
Antibacterial Activity of Chinese Red Propolis against Staphylococcus aureus and MRSA

 ,
, 
 and
and
Abstract
1. Introduction
2. Results
2.1. Results of the Quantitative Analysis of Polyphenols in RPE
2.2. RPE Has Anti-Bacterial Effect on S. aureus and MRSA
2.3. Time–Growth Curve Analysis
2.4. Inhibitory Effect of RPE on Intracellular Protein and DNA Leakage Analysis
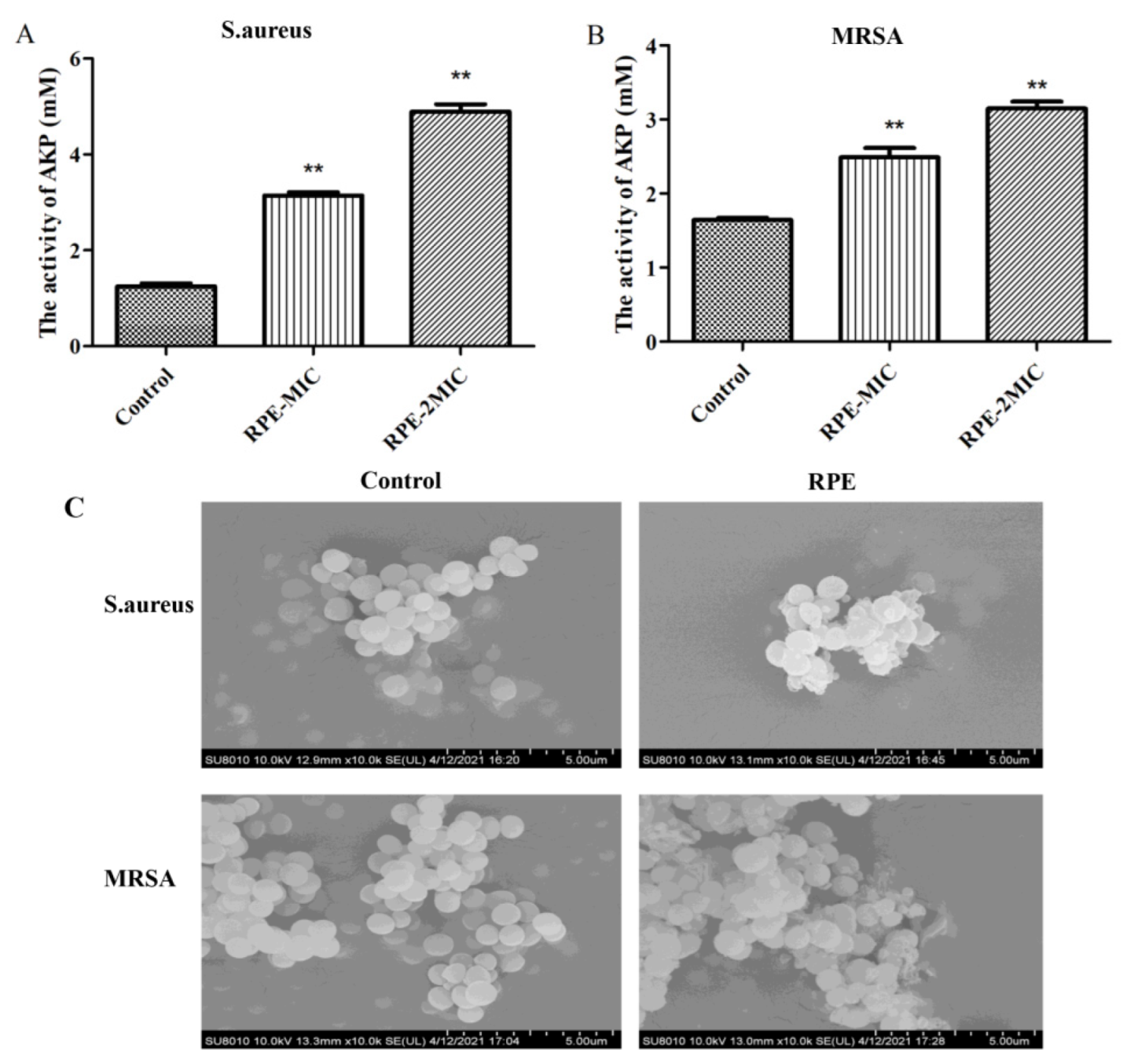
2.5. Cell Wall Integrity Analysis
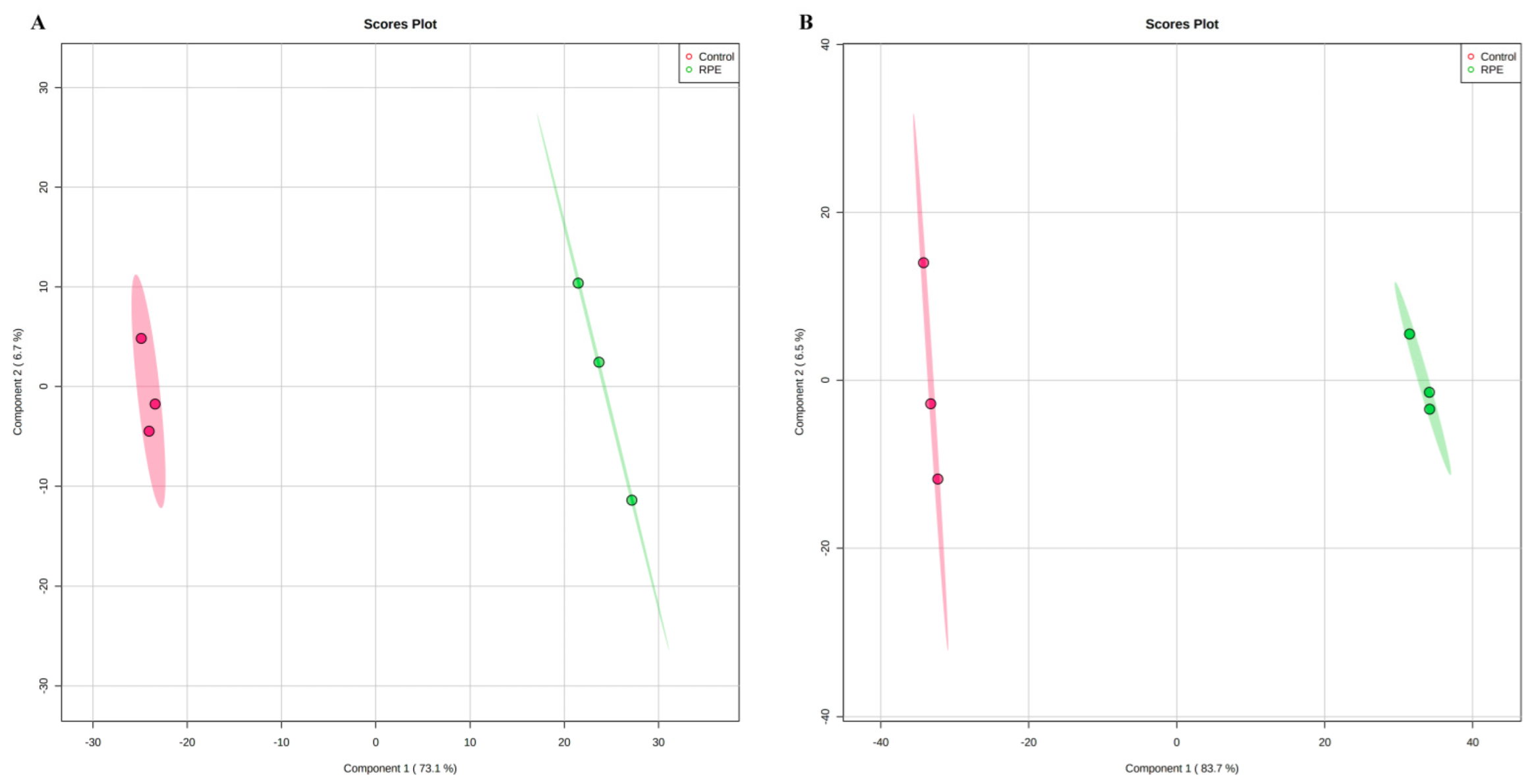
2.6. Metabolomic Analysis of the Effect of RPE Treated on S. aureus and MRSA
2.7. Effect of RPE on the Related Gene Expression of MRSA
3. Discussion
4. Materials and Methods
4.1. Extraction of CRP
4.2. Calibration and Quantification of the Main Polyphenols in RPE
4.3. Bacterial Strains and Growth Conditions
4.4. Determination of Antibacterial Activity of RPE
4.5. Determination of Minimal Inhibitory Concentration and Minimum Bactericidal Concentration (MBC)
4.6. Time–Growth Curve Assay
4.7. Intracellular Protein and Nucleic Acid Leakage Assay
4.8. Alkaline Phosphatase (AKP) Release Determination and Scanning Electron Microscopy (SEM)
4.9. Metabolomic Analysis
4.10. Real-Time PCR (RT-PCR) Analysis Related Gene Expressions of MRSA
4.11. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
Sample Availability
References
- Silva, J.C.; Rodrigues, S.S.Q.; Feás, X.; Estevinho, M.L.M.F. Antimicrobial activity, phenolic profile and role in the inflammation of propolis. Food Chem. Toxicol. 2012, 50, 1790–1795. [Google Scholar] [CrossRef] [PubMed]
- Rufatto, L.C.; Luchtenberg, P.; Garcia, C.; Thomassigny, C.; Bouttier, S.; Henriques, J.A.P.; Roesch-Ely, M.; Dumas, F.; Moura, S. Brazilian red propolis: Chemical composition and antibacterial activity determined using bioguided fractionation. Microbiol. Res. 2018, 214, 74–82. [Google Scholar] [CrossRef] [PubMed]
- Tosic, S.; Stojanovic, G.; Mitic, S.; Pavlovic, A.; Alagic, S. Mineral Composition of Selected Serbian Propolis Samples. J. Apic. Sci. 2017, 61, 5–15. [Google Scholar] [CrossRef]
- Machado, B.A.S.; Silva, R.P.D.; Barreto, G.; Costa, S.S.; da Silva, D.F.; Brandão, H.N.; Da Rocha, J.L.C.; Dellagostin, O.A.; Henriques, J.A.P.; Umsza-Guez, M.A.; et al. Chemical Composition and Biological Activity of Extracts Obtained by Supercritical Extraction and Ethanolic Extraction of Brown, Green and Red Propolis Derived from Different Geographic Regions in Brazil. PLoS ONE 2016, 11, e0145954. [Google Scholar] [CrossRef]
- Teixeira, É.W.; Negri, G.; Meira, R.M.; Salatino, A. Plant Origin of Green Propolis: Bee Behavior, Plant Anatomy and Chemistry. Evid.-Based Complement. Altern. Med. 2005, 2, 85–92. [Google Scholar] [CrossRef]
- Silva, B.B.; Rosalen, P.L.; Cury, J.A.; Ikegaki, M.; Souza, V.C.; Esteves, A.; Alencar, S.M. Chemical Composition and Botanical Origin of Red Propolis, a New Type of Brazilian Propolis. Evid.-Based Complement. Altern. Med. 2008, 5, 313–316. [Google Scholar] [CrossRef]
- Gong, S.; Luo, L.; Gong, W.; Gao, Y.; Xie, M. Multivariate analyses of element concentrations revealed the groupings of propolis from different regions in China. Food Chem. 2012, 134, 583–588. [Google Scholar] [CrossRef]
- Wu, Y.-W.; Sun, S.-Q.; Zhao, J.; Li, Y.; Zhou, Q. Rapid discrimination of extracts of Chinese propolis and poplar buds by FT-IR and 2D IR correlation spectroscopy. J. Mol. Struct. 2008, 883, 48–54. [Google Scholar] [CrossRef]
- Jiang, X.; Tian, J.; Zheng, Y.; Zhang, Y.; Wu, Y.; Zhang, C.; Zheng, H.; Hu, F. A New Propolis Type from Changbai Mountains in North-east China: Chemical Composition, Botanical Origin and Biological Activity. Molecules 2019, 24, 1369. [Google Scholar] [CrossRef]
- Wolska, K.; Gorska, A.; Antosik, K.; Lugowska, K. Immunomodulatory Effects of Propolis and its Components on Basic Immune Cell Functions. Indian J. Pharm. Sci. 2019, 81, 575–588. [Google Scholar] [CrossRef]
- Anjum, S.I.; Ullah, A.; Khan, K.A.; Attaullah, M.; Khan, H.; Ali, H.; Bashir, M.A.; Tahir, M.; Ansari, M.J.; Ghramh, H.A.; et al. Composition and functional properties of propolis (bee glue): A review. Saudi J. Biol. Sci. 2019, 26, 1695–1703. [Google Scholar] [CrossRef]
- Pimenta, H.C.; Violante, I.M.P.; De Musis, C.R.; Borges, Á.H.; Aranha, A.M.F. In vitro effectiveness of Brazilian brown propolis against Enterococcus faecalis. Braz. Oral Res. 2015, 29, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Silici, S.; Kutluca, S. Chemical composition and antibacterial activity of propolis collected by three different races of honeybees in the same region. J. Ethnopharmacol. 2005, 99, 69–73. [Google Scholar] [CrossRef] [PubMed]
- Przybyłek, I.; Karpiński, T.M. Antibacterial Properties of Propolis. Molecules 2019, 24, 2047. [Google Scholar] [CrossRef] [PubMed]
- Yuan, Z.; Ouyang, P.; Gu, K.; Rehman, T.; Zhang, T.; Yin, Z.; Fu, H.; Lin, J.; He, C.; Shu, G.; et al. The antibacterial mechanism of oridonin against methicillin-resistant Staphylococcus aureus (MRSA). Pharm. Biol. 2019, 57, 710–716. [Google Scholar] [CrossRef] [PubMed]
- Ferrera-Suanzes, M.; Prieto, V.; Medina-Olivera, A.J.; Botubol-Ares, J.M.; Galán-Sánchez, F.; Rodríguez-Iglesias, M.A.; Hernández-Galán, R.; Durán-Peña, M.J. Synthesis of Degraded Limonoid Analogs as New Antibacterial Scaffolds against Staphylococcus aureus. Antibiotics 2020, 9, 488. [Google Scholar] [CrossRef] [PubMed]
- Periasamy, H.; Iswarya, S.; Pavithra, N.; Senthilnathan, S.; Gnanamani, A. In vitro antibacterial activity of plumbagin isolated from Plumbago zeylanica L. against methicillin-resistant Staphylococcus aureus. Lett. Appl. Microbiol. 2019, 69, 41–49. [Google Scholar] [CrossRef]
- DeLeo, F.R.; Chambers, H.F. Reemergence of antibiotic-resistant Staphylococcus aureus in the genomics era. J. Clin. Investig. 2009, 119, 2464–2474. [Google Scholar] [CrossRef]
- Nguyen, H.M.; Graber, C.J. Limitations of antibiotic options for invasive infections caused by methicillin-resistant Staphylococcus aureus: Is combination therapy the answer? J. Antimicrob. Chemother. 2010, 65, 24–36. [Google Scholar] [CrossRef]
- Iwata, Y.; Sakai, N.; Yoneda, I.; Satou, K.; Furuichi, K.; Senda, Y.; Sakai-Takemori, Y.; Wada, T.; Fujita, S.; Ogura, H.; et al. The increased frequency of methicillin-resistant Staphylococcus aureus with low MIC of beta-lactam antibiotics isolated from hospitalized patients. J. Infect. Chemother. 2020, 26, 604–610. [Google Scholar] [CrossRef]
- Zhang, W.; Qi, S.; Xue, X.; Al Naggar, Y.; Wu, L.; Wang, K. Understanding the Gastrointestinal Protective Effects of Polyphenols using Foodomics-Based Approaches. Front. Immunol. 2021, 12, 671150. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Tang, C.; Zhang, R.; Ye, S.; Zhao, Z.; Huang, Y.; Xu, X.; Lan, W.; Yang, D. Metabolomics analysis to evaluate the antibacterial activity of the essential oil from the leaves of Cinnamomum camphora (Linn.) Presl. J. Ethnopharmacol. 2020, 253, 112652. [Google Scholar] [CrossRef] [PubMed]
- He, R.; Chen, W.; Zhang, M.; Zhong, Q.; Zhang, H.; Chen, H.; Chen, W. Antibacterial mechanism of linalool against L. monocytogenes, a metabolomic study. Food Control 2021, 132, 108533. [Google Scholar] [CrossRef]
- Alencar, S.; Oldoni, T.; Castro, M.; Cabral, I.; Costa-Neto, C.; Cury, J.; Rosalen, P.L.; Ikegaki, M. Chemical composition and biological activity of a new type of Brazilian propolis: Red propolis. J. Ethnopharmacol. 2007, 113, 278–283. [Google Scholar] [CrossRef]
- Hatano, A.; Nonaka, T.; Yoshino, M.; Ahn, M.-R.; Tazawa, S.; Araki, Y.; Kumazawa, S. Antioxidant Activity and Phenolic Constituents of Red Propolis from Shandong, China. Food Sci. Technol. Res. 2012, 18, 577–584. [Google Scholar] [CrossRef]
- Izuta, H.; Shimazawa, M.; Tsuruma, K.; Araki, Y.; Mishima, S.; Hara, H. Bee products prevent VEGF-induced angiogenesis in human umbilical vein endothelial cells. BMC Complement. Altern. Med. 2009, 9, 45. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.-H.; Cai, M.; Liu, Y.-S.; Sun, P.L.; Luo, S.-L. Antibacterial Activity and Mechanisms of Essential Oil from Citrus medica L. var. sarcodactylis. Molecules 2019, 24, 1577. [Google Scholar] [CrossRef]
- Hu, F.L.; Zhu, W.; Li, Y.J.; Zheng, H.Q. Study on Chemical and Biological Active Diversity of Propolis. J. Bee 2006, 1. [Google Scholar]
- Bankova, V.; Popova, M.; Trusheva, B. Propolis volatile compounds: Chemical diversity and biological activity: A review. Chem. Cent. J. 2014, 8, 28. [Google Scholar] [CrossRef]
- Darwish, R.M.; Abu Fares, R.J.; Abu Zarga, M.H.; Nazer, I.K. Antibacterial effect of Jordanian propolis and isolated flavonoids against human pathogenic bacteria. Afr. J. Biotechnol. 2010, 9, 5966–5974. [Google Scholar] [CrossRef]
- Kopacz, M.; Woźnicka, E.; Gruszecka, J. Antibacterial activity of morin and its complexes with La(III), Gd(III) and Lu(III) ions. Acta Pol. Pharm.-Drug Res. 2005, 62, 65–67. [Google Scholar]
- Kishimoto, N.; Kakino, Y.; Iwai, K.; Mochida, K.; Fujita, T. In Vitro Antibacterial, Antimutagenic and Anti-Influenza Virus Activity of Caffeic Acid Phenethyl Esters. Biocontrol. Sci. 2005, 10, 155–161. [Google Scholar] [CrossRef]
- Kharsany, K. The Antimicrobial Properties of The Major Compounds Found in South African Propolis; University of the Witwatersrand, Faculty of Health Sciences: Johannesburg, South Africa, 2019. [Google Scholar]
- Kheadr, E.; Bernoussi, N.; Lacroix, C.; Fliss, I. Comparison of the sensitivity of commercial strains and infant isolates of bifidobacteria to antibiotics and bacteriocins. Int. Dairy J. 2004, 14, 1041–1053. [Google Scholar] [CrossRef]
- Phillips, C.; Duggan, J. The effect of temperature and citric acid, alone, and in combination with nisin, on the growth of Arcobacter butzleri in culture. Food Control. 2002, 13, 463–468. [Google Scholar] [CrossRef]
- Huber, B.; Eberl, L.; Feucht, W.; Polster, J. Influence of Polyphenols on Bacterial Biofilm Formation and Quorum-sensing. Z. Für Nat. C 2003, 58, 879–884. [Google Scholar] [CrossRef]
- Almajano, M.; Carbó, R.; Delgado, M.; Gordon, M. Effect of pH on the Antimicrobial Activity and Oxidative Stability of Oil-in-Water Emulsions Containing Caffeic Acid. J. Food Sci. 2007, 72, C258–C263. [Google Scholar] [CrossRef]
- Bankova, V.; Marcucci, M.; Simova, S.; Nikolova, N.; Kujumgiev, A.; Popov, S. Antibacterial Diterpenic Acids from Brazilian Propolis. Z. Für Nat. C 1996, 51, 277–280. [Google Scholar] [CrossRef]
- Rahman, M.M.; Richardson, A.; Sofian-Azirun, M. Antibacterial activity of propolis and honey against Staphylococcus aureus and Escherichia coli. Afr. J. Microbiol. Res. 2010, 4, 1872. [Google Scholar] [CrossRef]
- Strahl, H.; Errington, J. Bacterial Membranes: Structure, Domains, and Function. Annu. Rev. Microbiol. 2017, 71, 519–538. [Google Scholar] [CrossRef]
- Typas, A.; Sourjik, V. Bacterial protein networks: Properties and functions. Nat. Rev. Genet. 2015, 13, 559–572. [Google Scholar] [CrossRef]
- Setlow, B.; Setlow, P. Binding of small, acid-soluble spore proteins to DNA plays a significant role in the resistance of Bacillus subtilis spores to hydrogen peroxide. Appl. Environ. Microb. 1993, 59, 3418–3423. [Google Scholar] [CrossRef] [PubMed]
- Diao, W.-R.; Hu, Q.-P.; Zhang, H.; Xu, J.-G. Chemical composition, antibacterial activity and mechanism of action of essential oil from seeds of fennel (Foeniculum vulgare Mill.). Food Control. 2014, 35, 109–116. [Google Scholar] [CrossRef]
- Zhou, D.; Liu, Z.-H.; Wang, D.-M.; Li, D.-W.; Yang, L.-N.; Wang, W. Chemical composition, antibacterial activity and related mechanism of valonia and shell from Quercus variabilis Blume (Fagaceae) against Salmonella paratyphi a and Staphylococcus aureus. BMC Complement. Altern. Med. 2019, 19, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Wang, F.; Wei, F.; Song, C.; Jiang, B.; Tian, S.; Yi, J.; Yu, C.; Song, Z.; Sun, L.; Bao, Y.; et al. Dodartia orientalis L. essential oil exerts antibacterial activity by mechanisms of disrupting cell structure and resisting biofilm. Ind. Crop. Prod. 2017, 109, 358–366. [Google Scholar] [CrossRef]
- Yuan, L.-H.; Hu, Q.-P.; Xu, J.-G. Antibacterial Mechanism of The Ehyl Acetate Extracts From Naked Oat Again Bacillus subtilis. Aceh Int. J. Sci. Technol. 2014, 4. [Google Scholar] [CrossRef]
- Wang, F.; Liu, H.; Li, J.; Zhang, W.; Jiang, B.; Xuan, H. Australian propolis ethanol extract exerts antibacterial activity against methicillin-resistant Staphylococcus aureus by mechanisms of disrupting cell structure, reversing resistance, and resisting biofilm. Braz. J. Microbiol. 2021, 52, 1651–1664. [Google Scholar] [CrossRef]
- Bryan, J.; Redden, P.; Traba, C. The mechanism of action of Russian propolis ethanol extracts against two antibiotic-resistant biofilm-forming bacteria. Lett. Appl. Microbiol. 2016, 62, 192–198. [Google Scholar] [CrossRef]
- He, F.; Li, K.; Zhang, X.; Yang, Y.; Fang, Y.; Xiang, F. Components and antibacterial activity of a novel essential oil from the nutrient broth of Eremothecium ashbyii H4565. LWT 2018, 101, 389–394. [Google Scholar] [CrossRef]
- An, P.; Yang, X.; Yu, J.; Qi, J.; Ren, X.; Kong, Q. α-terpineol and terpene-4-ol, the critical components of tea tree oil, exert antifungal activities in vitro and in vivo against Aspergillus niger in grapes by inducing morphous damage and metabolic changes of fungus. Food Control 2019, 98, 42–53. [Google Scholar] [CrossRef]
- Belenky, P.; Ye, J.D.; Porter, C.B.; Cohen, N.R.; Lobritz, M.A.; Ferrante, T.; Jain, S.; Korry, B.J.; Schwarz, E.G.; Walker, G.C.; et al. Bactericidal Antibiotics Induce Toxic Metabolic Perturbations that Lead to Cellular Damage. Cell Rep. 2015, 13, 968–980. [Google Scholar] [CrossRef]
- Schelli, K.; Zhong, F.; Zhu, J. Comparative metabolomics revealing Staphylococcus aureus metabolic response to different antibiotics. Microb. Biotechnol. 2017, 10, 1764–1774. [Google Scholar] [CrossRef] [PubMed]
- Liu, M.; Feng, M.; Yang, K.; Cao, Y.; Zhang, J.; Xu, J.; Hernández, S.H.; Wei, X.; Fan, M. Transcriptomic and metabolomic analyses reveal antibacterial mechanism of astringent persimmon tannin against Methicillin-resistant Staphylococcus aureus isolated from pork. Food Chem. 2020, 309, 125692. [Google Scholar] [CrossRef]
- Hussein, M.; Karas, J.A.; Schneider-Futschik, E.K.; Chen, F.; Swarbrick, J.; Paulin, O.K.A.; Hoyer, D.; Baker, M.; Zhu, Y.; Li, J.; et al. The Killing Mechanism of Teixobactin against Methicillin-Resistant Staphylococcus aureus: An Untargeted Metabolomics Study. MSystems 2020, 5, 00077-20. [Google Scholar] [CrossRef] [PubMed]
- Selvaraj, A.; Jayasree, T.; Valliammai, A.; Pandian, S.K. Myrtenol Attenuates MRSA Biofilm and Virulence by Suppressing sarA Expression Dynamism. Front. Microbiol. 2019, 10, 2027. [Google Scholar] [CrossRef] [PubMed]
- Arciola, C.R.; Campoccia, D.; Baldassarri, L.; Donati, M.E.; Pirini, V.; Gamberini, S.; Montanaro, L. Detection of biofilm formation inStaphylococcus epidermidis from implant infections. Comparison of a PCR-method that recognizes the presence ofica genes with two classic phenotypic methods. J. Biomed. Mater. Res. Part A 2005, 76, 425–430. [Google Scholar] [CrossRef] [PubMed]
- Ahmadrajabi, R.; Layegh-Khavidaki, S.; Kalantar-Neyestanaki, D.; Fasihi, Y. Molecular analysis of immune evasion cluster (IEC) genes and intercellular adhesion gene cluster (ICA) among methicillin-resistant and methicillin-sensitive isolates of Staphylococcus aureus. J. Prev. Med. Hyg. 2017, 58, E308–E314. [Google Scholar] [CrossRef] [PubMed]
- Bronner, S.; Monteil, H.; Prévost, G. Regulation of virulence determinants in Staphylococcus aureus: Complexity and applications. FEMS Microbiol. Rev. 2004, 28, 183–200. [Google Scholar] [CrossRef]
- Bai, J.; Wu, Y.; Liu, X.; Zhong, K.; Huang, Y.; Gao, H. Antibacterial Activity of Shikimic Acid from Pine Needles of Cedrus deodara against Staphylococcus aureus through Damage to Cell Membrane. Int. J. Mol. Sci. 2015, 16, 27145–27155. [Google Scholar] [CrossRef]
- Wang, K.; Zhang, J.; Ping, S.; Ma, Q.; Chen, X.; Xuan, H.; Shi, J.; Zhang, C.; Hu, F. Anti-inflammatory effects of ethanol extracts of Chinese propolis and buds from poplar (Populus × canadensis). J. Ethnopharmacol. 2014, 155, 300–311. [Google Scholar] [CrossRef]
- Liu, X.; Zhong, S.; Wu, K.; Yu, H.; Chai, X. Antimicrobial activity in the vapour phase of a combination of cinnamon and clove essential oils. Food Chem. 2009, 116, 982–989. [Google Scholar]
- Elnakady, Y.A.; Chatterjee, I.; Bischoff, M.; Rohde, M.; Josten, M.; Sahl, H.-G.; Herrmann, M.; Müller, R. Investigations to the Antibacterial Mechanism of Action of Kendomycin. PLoS ONE 2016, 11, e0146165. [Google Scholar] [CrossRef] [PubMed]
- Xu, C.; Li, J.; Yang, L.; Shi, F.; Yang, L.; Ye, M. Antibacterial activity and a membrane damage mechanism of Lachnum YM30 melanin against Vibrio parahaemolyticus and Staphylococcus aureus. Food Control 2017, 73, 1445–1451. [Google Scholar] [CrossRef]
- Sannasiddappa, T.H.; Lund, P.A.; Clarke, S.R.; Sannasiddappa, T. In Vitro Antibacterial Activity of Unconjugated and Conjugated Bile Salts on Staphylococcus aureus. Front. Microbiol. 2017, 8, 1581. [Google Scholar] [CrossRef] [PubMed]
- Hu, W.; Li, C.; Dai, J.; Cui, H.; Lin, L. Antibacterial activity and mechanism of Litsea cubeba essential oil against methicillin-resistant Staphylococcus aureus (MRSA). Ind. Crop. Prod. 2019, 130, 34–41. [Google Scholar] [CrossRef]
- Tang, C.; Chen, J.; Zhang, L.; Zhang, R.; Zhang, S.; Ye, S.; Zhao, Z.; Yang, D. Exploring the antibacterial mechanism of essential oils by membrane permeability, apoptosis and biofilm formation combination with proteomics analysis against methicillin-resistant staphylococcus aureus. Int. J. Med. Microbiol. 2020, 310, 151435. [Google Scholar] [CrossRef] [PubMed]
- Kot, B.; Sytykiewicz, H.; Sprawka, I. Expression of the Biofilm-Associated Genes in Methicillin-Resistant Staphylococcus aureus in Biofilm and Planktonic Conditions. Int. J. Mol. Sci. 2018, 19, 3487. [Google Scholar] [CrossRef]
- Qu, X.; Yang, H.; Jia, B.; Yu, Z.; Zheng, Y.; Dai, K. Biodegradable Zn–Cu alloys show antibacterial activity against MRSA bone infection by inhibiting pathogen adhesion and biofilm formation. Acta Biomater. 2020, 117, 400–417. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Compound Name | Retention Time | Precursor (m/z) | Product Ions (m/z) | CE | RPE (mg/g) |
|---|---|---|---|---|---|
| Pinobanksin-3-acetate | 7.813 | 313.1 | 253.1 | 15 | 53.79 |
| 271.0 | 15 | ||||
| Morin | 6.609 | 301 | 151 | 16 | 1.04 |
| 125 | 16 | ||||
| Ferulic acid | 5.487 | 192.9 | 178.1 | 15 | 2.82 |
| 134.2 | 10 | ||||
| Caffeic acid | 4.798 | 179 | 135 | 10 | 8.75 |
| 107.2 | 20 | ||||
| Caffeic acid phenethyl ester | 7.995 | 283.1 | 179 | 15 | 14.39 |
| 135.1 | 25 | ||||
| Pinobanksin | 6.819 | 271.1 | 253 | 15 | 194.05 |
| 125 | 20 | ||||
| Galangin | 8.268 | 269.1 | 213.1 | 20 | 25.33 |
| 171 | 25 | ||||
| Quercetin | 6.609 | 301 | 179 | 18 | 1.01 |
| 151 | 22 | ||||
| P-coumaric acid | 5.384 | 163.1 | 116.8 | 30 | 8.75 |
| 119 | 10 | ||||
| Apigenin | 7.242 | 269 | 151.1 | 20 | 14.39 |
| 117.1 | 40 | ||||
| Kaempferol | 7.114 | 285 | 257 | 20 | 1.74 |
| 93 | 35 | ||||
| Chrysin | 8.188 | 253.1 | 143 | 24 | 44.51 |
| 63 | 32 | ||||
| Pinocembrin | 7.811 | 255.1 | 213 | 15 | 18.66 |
| 151.2 | 20 | ||||
| Naringenin | 6.685 | 271.1 | 151 | 10 | 0.67 |
| 119 | 25 |
| Sample | S. aureus ATCC25923 | MRSA ATCC43300 | ||
|---|---|---|---|---|
| MIC (µg/mL) | MBC (µg/mL) | MIC (µg/mL) | MBC (µg/mL) | |
| RPE | 100 | 300 | 50 | 200 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, W.; Margarita, G.E.; Wu, D.; Yuan, W.; Yan, S.; Qi, S.; Xue, X.; Wang, K.; Wu, L. Antibacterial Activity of Chinese Red Propolis against Staphylococcus aureus and MRSA. Molecules 2022, 27, 1693. https://doi.org/10.3390/molecules27051693
Zhang W, Margarita GE, Wu D, Yuan W, Yan S, Qi S, Xue X, Wang K, Wu L. Antibacterial Activity of Chinese Red Propolis against Staphylococcus aureus and MRSA. Molecules. 2022; 27(5):1693. https://doi.org/10.3390/molecules27051693
Chicago/Turabian StyleZhang, Wenwen, Gomez Escalada Margarita, Di Wu, Wenqin Yuan, Sha Yan, Suzhen Qi, Xiaofeng Xue, Kai Wang, and Liming Wu. 2022. "Antibacterial Activity of Chinese Red Propolis against Staphylococcus aureus and MRSA" Molecules 27, no. 5: 1693. https://doi.org/10.3390/molecules27051693
APA StyleZhang, W., Margarita, G. E., Wu, D., Yuan, W., Yan, S., Qi, S., Xue, X., Wang, K., & Wu, L. (2022). Antibacterial Activity of Chinese Red Propolis against Staphylococcus aureus and MRSA. Molecules, 27(5), 1693. https://doi.org/10.3390/molecules27051693