed HK, Awad NS, Khodeer D, Abd El-Motelp BA. Ameliorative Effect of Citrus Lemon Peel Extract and Resveratrol on Premature Ovarian Failure Rat Model: Role of iNOS/Caspase-3 Pathway. Molecules. 2023; 28(1):122. https://doi.org/10.3390/molecules28010122
Chicago/Turabian StyleMobasher, Maysa A., Marwa T. Hassen, Rasha A. Ebiya, Norah A. Alturki, Ahmad Alzamami, Hanaa K. Mohamed, Nabil S. Awad, Dina Khodeer, and Bosy A. Abd El-Motelp. 2023. "Ameliorative Effect of Citrus Lemon Peel Extract and Resveratrol on Premature Ovarian Failure Rat Model: Role of iNOS/Caspase-3 Pathway" Molecules 28, no. 1: 122. https://doi.org/10.3390/molecules28010122
APA StyleMobasher, M. A., Hassen, M. T., Ebiya, R. A., Alturki, N. A., Alzamami, A., Mohamed, H. K., Awad, N. S., Khodeer, D., & Abd El-Motelp, B. A. (2023). Ameliorative Effect of Citrus Lemon Peel Extract and Resveratrol on Premature Ovarian Failure Rat Model: Role of iNOS/Caspase-3 Pathway. Molecules, 28(1), 122. https://doi.org/10.3390/molecules28010122