
Improved Wound Healing by Naringin Associated with MMP and the VEGF Pathway

 and
and {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
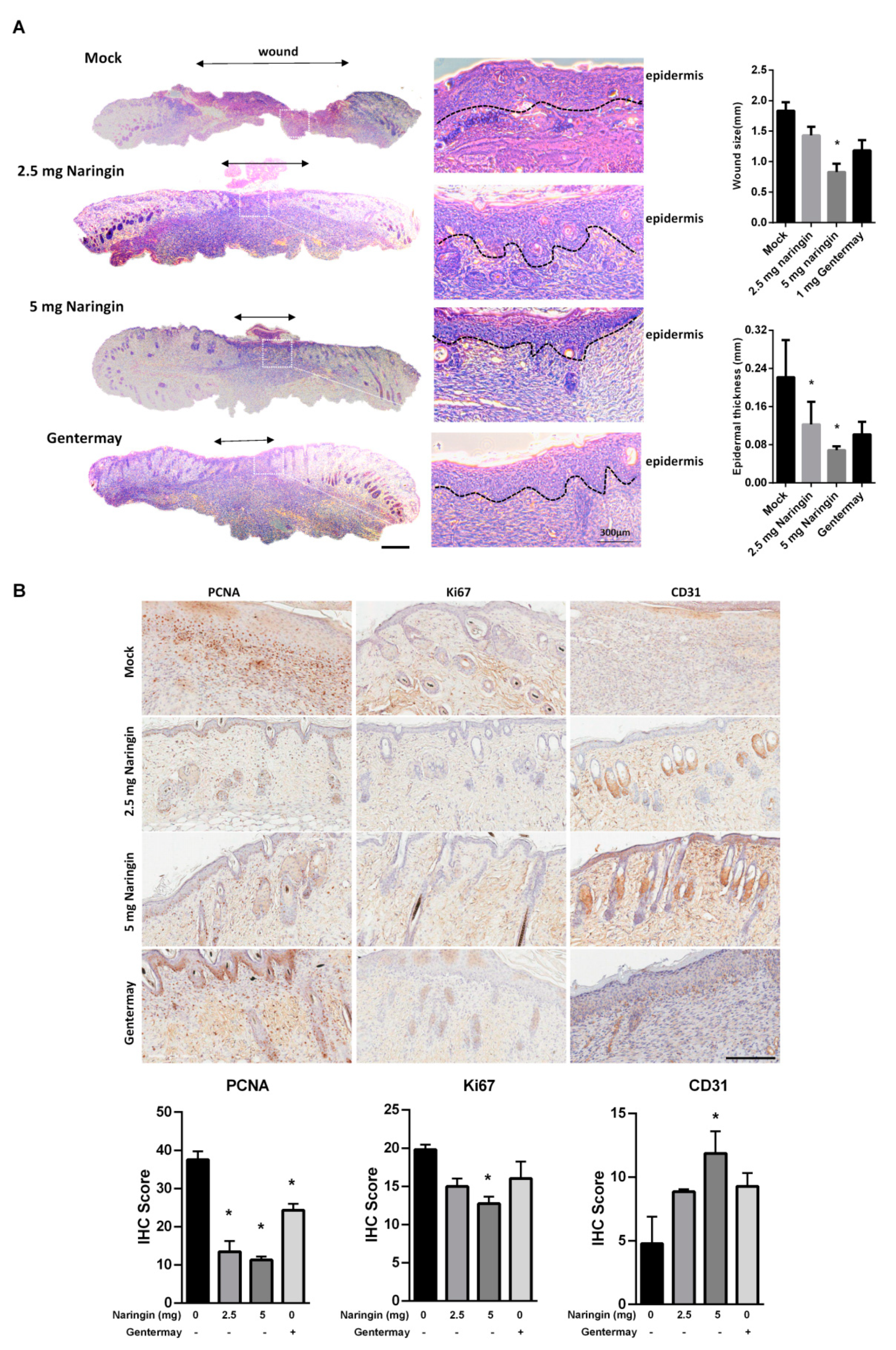
2.1. Naringin Cream Improves Wound Healing in a Mouse Model
2.2. Naringin Cream Treatment In Vivo
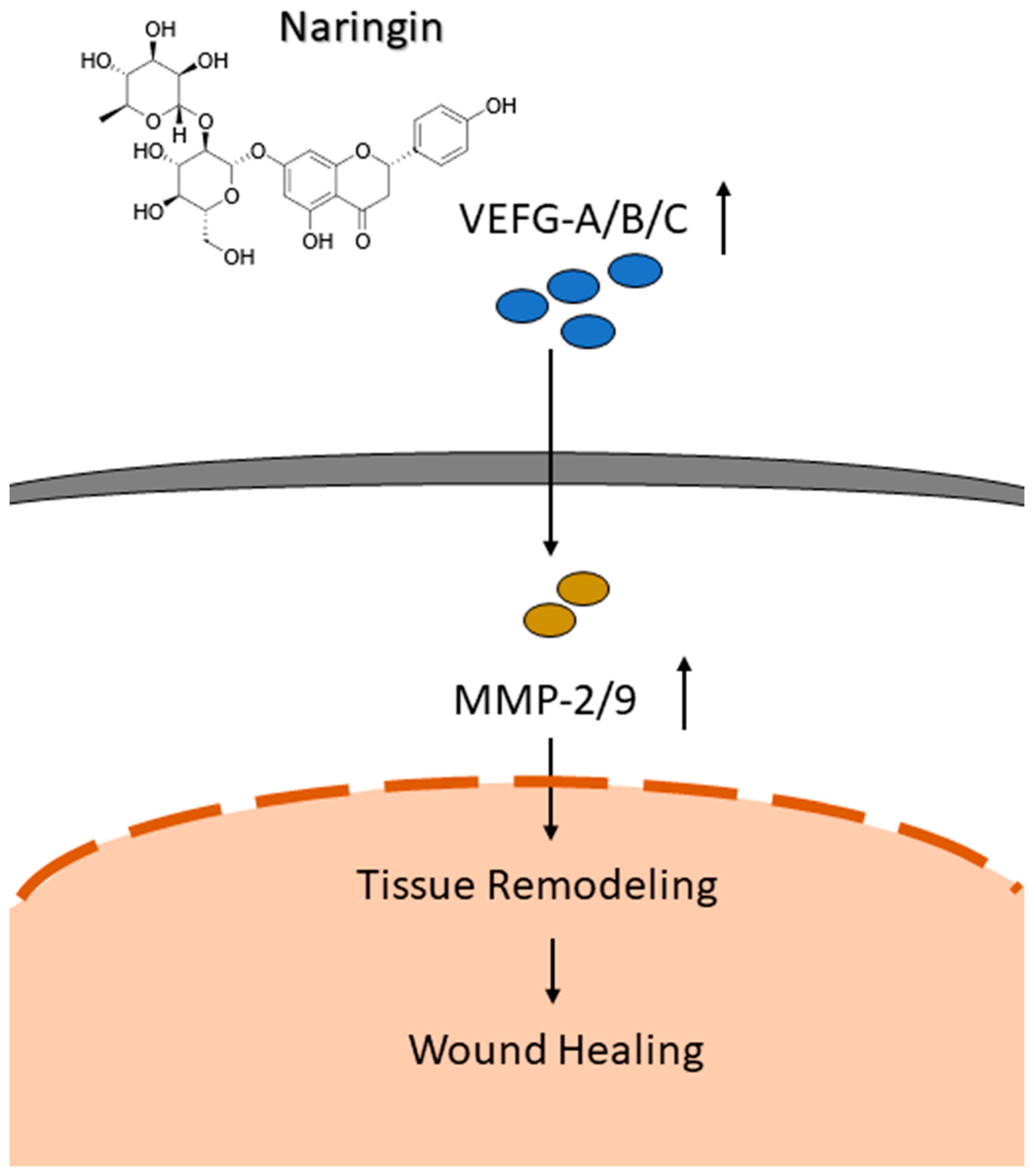
2.3. Upregulated Expressions of VEGFs and VEGFRs via Naringin Cream Treatment
2.4. Naringin Activates HaCaT Keratinocytes Migration
3. Discussion
4. Materials and Methods
4.1. Animals and Cell Culture
4.2. Preparation of Naringin Ointment
4.3. In Vivo Wound Model
4.4. Histopathological Evaluation
4.5. Western Blot
4.6. Wound Healing Assay
4.7. Migration Assay
4.8. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Eming, S.A.; Krieg, T.; Davidson, J.M. Inflammation in wound repair: Molecular and cellular mechanisms. J. Investig. Dermatol. 2007, 127, 514–525. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Worley, C.A. The wound healing process: Part III—The finale. Dermatol. Nurs. 2004, 16, 274–295. [Google Scholar] [PubMed]
- Worley, C.A. The wound healing process symphony: Part II. Dermatol. Nurs. 2004, 16, 179–180. [Google Scholar]
- Worley, C.A. The wound healing process symphony: Part I. Dermatol. Nurs. 2004, 16, 67–72. [Google Scholar]
- Sen, C.K. Human Wounds and Its Burden: An Updated Compendium of Estimates. Adv. Wound Care 2019, 8, 39–48. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Frykberg, R.G.; Banks, J. Challenges in the Treatment of Chronic Wounds. Adv. Wound Care 2015, 4, 560–582. [Google Scholar] [CrossRef] [Green Version]
- Ramanujam, C.L.; Han, D.; Fowler, S.; Kilpadi, K.; Zgonis, T. Impact of diabetes and comorbidities on split-thickness skin grafts for foot wounds. J. Am. Podiatr. Med. Assoc. 2013, 103, 223–232. [Google Scholar] [CrossRef]
- Usui, M.L.; Mansbridge, J.N.; Carter, W.G.; Fujita, M.; Olerud, J.E. Keratinocyte migration, proliferation, and differentiation in chronic ulcers from patients with diabetes and normal wounds. J. Histochem. Cytochem. Off. J. Histochem. Soc. 2008, 56, 687–696. [Google Scholar] [CrossRef] [Green Version]
- Kumar, M.; Dhatwalia, S.K.; Dhawan, D.K. Role of angiogenic factors of herbal origin in regulation of molecular pathways that control tumor angiogenesis. Tumour Biol. J. Int. Soc. Oncodev. Biol. Med. 2016, 37, 14341–14354. [Google Scholar] [CrossRef]
- Maeshima, Y.; Makino, H. The role of pro-angiogenic factors and angiogenesis inhibitors in diabetic nephropathy. Nihon Rinsho Jpn. J. Clin. Med. 2010, 68 (Suppl. S9), 396–400. [Google Scholar]
- Brown, L.F.; Yeo, K.T.; Berse, B.; Yeo, T.K.; Senger, D.R.; Dvorak, H.F.; van de Water, L. Expression of vascular permeability factor (vascular endothelial growth factor) by epidermal keratinocytes during wound healing. J. Exp. Med. 1992, 176, 1375–1379. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Karkkainen, M.J.; Haiko, P.; Sainio, K.; Partanen, J.; Taipale, J.; Petrova, T.V.; Jeltsch, M.; Jackson, D.G.; Talikka, M.; Rauvala, H.; et al. Vascular endothelial growth factor C is required for sprouting of the first lymphatic vessels from embryonic veins. Nat. Immunol. 2004, 5, 74–80. [Google Scholar] [CrossRef] [PubMed]
- Makinen, T.; Jussila, L.; Veikkola, T.; Karpanen, T.; Kettunen, M.I.; Pulkkanen, K.J.; Kauppinen, R.; Jackson, D.G.; Kubo, H.; Nishikawa, S.; et al. Inhibition of lymphangiogenesis with resulting lymphedema in transgenic mice expressing soluble VEGF receptor-3. Nat. Med. 2001, 7, 199–205. [Google Scholar] [CrossRef] [PubMed]
- Partanen, T.A.; Arola, J.; Saaristo, A.; Jussila, L.; Ora, A.; Miettinen, M.; Stacker, S.A.; Achen, M.G.; Alitalo, K. VEGF-C and VEGF-D expression in neuroendocrine cells and their receptor, VEGFR-3, in fenestrated blood vessels in human tissues. FASEB J. Off. Publ. Fed. Am. Soc. Exp. Biol. 2000, 14, 2087–2096. [Google Scholar] [CrossRef] [PubMed]
- Skobe, M.; Hamberg, L.M.; Hawighorst, T.; Schirner, M.; Wolf, G.L.; Alitalo, K.; Detmar, M. Concurrent induction of lymphangiogenesis, angiogenesis, and macrophage recruitment by vascular endothelial growth factor-C in melanoma. Am. J. Pathol. 2001, 159, 893–903. [Google Scholar] [CrossRef] [Green Version]
- Shibuya, M. Vascular Endothelial Growth Factor (VEGF) and Its Receptor (VEGFR) Signaling in Angiogenesis: A Crucial Target for Anti- and Pro-Angiogenic Therapies. Genes Cancer 2011, 2, 1097–1105. [Google Scholar] [CrossRef]
- Saaristo, A.; Tammela, T.; Farkkila, A.; Karkkainen, M.; Suominen, E.; Yla-Herttuala, S.; Alitalo, K. Vascular endothelial growth factor-C accelerates diabetic wound healing. Am. J. Pathol. 2006, 169, 1080–1087. [Google Scholar] [CrossRef] [Green Version]
- Akintunde, J.K.; Akintola, T.E.; Aliu, F.H.; Fajoye, M.O.; Adimchi, S.O. Naringin regulates erectile dysfunction by abolition of apoptosis and inflammation through NOS/cGMP/PKG signalling pathway on exposure to Bisphenol-A in hypertensive rat model. Reprod. Toxicol. 2020, 95, 123–136. [Google Scholar] [CrossRef]
- Elsawy, H.; Algefare, A.I.; Alfwuaires, M.; Khalil, M.; Elmenshawy, O.M.; Sedky, A.; Abdel-Moneim, A.M. Naringin alleviates methotrexate-induced liver injury in male albino rats and enhances its antitumor efficacy in HepG2 cells. Biosci. Rep. 2020, 40, BSR20193686. [Google Scholar] [CrossRef]
- Zhou, J.; Xia, L.; Zhang, Y. Naringin inhibits thyroid cancer cell proliferation and induces cell apoptosis through repressing PI3K/AKT pathway. Pathol. Res. Pract. 2019, 215, 152707. [Google Scholar] [CrossRef]
- Bai, J.; Li, S.; Wu, G.; Ma, L.; Li, C.; Liu, H.; Zhang, P. Naringin inhibits lipopolysaccharide-induced activation of microglia cells. Cell. Mol. Biol. 2019, 65, 38–42. [Google Scholar] [CrossRef] [PubMed]
- Nan, L.P.; Wang, F.; Ran, D.; Zhou, S.F.; Liu, Y.; Zhang, Z.; Huang, Z.N.; Wang, Z.Y.; Wang, J.C.; Feng, X.M.; et al. Naringin alleviates H2O2-induced apoptosis via the PI3K/Akt pathway in rat nucleus pulposus-derived mesenchymal stem cells. Connect. Tissue Res. 2020, 61, 554–567. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.; Gao, M.; Yang, B.; Zhang, H.; Wang, K.; Liu, Z.; Xiao, X.; Yang, M. Naringin attenuates MLC phosphorylation and NF-kappaB activation to protect sepsis-induced intestinal injury via RhoA/ROCK pathway. Biomed. Pharmacother. 2018, 103, 50–58. [Google Scholar] [CrossRef] [PubMed]
- Lv, Z.; Wu, W.; Ge, S.; Jia, R.; Lin, T.; Yuan, Y.; Kuang, H.; Yang, B.; Wu, L.; Wei, J.; et al. Naringin protects against perfluorooctane sulfonate-induced liver injury by modulating NRF2 and NF-kappaB in mice. Int. Immunopharmacol. 2018, 65, 140–147. [Google Scholar] [CrossRef] [PubMed]
- Heidary Moghaddam, R.; Samimi, Z.; Moradi, S.Z.; Little, P.J.; Xu, S.; Farzaei, M.H. Naringenin and naringin in cardiovascular disease prevention: A preclinical review. Eur. J. Pharmacol. 2020, 887, 173535. [Google Scholar] [CrossRef]
- Amini, N.; Sarkaki, A.; Dianat, M.; Mard, S.A.; Ahangarpour, A.; Badavi, M. The renoprotective effects of naringin and trimetazidine on renal ischemia/reperfusion injury in rats through inhibition of apoptosis and downregulation of micoRNA-10a. Biomed. Pharmacother. 2019, 112, 108568. [Google Scholar] [CrossRef]
- Gopinath, K.; Prakash, D.; Sudhandiran, G. Neuroprotective effect of naringin, a dietary flavonoid against 3-nitropropionic acid-induced neuronal apoptosis. Neurochem. Int. 2011, 59, 1066–1073. [Google Scholar] [CrossRef]
- Wallace, H.A.; Basehore, B.M.; Zito, P.M. Wound Healing Phases. In StatPearls; StatPearls Publishing: Treasure Island, FL, USA, 2021. [Google Scholar]
- Wernick, B.; Nahirniak, P.; Stawicki, S.P. Impaired Wound Healing. In StatPearls; StatPearls Publishing: Treasure Island, FL, USA, 2021. [Google Scholar]
- Roine, E.; Bjork, I.T.; Oyen, O. Targeting risk factors for impaired wound healing and wound complications after kidney transplantation. Transplant. Proc. 2010, 42, 2542–2546. [Google Scholar] [CrossRef]
- Menke, N.B.; Ward, K.R.; Witten, T.M.; Bonchev, D.G.; Diegelmann, R.F. Impaired wound healing. Clin. Dermatol. 2007, 25, 19–25. [Google Scholar] [CrossRef]
- Knight, R.J.; Villa, M.; Laskey, R.; Benavides, C.; Schoenberg, L.; Welsh, M.; Kerman, R.H.; Podder, H.; Van Buren, C.T.; Katz, S.M.; et al. Risk factors for impaired wound healing in sirolimus-treated renal transplant recipients. Clin. Transplant. 2007, 21, 460–465. [Google Scholar] [CrossRef]
- Song, N.; Zhao, Z.; Ma, X.; Sun, X.; Ma, J.; Li, F.; Sun, L.; Lv, J. Naringin promotes fracture healing through stimulation of angiogenesis by regulating the VEGF/VEGFR-2 signaling pathway in osteoporotic rats. Chem.-Biol. Interact. 2017, 261, 11–17. [Google Scholar] [CrossRef] [PubMed]
- Zhang, P.; Dai, K.R.; Yan, S.G.; Yan, W.Q.; Zhang, C.; Chen, D.Q.; Xu, B.; Xu, Z.W. Effects of naringin on the proliferation and osteogenic differentiation of human bone mesenchymal stem cell. Eur. J. Pharmacol. 2009, 607, 1–5. [Google Scholar] [CrossRef] [PubMed]
- Wu, G.J.; Chen, K.Y.; Yang, J.D.; Liu, S.H.; Chen, R.M. Naringin Improves Osteoblast Mineralization and Bone Healing and Strength through Regulating Estrogen Receptor Alpha-Dependent Alkaline Phosphatase Gene Expression. J. Agric. Food Chem. 2021, 69, 13020–13033. [Google Scholar] [CrossRef] [PubMed]
- Arpino, V.; Brock, M.; Gill, S.E. The role of TIMPs in regulation of extracellular matrix proteolysis. Matrix Biol. J. Int. Soc. Matrix Biol. 2015, 44–46, 247–254. [Google Scholar] [CrossRef] [PubMed]




Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yen, J.-H.; Chio, W.-T.; Chuang, C.-J.; Yang, H.-L.; Huang, S.-T. Improved Wound Healing by Naringin Associated with MMP and the VEGF Pathway. Molecules 2022, 27, 1695. https://doi.org/10.3390/molecules27051695
Yen J-H, Chio W-T, Chuang C-J, Yang H-L, Huang S-T. Improved Wound Healing by Naringin Associated with MMP and the VEGF Pathway. Molecules. 2022; 27(5):1695. https://doi.org/10.3390/molecules27051695
Chicago/Turabian StyleYen, Jia-Hau, Wan-Ting Chio, Chia-Ju Chuang, Hsin-Ling Yang, and Sheng-Teng Huang. 2022. "Improved Wound Healing by Naringin Associated with MMP and the VEGF Pathway" Molecules 27, no. 5: 1695. https://doi.org/10.3390/molecules27051695
APA StyleYen, J.-H., Chio, W.-T., Chuang, C.-J., Yang, H.-L., & Huang, S.-T. (2022). Improved Wound Healing by Naringin Associated with MMP and the VEGF Pathway. Molecules, 27(5), 1695. https://doi.org/10.3390/molecules27051695