
Flavonoids and Phenols, the Potential Anti-Diabetic Compounds from Bauhinia strychnifolia Craib. Stem.


Abstract
:1. Introduction
2. Results and Discussions
2.1. B. strychnifolia Stem Extraction
2.2. Total Phenolic Contents (TPC) and Total Flavonoid Contents (TFC)
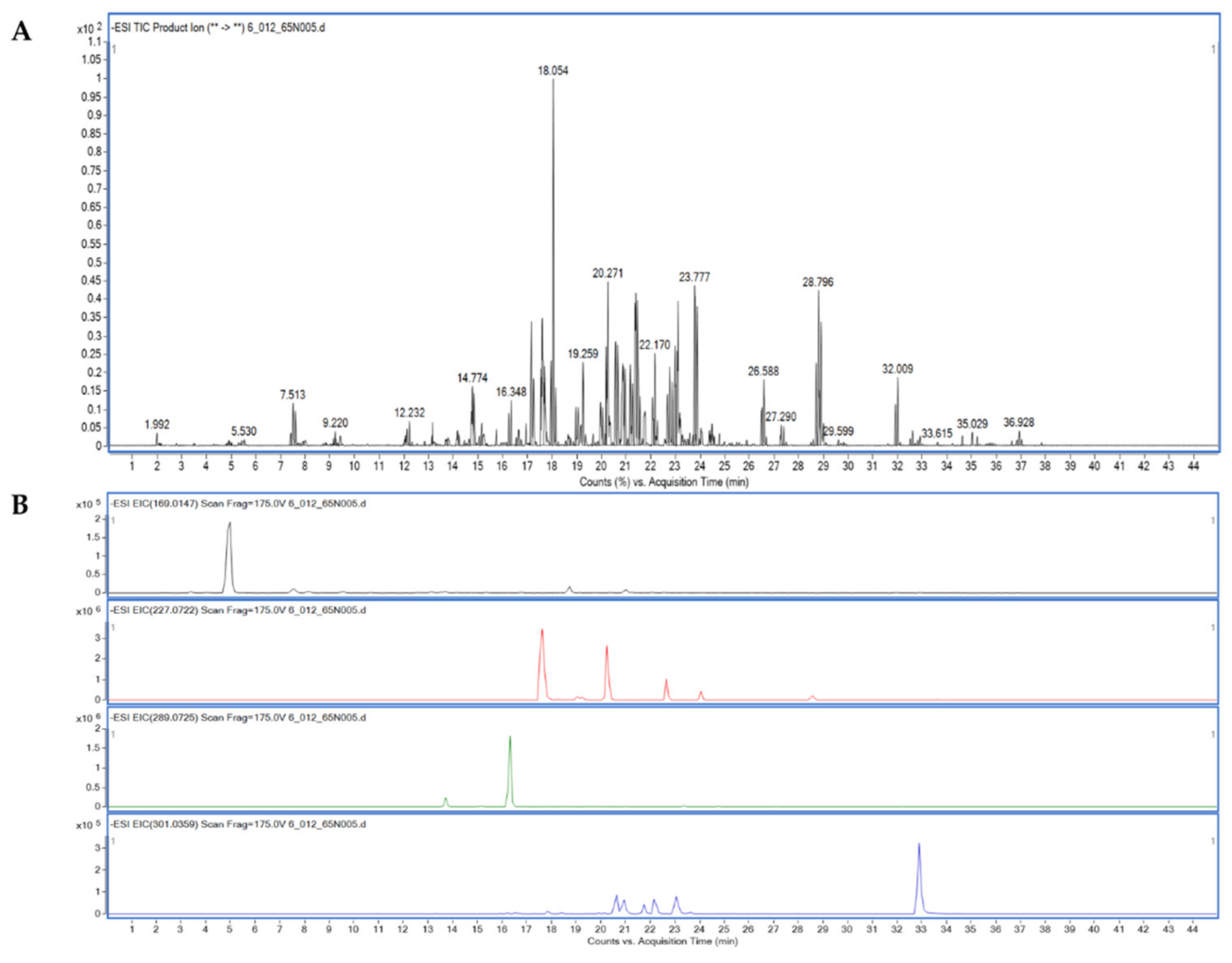
2.3. Liquid Chromatography Tandem Mass Spectrometry (LC-MS/MS) Analysis
2.4. Isolation
2.4.1. Resveratrol
2.4.2. Epicatechin
2.4.3. Quercetin
2.4.4. Gallic Acid
2.5. α-Glucosidase Inhibitory Assay
2.6. Dipeptidyl Peptidase-IV (DPP-IV) Inhibitory Assay
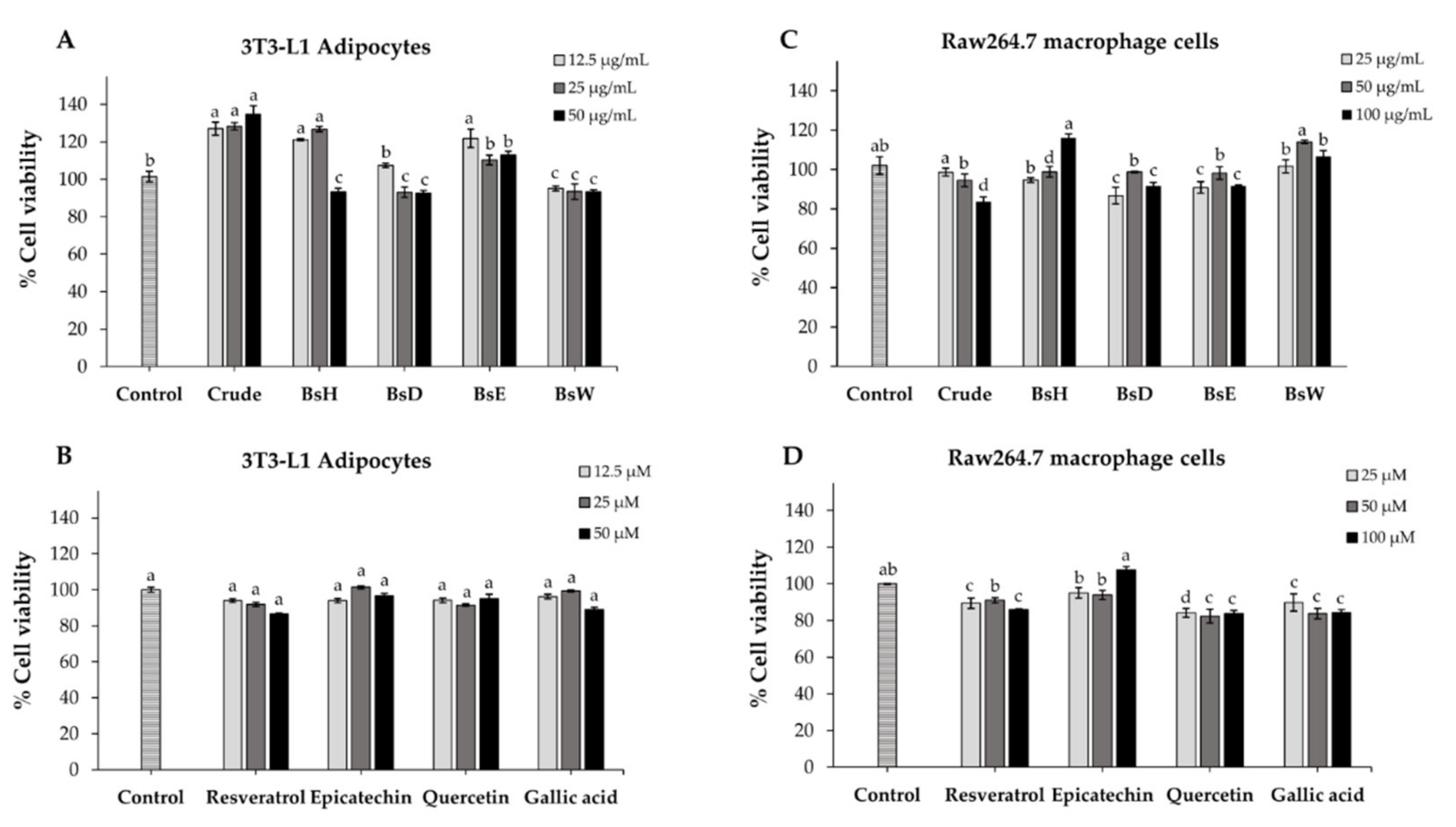
2.7. Cell Viability
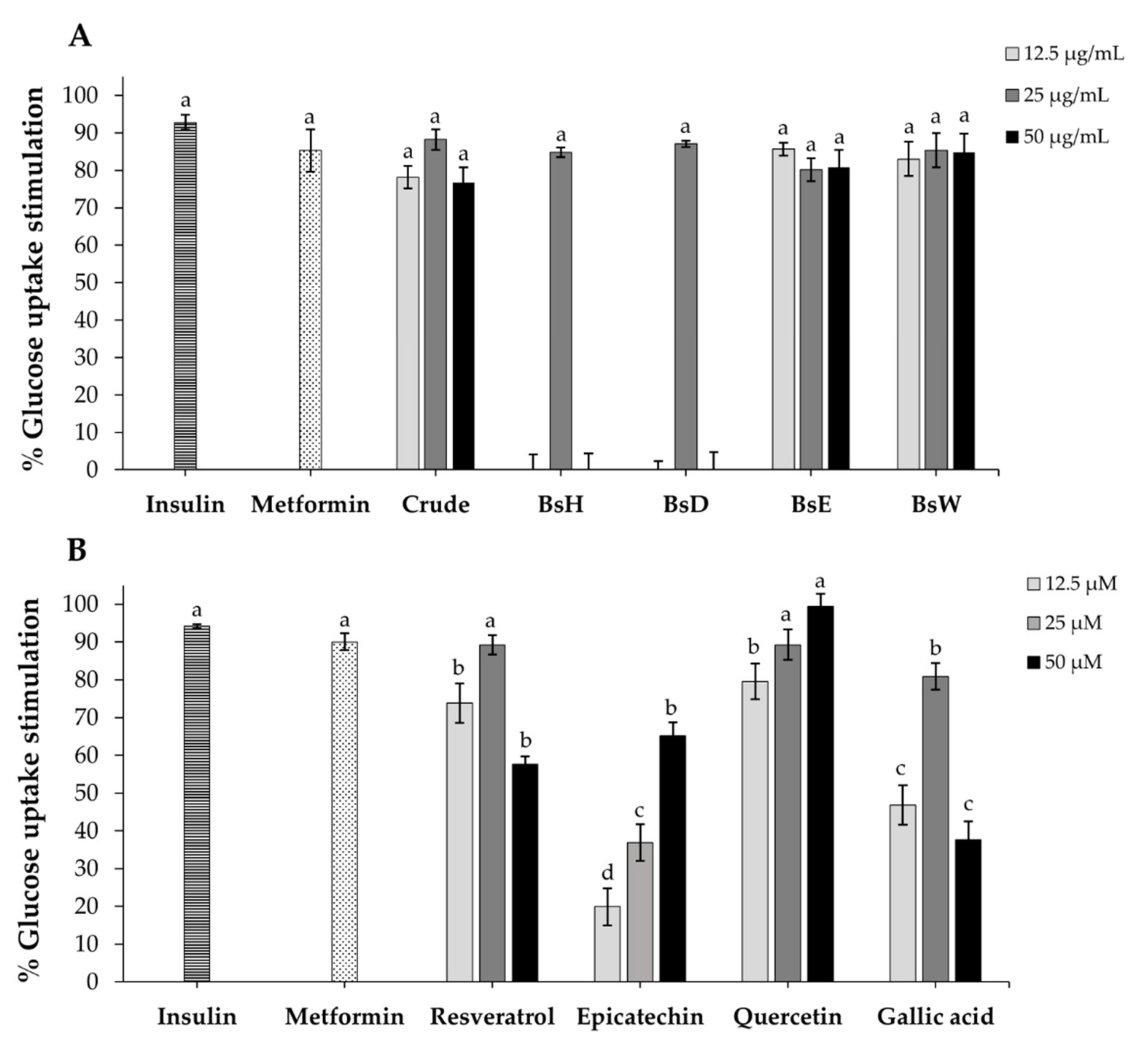
2.8. Glucose Uptake in 3T3-L1 Adipocytes
2.9. Nitric Oxide (NO) Production
3. Materials and Methods
3.1. Chemicals and Reagents
3.2. Plant Materials
3.3. Preparation of B. strychnifolia Extracts and Its Isolated Compounds
3.4. Total Phenolic Contents (TPC)
3.5. Total Flavonoid Contents (TFC)
3.6. Liquid Chromatography with Tandem Mass Spectrometry (LC-MS/MS) Analysis
3.7. α-Glucosidase Inhibitory Assay
3.8. Dipeptidyl Peptidase-IV (DPP-IV) Inhibitory Assay
3.9. Cell Culture
3.10. Cell Differentiation
3.11. Cell Viability Assay
3.12. Glucose Uptake Assay
3.13. Inhibition of Nitric Oxide (NO) Production Using Griess Renitricaction Assay
3.14. Statistics
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Sample Availability
Abbreviations
| A | absorbance |
| ANOVA | analysis of variance |
| °C | degree celsius |
| DMEM | Dulbecco’s modified Eagle’s medium |
| DMSO | dimethyl sulfoxide |
| GP-p-NA | gly-pro-p-nitroanilide |
| IBMX | isobutyl-methylxanthine |
| LPS | lipopolysaccharide |
| MTT | 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetra-zolium bromide |
| nm | nanometer |
| NMR | nuclear magnetic resonance |
| NO | nitric oxide |
| pNPG | para-nirophenyl-α-D-glucopyranoside |
| RPMI | Roswell Park Memorial Institute |
| UV | ultraviolet |
References
- American Diabetes Association. Diagnosis and Classification of Diabetes Mellitus. Diabetes Care 2007, 30, S42–S47. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Saeedi, P.; Petersohn, I.; Salpea, P.; Malanda, B.; Karuranga, S.; Unwin, N.; Colagiuri, S.; Guariguata, L.; Motala, A.A.; Ogurtsova, K.; et al. Global and regional diabetes prevalence estimates for 2019 and projections for 2030 and 2045: Results from the International Diabetes Federation Diabetes Atlas, 9th edition. Diabetes Res. Clin. Pract. 2019, 157, 107843. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sun, H.; Saeedi, P.; Karuranga, S.; Pinkepank, M.; Ogurtsova, K.; Duncan, B.B.; Stein, C.; Stein, A.; Chan, J.C.N.; Mbanya, J.C.; et al. IDF diabetes atlas: Global, regional, and country-level diabetes prevalence estimates for 2021 and projections for 2045. Diabetes Res. Clin. Pract. 2022, 183, 109119. [Google Scholar] [CrossRef]
- Galicia-Garcia, U.; Benito-Vicente, A.; Jebari, S.; Larrea-Sebal, A.; Siddiqi, H.; Uribe, K.B.; Ostolaza, H.; Martín, C. Pathophysiology of type 2 diabetes mellitus. Int. J. Mol. Sci. 2020, 21, 6275. [Google Scholar] [CrossRef] [PubMed]
- Deerochanawong, C.; Ferrario, A. Diabetes management in Thailand: A literature review of the burden, costs, and outcomes. Glob. Health 2013, 9, 11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Manukumar, H.M.; Kumar, J.S.; Chandrasekhar, B.; Raghava, S.; Umesha, S. Evidences for diabetes and insulin mimetic activity of medicinal plants: Present status and future prospects. Crit. Rev. Food Sci. 2017, 57, 2712–2729. [Google Scholar] [CrossRef] [PubMed]
- Aryangat, A.V.; Gerich, J.E. Type 2 diabetes: Postprandial hyperglycemia and increased cardiovascular risk. Vasc. Health Risk Manag. 2012, 6, 145–155. [Google Scholar] [CrossRef] [Green Version]
- Rehani, P.R.; Iftikhar, H.; Nakajima, M.; Tanaka, T.; Jabbar, Z.; Rehani, R.N. Safety and Mode of Action of Diabetes Medications in comparison with 5-Aminolevulinic Acid (5-ALA). J. Diabetes Res. 2019, 2019, 4267357. [Google Scholar] [CrossRef] [Green Version]
- Meneses, M.J.; Silva, B.M.; Sousa, M.; Sá, R.; Oliveira, P.F.; Alves, M.G. Antidiabetic Drugs: Mechanisms of Action and Potential Outcomes on Cellular Metabolism. Curr. Pharm. Des. 2015, 21, 3606–3620. [Google Scholar] [CrossRef]
- Hollander, P. Safety profile of acabose, an alpha-glucosidase inhibitor. Drugs 1992, 44, 47–53. [Google Scholar] [CrossRef]
- Chaudhury, A.; Duvoor, C.; Reddy Dendi, V.S.; Kraleti, S.; Chada, A.; Ravilla, R.; Marco, A.; Shekhawat, N.S.; Montales, M.T.; Kuriakose, K.; et al. Clinical Review of Antidiabetic Drugs: Implications for Type 2 Diabetes Mellitus Management. Front. Endocrinol. 2017, 8, 6. [Google Scholar] [CrossRef] [Green Version]
- Abdullah, K.M.; Arefeen, A.; Shamsi, A.; Alhumaydhi, F.A.; Naseem, I. Insight into the In Vitro Antiglycation and In Vivo Antidiabetic Effects of Thiamine: Implications of Vitamin B1 in Controlling Diabetes. ACS Omega. 2021, 6, 12605–12614. [Google Scholar] [CrossRef]
- Adeshara, K.A.; Bangar, N.; Diwan, A.G.; Tupe, R.S. Plasma glycation adducts and various RAGE isoforms are intricately associated with oxidative stress and inflammatory markers in type 2 diabe. Diabetes Metab. Syndr. Clin. Res. Rev. 2022, 102441. [Google Scholar] [CrossRef]
- Panchinda, C.; Ruangnoo, S.; Itharat, A. Cytotoxic activity against cancer cell lines from the ethanolic extracts and its VLC fractions of Bauhinia strychnifolia leaves. J. Med. Assoc. Thail. 2016, 99 (Suppl. S4), S110–S115. [Google Scholar]
- Kaewpiboon, C.; Lirdprapamongkol, K.; Srisomsap, C.; Winayanuwattikun, P.; Yongvanich, T.; Puwaprisirisan, P.; Svasti, J.; Assavalapsakul, W. Studies of the in vitro cytotoxic, antioxidant, lipase inhibitory and antimicrobial activities of selected Thai medicinal plants. BMC Complemetary Altern. Med. 2012, 12, 217. [Google Scholar] [CrossRef] [Green Version]
- Itharat, A.; Sayompark, S.; Hansakul, P.; Dechayont, B. In vitro antioxidant activities of extracts of Bauhinia strychnifolia stems and leaves: Comparison with activities in green tea extracts. Med. Aromat Plants 2016, 5, 2167-0412. [Google Scholar] [CrossRef]
- Yuenyongsawad, S.; Bunluepuech, K.; Wattanapiromsakul, C.; Tewtrakul, S. Anti-cancer activity of compounds from Bauhinia strychnifolia stem. J. Ethnopharmacol. 2013, 150, 765–769. [Google Scholar] [CrossRef]
- Bunluepuech, K.; Tewtrakul, S.; Wattanapiromsakul, C. Alpha-glucosidase inhibitory activity of compounds from Bauhinia strychnifolia. J. Chem. Pharm. Res. 2019, 11, 22–26. [Google Scholar]
- Thaina, P.; Rattanamusik, N.; Saraphee, T.; Tengyai, S.; Pattanawongsa, A.; Puttarak, P. Anti-diabetic activity of Bauhinia strychnifolia Craib stem extract in Alloxan-induced diabetic rat. Thai J. Pharm. Sci. 2022, 46, accepted. [Google Scholar]
- Do, Q.D.; Angkawijaya, A.E.; Tran-Nguyen, P.L.; Huynh, L.H.; Soetaredjo, F.E.; Ismadji, S.; Ju, Y.H. Effect of extraction solvent on total phenol content, total flavonoid content, and antioxidant activity of Limnophila aromatica. J. Food Drug Anal. 2014, 22, 296–302. [Google Scholar] [CrossRef] [Green Version]
- Vinayagam, R.; Xu, B. Antidiabetic properties of dietary flavonoids: A cellular mechanism review. Nutr. Metab. 2015, 12, 60. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ong, K.C.; Khoo, H.E. Effects of myricetin on glycemia and glycogen metabolism in diabetic rats. Life Sci. 2000, 67, 1695–1705. [Google Scholar] [CrossRef]
- Al-Ishaq, R.K.; Abotaleb, M.; Kubatka, P.; Kajo, K.; Büsselberg, D. Flavonoids and Their Anti-Diabetic Effects: Cellular Mechanisms and Effects to Improve Blood Sugar Levels. Biomolecules 2019, 9, 430. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Babbar, N.; Oberoi, H.S.; Sandhu, S.K.; Bhargav, V.K. Influence of different solvents in extraction of phenolic compounds from vegetable residues and their evaluation as natural sources of antioxidants. J. Food Sci. Technol. 2014, 51, 2568–2575. [Google Scholar] [CrossRef] [Green Version]
- Pandey, K.B.; Rizvi, S.I. Plant polyphenols as dietary antioxidants in human health and disease. Oxidative Med. Cell. Longev. 2009, 2, 270–278. [Google Scholar] [CrossRef] [Green Version]
- Bule, M.; Abdurahman, A.; Nikfar, S.; Abdollahi, M.; Amini, M. Antidiabetic effect of quercetin: A systematic review and meta-analysis of animal studies. Food Chem. Toxicol. 2019, 125, 494–502. [Google Scholar] [CrossRef]
- Eid, H.M.; Martineau, L.C.; Saleem, A.; Muhammad, A.; Vallerand, D.; Benhaddou-Andaloussi, A.; Haddad, P.S. Stimulation of AMP-activated protein kinase and enhancement of basal glucose uptake in muscle cells by quercetin and quercetin glycosides, active principles of the antidiabetic medicinal plant Vaccinium vitis-idaea. Mol. Nutr. Food Res. 2010, 54, 991–1003. [Google Scholar] [CrossRef]
- Zanatta, L.; Rosso, A.; Folador, P.; Figueiredo, M.S.; Pizzolatti, M.G.; Leite, L.D.; Silva, F.R. Insulinomimetic effect of kaempferol 3-neohesperidoside on the rat soleus muscle. J. Nat. Prod. 2008, 71, 532–535. [Google Scholar] [CrossRef]
- Zhang, Y.; Liu, D. Flavonol kaempferol improves chronic hyperglycemia-impaired pancreatic beta-cell viability and insulin secretory function. Eur. J. Pharmacol. 2011, 670, 325–332. [Google Scholar] [CrossRef]
- Campanero, M.A.; Escolar, M.; Perez, G.; Garcia-Quetglas, E.; Sadaba, B.; Azanza, J.R. Simultaneous determination of diosmin and diosmetin in human plasma by ion trap liquid chromatography-atmospheric pressure chemical ionization tandem mass spectrometry: Application to a clinical pharmacokinetic study. J. Pharm. Biomed. Anal. 2010, 51, 875–881. [Google Scholar] [CrossRef]
- Zang, Y.; Igarashi, K.; Li, Y. Anti-diabetic effects of luteolin and luteolin-7-O-glucoside on KK-A(y) mice. Biosci. Biotechnol. Biochem. 2016, 80, 1580–1586. [Google Scholar] [CrossRef] [Green Version]
- Baek, Y.; Lee, M.N.; Wu, D.; Pae, M. Luteolin Improves Insulin Resistance in Postmenopausal Obese Mice by Altering Macrophage Polarization. Curr. Dev. Nutr. 2019, 13, FS12–FS19. [Google Scholar] [CrossRef]
- Rauter, A.P.; Martins, A.; Borges, C.; Mota-Filipe, H.; Pinto, R.; Sepodes, B.; Justino, J. Antihyperglycaemic and protective effects of flavonoids on streptozotocin-induced diabetic rats. Phytother. Res. 2010, 24, S133–S138. [Google Scholar] [CrossRef] [Green Version]
- Kim, E.K.; Kwon, K.B.; Song, M.Y.; Han, M.J.; Lee, J.H.; Lee, Y.R.; Park, J.W. Flavonoids protect against cytokine-induced pancreatic beta-cell damage through suppression of nuclear factor kappaB activation. Pancreas 2007, 35, e1–e9. [Google Scholar] [CrossRef]
- Zang, M.; Xu, S.; Maitland-Toolan, K.A.; Zuccollo, A.; Hou, X.; Jiang, B.; Cohen, R.A. Polyphenols stimulate AMP-activated protein kinase, lower lipids, and inhibit accelerated atherosclerosis in diabetic LDL receptor-deficient mice. Diabetes 2006, 5, 2180–2191. [Google Scholar] [CrossRef] [Green Version]
- Priscilla, D.H.; Roy, D.; Suresh, A.; Kumar, V.; Thirumurugan, K. Naringenin inhibits alpha-glucosidase activity: A promising strategy for the regulation of postprandial hyperglycemia in high fat diet fed streptozotocin induced diabetic rats. Chem. Biol. Interact. 2014, 210, 77–85. [Google Scholar] [CrossRef]
- Zygmunt, K.; Faubert, B.; MacNeil, J.; Tsiani, E. Naringenin, a citrus flavonoid, increases muscle cell glucose uptake via AMPK. Biochem. Biophys. Res. Commun. 2010, 398, 178–183. [Google Scholar] [CrossRef]
- Cushnie, T.P.; Lamb, A.J. Antimicrobial activity of flavonoids. Int. J. Antimicrob. Agents. 2005, 26, 343–356. [Google Scholar] [CrossRef]
- Martin, A.E.; Montgomery, P.A. Acarbose: An alpha-glucosidase inhibitor. Am. J. Health Syst Pharm. 1996, 53, 2277–2337. [Google Scholar] [CrossRef]
- Ortiz-Andrade, R.R.; Garcia-Jimenez, S.; Castillo-Espana, P.; Ramirez-Avila, G.; Vila-lobosMolina, R.; Estrada-Soto, S. Alpha-glucosidase inhibitory activity of the methanolic extract from Tournefortia hartwegiana: An anti-hyperglycemic agent. J. Ethnopharmacol. 2007, 109, 48–53. [Google Scholar] [CrossRef]
- Shang, Q.; Xiang, J.F.; Tang, Y.L. Screening α-glucosidase inhibitors from mulberry extracts via DOSY and relaxation-edited NMR. Talanta 2012, 97, 362–367. [Google Scholar] [CrossRef]
- Dipeptidyl Peptidase IV (DPP IV) Inhibitors. Available online: https://www.ncbi.nlm.nih.gov/books/NBK542331/ (accessed on 22 January 2022).
- Paliwal, G.; Sharma, A.; Upadhyay, N.; Das, M.; Tiwari, A. Therapeutic stimulation of glp-1 protein by implementing in silico to in vitro approach for type-2 diabetes treatment. Middle East J. Sci. Res. 2015, 23, 1005–1011. [Google Scholar]
- Ng, V.W.S.; Glasg, F. Dipeptidyl peptidase (dpp) -iv inhibitor: A novel class of oral anti-hyperglycemic agents. Clin. Pharmacol. Ther. 2007, 11, 33–34. [Google Scholar]
- Rahman, M.S.; Hossain, K.S.; Das, S.; Kundu, S.; Adegoke, E.O.; Rahman, M.A.; Hannan, M.A.; Uddin, M.J.; Pang, M.G. Role of Insulin in Health and Disease: An Update. Int. J. Mol. Sci. 2021, 22, 6403. [Google Scholar] [CrossRef]
- Singh, A.-K.; Patel, P.K.; Choudhary, K.; Joshi, J.; Yadav, D.; Jin, J.-O. Quercetin and Coumarin Inhibit Dipeptidyl Peptidase-IV and Exhibits Antioxidant Properties: In Silico, In Vitro, Ex Vivo. Biomolecules 2020, 10, 207. [Google Scholar] [CrossRef] [Green Version]
- Huang, P.K.; Lin, S.R.; Chang, C.H.; Tsai, M.J.; Lee, D.N.; Weng, C.F. Natural phenolic compounds potentiate hypoglycemia via inhibition of Dipeptidyl peptidase IV. Sci. Rep. 2019, 9, 15585. [Google Scholar] [CrossRef]
- Rena, G.; Hardie, D.G.; Pearson, E.R. The mechanisms of action of metformin. Diabetologia 2017, 60, 1577–1585. [Google Scholar] [CrossRef] [Green Version]
- Boesch-Saadatmandi, C.; Loboda, A.; Wagner, A.E.; Stachurska, A.; Jozkowicz, A.; Dulak, J.; Döring, F.; Wolffram, S.; Rimbach, G. Effect of quercetin and its metabolites isorhamnetin and quercetin-3-glucuronide on inflammatory gene expression: Role of miR-155. J. Nutr. Biochem. 2011, 22, 293–299. [Google Scholar] [CrossRef]
- Ueda-Wakagi, M.; Mukai, R.; Fuse, N.; Mizushina, Y.; Ashida, H. 3-O-Acyl-epicatechins Increase Glucose Uptake Activity and GLUT4 Translocation through Activation of PI3K Signaling in Skeletal Muscle Cells. Int. J. Mol. Sci. 2015, 16, 16288–16299. [Google Scholar] [CrossRef] [Green Version]
- Prasad, C.N.; Anjana, T.; Banerji, A.; Gopalakrishnapillai, A. Gallic acid induces GLUT4 translocation and glucose uptake activity in 3T3-L1 cells. FEBS Lett. 2010, 584, 531–536. [Google Scholar] [CrossRef] [Green Version]
- Lee, H.; Kim, J.W. High-dose Resveratrol Inhibits Insulin Signaling Pathway in 3T3-L1 Adipocytes. J. Lifestyle Med. 2013, 3, 41–47. [Google Scholar] [PubMed]
- Katashima, C.K.; Silva, V.R.R.; Lenhare, L.; Marin, R.M.; Carvalheira, J.B.C. iNOS promotes hypothalamic insulin resistance associated with deregulation of energy balance and obesity in rodents. Sci. Rep. 2017, 7, 9265. [Google Scholar] [CrossRef] [PubMed]
- Aggarwal, H.; Pathak, P.; Singh, P.; Gayen, J.R.; Jagavelu, K.; Dikshit, M. Systemic Insulin Resistance and Metabolic Perturbations in Chow Fed Inducible Nitric Oxide Synthase Knockout Male Mice: Partial Reversal by Nitrite Supplementation. Antioxidants 2020, 9, 736. [Google Scholar] [CrossRef] [PubMed]
- Hatami, T.; Emami, S.A.; Miraghaee, S.S.; Mojarrab, M. Total Phenolic Contents and Antioxidant Activities of Different Extracts and Fractions from the Aerial Parts of Artemisia biennis Willd. Iran. J. Pharm. Res. 2014, 13, 551–559. [Google Scholar]
- Jia, Z.; Tang, M.; Wu, J. The determination of flavonoid contents in mulberry and their scavenging effects on superoxides radicals. Food Chem. 1998, 64, 555–559. [Google Scholar]
- Bachhawat, A.; Shihabudeen, M.S.; Thirumurugan, K. Screening of fifteen Indian Ayurvedic plants for alpha-glucosidase inhibitory activity and enzyme kinetics. Int. J. Pharm Pharm Sci. 2011, 3, 267–274. [Google Scholar]
- Al-Masri, I.M.; Mohammad, M.K.; Tahaa, M.O. Inhibition of dipeptidyl peptidase-IV (DPP-IV) is one of the mechanisms explaining the hypoglycemic effect of berberine. J. Enzym Inhib. Med. Chem. 2009, 24, 1061–1066. [Google Scholar] [CrossRef]
- Choi, J.S.; Nurul Islam, M.; Yousof Ali, M.; Kim, E.J.; Kim, Y.M.; Jung, H.A. Effects of C-glycosylation on anti-diabetic, antiAlzheimer’s disease and anti-inflammatory potential of apigenin. Food Chem. Toxicol. 2014, 64, 27–33. [Google Scholar] [CrossRef]
- Vishwanath, D.; Srinivasan, H.; Patil, M.S.; Seetarama, S.; Agrawal, S.K.; Dixit, M.N.; Dhar, K. Novel method to differentiate 3T3 L1 cells in vitro to produce highly sensitive adipocytes for a GLUT4 mediated glucose uptake using fluorescent glucose analog. J. Cell Commun. Signal. 2013, 7, 129–140. [Google Scholar] [CrossRef] [Green Version]
- Sudsai, T.; Wattanapiromsakul, C.; Nakpheng, T.; Tewtrakul, S. Evaluation of the wound healing property of Boesenbergia longiflora rhizomes. J. Ethnopharmacol. 2013, 150, 223–231. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Extracts | % Yield Extraction | TPC mg GAE/g Extract | TFC mg QE/g Extract |
|---|---|---|---|
| Crude | 10.30 | 562.33 ± 2.89 | 79.70 ± 0.72 |
| BsH | 0.40 | 12.33 ± 0.76 | 55.10 ± 0.00 |
| BsD | 8.15 | 210.67 ± 1.88 | 65.52 ± 0.72 |
| BsE | 5.90 | 620.67 ± 0.99 * | 131.35 ± 2.50 * |
| BsW | 60.00 | 392.33 ± 1.65 | 71.77 ± 0.72 |
| Compounds | Formula | Retention Time (min) | Mode of Ionization (ESI−/ESI+) | Mass | m/z | Mass Error (ppm) |
|---|---|---|---|---|---|---|
| Flavonoids | ||||||
| 1. Epigallocatechin | C15H14O7 | 12.54 | [M-H]− | 306.07 | 305.07 | 0.02 |
| 2. Catechin | C15H14O6 | 13.72 | [M-H]− | 290.07 | 289.07 | −0.05 |
| 3. Apuleirin | C20H20O9 | 15.00 | [M-H]− | 404.11 | 403.10 | −0.40 |
| 4. Phloridzin | C21H24O10 | 15.11 | [M-H]− | 436.13 | 435.13 | −0.05 |
| 5. Epicatechin | C15H14O6 | 16.35 | [M-H]− | 290.07 | 289.07 | −0.57 |
| 6. Astilbin | C21H22O11 | 16.71 | [M-H]− | 450.11 | 449.11 | −0.39 |
| 7. Rhapontin | C21H24O9 | 18.86 | [M-H]− | 420.14 | 419.13 | −0.29 |
| 8. Quercetin3-(2-galloylglucoside) | C28H24O16 | 19.41 | [M-H]− | 616.11 | 615.10 | 0.18 |
| 9. Quercetin 3-galactoside | C21H20O12 | 20.68 | [M-H]− | 464.09 | 463.08 | −1.01 |
| 10. Epicatechin Monogallate | C22H18O10 | 21.12 | [M-H]− | 442.09 | 441.08 | −0.08 |
| 11. Taxifolin | C15H12O7 | 21.70 | [M-H]− | 304.05 | 303.05 | −0.25 |
| 12. Kaempferol−7-o-glucoside | C21H20O11 | 22.21 | [M-H]− | 448.10 | 447.09 | −1.07 |
| 13. Naringenin | C15H12O5 | 23.53 | [M-H]− | 272.07 | 271.06 | −0.19 |
| 14. Alphitonin | C15H12O7 | 24.23 | [M-H]− | 304.06 | 303.05 | −0.17 |
| 15. Apigenin 7-O-glucoside | C21H20O10 | 26.12 | [M-H]− | 432.11 | 431.11 | −0.46 |
| 16. Phloretin | C15H14O5 | 28.84 | [M-H]− | 274.08 | 273.08 | −0.58 |
| 17. Agehoustin C | C22H24O10 | 31.16 | [M-H]− | 448.13 | 447.13 | 0.31 |
| 18. Theasinensin C | C30H26O14 | 32.04 | [M-H]− | 610.13 | 609.12 | 3.00 |
| 19. Luteolin | C15H10O6 | 32.62 | [M-H]− | 286.04 | 285.04 | −0.27 |
| 20. Quercetin | C15H10O7 | 32.92 | [M-H]− | 302.04 | 301.04 | −0.12 |
| 21. Dichamanetin | C29H24O6 | 33.62 | [M+HCOO]− | 468.16 | 513.15 | 0.28 |
| 22. Viniferal | C35H26O8 | 35.07 | [M+HCOO]− | 574.17 | 633.18 | −2.57 |
| 23. Cnidilin | C17H16O5 | 35.65 | [M-H]− | 300.10 | 299.09 | 0.09 |
| 24. Apigenin | C15H10O5 | 36.89 | [M-H]− | 270.05 | 269.04 | 0.14 |
| 25. Kaempferol | C15H10O6 | 37.90 | [M-H]− | 286.05 | 285.04 | −0.17 |
| Phenolics | ||||||
| 26. Gallic acid | C7H6O5 | 4.92 | [M-H]− | 170.22 | 169.01 | −0.20 |
| 27. Caffeic acid | C9H8O4 | 10.58 | [M-H]− | 180.04 | 179.03 | 0.06 |
| 28. Gentisic acid | C7H6O4 | 13.19 | [M-H]− | 154.02 | 153.19 | 0.02 |
| 29. Kelampayoside A | C20H30O13 | 13.82 | [M+HCOO]− | 478.19 | 523.17 | −0.08 |
| 30. Pyrocatechol | C23H34O14 | 15.41 | [M-H]− | 110.03 | 109.03 | −0.04 |
| 31. Irisxanthone | C20H20O11 | 22.63 | [M-H]− | 436.10 | 435.09 | −0.28 |
| 32. Resveratrol | C14H12O3 | 22.70 | [M-H]− | 228.07 | 227.07 | −0.53 |
| 33. Isosyringinoside | C23H34O14 | 23.43 | [M-H]− | 534.19 | 533.19 | 0.56 |
| Samples | IC50 | |
|---|---|---|
| α-Glucosidase (µg/mL) | DPP-IV (µg/mL) | |
| Crude extract | 2.37 ± 0.13 a | >50 |
| BsH | 10.61 ± 0.93 a | >50 |
| BsD | 10.09 ± 0.75 a | >50 |
| BsE | 1.51 ± 0.01 a | 2.62 ± 0.03 a |
| BsW | 2.42 ± 0.10 a | 3.20 ± 0.02 a |
| Resveratrol | 1.41 ± 0.02 a (8.16 µM) | 3.22 ± 0.02 a (14.11 µM) |
| Epicatechin | >7.26 (25 µM) | >14.52 (50 µM) |
| Quercetin | 1.89 ± 0.11a (6.26 µM) | 2.49 ± 0.01a (8.25 µM) |
| Gallic acid | >4.25 (25 µM) | >8.50 (50 µM) |
| Acarbose | 329.48 ± 6.91 b (509.60 µM) | - |
| Diprotin A | - | 1.14 ± 0.05 a (3.26 µM) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Praparatana, R.; Maliyam, P.; Barrows, L.R.; Puttarak, P. Flavonoids and Phenols, the Potential Anti-Diabetic Compounds from Bauhinia strychnifolia Craib. Stem. Molecules 2022, 27, 2393. https://doi.org/10.3390/molecules27082393
Praparatana R, Maliyam P, Barrows LR, Puttarak P. Flavonoids and Phenols, the Potential Anti-Diabetic Compounds from Bauhinia strychnifolia Craib. Stem. Molecules. 2022; 27(8):2393. https://doi.org/10.3390/molecules27082393
Chicago/Turabian StylePraparatana, Rachanida, Pattaravan Maliyam, Louis R. Barrows, and Panupong Puttarak. 2022. "Flavonoids and Phenols, the Potential Anti-Diabetic Compounds from Bauhinia strychnifolia Craib. Stem." Molecules 27, no. 8: 2393. https://doi.org/10.3390/molecules27082393
APA StylePraparatana, R., Maliyam, P., Barrows, L. R., & Puttarak, P. (2022). Flavonoids and Phenols, the Potential Anti-Diabetic Compounds from Bauhinia strychnifolia Craib. Stem. Molecules, 27(8), 2393. https://doi.org/10.3390/molecules27082393