
Metabolomic Profiling Reveals Protective Effects and Mechanisms of Sea Buckthorn Sterol against Carbon Tetrachloride-Induced Acute Liver Injury in Rats

Abstract
:1. Introduction
2. Materials and Methods
2.1. Animals, Reagents, and Drugs
2.2. Preparations of the Acute Liver Injury Model
2.3. Pathological Examination of Liver Tissue
2.4. Measurement of Biochemical Index in Liver Tissue Homogenate
2.5. Sample Preparation
2.5.1. Extraction of Hydrophilic Compounds
2.5.2. Extraction of Hydrophobic Compounds
2.6. Analysis by UPLC
2.7. Analysis by ESI-Q TRAP-MS/MS
2.7.1. ESI-Q TRAP-MS/MS of Hydrophilic Compounds
2.7.2. ESI-Q TRAP-MS/MS of Hydrophobic Compounds
2.8. Transcriptome Analysis
2.8.1. RNA Detection
2.8.2. Library Setup
Library Quality Check
- (i)
- Qubit2.0 was used for preliminary quantification, and Agilent 2100 was used to detect the insert size of the library. The next experiment could only be carried out after the insert size met expectations;
- (ii)
- The Q-PCR method accurately quantified the effective concentration of the library (the effective concentration of the library was >2 nM). Then, the library check was successfully performed.
Sequencing on the Machine
2.9. Statistical Analysis
3. Results
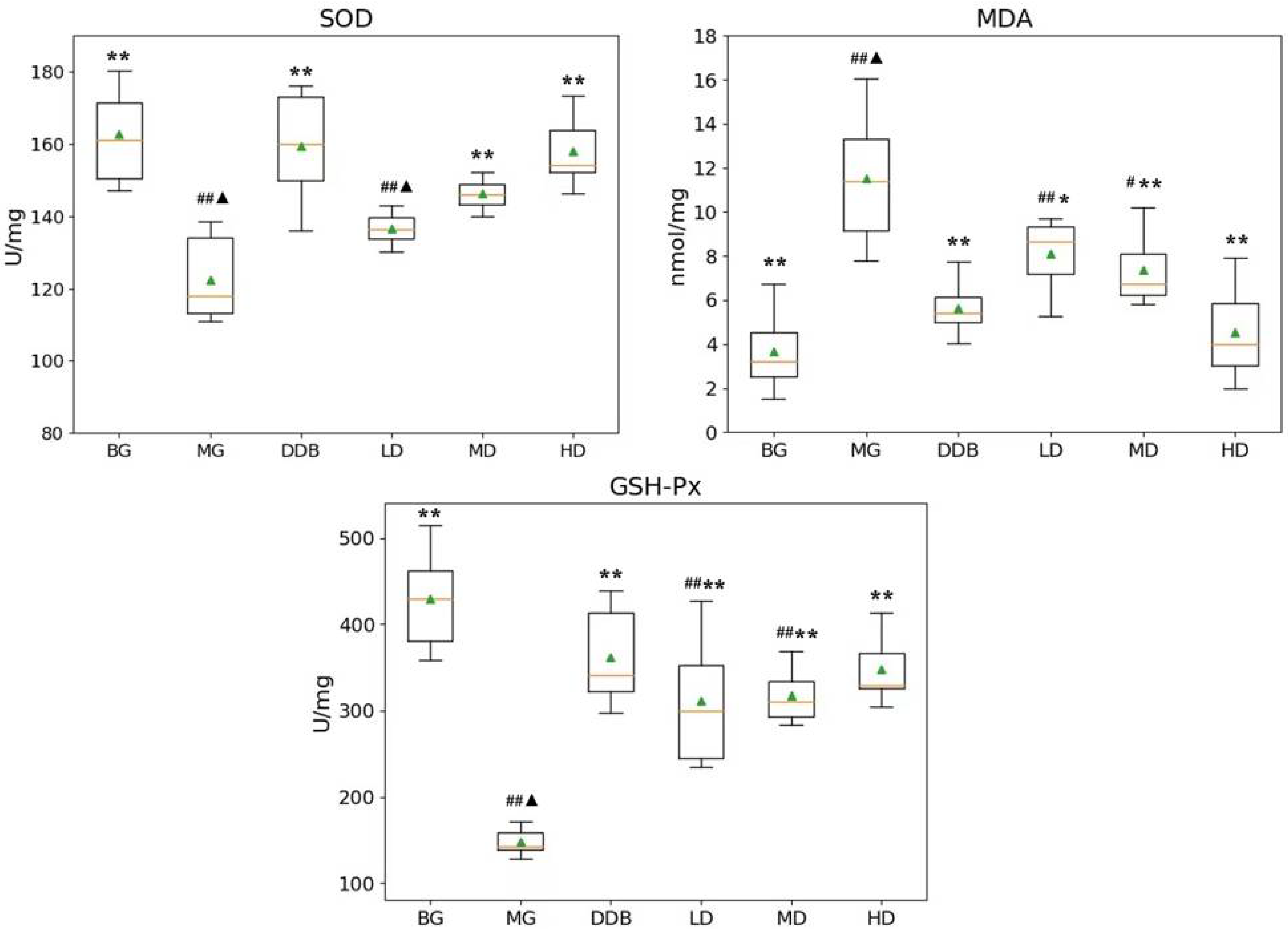
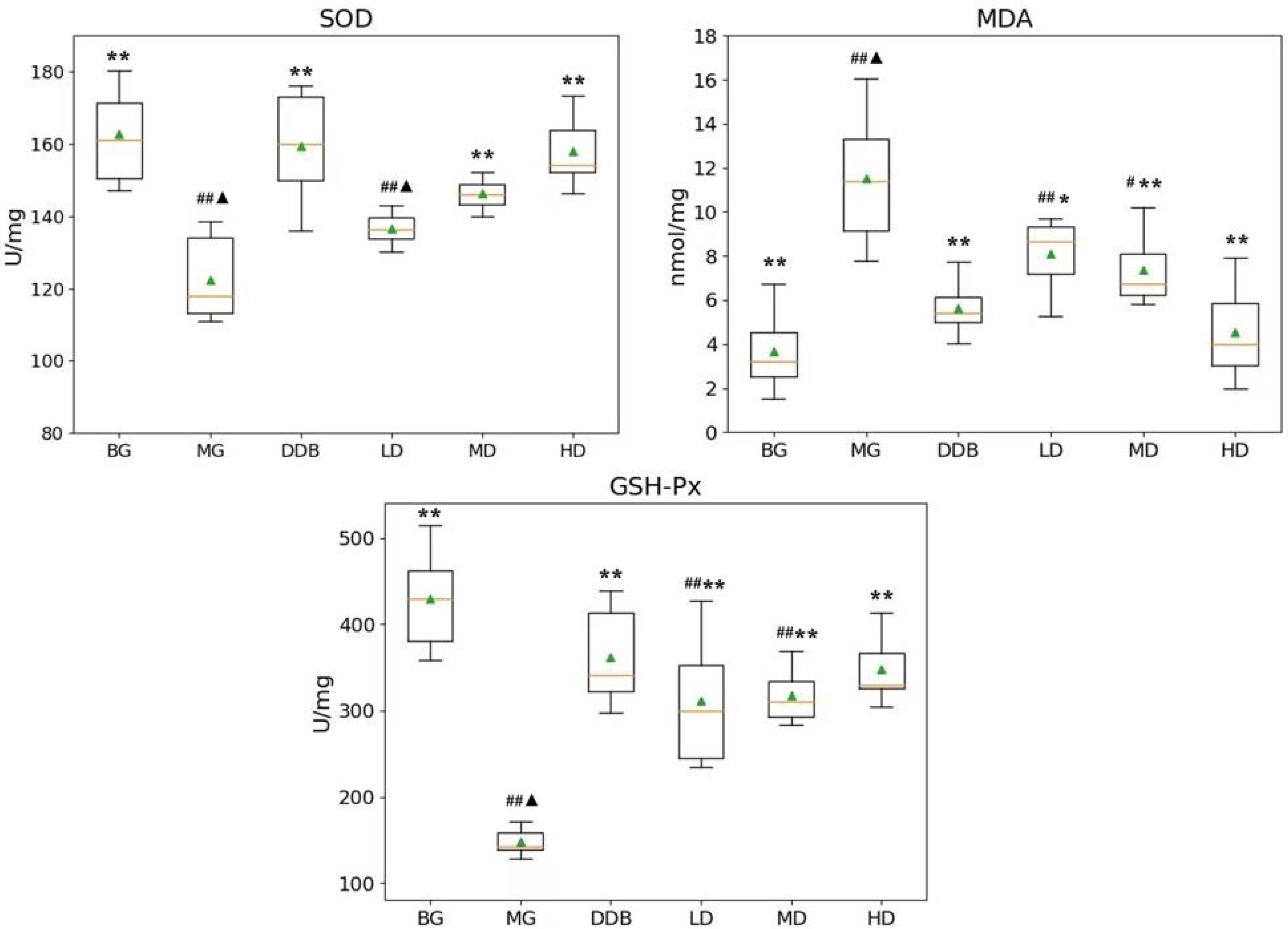
3.1. Effects of SBS on SOD, MDA, and GSH-PX in Rats with CCl4-Induced Liver Injury
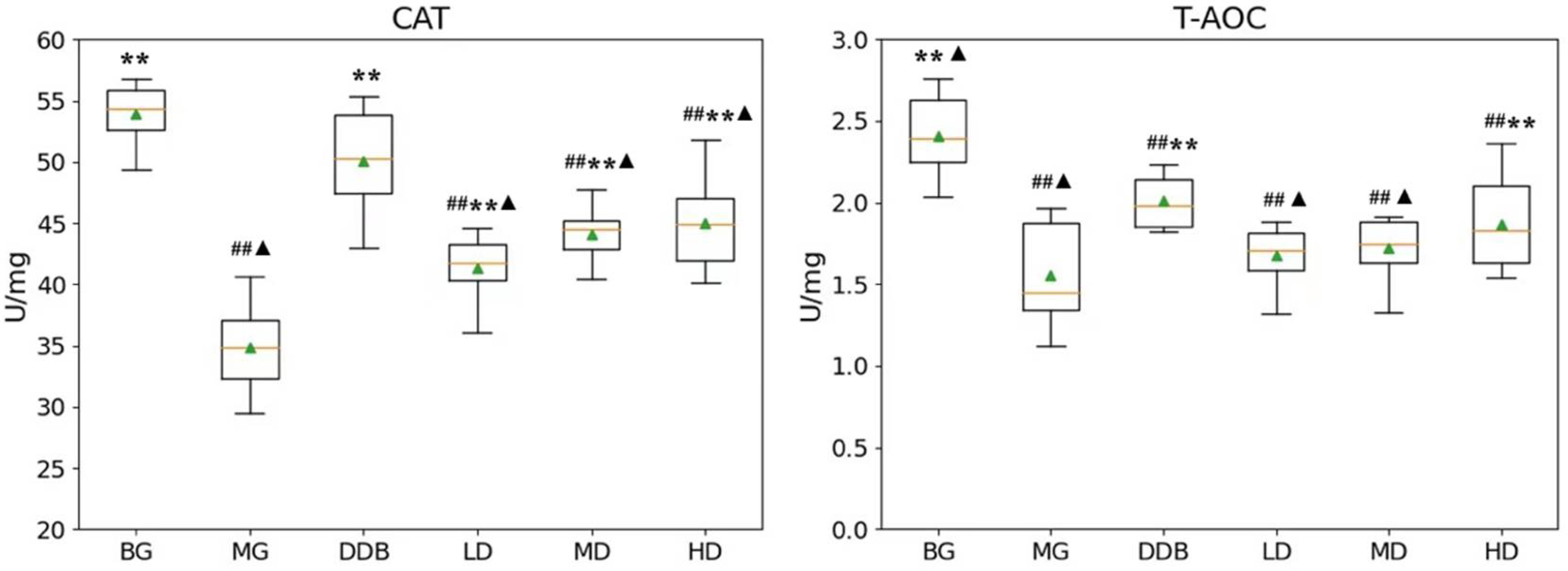
3.2. Effects of SBS on CAT and T-AOC in Rats with CCl4-Induced Liver Injury
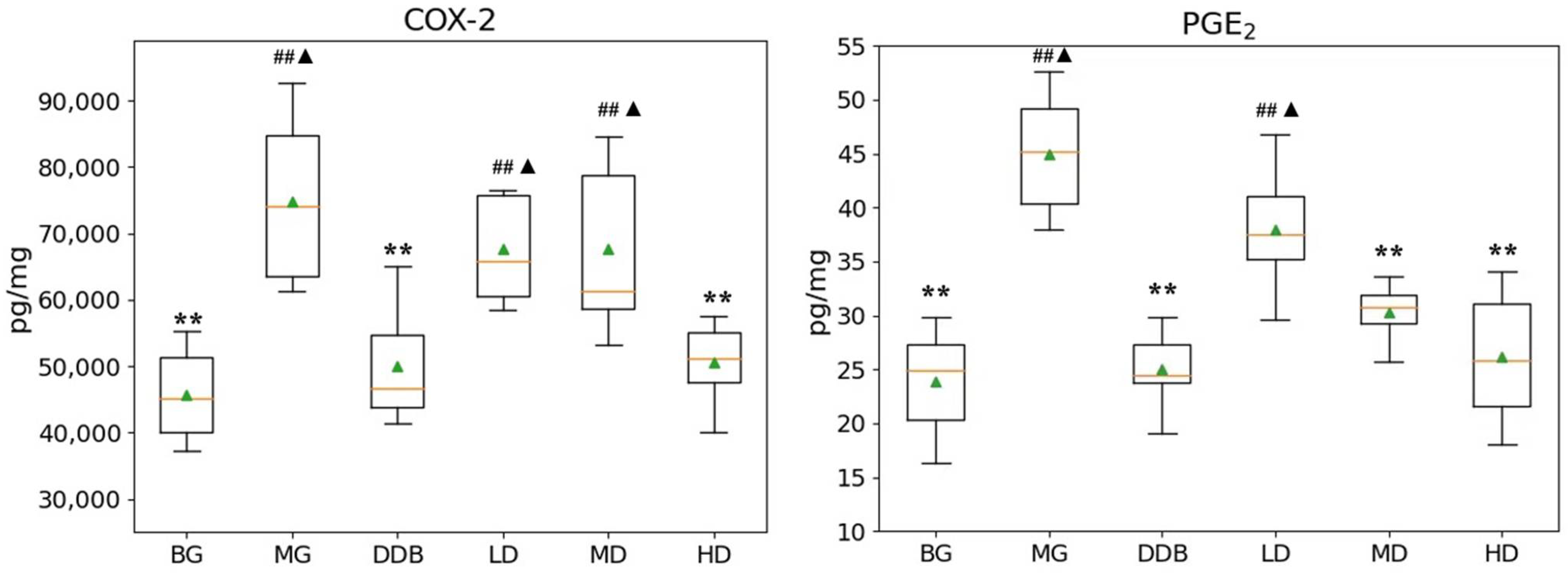
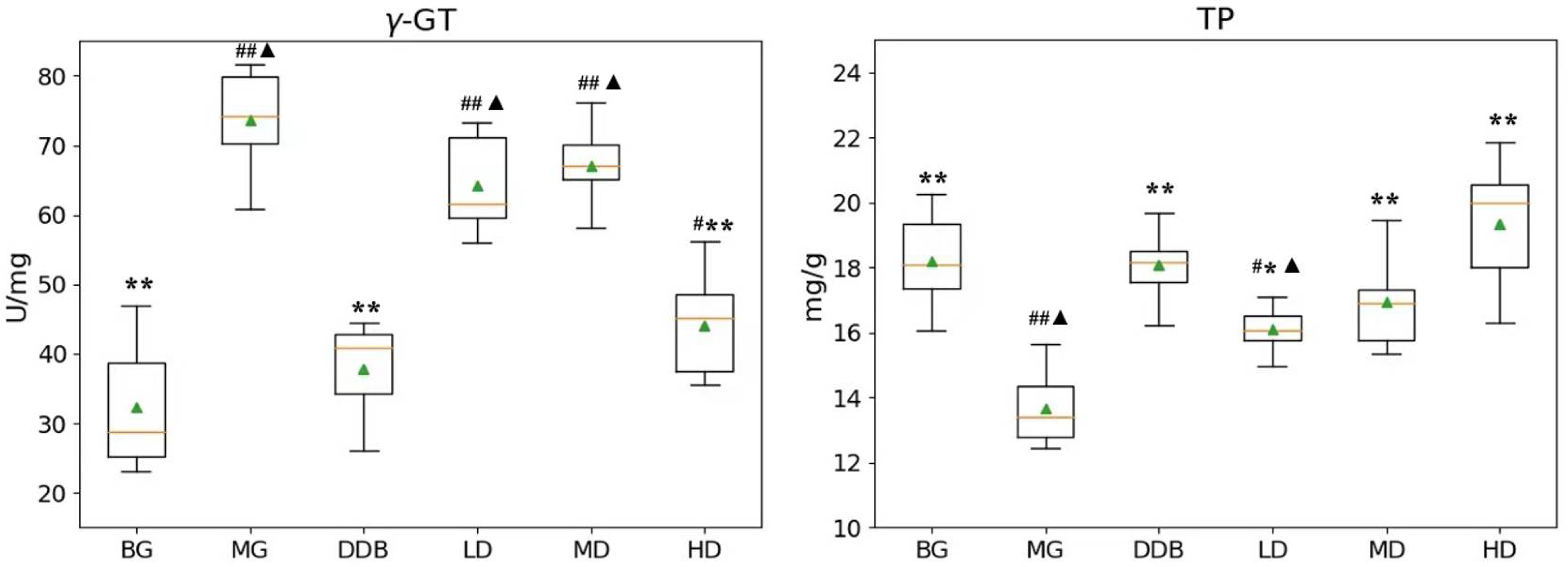
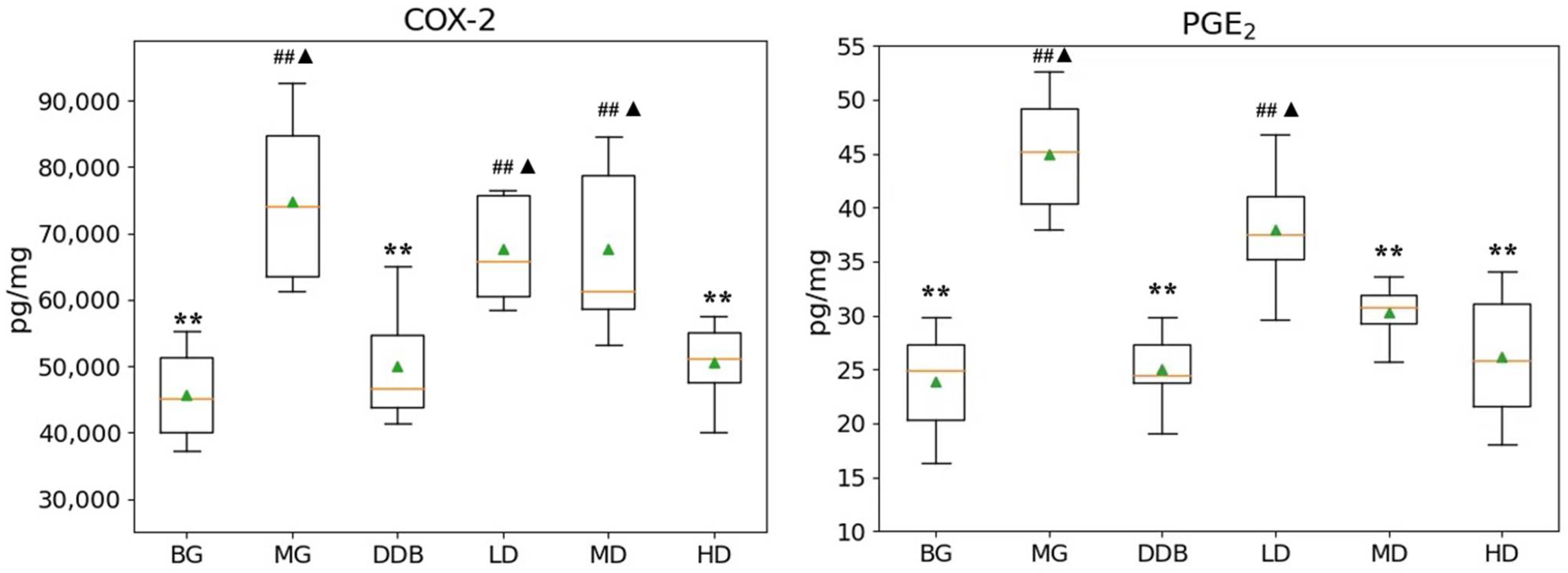
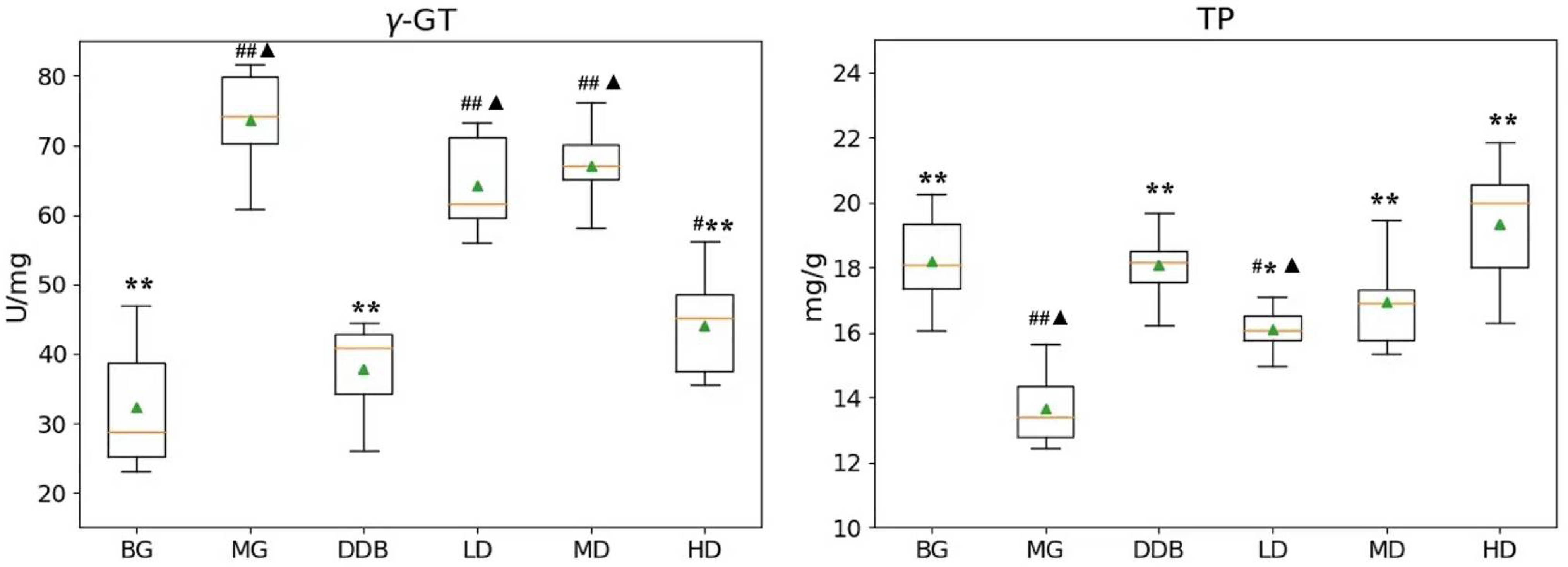
3.3. Effects of SBS on γ-GT, TP, PGE2, and COX-2 in Rats with CCl4-Induced Liver Injury
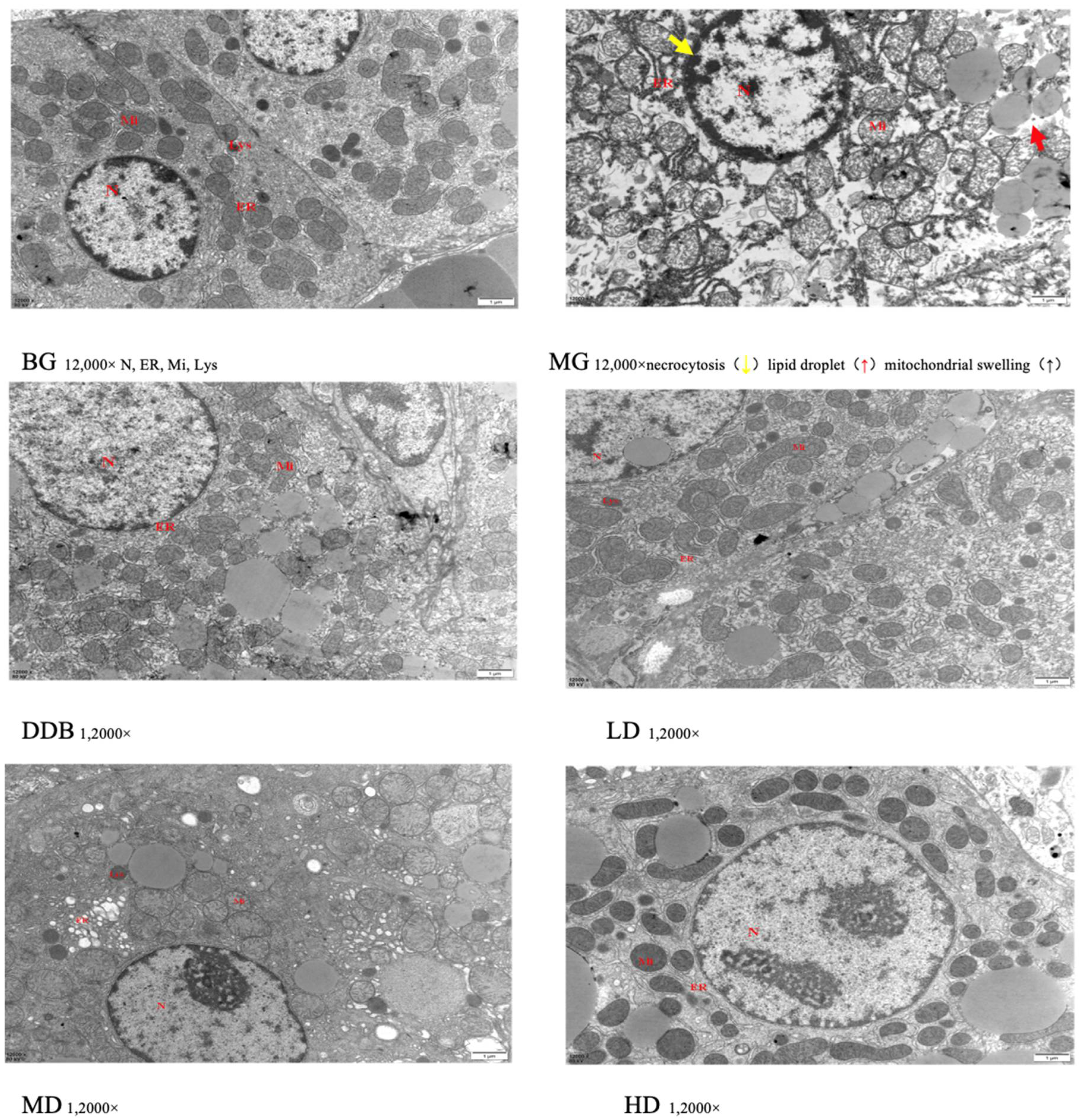
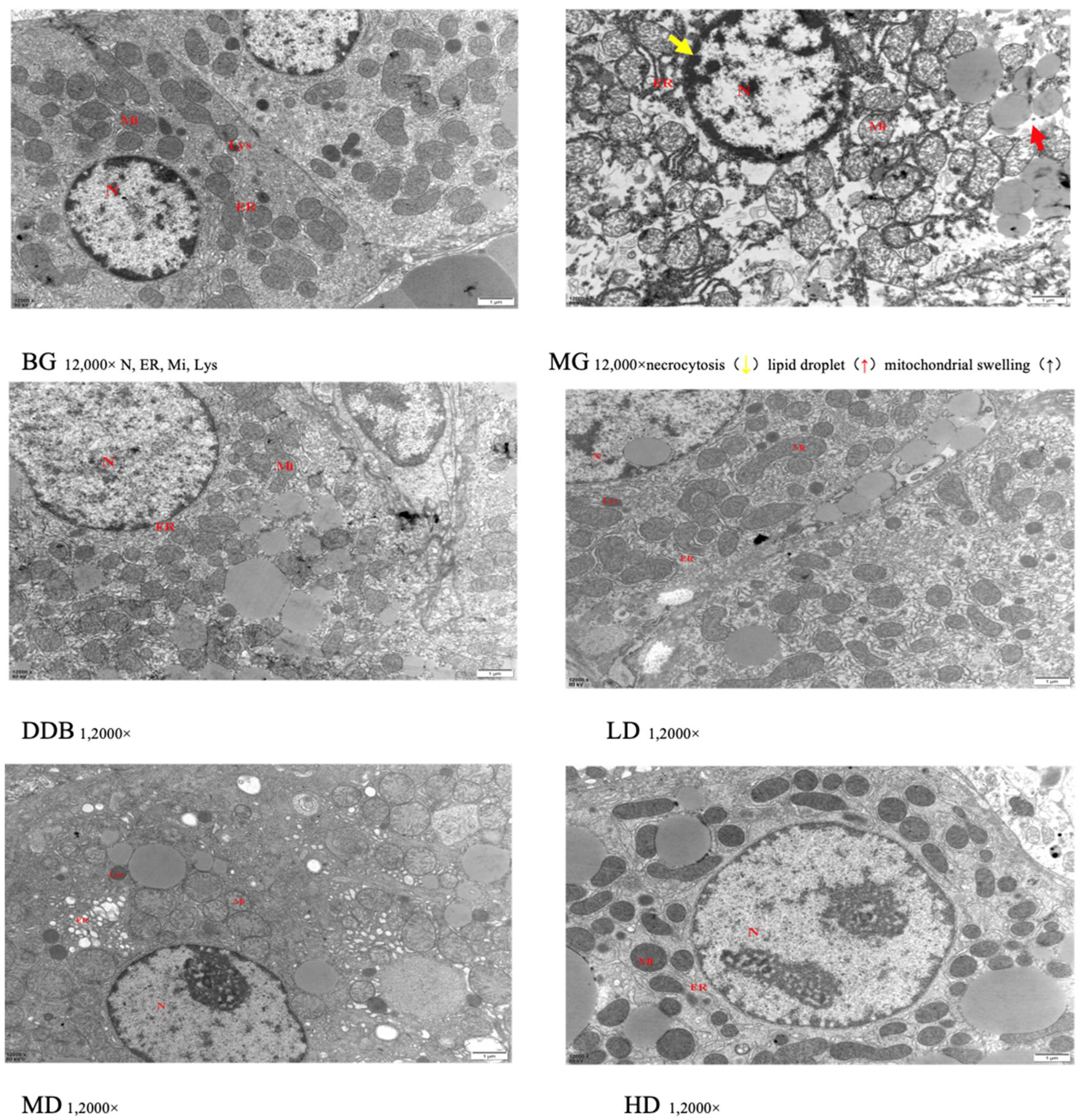
3.4. Electron Microscopic Observations of Liver Tissue
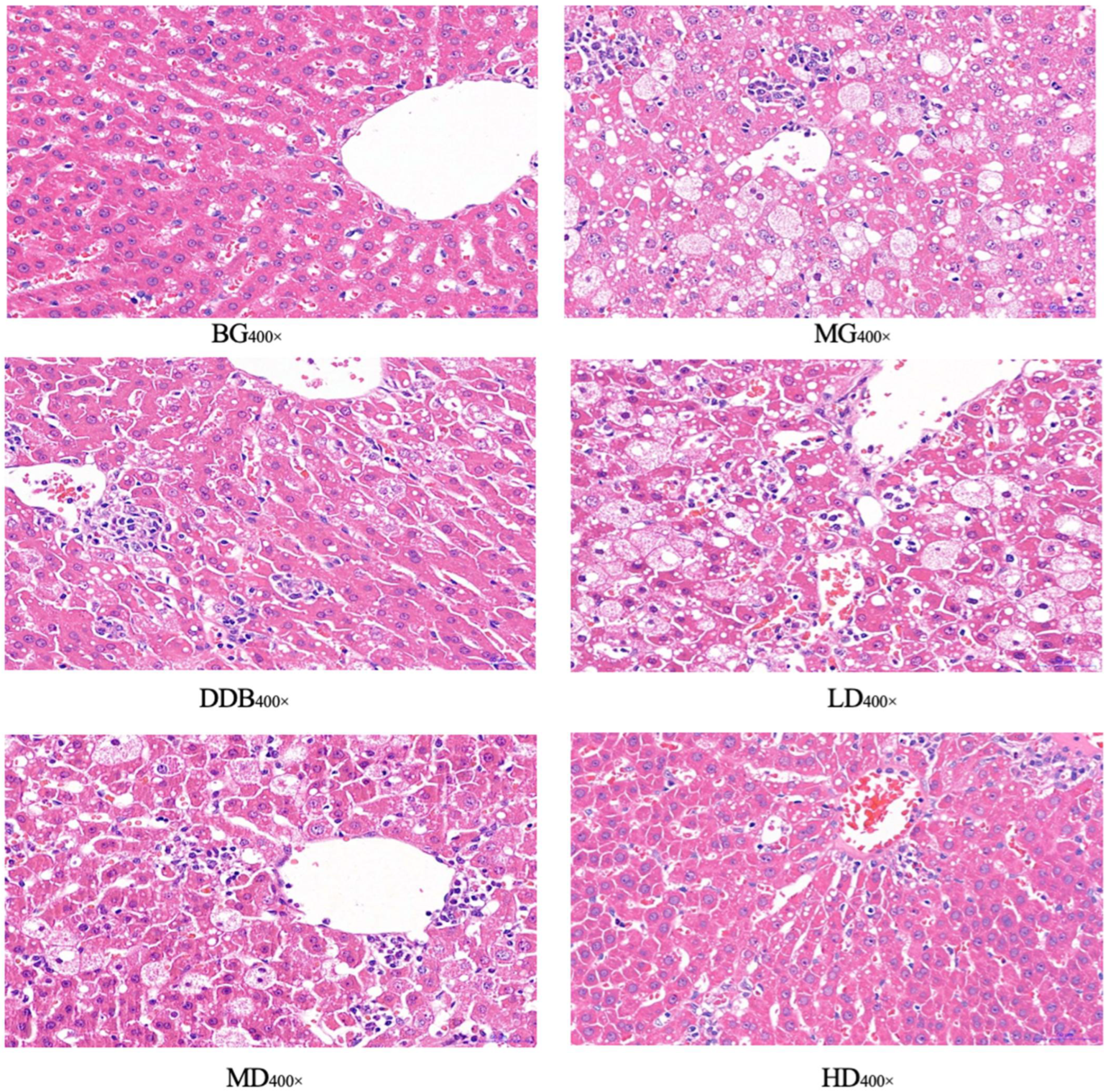
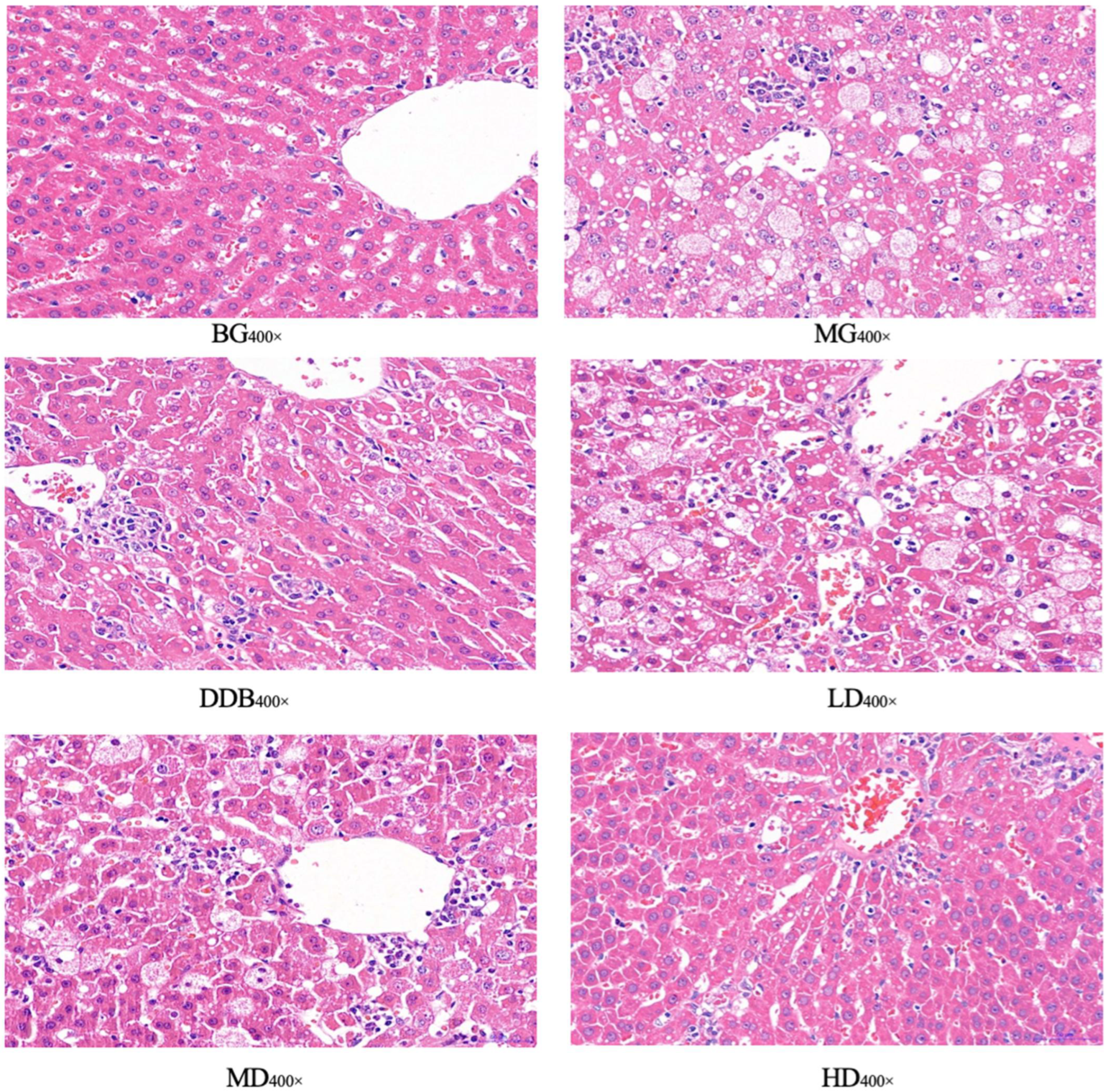
3.5. Light Microscopic Observations of Liver Tissue
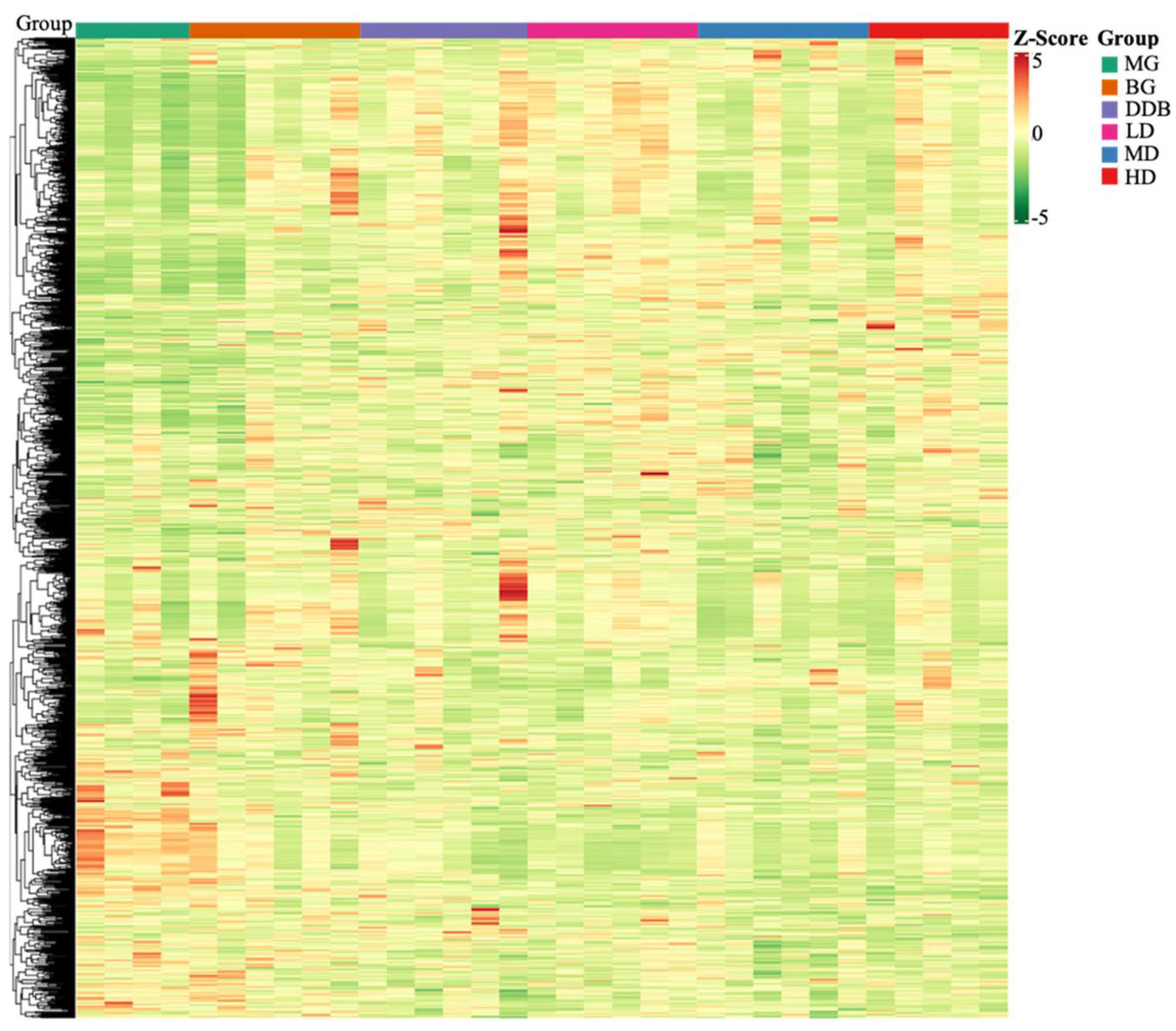
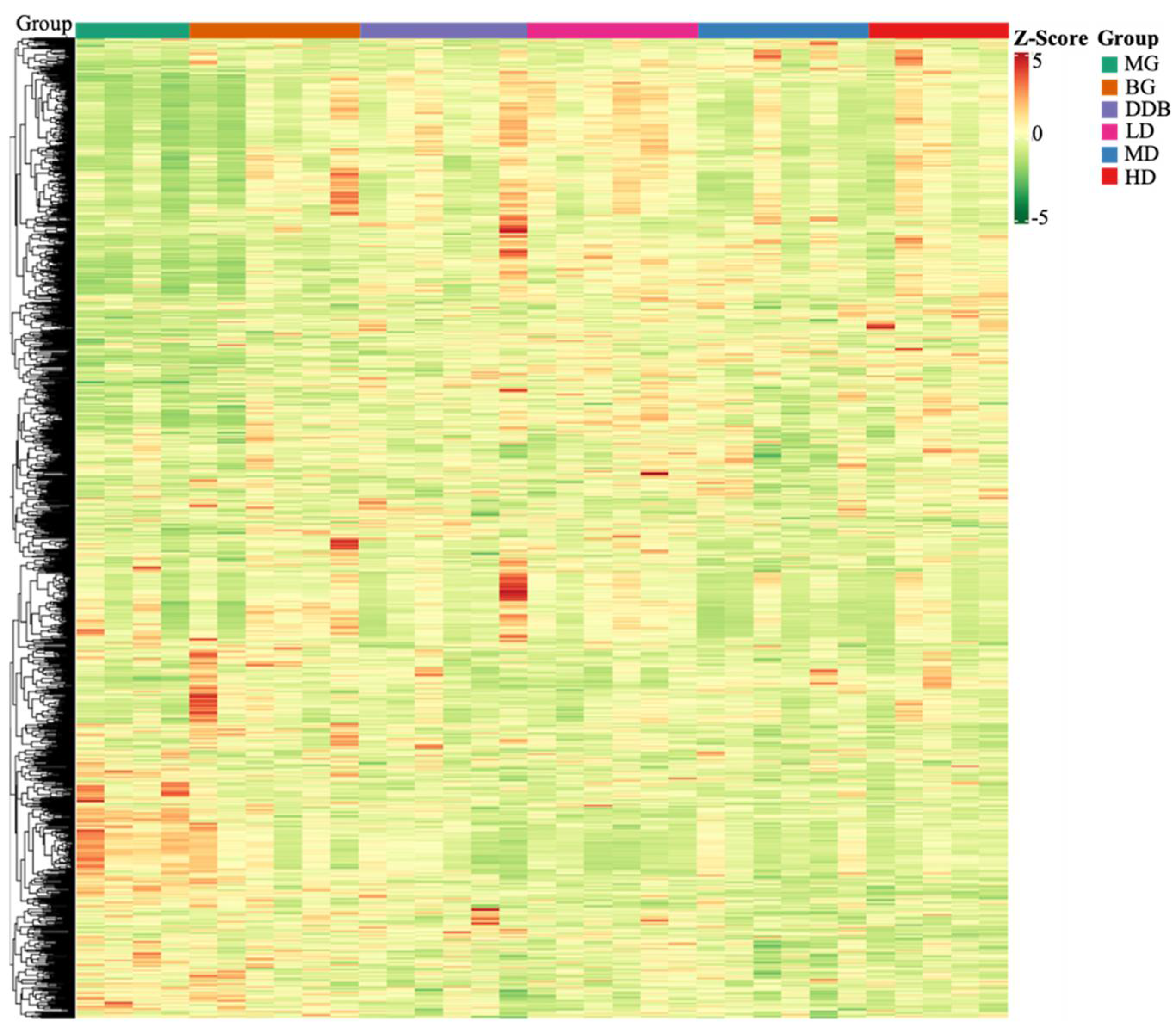
3.6. Identification and Analysis of Metabolites
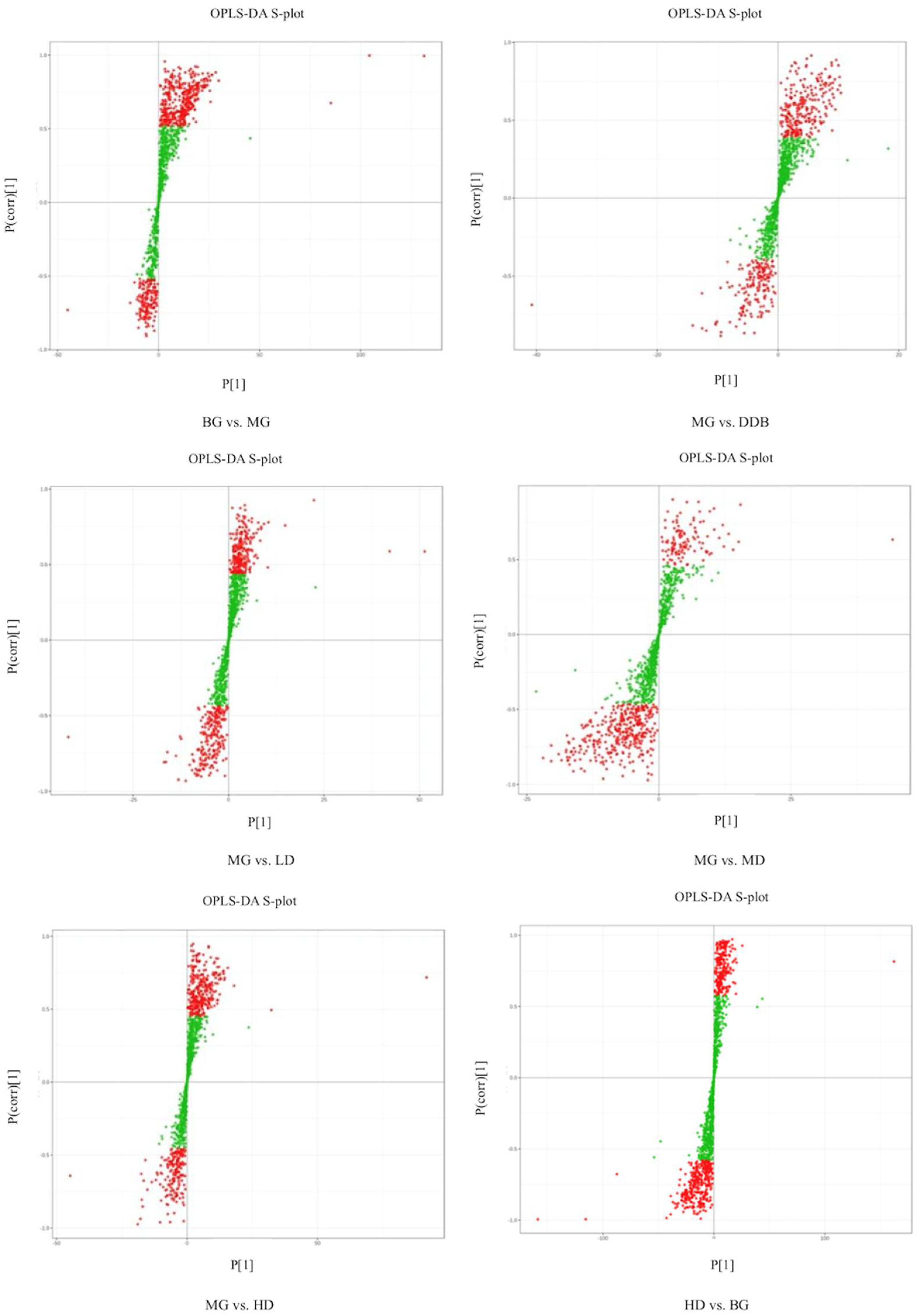
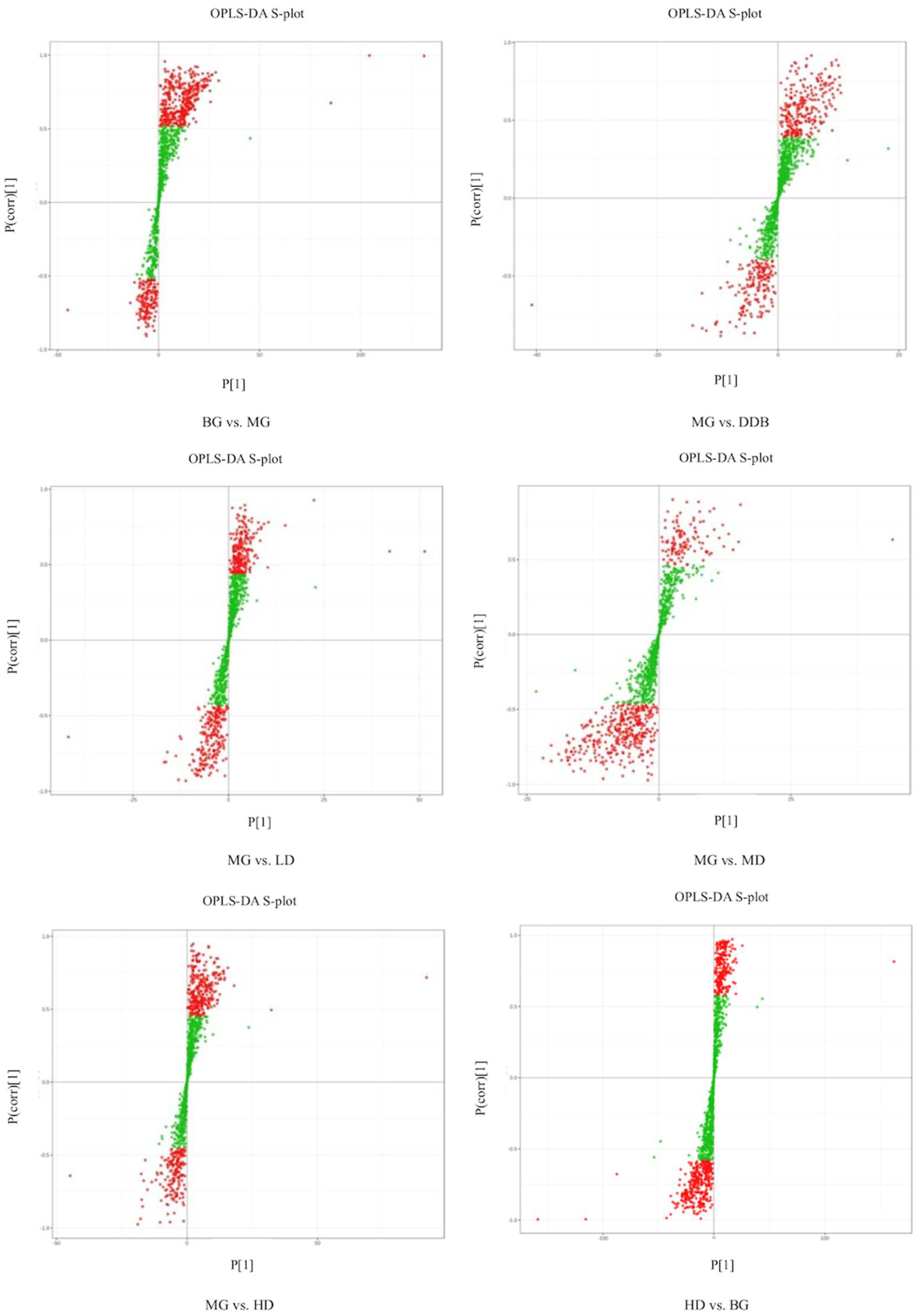
3.6.1. Discriminant Analysis of Orthogonal Partial Least Squares Method
3.6.2. Determination of Relevant Metabolite
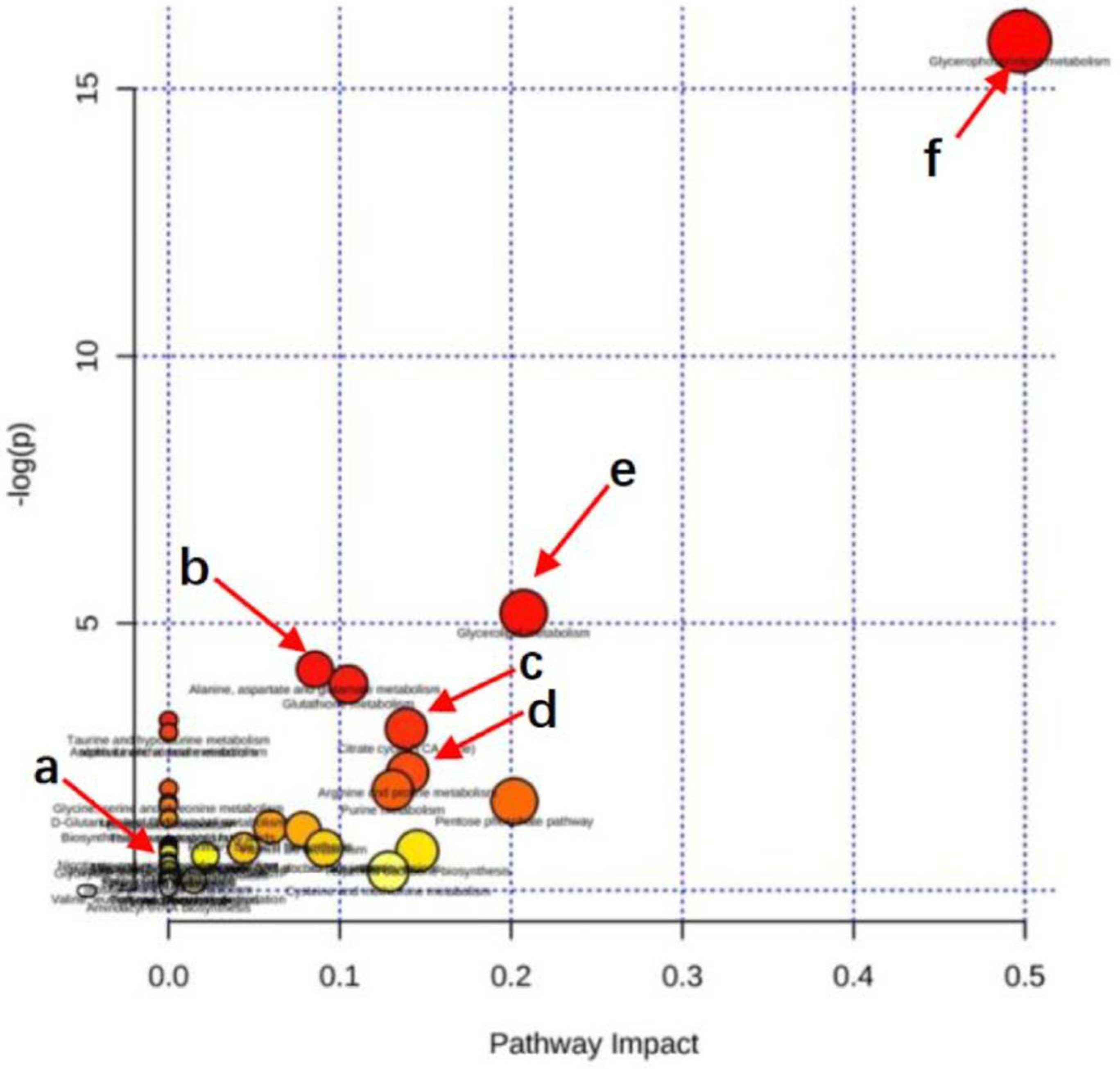
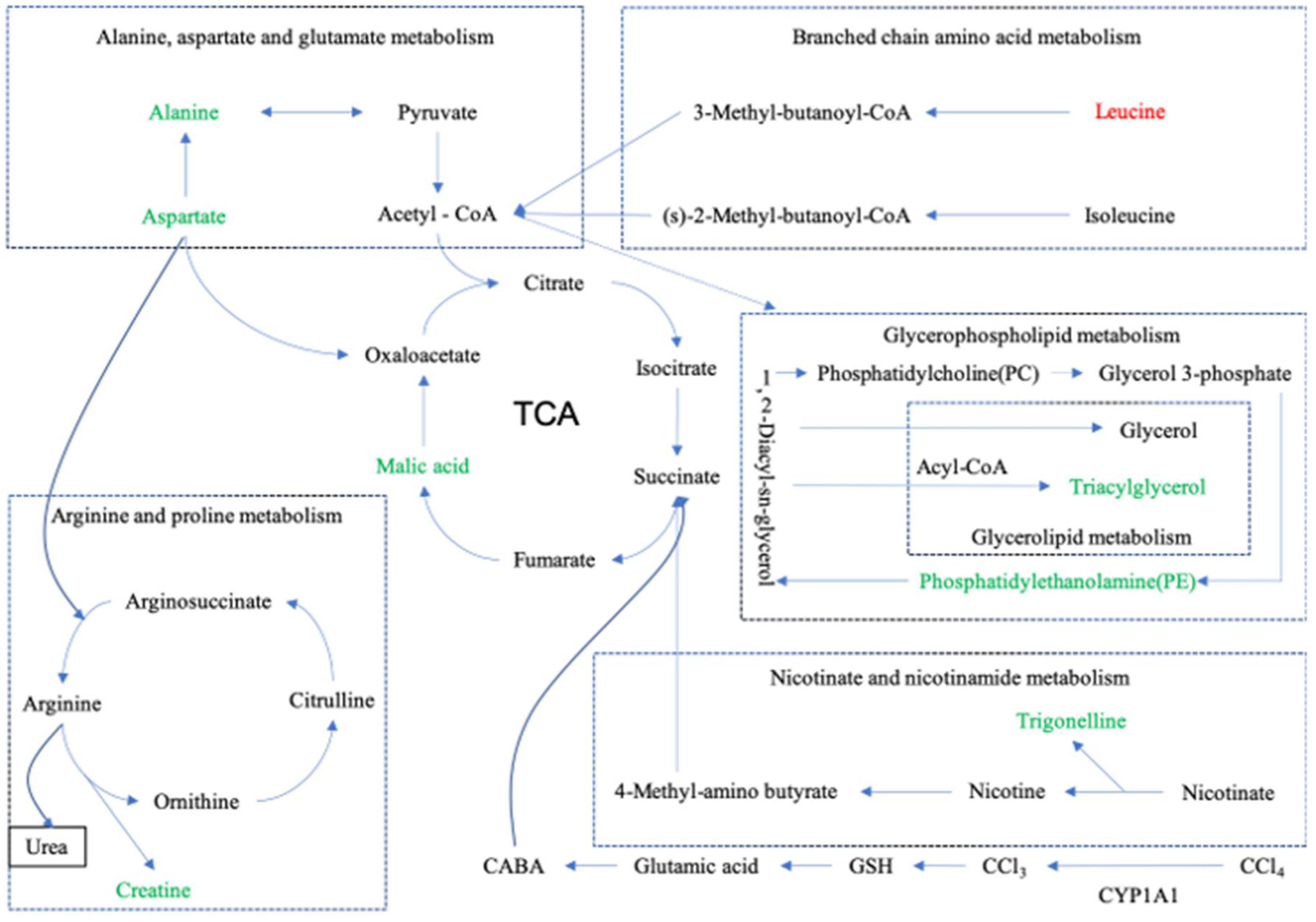
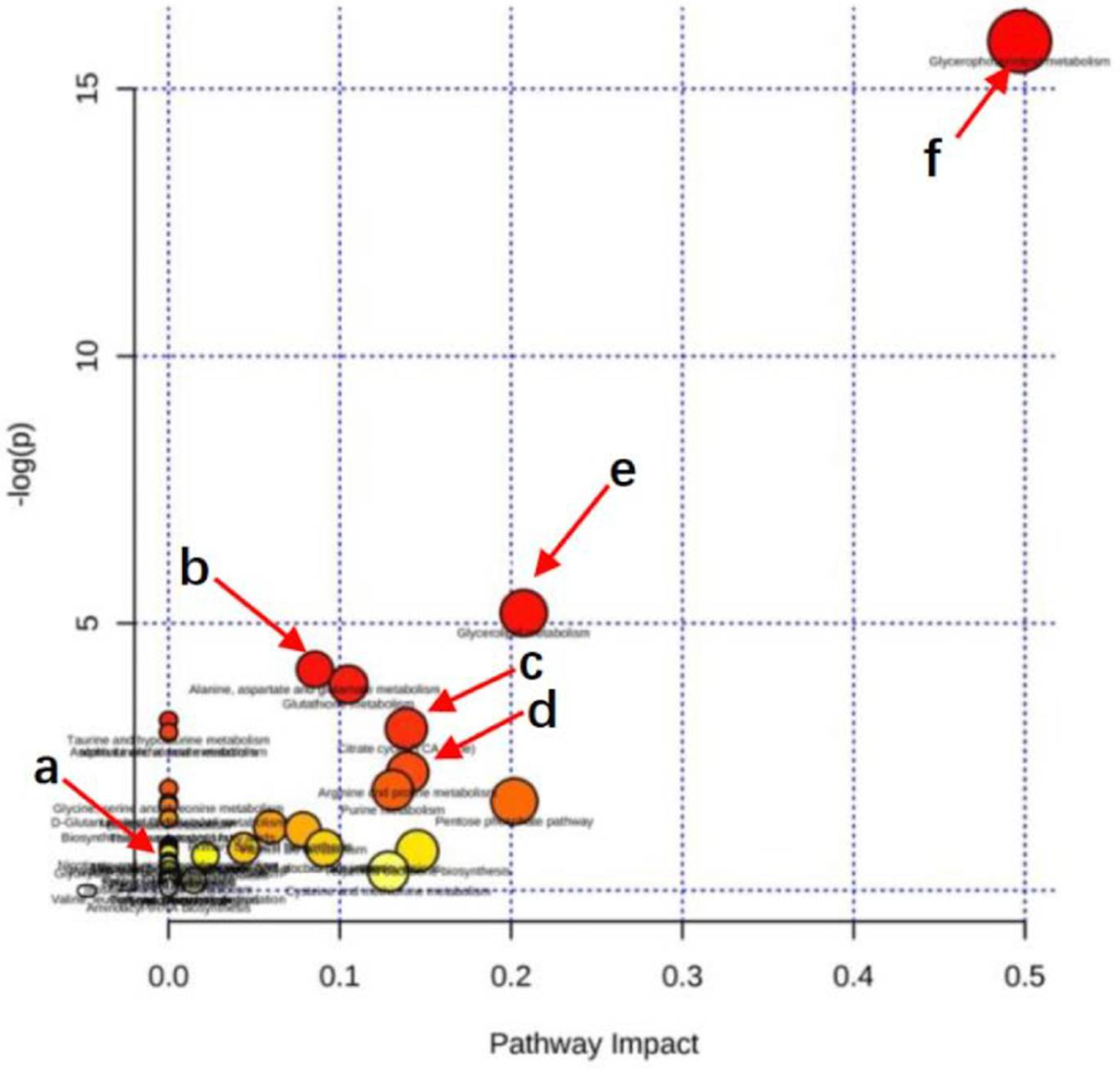
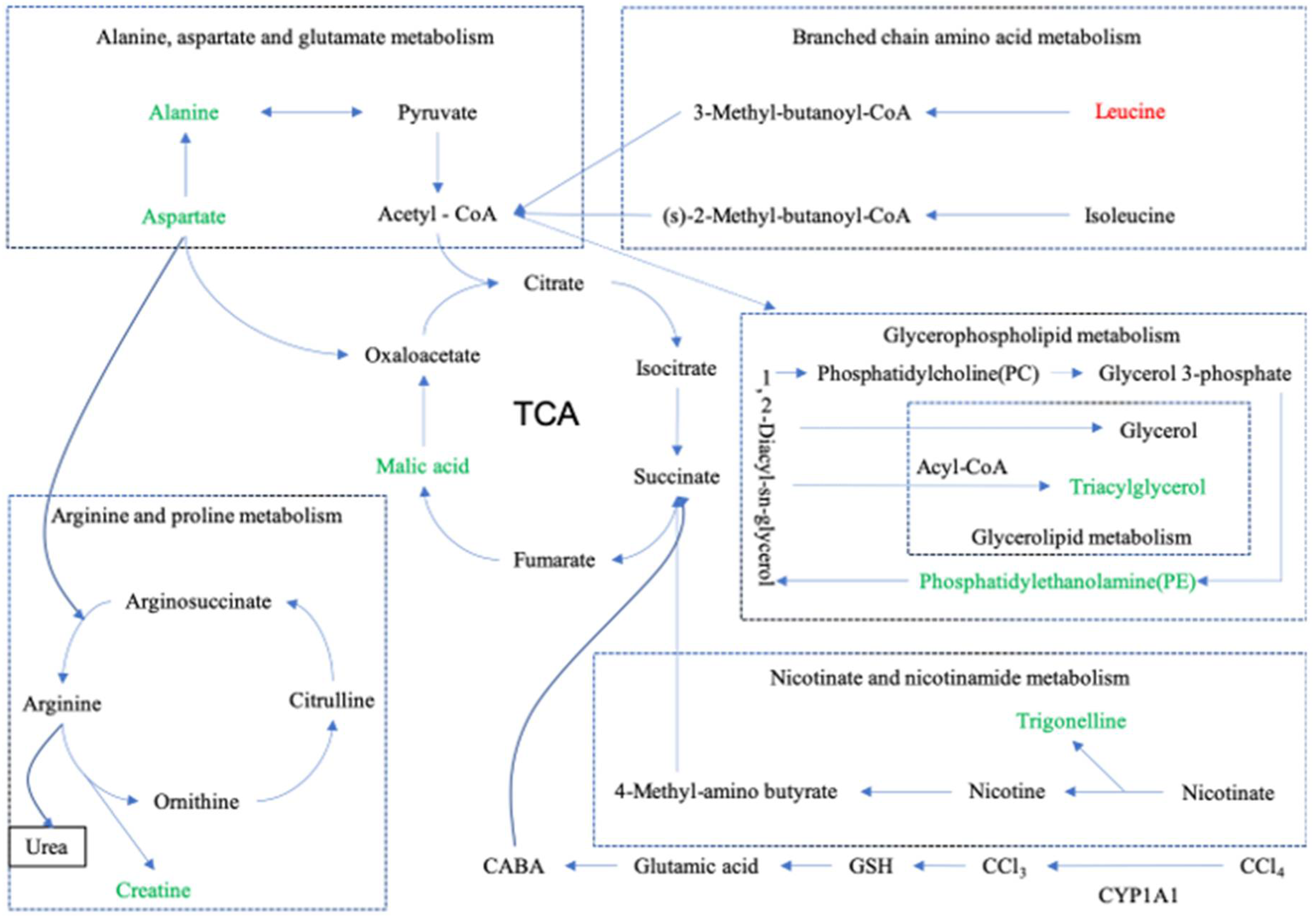
3.6.3. Metabolic Pathway Enrichment Analysis
3.7. Identification and Selection of Differential Genes
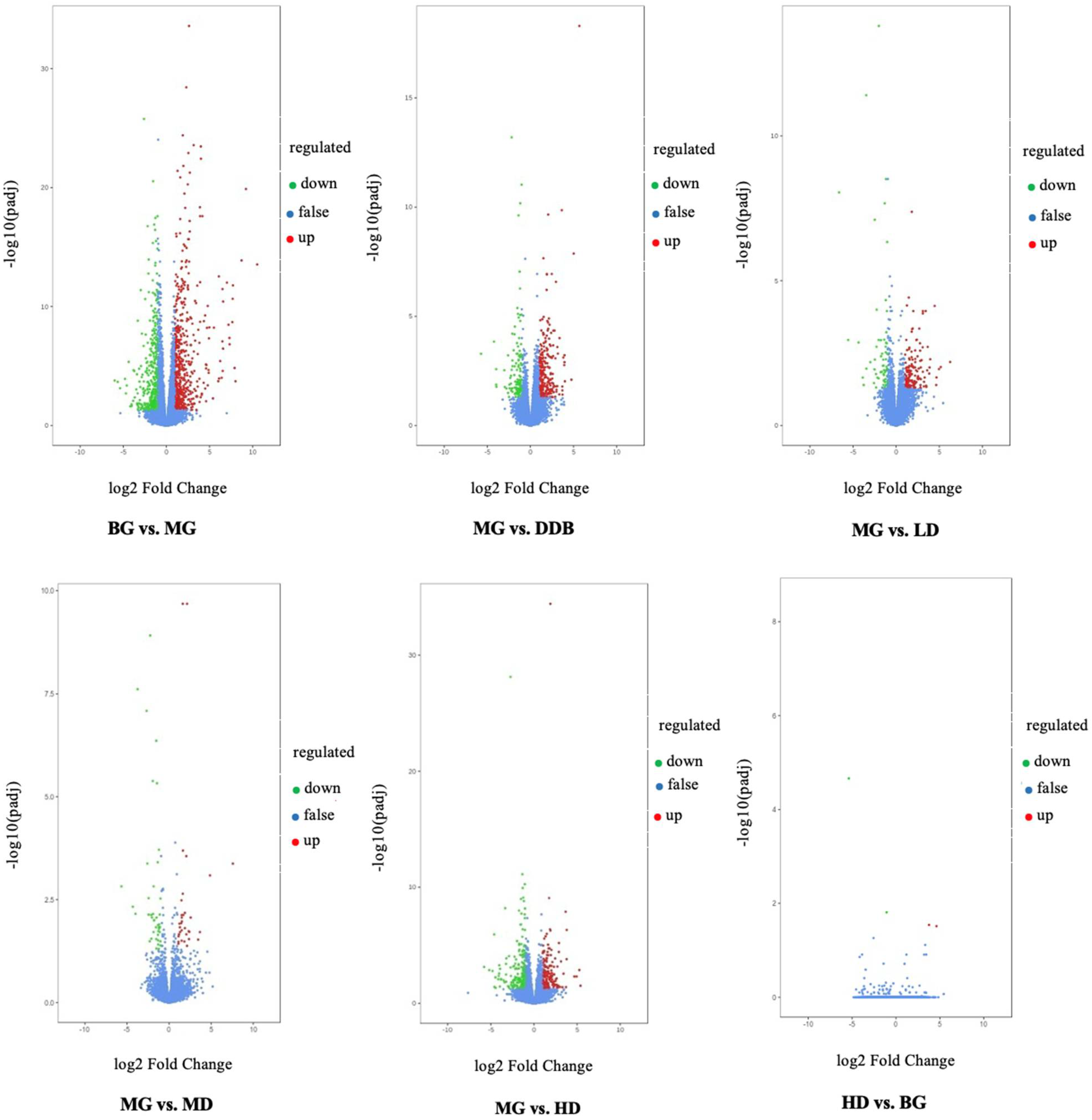
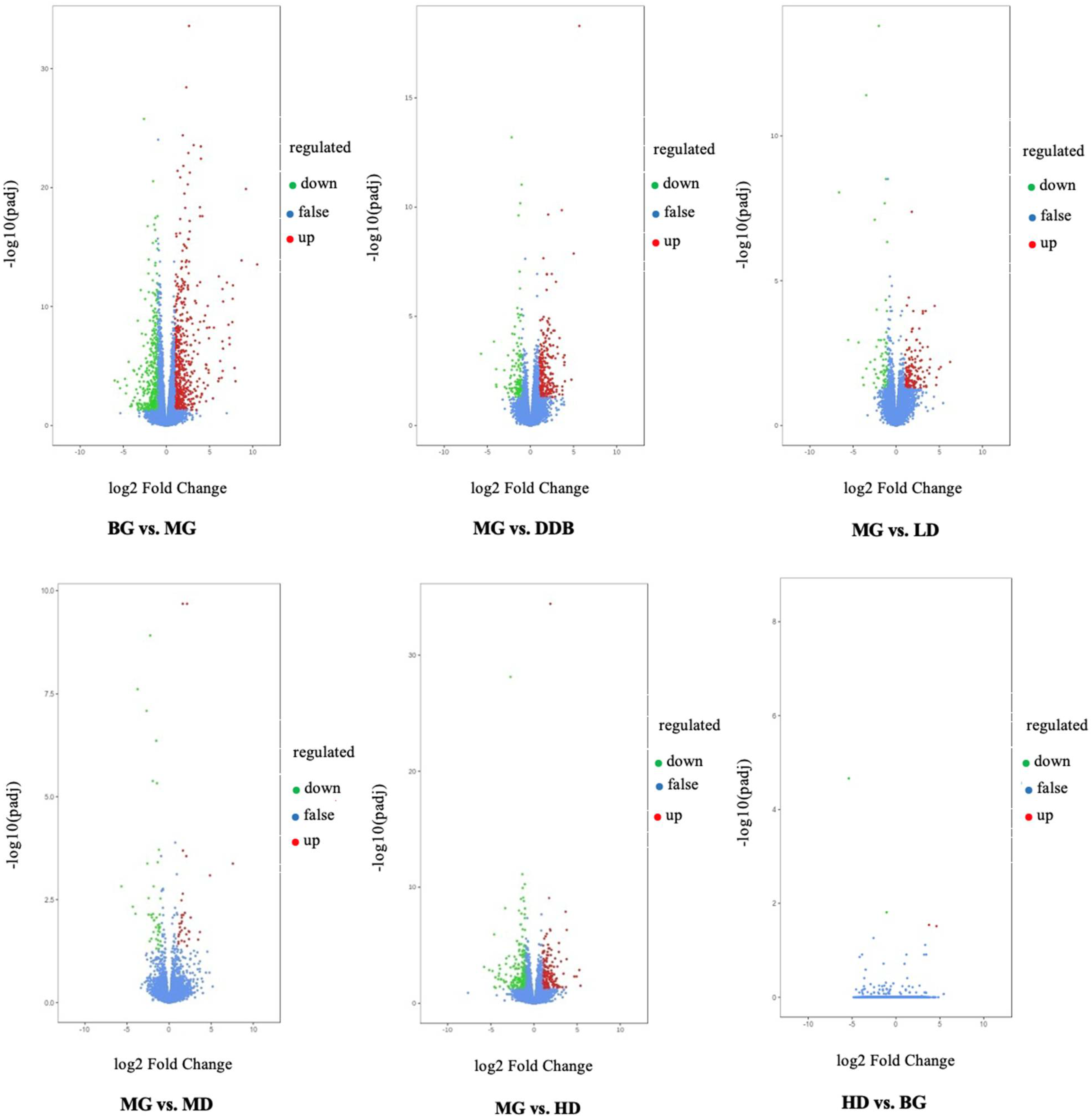
3.7.1. Differential Gene Volcano Map
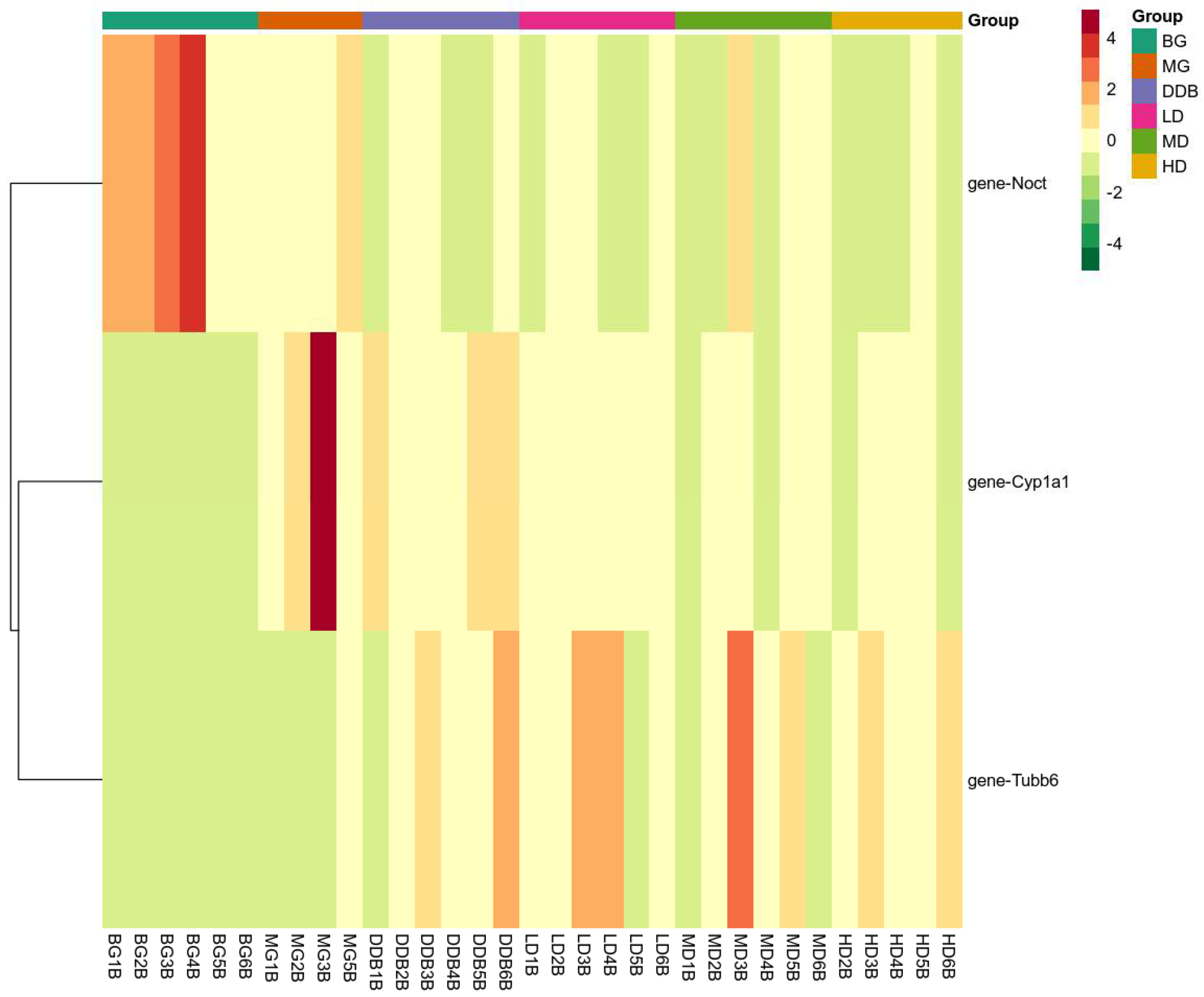
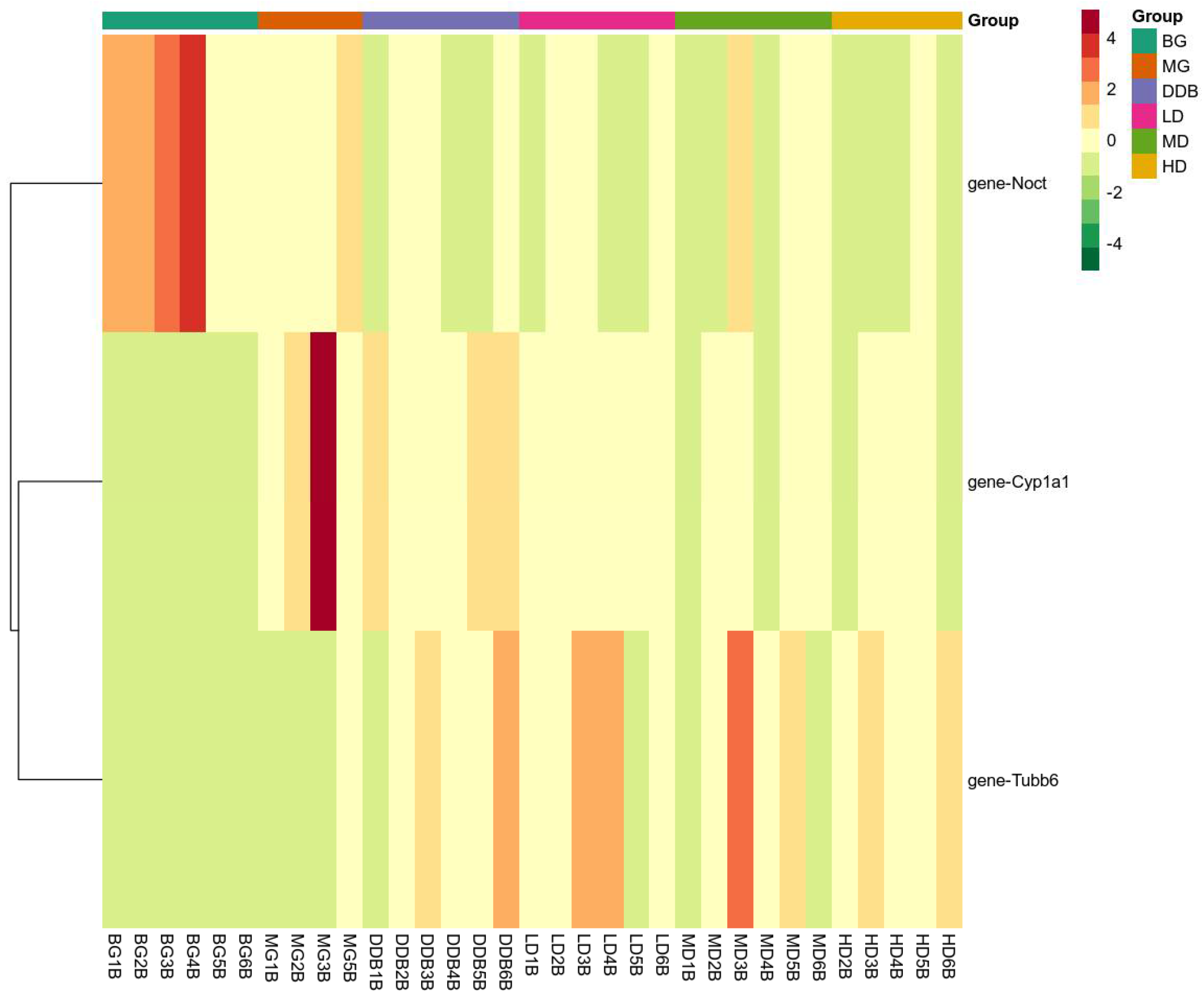
3.7.2. Expression of Key Genes
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Sample Availability
References
- Hao, W.; He, Z.; Zhu, H.; Liu, J.; Kwek, E.; Zhao, Y.; Ma, K.Y.; He, W.-S.; Chen, Z.-Y. Sea buckthorn seed oil reduces blood cholesterol and modulates gut microbiota. Food Funct. 2019, 10, 5669–5681. [Google Scholar] [CrossRef]
- Negi, B.; Kaur, R.; Dey, G. Protective effects of a novel sea buckthorn wine on oxidative stress and hypercholesterolemia. Food Funct. 2012, 4, 240–248. [Google Scholar] [CrossRef]
- Larmo, P.S.; Kangas, A.J.; Soininen, P.; Lehtonen, H.-M.; Suomela, J.-P.; Yang, B.; Viikari, J.; Ala-Korpela, M.; Kallio, H.P. Effects of sea buckthorn and bilberry on serum metabolites differ according to baseline metabolic profiles in overweight women: A randomized crossover trial. Am. J. Clin. Nutr. 2013, 98, 941–951. [Google Scholar] [CrossRef] [Green Version]
- Teleszko, M.; Wojdyło, A.; Rudzińska, M.; Oszmiański, J.; Golis, T. Analysis of Lipophilic and Hydrophilic Bioactive Compounds Content in Sea Buckthorn (Hippophaë rhamnoides L.) Berries. J. Agric. Food Chem. 2015, 63, 4120–4129. [Google Scholar] [CrossRef]
- Sajfrtova, M.; Ličková, I.; Wimmerová, M.; Sovová, H.; Wimmer, Z. β-Sitosterol: Supercritical Carbon Dioxide Extraction from Sea Buckthorn (Hippophae rhamnoides L.) Seeds. Int. J. Mol. Sci. 2010, 11, 1842–1850. [Google Scholar] [CrossRef]
- Derakhshesh, N.; Salamat, N.; Movahedinia, A.; Hashemitabar, M.; Bayati, V. Corrigendum to “exposure of liver cell culture from the orange-spotted grouper, Epinephelus coioides, to benzo[a]pyrene and light results in oxidative damage as measured by antioxidant enzymes”. Chemosphere 2019, 226, 534–544. [Google Scholar] [CrossRef]
- Paulpriya, K.; Tresina, P.S.; Mohan, V.R. Hepatoprotective effect of crotalaria longipes wight and arn, ethanol extract in CCL4 induced hepatotoxicity in wistar rats. Int. J. Toxicol. Pharmacol. Res. 2016, 8, 45–52. [Google Scholar]
- Baskar, A.A.; Al Numair, K.S.; Gabriel Paulraj, M.; Alsaif, M.A.; Muamar, M.A.; Ignacimuthu, S. β-sitosterol prevents lipid peroxidation and improves antioxidant status and histoarchitecture in rats with 1, 2-dimethylhydrazine-induced colon cancer. J. Med. Food 2012, 15, 335–343. [Google Scholar] [CrossRef]
- Song, L.; Zhou, H.; Yu, W.; Ding, X.; Yang, L.; Wu, J.; Song, C. Effects of phytosterol ester on the fatty acid profiles in rats with nonalcoholic fatty liver disease. J. Med. Food 2020, 23, 161–172. [Google Scholar] [CrossRef]
- Ran, B.; Guo, C.; Li, W.; Li, W.; Wang, Q.; Qian, J.; Li, H. Sea buckthorn (Hippophae rhamnoides L.) fermentation liquid protects against alcoholic liver disease linked to regulation of liver metabolome and the abundance of gut microbiota. J. Sci. Food Agric. 2021, 101, 2846–2854. [Google Scholar] [CrossRef]
- Mohamed, E.A.; Bordean, D.M.; Radulov, I.; Moruzi, R.F.; Hulea, C.I.; Orășan, S.A.; Dumitrescu, E.; Muselin, F.; Herman, H.; Brezovan, D.; et al. Sea Buckthorn and Grape Antioxidant Effects in Hyperlipidemic Rats: Relationship with the Atorvastatin Therapy. Evid.-Based Complement. Altern. Med. 2020, 2020, 1736803. [Google Scholar] [CrossRef]
- Liu, H.; Zhang, W.; Dong, S.; Song, L.; Zhao, S.; Wu, C.; Wang, X.; Liu, F.; Xie, J.; Wang, J.; et al. Protective effects of sea buckthorn polysaccharide extracts against LPS/d-GalN-induced acute liver failure in mice via suppressing TLR4-NF-κB signaling. J. Ethnopharmacol. 2015, 176, 69–78. [Google Scholar] [CrossRef]
- Liu, F.; Wang, T.; Li, X.; Jia, J.; Lin, Q.; Xue, Y. Involvement of NF-κB in the reversal of CYP3A down-regulation induced by sea buckthorn in BCG-induced rats. PLoS ONE 2020, 15, e0238810. [Google Scholar] [CrossRef]
- Wu, D.; Meng, Z. Effect of Sulfur Dioxide Inhalation on the Glutathione Redox System in Mice and Protective Role of Sea Buckthorn Seed Oil. Arch. Environ. Contam. Toxicol. 2003, 45, 423–428. [Google Scholar] [CrossRef]
- Zhao, S.; Sun, H.; Liu, Q.; Shen, Y.; Jiang, Y.; Li, Y.; Liu, T.; Liu, T.; Xu, H.; Shao, M. Protective effect of seabuckthorn berry juice against acrylamide-induced oxidative damage in rats. J. Food Sci. 2020, 85, 2245–2254. [Google Scholar] [CrossRef]
- Taysi, S.; Gumustekin, K.; Demircan, B.; Aktas, O.; Oztasan, N.; Akcay, F.; Suleyman, H.; Akar, S.; Dane, S.; Gul, M. Hippophae rhamnoides attenuates nicotine-induced oxidative stress in rat liver. Pharm. Biol. 2010, 48, 488–493. [Google Scholar] [CrossRef]
- Jiang, W.; Gao, M.; Sun, S.; Bi, A.; Xin, Y.; Han, X.; Wang, L.; Yin, Z.; Luo, L. Protective effect of l-theanine on carbon tetrachloride-induced acute liver injury in mice. Biochem. Biophys. Res. Commun. 2012, 422, 344–350. [Google Scholar] [CrossRef]
- Sheng, C.T.; Guo, Y.; Zhang, B.Y.; Yang, Y.J.; Ma, J.; Zhang, X.F.; Zhang, D.J. Protective effect of sea buckthorn sterol on acute liver injury induced by carbon tetrachloride in rats. China Food Addit. 2021, 32, 63–69. [Google Scholar]
- Xue, Y.T.; Zhang, X.F.; Zhang, Y.H.; Zhang, B.Y.; Zhang, D.J. Effect of sea buckthorn sterol on lowering blood lipid. China Food Addit. 2019, 30, 129–137. [Google Scholar]
- Liu, Y.; Wen, P.H.; Zhang, X.X.; Dai, Y.; He, Q. Breviscapine ameliorates CCl4 induced liver injury in mice through inhibiting inflammatory apoptotic response and ROS generation. Int. J. Mol. Med. 2018, 42, 755–768. [Google Scholar] [CrossRef] [Green Version]
- Xiong, X.; Ren, Y.; Cui, Y.; Li, R.; Wang, C.; Zhang, Y. Obeticholic acid protects mice against lipopolysaccharide-induced liver injury and inflammation. Biomed. Pharmacother. 2017, 96, 1292–1298. [Google Scholar] [CrossRef]
- Xia, H.; He, Q.; Wang, H.; Wang, Y.; Yang, Y.; Li, Y.; Zhang, J.; Chen, Z.; Yang, J. Treatment with either COX-2 inhibitor or 5-LOX inhibitor causes no compensation between COX-2 pathway and 5-LOX pathway in chronic aluminum overload-induced liver injury in rats. Fundam. Clin. Pharmacol. 2019, 33, 535–543. [Google Scholar] [CrossRef]
- Lin, S.; Wang, T.-Y.; Xu, H.-R.; Zhang, X.-N.; Wang, Q.; Liu, R.; Li, Q.; Bi, K.-S. A systemic combined nontargeted and targeted LC-MS based metabolomic strategy of plasma and liver on pathology exploration of alpha-naphthylisothiocyanate induced cholestatic liver injury in mice. J. Pharm. Biomed. Anal. 2019, 171, 180–192. [Google Scholar] [CrossRef]
- Xu, L.; Yu, Y.; Sang, R.; Li, J.; Ge, B.; Zhang, X. Protective Effects of Taraxasterol against Ethanol-Induced Liver Injury by Regulating CYP2E1/Nrf2/HO-1 and NF-κB Signaling Pathways in Mice. Oxid. Med. Cell. Longev. 2018, 2018, 8284107. [Google Scholar] [CrossRef] [Green Version]
- Zhang, C.; Wang, N.; Xu, Y.; Tan, H.-Y.; Li, S.; Feng, Y. Molecular Mechanisms Involved in Oxidative Stress-Associated Liver Injury Induced by Chinese Herbal Medicine: An Experimental Evidence-Based Literature Review and Network Pharmacology Study. Int. J. Mol. Sci. 2018, 19, 2745. [Google Scholar] [CrossRef] [Green Version]
- Aydemir, D.; Ulusu, N.N. Comment on the: Molecular mechanism of CAT and SOD activity change under MPA-CdTe quantum dots induced oxidative stress in the mouse primary hepatocytes. Spectrochim. Acta A Mol. Biomol. Spectrosc. 2019, 220, 117104. [Google Scholar]
- Jia, R.; Du, J.; Cao, L.; Li, Y.; Johnson, O.; Gu, Z.; Jeney, G.; Xu, P.; Yin, G. Antioxidative, inflammatory and immune responses in hydrogen peroxide-induced liver injury of tilapia (GIFT, Oreochromis niloticus). Fish Shellfish Immunol. 2019, 84, 894–905. [Google Scholar] [CrossRef]
- Zhang, X.; Wu, J.-Z.; Lin, Z.-X.; Yuan, Q.-J.; Li, Y.-C.; Liang, J.-L.; Zhan, J.Y.-X.; Xie, Y.-L.; Su, Z.-R.; Liu, Y.-H. Ameliorative effect of supercritical fluid extract of Chrysanthemum indicum Linnén against D-galactose induced brain and liver injury in senescent mice via suppression of oxidative stress, inflammation and apoptosis. J. Ethnopharmacol. 2019, 234, 44–56. [Google Scholar] [CrossRef]
- Zhang, Y.; Li, M.M.; Hua, T.M.; Sun, Q.Y. The protective effect of tea polyphenols on chronic alcoholic liver injury in rats. Chin. J. Appl. Physiol. 2018, 34, 481–484. (In Chinese) [Google Scholar]
- Kapelle, I.B.D.; Manalu, W.; Mainassy, M.C. Influence of Process Methods on the Hepatoprotective Effect of Curcumin Analogs Synthesized from Culilawan Oil in Mice (Mus musculus L.) with CCl4 Induced Liver Damage. Ohio J. Sci. 2019, 119, 28–37. [Google Scholar] [CrossRef]
- Liu, W.; Fan, T.; Li, M.; Zhang, G.; Guo, W.; Yang, X.; Jiang, C.; Li, X.; Xu, X.; Tang, A.; et al. Andrographolide potentiates PD-1 blockade immunotherapy by inhibiting COX-2 mediated PGE2 release. Int. Immunopharmacol. 2020, 81, 11–23. [Google Scholar] [CrossRef] [PubMed]
- Yao, L.; Chen, W.; Han, C.; Wu, T. Microsomal prostaglandin E synthase-1 protects against Fas-induced liver injury. Am. J. Physiol. Gastrointest. Liver Physiol. 2016, 310, 223–230. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, K.; Kong, J.; Liu, B.; Meng, X. Regulatory T cells suppress the expression of COX-2 in vulnerable plaque. Heart Vessel. 2020, 35, 456–460. [Google Scholar] [CrossRef] [PubMed]
- Elhosseiny, L.S.; Alqurashy, N.N.; Sheweita, S. A Oxidative stress alleviation by sage essential oil in co-amoxiclav induced hepatotoxicity in rats. Int. J. Biomed. Sci. 2016, 12, 71–78. [Google Scholar]
- Hu, D.-D.; Zhao, Q.; Cheng, Y.; Xiao, X.-R.; Huang, J.-F.; Qu, Y.; Li, X.; Tang, Y.-M.; Bao, W.-M.; Yang, J.-H.; et al. The Protective Roles of PPARα Activation in Triptolide-Induced Liver Injury. Toxicol. Sci. 2019, 171, 1–12. [Google Scholar] [CrossRef]
- Kuang, Y.; Han, X.; Xu, M.; Wang, Y.; Zhao, Y.; Yang, Q. Oxaloacetate Ameliorates Chemical Liver Injury via Oxidative Stress Reduction and Enhancement of Bioenergetic Fluxes. Int. J. Mol. Sci. 2018, 19, 1626. [Google Scholar] [CrossRef] [Green Version]
- Akram, M. Citric acid cycle and role of its intermediates in metabolism. Cell Biochem. Biophys. 2014, 68, 475–478. [Google Scholar] [CrossRef]
- Choiniere, J.; Lin, M.; Wang, L.; Wu, J. Deficiency of pyruvate dehydrogenase kinase 4 sensitizes mouse liver to diethylnitrosamine and arsenic toxicity through inducing apoptosis. Liver Res. 2018, 2, 100–107. [Google Scholar] [CrossRef]
- Liu, F.; Sun, Z.; Hu, P.; Tian, Q.; Xu, Z.; Li, Z.; Tian, X.; Chen, M.; Huang, C. Determining the protective effects of Yin-Chen-Hao Tang against acute liver injury induced by carbon tetrachloride using 16S rRNA gene sequencing and LC/MS-based metabolomics. J. Pharm. Biomed. Anal. 2019, 174, 567–577. [Google Scholar] [CrossRef]
- Ranjbar, K.; Nazem, F.; Sabrinezhad, R.; Nazari, A. Aerobic training and L-arginine supplement attenuates myocardial infarction-induced kidney and liver injury in rats via reduced oxidative stress. Indian Heart J. 2018, 70, 538–543. [Google Scholar] [CrossRef]
- Gaudet, A.D.; Fonken, L.K.; Ayala, M.T.; Dangelo, H.M.; Smith, E.J.; Bateman, E.M.; Schleicher, W.E.; Maier, S.F.; Watkins, L.R. Spinal Cord Injury in Rats Dysregulates Diurnal Rhythms of Fecal Output and Liver Metabolic Indicators. J. Neurotrauma 2019, 36, 1923–1934. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.-W.; Tang, C.-L.; Zheng, H.; Wu, J.-X.; Wu, F.; Mo, Y.-Y.; Liu, X.; Zhu, H.-J.; Yin, C.-L.; Cheng, B.; et al. Investigation of the hepatoprotective effect of Corydalis saxicola Bunting on carbon tetrachloride-induced liver fibrosis in rats by 1H-NMR-based metabonomics and network pharmacology approaches. J. Pharm. Biomed. Anal. 2018, 159, 252–261. [Google Scholar] [CrossRef]
- Rankin, K.C.; O’Brien, L.C.; Segal, L.; Khan, M.R.; Gorgey, A.S. Liver Adiposity and Metabolic Profile in Individuals with Chronic Spinal Cord Injury. BioMed Res. Int. 2017, 2017, 1364818. [Google Scholar] [CrossRef] [PubMed]
- Dong, S.; Chen, Q.-L.; Song, Y.-N.; Sun, Y.; Wei, B.; Li, X.-Y.; Hu, Y.-Y.; Liu, P.; Su, S.-B. Mechanisms of CCl4-induced liver fibrosis with combined transcriptomic and proteomic analysis. J. Toxicol. Sci. 2016, 41, 561–572. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Musso, G.; Cassader, M.; Paschetta, E.; Gambino, R. Bioactive Lipid Species and Metabolic Pathways in Progression and Resolution of Nonalcoholic Steatohepatitis. Gastroenterology 2018, 155, 282–302.e8. [Google Scholar] [CrossRef]
- Shi, C.; Han, X.; Mao, X.; Fan, C.; Jin, M. Metabolic profiling of liver tissues in mice after instillation of fine particulate matter. Sci. Total Environ. 2019, 696, 133974. [Google Scholar] [CrossRef]
- Singaravelu, R.; Quan, C.; Powdrill, M.H.; Shaw, T.A.; Srinivasan, P.; Lyn, R.K.; Alonzi, R.C.; Jones, D.M.; Filip, R.; Russell, R.S.; et al. MicroRNA-7 mediates cross-talk between metabolic signaling pathways in the liver. Sci. Rep. 2018, 8, 361. [Google Scholar] [CrossRef]
- Fonseca, L.D.; Eler, J.; Pereira, M.A.; Rosa, A.; Alexandre, P.A.; Moncau, C.T.; Salvato, F.; Rosa-Fernandes, L.; Palmisano, G.; Ferraz, J.B.S.; et al. Liver proteomics unravel the metabolic pathways related to Feed Efficiency in beef cattle. Sci. Rep. 2019, 9, 5364. [Google Scholar] [CrossRef] [Green Version]
- Lang, D.; Radtke, M.; Bairlein, M. Highly Variable Expression of CYP1A1 in Human Liver and Impact on Pharmacokinetics of Riociguat and Granisetron in Humans. Chem. Res. Toxicol. 2019, 32, 1115–1122. [Google Scholar] [CrossRef]
- Huang, B.; Bao, J.; Cao, Y.-R.; Gao, H.-F.; Jin, Y. Cytochrome P450 1A1 (CYP1A1) Catalyzes Lipid Peroxidation of Oleic Acid-Induced HepG2 Cells. Biochemistry 2018, 83, 595–602. [Google Scholar] [CrossRef]
- Li, Y.; Li, Y.; Zheng, G.; Zhu, L.; Wang, J.; Mu, S.; Ren, Q.; Feng, F. Cytochrome P450 1A1 and 1B1 promoter CpG island methylation regulates rat liver injury induced by isoniazid. Mol. Med. Rep. 2017, 17, 753–762. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xia, H.; Zhu, X.; Zhang, X.; Jiang, H.; Li, B.; Wang, Z.; Li, D.; Jin, Y. Alpha-naphthoflavone attenuates non-alcoholic fatty liver disease in oleic acid-treated HepG2 hepatocytes and in high fat diet-fed mice. Biomed. Pharmacother. 2019, 118, 109287. [Google Scholar] [CrossRef] [PubMed]
- He, L.; Gao, L.; Shi, Z.; Li, Y.; Zhu, L.; Li, S.; Zhang, P.; Zheng, G.; Ren, Q.; Li, Y.; et al. Involvement of Cytochrome P450 1A1 and Glutathione S-Transferase P1 Polymorphisms and Promoter Hypermethylation in the Progression of Anti-Tuberculosis Drug-Induced Liver Injury: A Case–Control Study. PLoS ONE 2015, 10, e0119481. [Google Scholar] [CrossRef]
- Xiong, X.; Xu, D.; Yang, Z.; Huang, H.; Cui, X. A single amino-acid substitution at lysine 40 of an Arabidopsis thalianaα-tubulin causes extensive cell proliferation and expansion defects. J. Integr. Plant Biol. 2013, 55, 209–220. [Google Scholar] [CrossRef] [PubMed]
- Salinas, R.E.; Ogohara, C.; Thomas, M.I.; Shukla, K.P.; Miller, S.I.; Ko, D.C. A cellular genome-wide association study reveals human variation in microtubule stability and a role in inflammatory cell death. Mol. Biol. Cell 2014, 25, 76–86. [Google Scholar] [CrossRef] [PubMed]
- Nami, B.; Wang, Z. Genetics and Expression Profile of the Tubulin Gene Superfamily in Breast Cancer Subtypes and Its Relation to Taxane Resistance. Cancers 2018, 10, 274. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Le, P.T.; Bornstein, S.A.; Motyl, K.J.; Tian, L.; Stubblefield, J.J.; Hong, H.; Takahashi, J.S.; Green, C.B.; Rosen, C.J.; Guntur, A.R. A novel mouse model overexpressing Nocturnin results in decreased fat mass in male mice. J. Cell. Physiol. 2019, 234, 20228–20239. [Google Scholar] [CrossRef]
- Abshire, E.; Chasseur, J.; Bohn, J.A.; Del Rizzo, P.A.; Freddolino, P.L.; Goldstrohm, A.C.; Trievel, R.C. The structure of human Nocturnin reveals a conserved ribonuclease domain that represses target transcript translation and abundance in cells. Nucleic Acids Res. 2018, 46, 6257–6270. [Google Scholar] [CrossRef]
- Xie, Q.; Guo, F.-F.; Zhou, W. Protective effects of cassia seed ethanol extract against carbon tetrachloride-induced liver injury in mice. Acta Biochim. Pol. 2012, 59, 265–270. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Group | Results |
|---|---|
| BG | (−) |
| MG | Feathery degeneration of hepatocyte (++) Fatty degeneration of hepatocytes (++) Hepatocyte necrosis (++) Inflammatory cell infiltration (+) |
| DDB | A small amount of feathery degeneration of hepatocytes (+) A small amount of fatty degeneration of hepatocytes (+) Hepatocyte necrosis (+) |
| LD | Feathery degeneration of hepatocytes (++) Fatty degeneration of hepatocytes (+) Hepatocyte necrosis (+) Inflammatory cell infiltration (+) |
| MD | Feathery degeneration of hepatocytes (++) Fatty degeneration of hepatocytes (+) Hepatocyte necrosis (++) Inflammatory cell infiltration (+) |
| HD | A small amount of feathery degeneration of hepatocytes (+) A small amount of fatty degeneration of hepatocytes (+) Hepatocyte necrosis (+) |
| NO. | Metabolites | RT | MG/BG | LD/MG | MD/MG | HD/MG | DDB/MG | Pathway |
|---|---|---|---|---|---|---|---|---|
| 1 | L-Malic Acid | 0.88 | ↑ * | ↓/- | ↓ * | ↓ * | ↓/- | Citrate cycle (TCA cycle) |
| 2 | 7Z, 10Z, 13Z, 16Z, 19Z-docosapentaenoic acid | 11.3 | ↑ * | ↓ * | ↓ * | ↓ * | ↓/- | Biosynthesis of unsaturated fatty acids |
| 3 | creatine | 0.78 | ↑ * | ↓/- | ↓ * | ↓ * | ↓ * | Arginine-proline metabolism |
| 4 | n-acetyl-l-alanine | 1.3 | ↑ * | ↓/- | ↓ * | ↓ * | ↓ * | - |
| 5 | N-Acetylaspartate | 0.69 | ↑ * | ↓/- | ↓ * | ↓ * | ↓ * | Alanine, aspartate and glutamate metabolism |
| 6 | Trigonelline | 0.77 | ↑ * | ↓/- | ↓ * | ↓ * | ↓/- | Nicotinate and nicotinamide metabolism |
| 7 | 4-guanidinobutyric acid | 0.85 | ↑ * | ↓ * | ↓ * | ↓ * | ↓ * | Arginine-proline metabolism |
| 8 | N-Amidino-L-Aspartate | 0.96 | ↑ * | ↓ * | ↓ * | ↓ * | ↓ * | - |
| 9 | n-glycyl-l-leucine | 1.8 | ↓ * | ↑/- | ↑ * | ↑ * | ↑/- | - |
| 10 | FFA(6:0) | 0.73 | ↓ * | ↑ * | ↑ * | ↑ * | ↑ * | Fat digestion and absorption |
| 11 | CE(16:1) | 13.12 | ↑ * | ↓ * | ↓ * | ↓ * | ↓ * | - |
| 12 | CE(18:2) | 13.21 | ↑ * | ↓ * | ↓ * | ↓ * | ↓ * | - |
| 13 | PE(16:1/16:0) | 6.25 | ↑ * | ↓ * | ↓ * | ↓ * | ↓ * | Glycerophospholipid metabolism |
| 14 | DG(16:0/20:2/0:0) | 9.18 | ↑ * | ↓/- | ↓ * | ↓ * | ↓/- | Glycerolipid metabolism |
| 15 | TG(14:0/18:0/18:2) | 11.85 | ↑ * | ↓/- | ↓ * | ↓ * | ↓ * | Glycerolipid metabolism |
| 16 | TG(14:0/18:0/20:4) | 11.81 | ↑ * | ↓/- | ↓ * | ↓ * | ↓ * | Glycerolipid metabolism |
| 17 | TG(16:0/16:1/22:5) | 11.41 | ↑ * | ↓/- | ↓ * | ↓ * | ↓ * | Glycerolipid metabolism |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sheng, C.; Guo, Y.; Ma, J.; Hong, E.-K.; Zhang, B.; Yang, Y.; Zhang, X.; Zhang, D. Metabolomic Profiling Reveals Protective Effects and Mechanisms of Sea Buckthorn Sterol against Carbon Tetrachloride-Induced Acute Liver Injury in Rats. Molecules 2022, 27, 2224. https://doi.org/10.3390/molecules27072224
Sheng C, Guo Y, Ma J, Hong E-K, Zhang B, Yang Y, Zhang X, Zhang D. Metabolomic Profiling Reveals Protective Effects and Mechanisms of Sea Buckthorn Sterol against Carbon Tetrachloride-Induced Acute Liver Injury in Rats. Molecules. 2022; 27(7):2224. https://doi.org/10.3390/molecules27072224
Chicago/Turabian StyleSheng, Changting, Yang Guo, Jing Ma, Eun-Kyung Hong, Benyin Zhang, Yongjing Yang, Xiaofeng Zhang, and Dejun Zhang. 2022. "Metabolomic Profiling Reveals Protective Effects and Mechanisms of Sea Buckthorn Sterol against Carbon Tetrachloride-Induced Acute Liver Injury in Rats" Molecules 27, no. 7: 2224. https://doi.org/10.3390/molecules27072224
APA StyleSheng, C., Guo, Y., Ma, J., Hong, E.-K., Zhang, B., Yang, Y., Zhang, X., & Zhang, D. (2022). Metabolomic Profiling Reveals Protective Effects and Mechanisms of Sea Buckthorn Sterol against Carbon Tetrachloride-Induced Acute Liver Injury in Rats. Molecules, 27(7), 2224. https://doi.org/10.3390/molecules27072224