
Cannabis sativa CBD Extract Shows Promising Antibacterial Activity against Salmonella typhimurium and S. newington

 , ,
, , {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
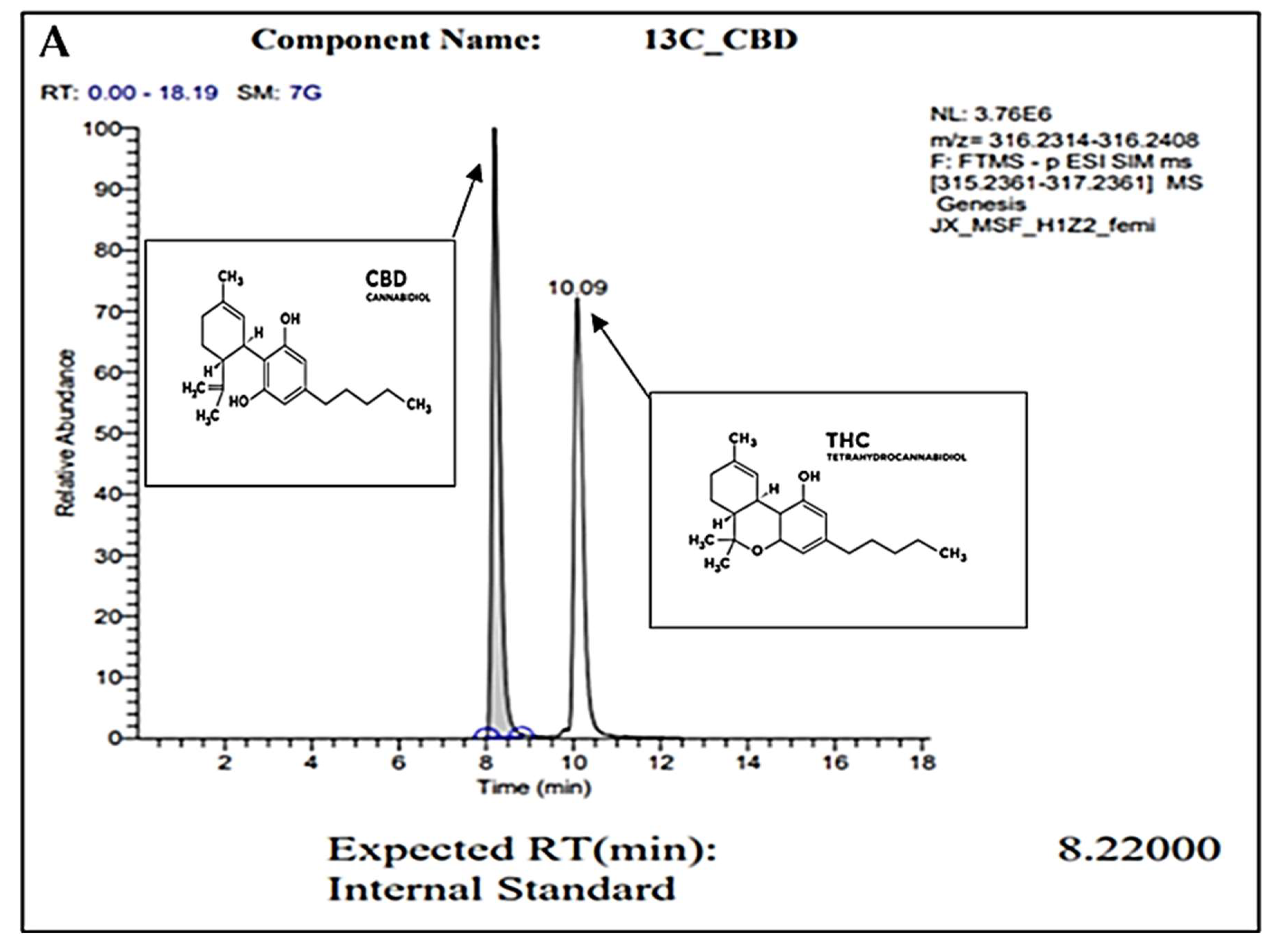
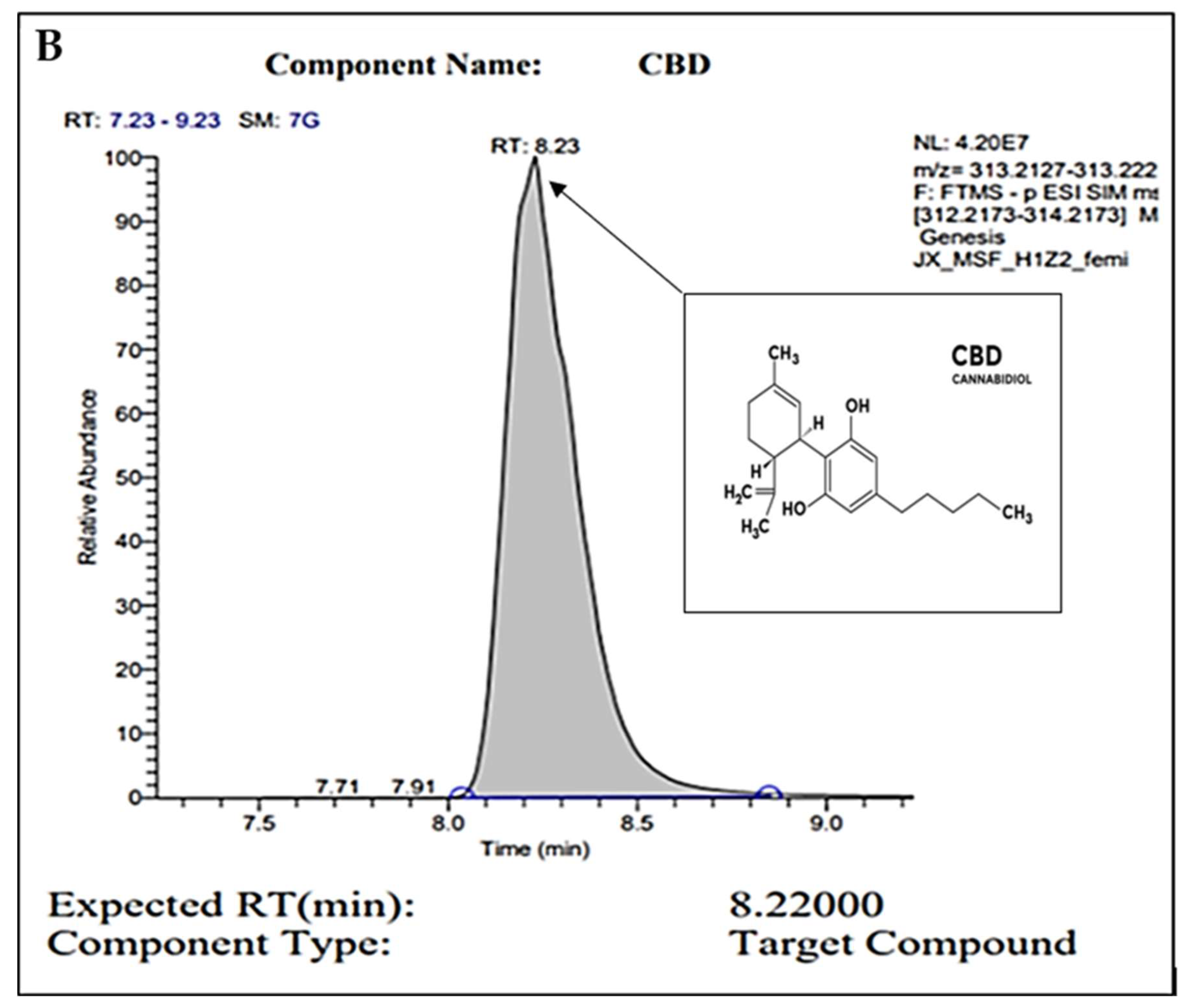
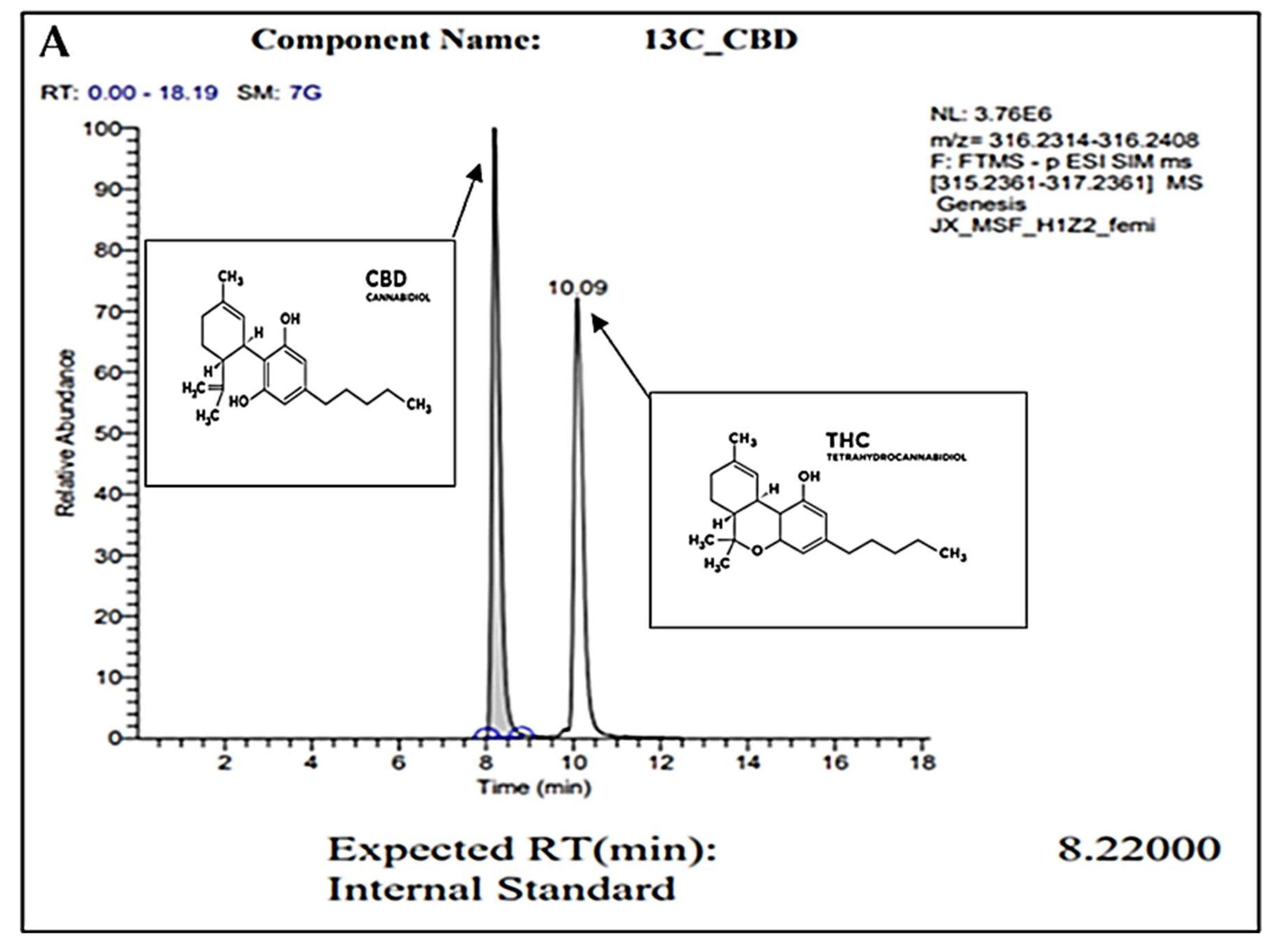
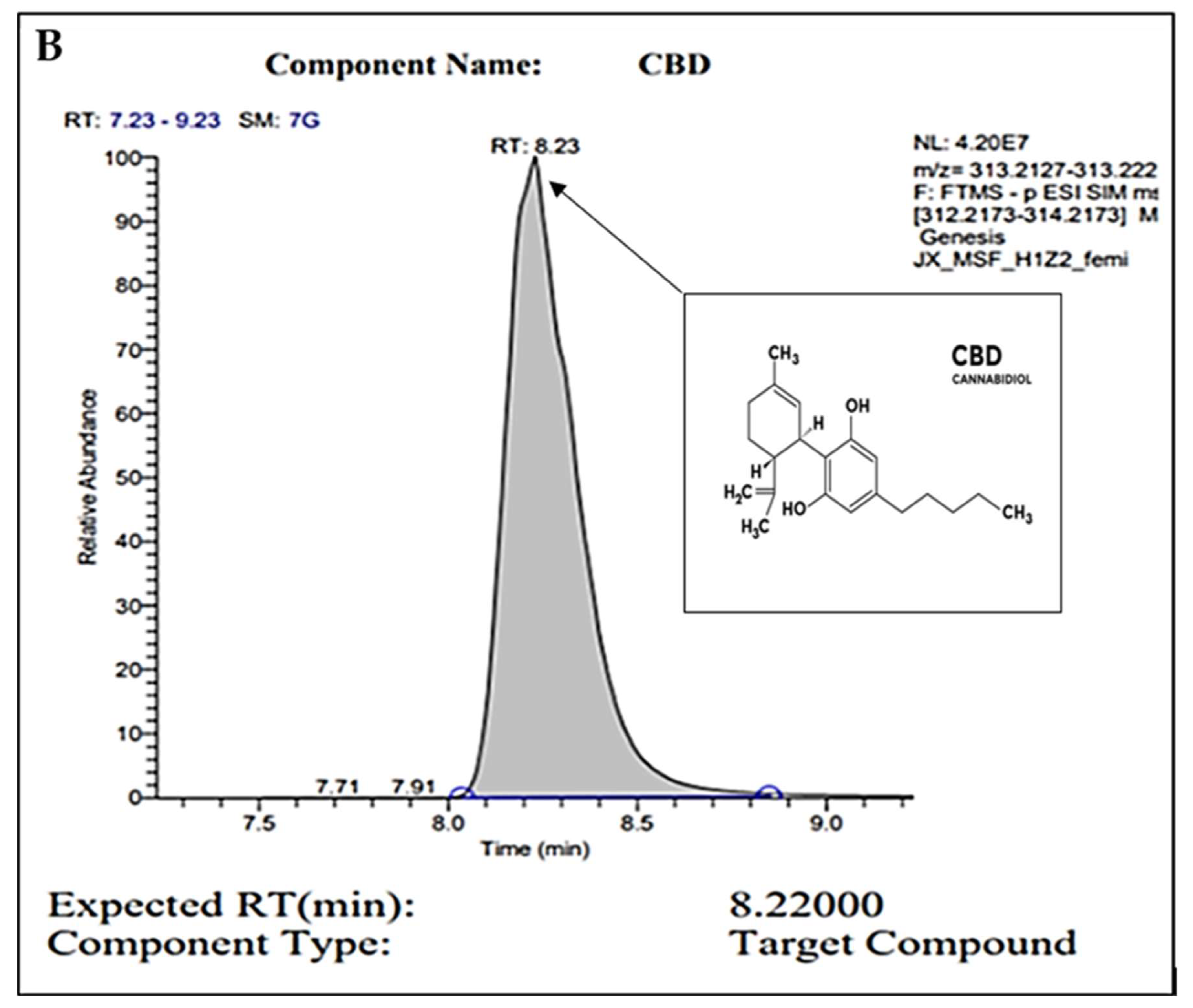
2.1. Gas Chromatography
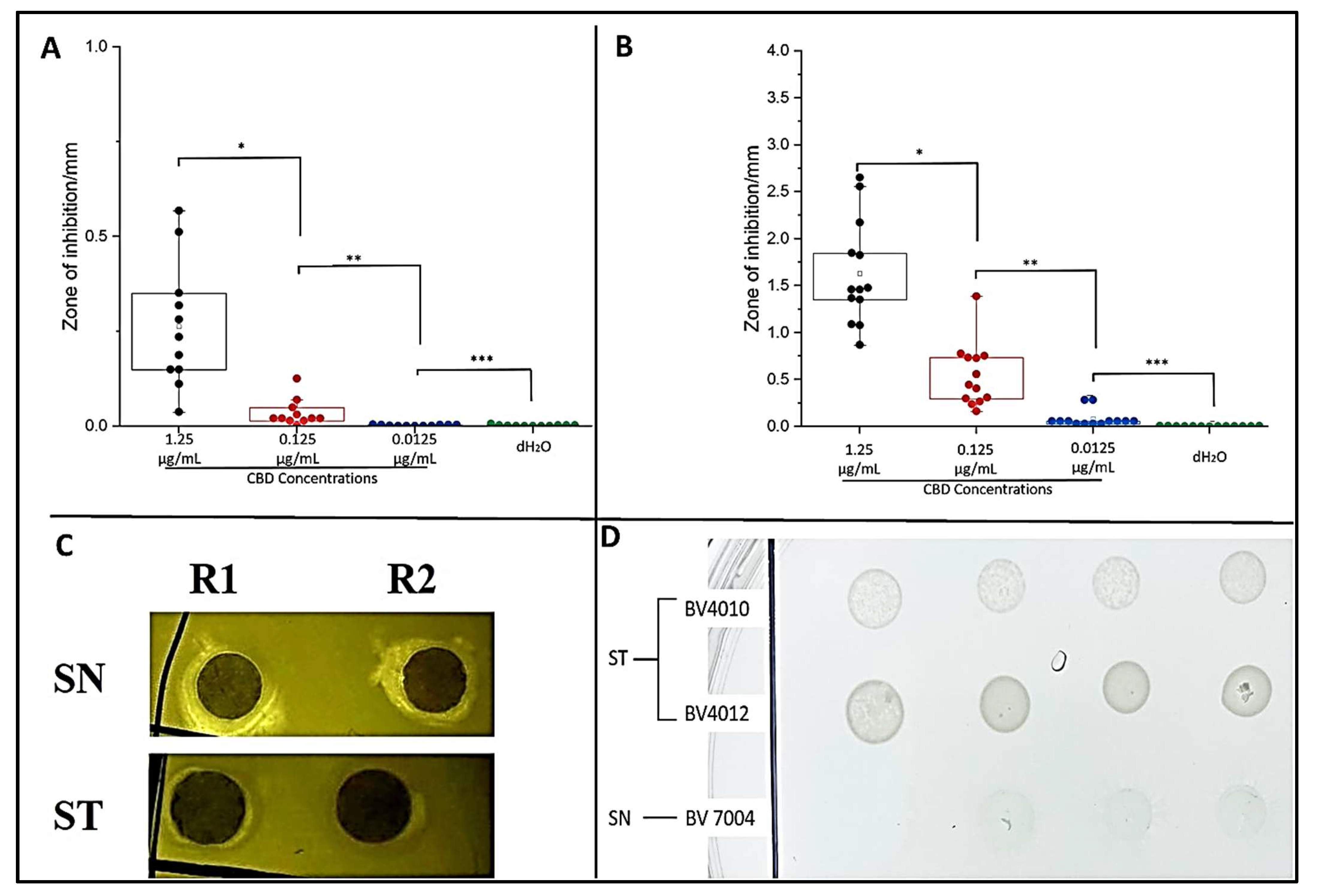
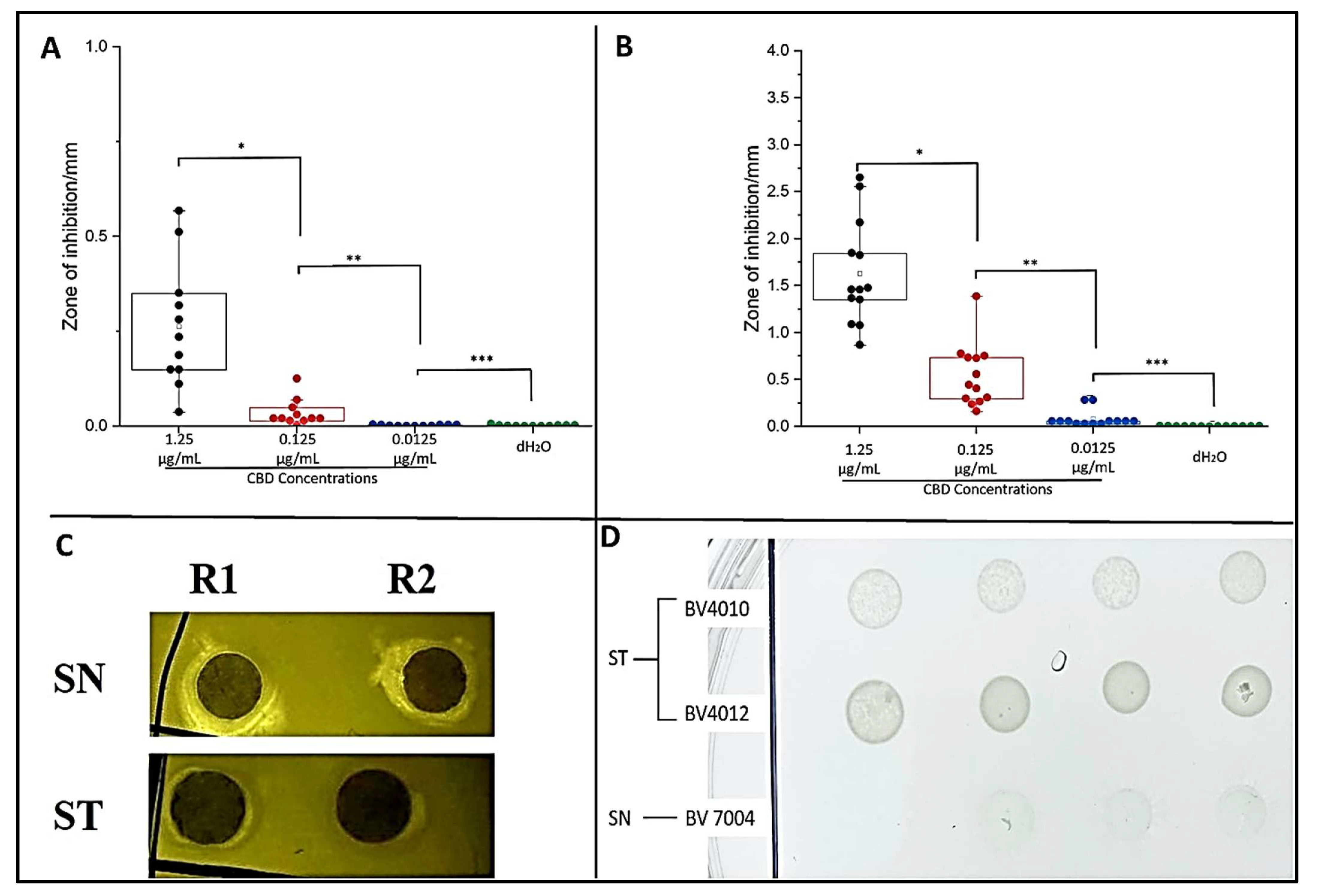
2.2. Plate Assays
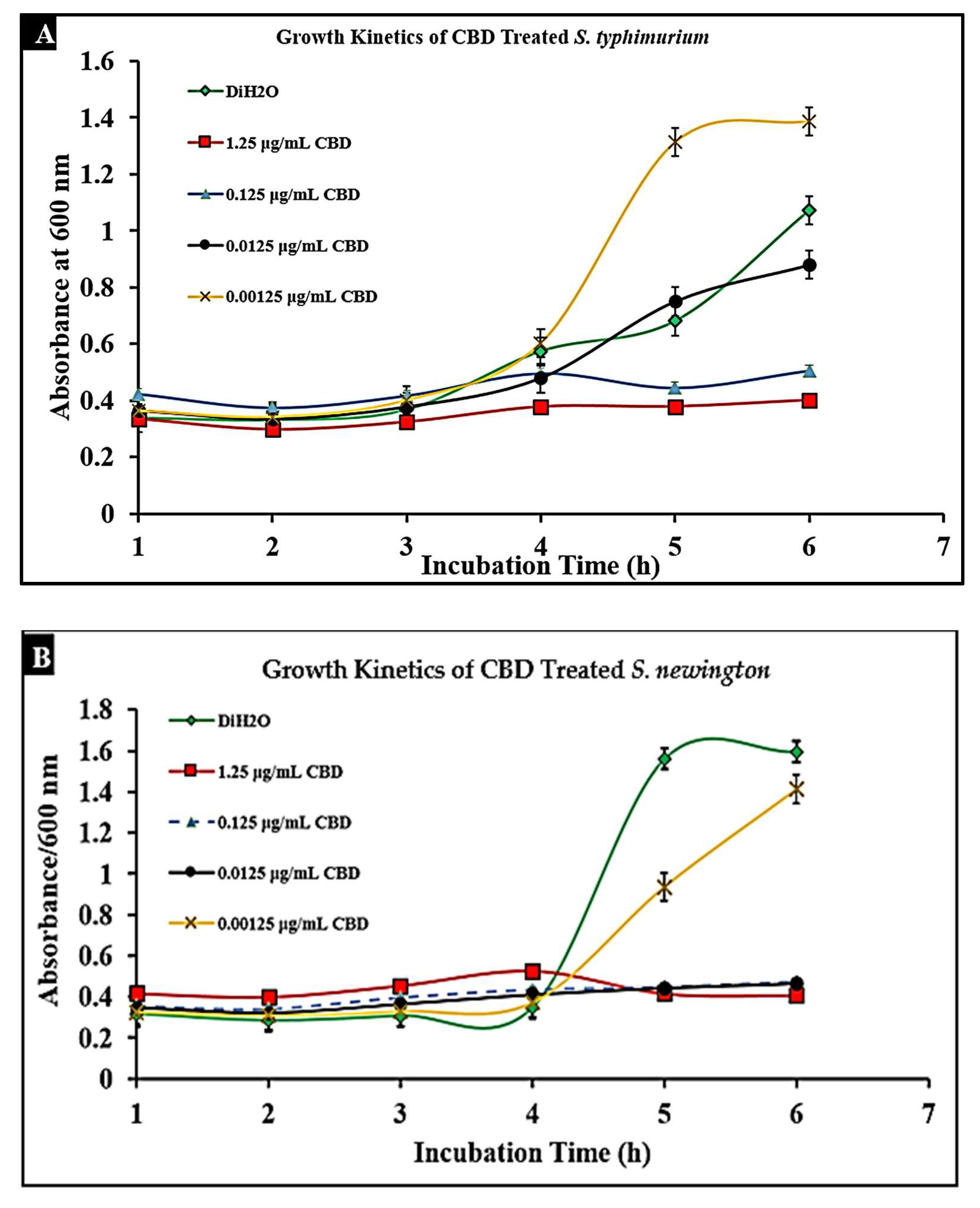
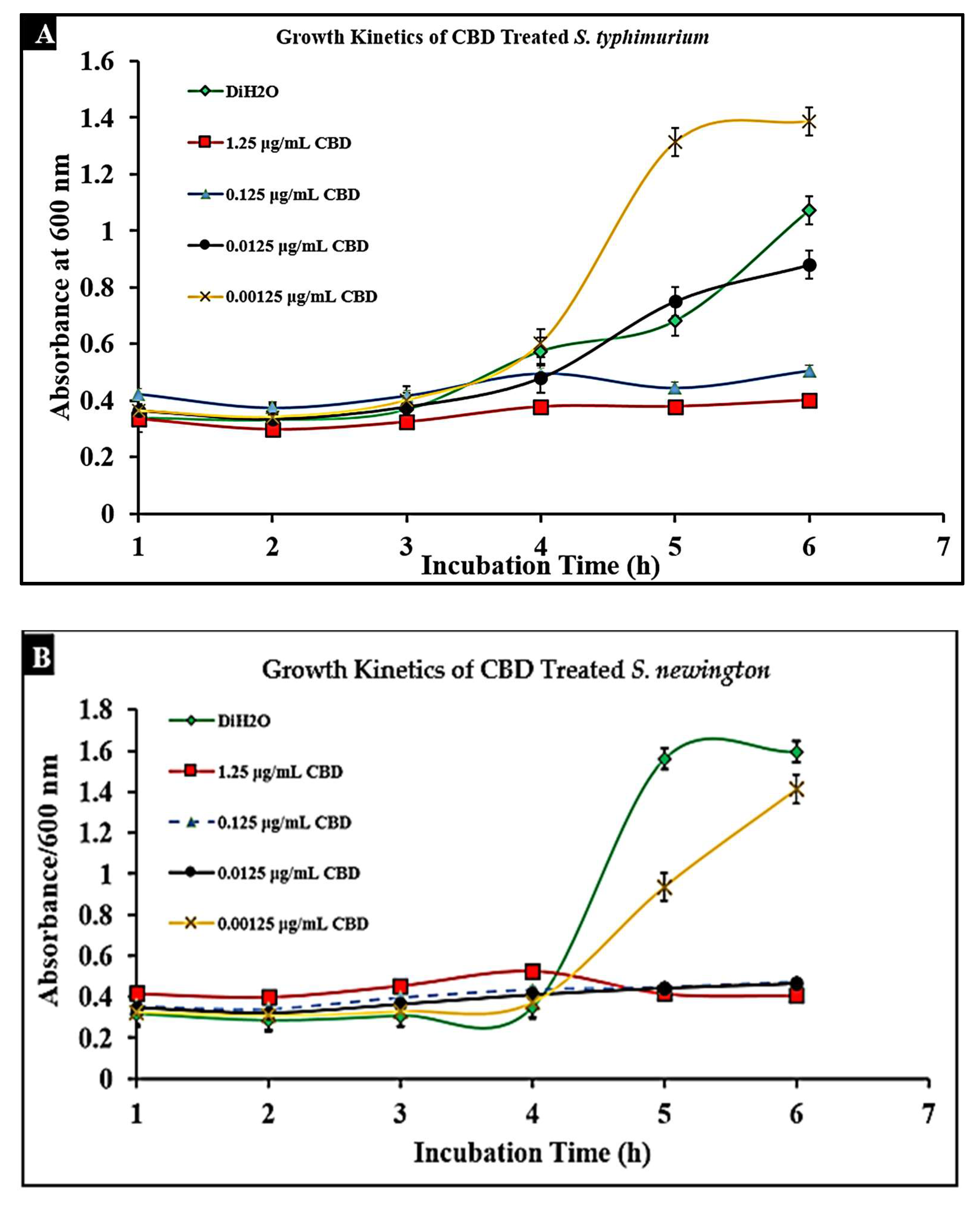
2.3. CBD Extract Reduces Bacterial Growth of Salmonella typhimurium and Salmonella newington
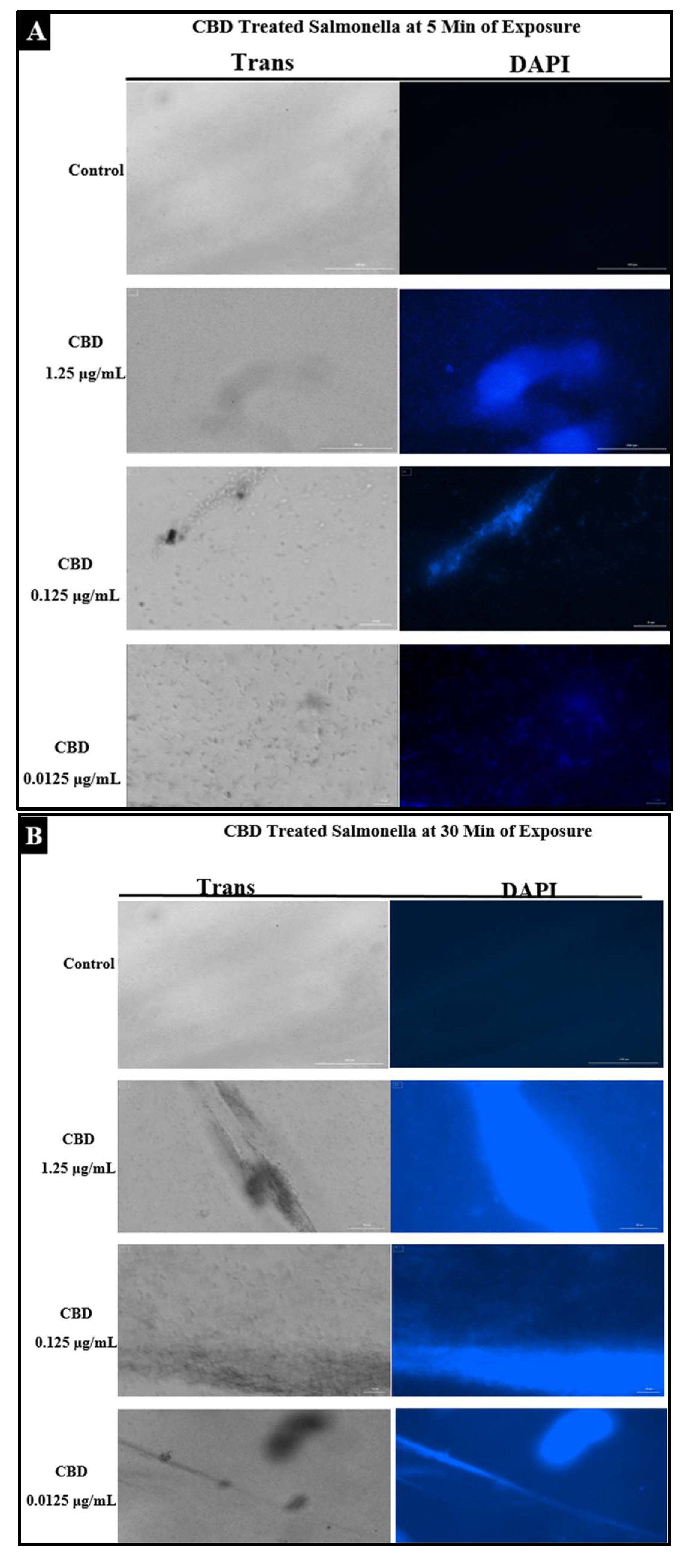
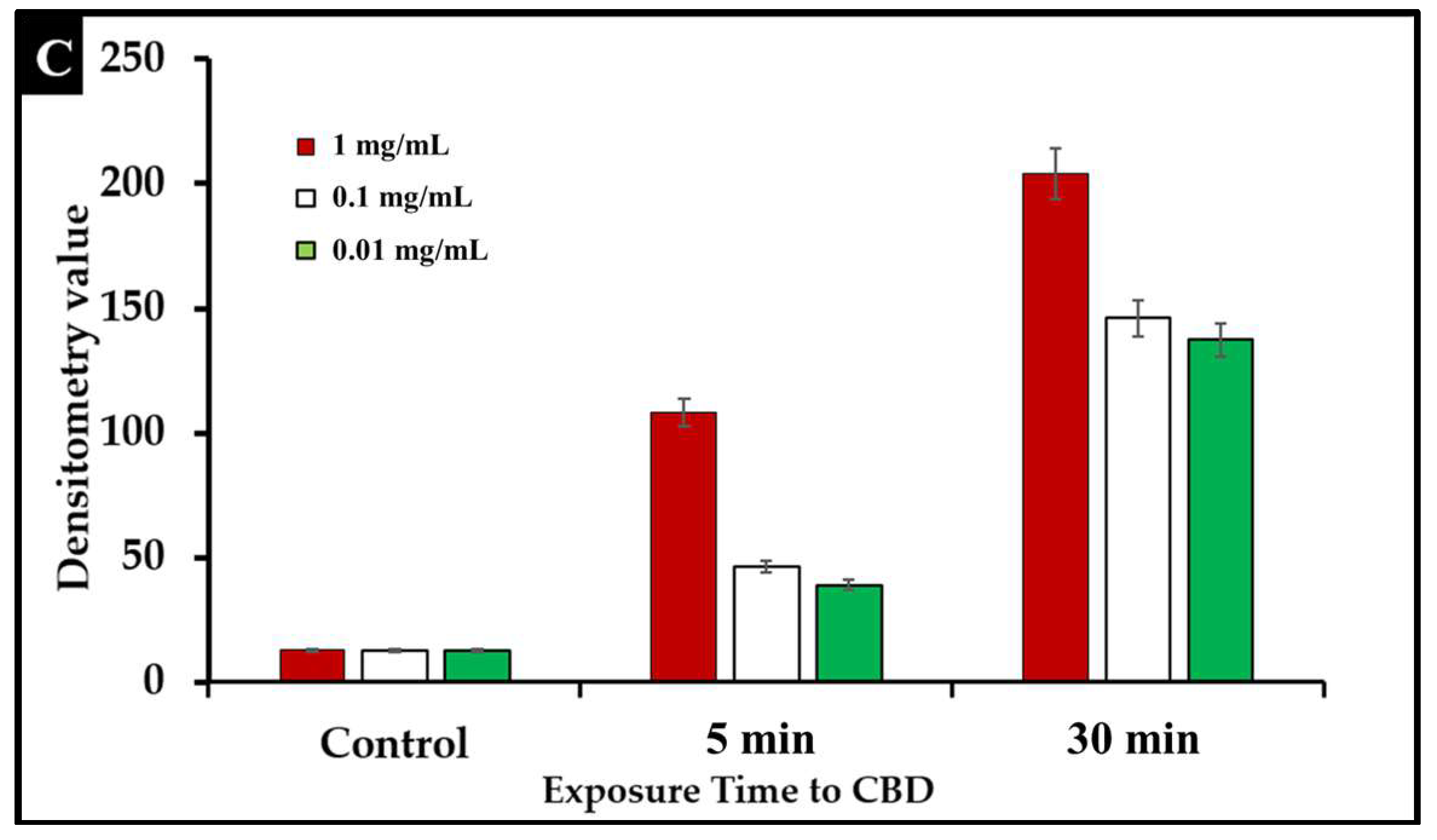
2.4. Assessment of Bacterial Membrane Integrity via Fluorescent Staining of CBD-Treated Salmonella Cells
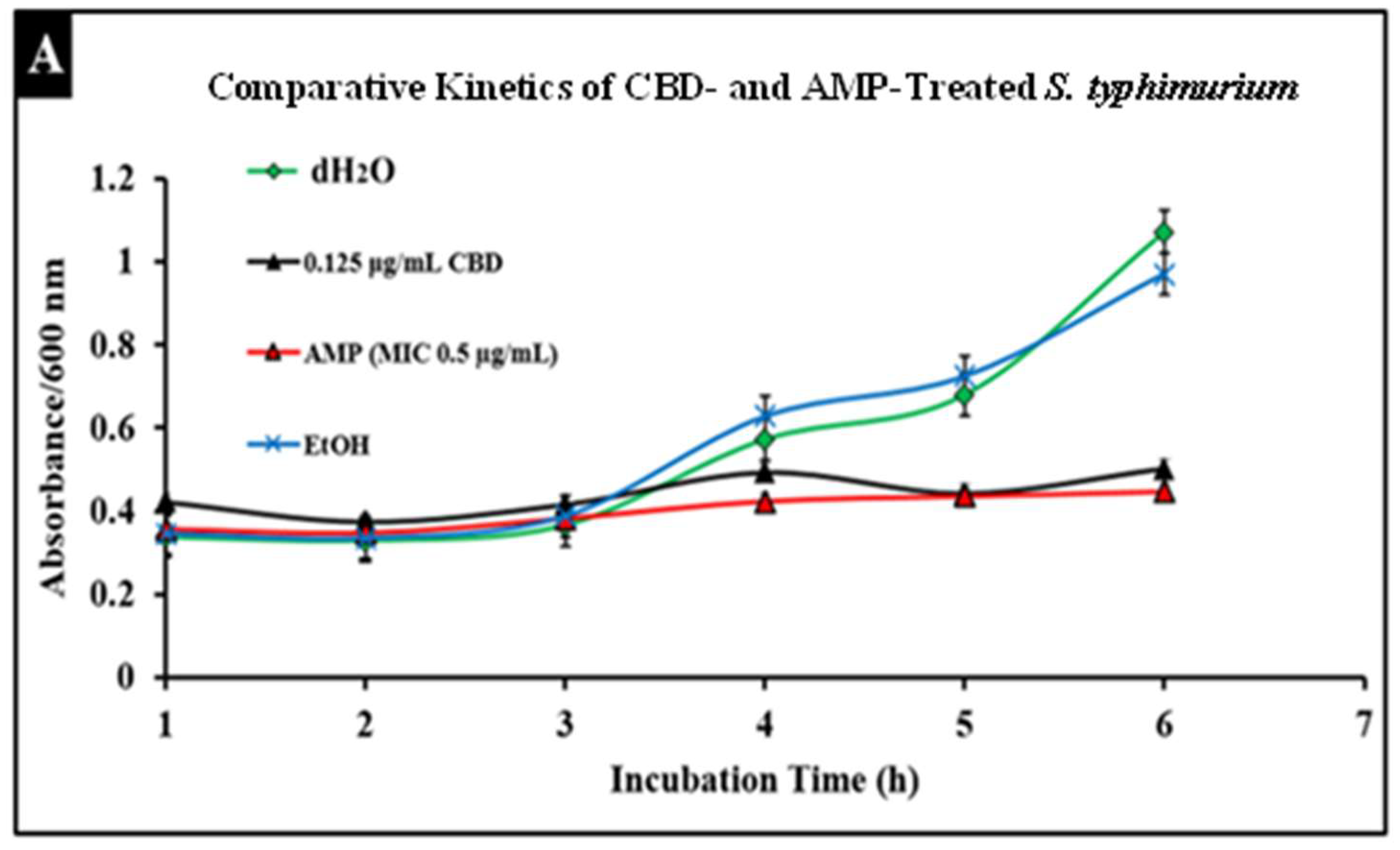
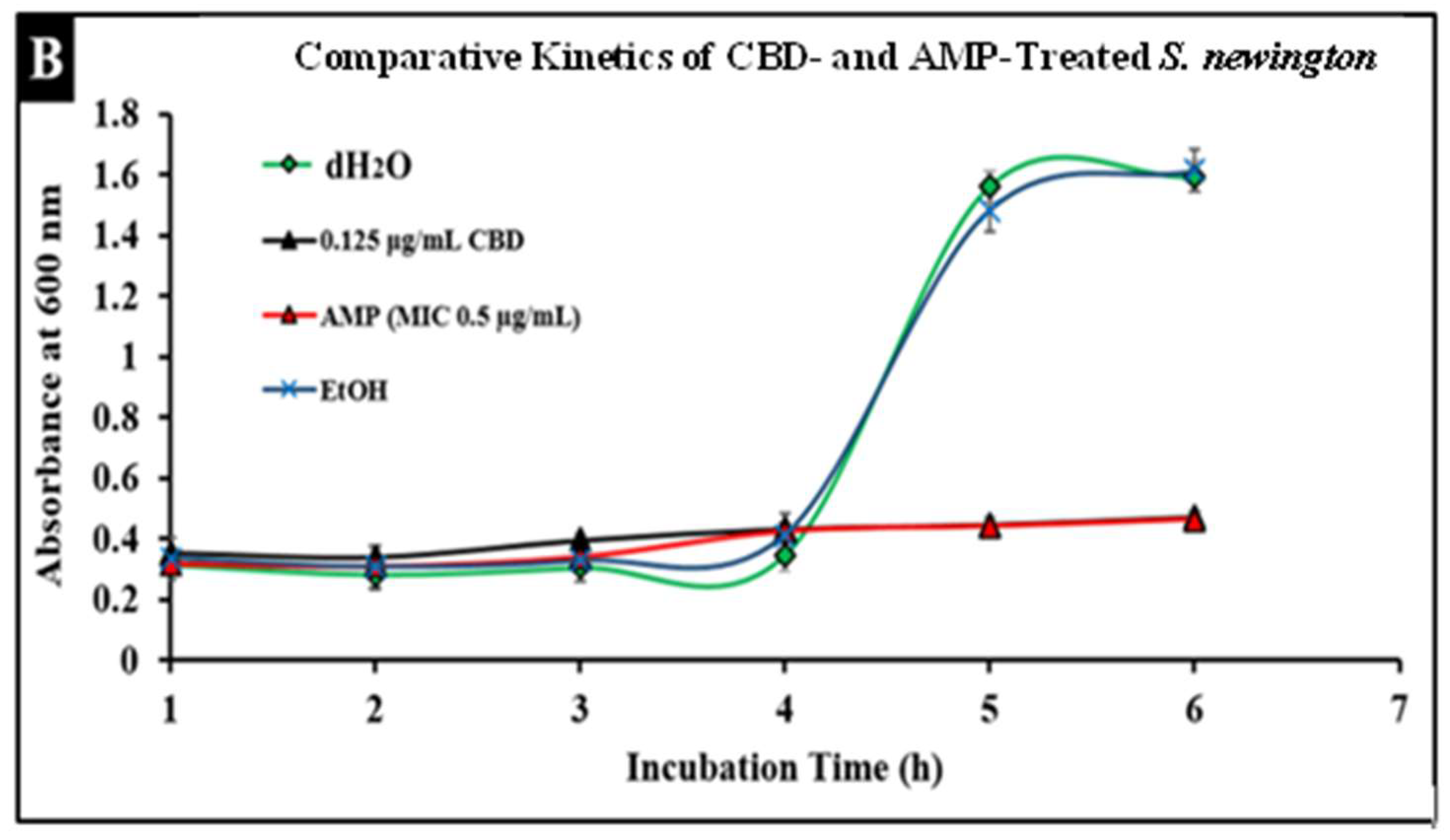
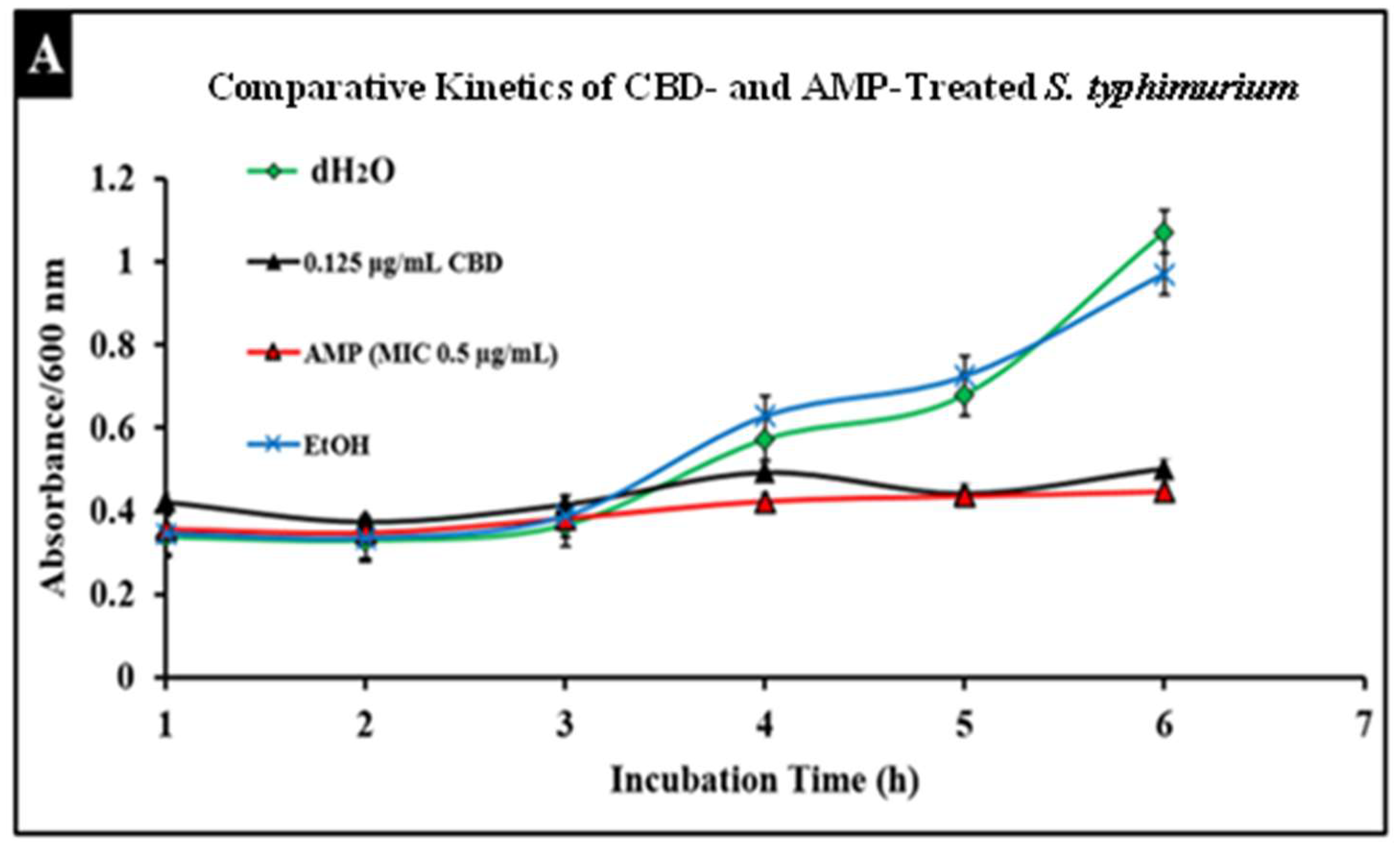
2.5. Comparative Study of CBD and Antibiotic Treatment against Salmonella
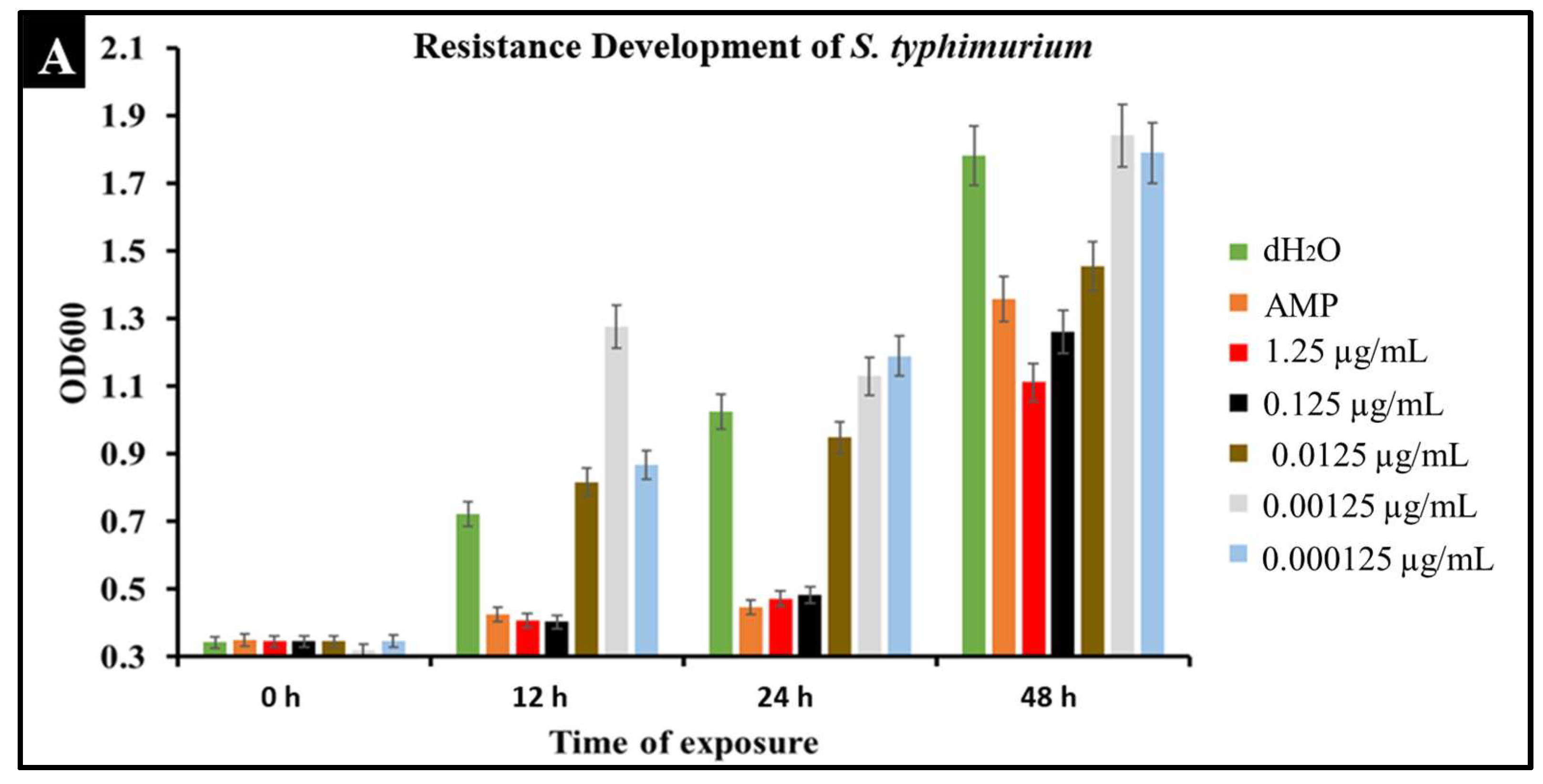
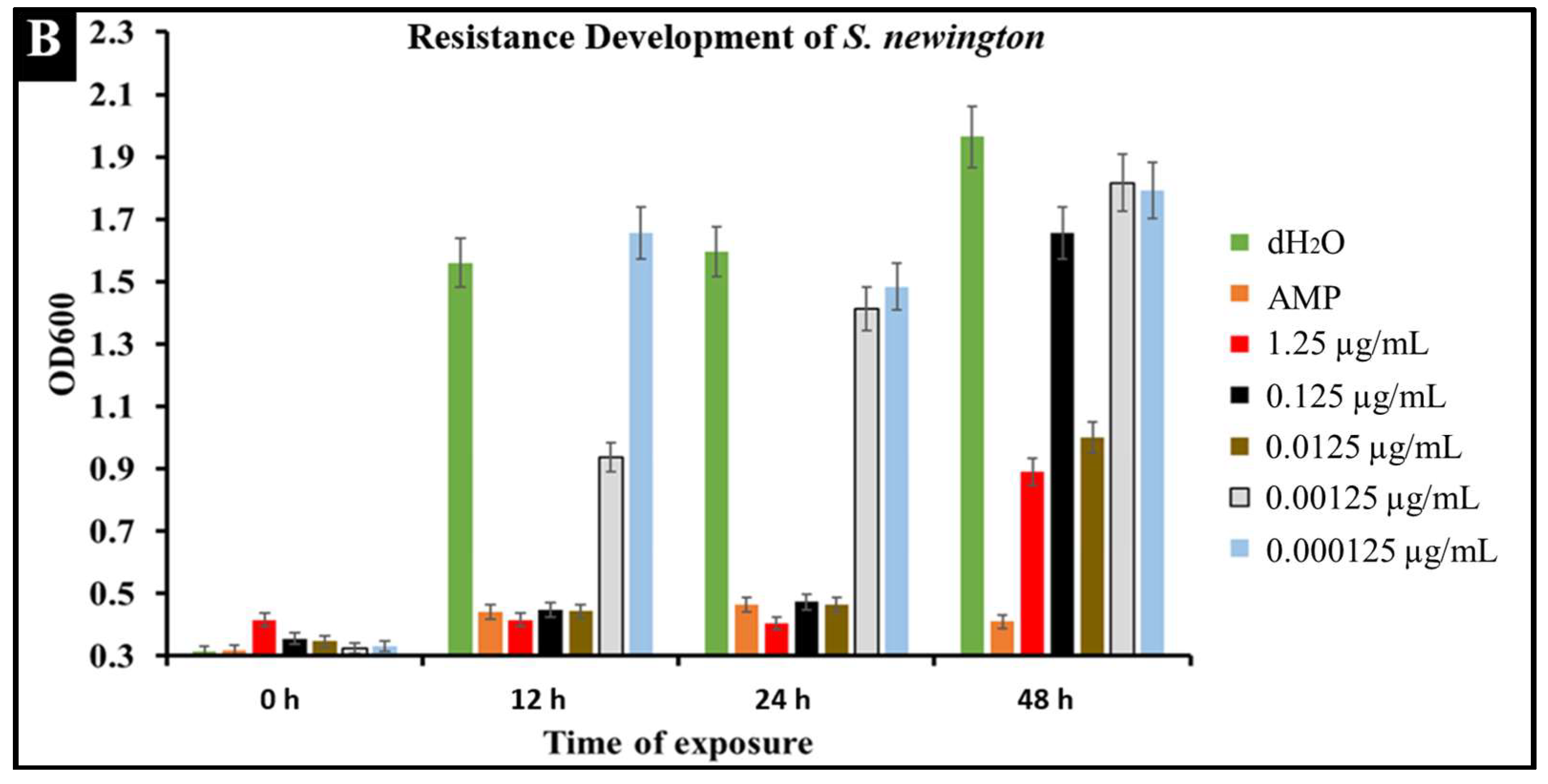
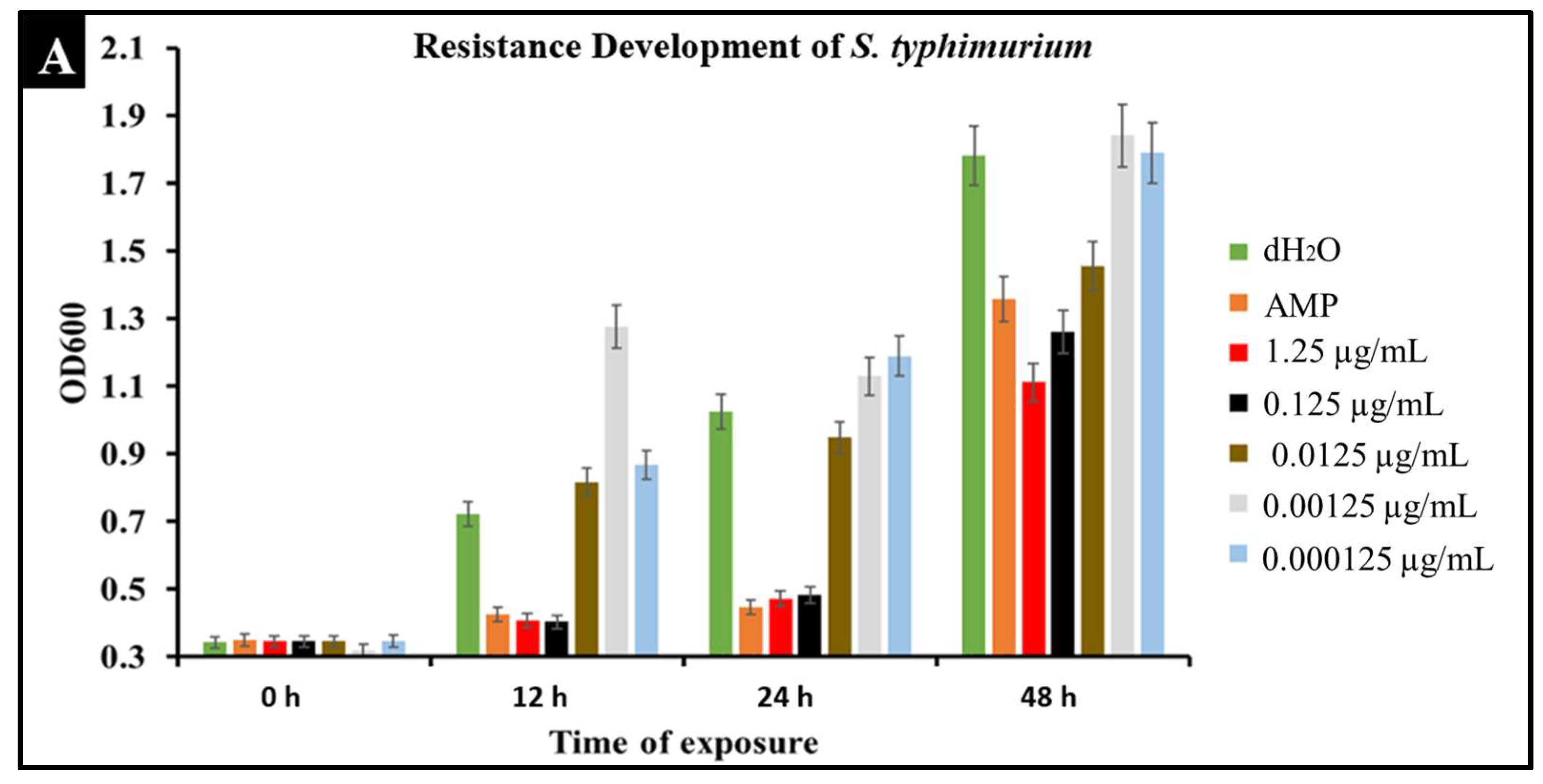
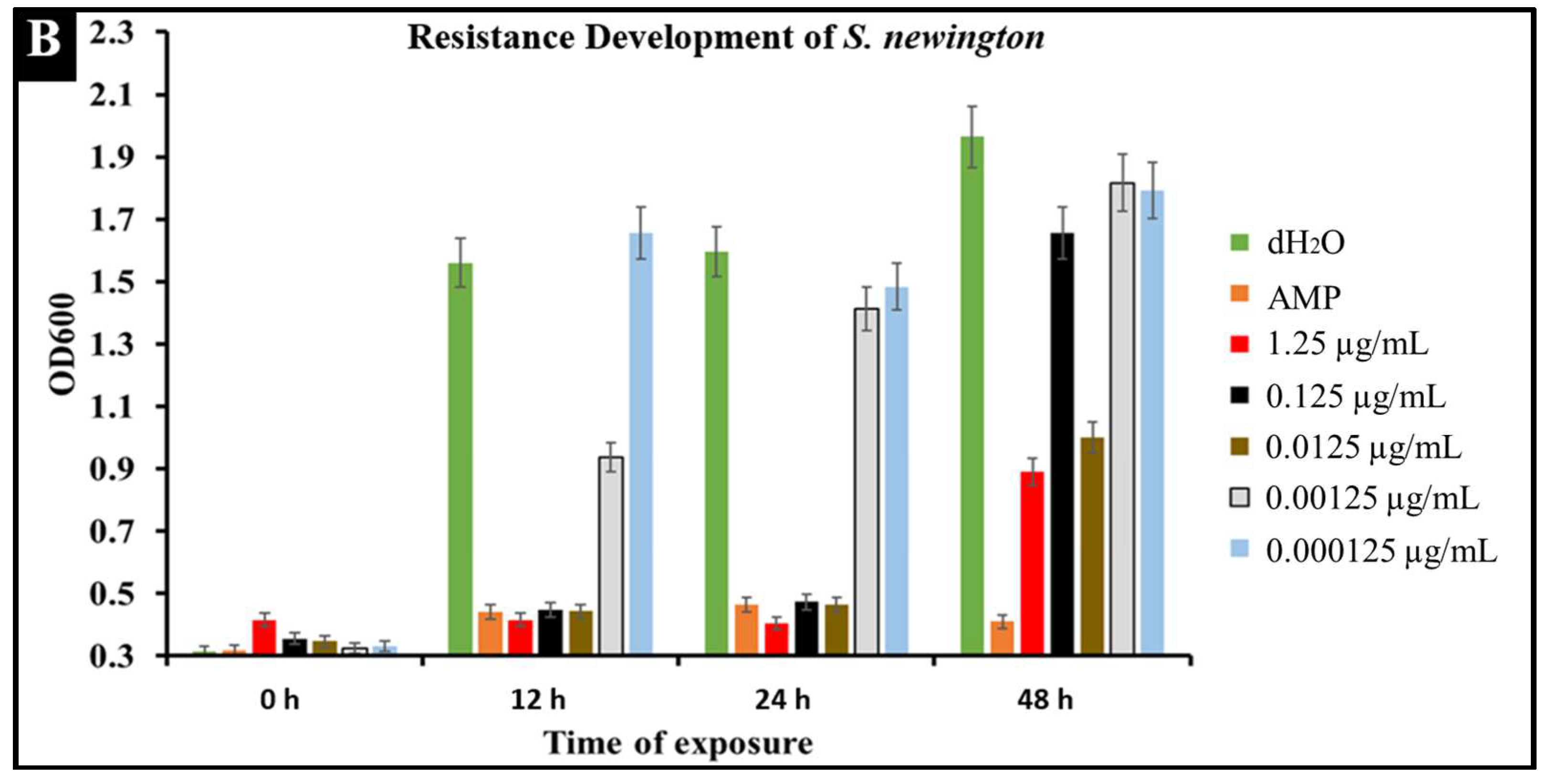
2.6. Developed Resistance of Salmonella to CBD Treatment
2.7. CBD Effectiveness against Salmonella Biofilm
3. Discussion
4. Materials and Methods
4.1. CBD Extraction
4.2. Gas Chromatography Analysis of CBD
4.3. Plate Assays for Antibacterial Screening of CBD
4.4. Fluorescence Microscopy of CBD-Treated Salmonella Cells
4.5. Bacterial Growth Kinetics
4.6. Bacterial Growth Kinetics in the Prescence of CBD
4.7. Comparative Bacterial Growth Kinetics in the Prescence of Ampicillin or CBD
4.8. SEM Imaging of CBD-Treated S. typhimurium Biofilm
4.9. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Sample Availability
References
- Long, T.; Wagner, M.; Demske, D.; Leipe, C.; Tarasov, P. Cannabis in Eurasia: Origin of human use and Bronze Age trans-continental connections. Veget. Hist. Archaeobot. 2017, 26, 245–258. [Google Scholar] [CrossRef]
- Zuardi, A.W. History of cannabis as a medicine: A review. Rev. Bras. Psiquiatr. 2006, 28, 153–157. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Crini, G.; Lichtfouse, E.; Chanet, G.; Crini, N. Applications of hemp in textiles, paper industry, insulation and building materials, horticulture, animal nutrition, food and beverages, nutraceuticals, cosmetics and hygiene, medicine, agrochemistry, energy production and environment: A review. Environ. Chem. Lett. 2020, 18, 1451–1476. [Google Scholar] [CrossRef]
- Rehman, M.; Fahad, S.; Du, G.; Cheng, X.; Yang, Y.; Tang, K.; Liu, L.; Liu, F.H.; Deng, C. Evaluation of hemp (Cannabis sativa L.) as an industrial crop: A review. Environ. Sci. Pollut. Res. 2021, 28, 52832–52843. [Google Scholar] [CrossRef] [PubMed]
- Aldrich, M. History of therapeutic cannabis. In Cannabis in Medical Practice; Mathre, M.L., Ed.; Mc Farland: Jefferson, NC, USA, 1997; pp. 35–55. [Google Scholar]
- Brand, J.E.; Zhao, Z. Cannabis in Chinese medicine: Are some traditional indications references in ancient literature related to cannabinoids. Front. Pharmacol. 2017, 8, 1663–9812. [Google Scholar] [CrossRef] [Green Version]
- Amar, M.B. Cannabinoids in medicine: A review of their therapeutic potential. J. Ethnopharmacol. 2006, 105, 1–25. [Google Scholar] [CrossRef]
- World Health Organization. Annex 1: Extract from the Report of the 41th Expert Committee on Drug Dependence: Cannabis and Cannabis Related Substances, p. 4. 2019. Available online: https://www.who.int/medicines/access/controlled-substances/Annex_1_41_ECDD_recommendations_cannabis_22Jan19pdf (accessed on 19 January 2022).
- Clementi, A. High time for a change: How the relationship between signatory countries and the United Nations Conventions governing narcotic drugs must adapt to foster a global shift in cannabis law. Brook. J. Int’L Law 2021, 46, 603. [Google Scholar]
- Scadron, T.E.; Koelble, T. Bongalong: Marketing Cannabis Products in a Stigmatized Environment; GSB: 2018-5; Graduate School of Business, University of Cape Town: Cape Town, South Africa, 2018. [Google Scholar]
- Geiger-Oneto, S.; Sprague, R. Cannabis regulatory confusion and its impact on consumer adoption. Am. Bus. Law. J. 2020, 57, 735–772. [Google Scholar] [CrossRef]
- Peters, J.; Chien, J. Contemporary routes of cannabis consumption: A primer for clinicians. J. Osteopath. Med. 2018, 118, 67–70. [Google Scholar] [CrossRef] [Green Version]
- Kalant, H. Medicinal use of cannabis: History and current status. Pain Res. Manag. 2001, 6, 12. [Google Scholar] [CrossRef] [Green Version]
- Solimini, R.; Rotolo, M.C.; Pichini, S.; Pacifici, R. Neurological disorders in medical use of cannabis: An update. CNS Neurol. Disord. Drug Targets 2017, 16, 527–533. [Google Scholar] [CrossRef] [PubMed]
- Fiani, B.; Sarhadi, K.J.; Soula, M.; Zafar, A.; Quadri, S.A. Current application of cannabidiol (CBD) in the management and treatment of neurological disorders. Neurol. Sci. 2020, 41, 3085–3098. [Google Scholar] [CrossRef] [PubMed]
- Waskow, I.E.; Olsson, J.E.; Salzman, C.; Katz, M.M. Psychological effects of tetrahydrocannabinol. Arch. Gen. Psychiatry 1970, 22, 97–107. [Google Scholar] [CrossRef] [PubMed]
- Karniol, I.G.; Shirakawa, I.; Takahashi, R.N.; Knobel, E.; Musty, R.E. Effects of delta9-tetrahydrocannabinol and cannabinol in man. Pharmacology 1975, 13, 502–512. [Google Scholar] [CrossRef]
- Gray, R.A.; Whalley, B.J. The proposed mechanisms of action of CBD in epilepsy. Epileptic Disord. 2020, 22, 10–15. [Google Scholar] [CrossRef]
- Pichini, S.; Malaca, S.; Gottardi, M.; Pérez-Acevedo, A.P.; Papaseit, E.; Perez-Maña, C.; Farré, M.; Pacifici, R.; Tagliabracci, A.; Mannocchi, G.; et al. F.P. UHPLC-MS/MS analysis of cannabidiol metabolites in serum and urine samples. Application to an individual treated with medical cannabis. Talanta 2021, 223, 121772. [Google Scholar] [CrossRef]
- Appendino, G.; Gibbons, S.; Giana, A.; Pagani, A.; Grassi, G.; Stavri, M.; Smith, E.; Rahman, M.M. Antibacterial cannabinoids from Cannabis sativa: A structure-activity study. J. Nat. Prod. 2008, 71, 1427–1430. [Google Scholar] [CrossRef]
- Blaskovich, M.A.T.; Kavanagh, A.M.; Elliott, A.G.; Zhang, B.; Ramu, S.; Amado, M.; Lowe, G.J.; Hinton, A.O.; Pham, D.M.T.; Zuegg, J.; et al. The antimicrobial potential of cannabidiol. Commun. Biol. 2021, 4, 7. [Google Scholar] [CrossRef]
- van Klingeren, B.; ten Ham, M. Antibacterial activity of Δ9-tetrahydrocannabinol and cannabidiol. Antonie. Van. Leeuwenhoek 1976, 42, 9–12. [Google Scholar] [CrossRef]
- Iseppi, R.; Brighenti, V.; Licata, M.; Lambertini, A.; Sabia, C.; Messi, P.; Pellati, F.; Benvenuti, S. Chemical characterization and evaluation of the antibacterial activity of essential oils from fibre-type Cannabis sativa L. (Hemp). Molecules 2019, 24, 2302. [Google Scholar] [CrossRef] [Green Version]
- Livermore, D.M. Has the era of untreatable infections arrived? J. Antimicrob. Chemother. 2009, 64, i29–i36. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- WHO. Antimicrobial Resistance. 2020. Available online: https://www.who.int/healthtopics/antimicrobial-resistance (accessed on 19 January 2022).
- Ayariga, J.A.; Gildea, L.; Wu, H.; Villafane, R. The Ɛ34 phage tailspike protein: An in vitro characterization, structure prediction, potential interaction with S. newington LPS and cytotoxicity assessment to animal cell line. bioRxiv 2021. [Google Scholar] [CrossRef]
- Gildea, L.; Ayariga, J.A.; Robertson, B.K.; Villafane, R. P22 phage shows promising antibacterial activity under pathophysiological conditions. Arch. Microbiol. Immunol. 2022, 6, 81–100. [Google Scholar] [CrossRef]
- Ardal, C.; Balasegaram, M.; Laxminarayan, R.; McAdams, D.; Outterson, K.; Rex, J.H.; Sumpradit, N. Antibiotic development–economic, regulatory, and societal challenges. Nat. Rev. Microbiol. 2020, 18, 267–274. [Google Scholar] [CrossRef]
- Sharland, M.; Gandra, S.; Huttner, B.; Moja, L.; Pulcini, C.; Zeng, M.; Mendelson, M.; Cappello, B.; Cooke, G.; Magrini, N.; et al. Encouraging AWaRe-ness and discouraging inappropriate antibiotic use-the new 2019 Essential medicines list becomes a global antibiotic stewardship tool. Lancet Infect. Dis. 2019, 19, 1278–1280. [Google Scholar] [CrossRef]
- CDC. Antibiotic Resistance Threats in the United States. 2019. Available online: https://www.cdc.gov/DrugResistance/Biggest-Threats.html (accessed on 19 January 2022).
- Slayton, R.B.; Toth, D.; Lee, B.Y.; Tanner, W.; Bartsch, S.M.; Khader, K.; Wong, K.; Brown, K.; McKinnell, J.A.; Ray, W.; et al. Vital signs: Estimated effects of a coordinated approach for action to reduce antibiotic-resistant infections in health care facilities-United States. MMWR Morb. Mortal. Wkly. Rep. 2015, 64, 826–831. [Google Scholar] [CrossRef]
- White, C.M.A. Review of human studies assessing cannabidiol’s (CBD) therapeutic actions and potential. J. Clin. Pharmacol. 2019, 59, 923–934. [Google Scholar] [CrossRef]
- Watt, G.; Karl, T. In vivo evidence for therapeutic properties of cannabidiol (CBD) for alzheimer’s disease. Front. Pharmocol. 2017, 8, 20. [Google Scholar] [CrossRef] [Green Version]
- Solowij, N.; Broyd, S.J.; Beale, C.; Prick, J.A.; Greenwood, L.M.; van Hell, H.; Suo, C.; Galettis, P.; Pai, N.; Fu, S.; et al. Therapeutic effects of prolonged cannabidiol treatment on psychological symptoms and cognitive function in regular cannabis users: A pragmatic open-label clinical trial. Cannabis Cannabinoid Res. 2018, 3, 21–34. [Google Scholar] [CrossRef] [Green Version]
- World Health Organization. Cannabidiol (CBD): Critical Review Report: Expert Committee on Drug Dependence, Fortieth Meeting; World Health Organization: Geneva, Switzerland, 2018. [Google Scholar]
- Kosgodage, U.S.; Matewele, P.; Awamaria, B.; Kraev, I.; Warde, P.; Mastroianni, G.; Nunn, A.V.; Guy, G.W. Cannabidiol is a novel modulator of bacterial membrane vesicles. Front. Cell. Infect. Microbiol. 2019, 9, 2235–2988. [Google Scholar] [CrossRef]
- Bryan, F.L. Foodborne diseases in the United States associated with meat and poultry. J. Food Prot. 1980, 43, 140–150. [Google Scholar] [CrossRef] [PubMed]
- Heinitz, M.L.; Ruble, R.D.; Wagner, D.E.; Tatini, S.R. Incidence of Salmonella in fish and seafood. J. Food. Prot. 2000, 63, 579–592. [Google Scholar] [CrossRef] [PubMed]
- Park, S.Y.; Woodward, C.L.; Kubena, L.F.; Nisbet, D.J.; Birkhold, S.G.; Ricke, S.C. Environmental dissemination of foodborne Salmonella in preharvest poultry production: Reservoirs, critical factors, and research strategies. Crit. Rev. Environ. Sci. Technol. 2008, 38, 73–111. [Google Scholar] [CrossRef]
- White, D.G.; Zhao, S.; Sudler, R.; Ayers, S.; Friedman, S.; Chen, S.; McDermott, P.; McDermott, S.; Wagner, D.; Meng, J. The isolation of antibiotic-resistant Salmonella from retail ground meats. N. Engl. J. Med. 2001, 345, 1147–1154. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Iseppi, R.; Mariani, M.; Condò, C.; Sabia, C.; Messi, P. Essential oils: A natural weapon against antibiotic-resistant bacteria responsible for nosocomial infections. Antibiotics 2021, 10, 417. [Google Scholar] [CrossRef]
- Harrell, J.E.; Hahn, M.M.; D’Souza, S.J.; Vasicek, E.M.; Sandala, J.L.; Gunn, J.S.; McLachlan, J.B. Salmonella biofilm formation, chronic infection, and immunity within the intestine and hepatobiliary tract. Front. Cell. Infect. Microbiol. 2021, 10, 910. [Google Scholar] [CrossRef] [PubMed]
- Atalay, S.; Jarocka-Karpowicz, I.; Skrzydlewska, E. Antioxidative and anti-inflammatory properties of cannabidiol. Antioxidants 2020, 9, 21. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- di Giacomo, V.; Recinella, L.; Chiavaroli, A.; Orlando, G.; Cataldi, A.; Rapino, M.; Di Valerio, V.; Politi, M.; Antolini, M.D.; Acquaviva, A.; et al. Metabolomic profile and antioxidant/anti-inflammatory effects of industrial hemp water extract in fibroblasts, keratinocytes and isolated mouse skin specimens. Antioxidants 2021, 10, 44. [Google Scholar] [CrossRef]
- Nikaido, H.; Basina, M.; Nguyen, V.; Rosenberg, E.Y. Multidrug efflux pump AcrAB of Salmonella typhimurium excretes only those β-Lactam antibiotics containing lipophilic side chains. J. Bacteriol. 1998, 180, 17. [Google Scholar] [CrossRef] [Green Version]
- Ruiz, J.; Capitano, L.; Nuñez, L.; Castro, D.; Sierra, J.M.; Hatha, M.; Borrego, J.J.; Vila, J. Mechanisms of resistance to ampicillin, chloramphenicol and quinolones in multiresistant Salmonella typhimurium strains isolated from fish. J. Antimicrob. Chemother. 1999, 43, 699–702. [Google Scholar] [CrossRef] [Green Version]
- Eguale, T.; Birungi, J.; Asrat, D.; Njahira, M.N.; Njuguna, J.; Gebreyes, W.A.; Gunn, J.S.; Djikeng, A.; Engidawork, E. Genetic markers associated with resistance to beta-lactam and quinolone antimicrobials in non-typhoidal Salmonella isolates from humans and animals in central Ethiopia. Antimicrob. Resist. Infect. Control 2017, 6, 13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lukacik, P.; Barnard, T.J.; Keller, P.W.; Chaturvedi, K.S.; Seddiki, N.; Fairman, J.W.; Noinaj, N.; Kirby, T.L.; Henderson, J.P.; Steven, A.C.; et al. Structural engineering of a phage lysin that targets gram negative pathogens. Proc. Natl. Acad. Sci. USA 2012, 109, 9857–9862. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lin, D.M.; Koskella, B.; Lin, H.C. Phage therapy: An alternative to antibiotics in the age of multi-drug resistance. World J. Gastrointest. Pharmacol. Ther. 2017, 8, 162–173. [Google Scholar] [CrossRef] [PubMed]
- Ghosh, C.; Sarkar, P.; Issa, R.; Haldar, J. Alternatives to conventional antibiotics in the era of antimicrobial resistance. Trends Microbiol. 2019, 27, 323–338. [Google Scholar] [CrossRef] [PubMed]
- D’Accolti, M.; Soffritti, I.; Mazzacane, S.; Caselli, E. Bacteriophages as a potential 360-degree pathogen control strategy. Microorganisms 2021, 9, 261. [Google Scholar] [CrossRef] [PubMed]
- Lorenzi, V.; Muselli, A.; Bernardini, A.F.; Berti, L.; Pagès, J.M.; Amaral, L.; Bolla, J.M. Geraniol restores antibiotic activities against multidrug-resistant isolates from gram-negative species. Antimicrob. Agents Chemother. 2009, 53, 2209–2211. [Google Scholar] [CrossRef] [Green Version]
- Sustainable CBD LLC. Sustainable CBD LLC Standard of Procedures (SOP) 1–6; Sustainable CBD LLC: Salem, AL, USA, 2020. [Google Scholar]
- Valizadehderakhshan, M.; Shahbazi, A.; Kazem-Rostami, M.; Todd, M.S.; Bhowmik, A.; Wang, L. Extraction of cannabinoids from Cannabis sativa L. (Hemp)—Review. Agriculture 2021, 11, 384. [Google Scholar] [CrossRef]
- Grijó, D.R.; Osorio, I.A.V.; Cardozo-Filho, L. Supercritical extraction strategies using CO2 and ethanol to obtain cannabinoid compounds from Cannabis hybrid flowers. J. CO2 Util. 2018, 28, 174–180. [Google Scholar] [CrossRef]
- Kirby, W.M.M.; Yoshihara, G.M.; Sundsted, K.S.; Warren, J.H. Clinical usefulness of a single disc method for antibiotic sensitivity testing. Antibiot. Annu. 1957, 892–897. Available online: https://pubmed.ncbi.nlm.nih.gov/13425478/ (accessed on 19 January 2022).
- Clinical Laboratory Standards Institute. Performance Standards for Antimicrobial Disk Susceptibility Tests, 9th ed.; CLSI Document M2-A9; Clinical Laboratory Standards Institute: Malvern, PA, USA, 2006. [Google Scholar]
- Jorgensen, J.H.; Turnidge, J.D. Susceptibility test methods: Dilution and disk diffusion methods. Man. Clin. Microbiol. 2007, 9, 1152–1172. [Google Scholar]
- Powell, R.; Neilan, J.; Gannon, F. Plaque dot assay. Nucleic Acids Res. 1986, 14, 1541. [Google Scholar] [CrossRef] [Green Version]
- Kalapala, Y.C.; Sharma, P.R.; Agarwal, R. Antimycobacterial potential of mycobacteriophage under disease-mimicking conditions. Front. Microbiol. 2020, 11, 583661. [Google Scholar] [CrossRef] [PubMed]
- Ayariga, J.A.; Gildea, L.; Villafane, R. Ɛ34 phage tailspike protein is resistant to trypsin and inhibits Salmonella biofilm formation. Enliven Microb. Microb. Tech. 2022, 9, 002. [Google Scholar]
- Mittrücker, H.W.; Kaufmann, S.H. Immune response to infection with Salmonella typhimurium in mice. J. Leukoc. Biol. 2000, 67, 457–463. [Google Scholar] [CrossRef]
- Pui, C.F.; Wong, W.C.; Chai, L.C.; Tunung, R.; Jeyaletchumi, P.; Hidayah, N.; Ubong, A.; Farinazleen, M.G.; Cheah, Y.K.; Son, R. Salmonella: A foodborne pathogen. Int. Food Res. J. 2011, 18, 465–473. [Google Scholar]
- Ayariga, J.A.; Abugri, D.; Amrutha, B.; Villafane, R. Capsaicin potently blocks Salmonella typhimurium invasion of Vero cells. bioRxiv 2022. [Google Scholar] [CrossRef]




Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gildea, L.; Ayariga, J.A.; Ajayi, O.S.; Xu, J.; Villafane, R.; Samuel-Foo, M. Cannabis sativa CBD Extract Shows Promising Antibacterial Activity against Salmonella typhimurium and S. newington. Molecules 2022, 27, 2669. https://doi.org/10.3390/molecules27092669
Gildea L, Ayariga JA, Ajayi OS, Xu J, Villafane R, Samuel-Foo M. Cannabis sativa CBD Extract Shows Promising Antibacterial Activity against Salmonella typhimurium and S. newington. Molecules. 2022; 27(9):2669. https://doi.org/10.3390/molecules27092669
Chicago/Turabian StyleGildea, Logan, Joseph Atia Ayariga, Olufemi S. Ajayi, Junhuan Xu, Robert Villafane, and Michelle Samuel-Foo. 2022. "Cannabis sativa CBD Extract Shows Promising Antibacterial Activity against Salmonella typhimurium and S. newington" Molecules 27, no. 9: 2669. https://doi.org/10.3390/molecules27092669
APA StyleGildea, L., Ayariga, J. A., Ajayi, O. S., Xu, J., Villafane, R., & Samuel-Foo, M. (2022). Cannabis sativa CBD Extract Shows Promising Antibacterial Activity against Salmonella typhimurium and S. newington. Molecules, 27(9), 2669. https://doi.org/10.3390/molecules27092669