
Machine Learning Analysis of Essential Oils from Cuban Plants: Potential Activity against Protozoa Parasites


 ,
,  ,
,  , and
, and 
Abstract
1. Introduction
2. Results and Discussion

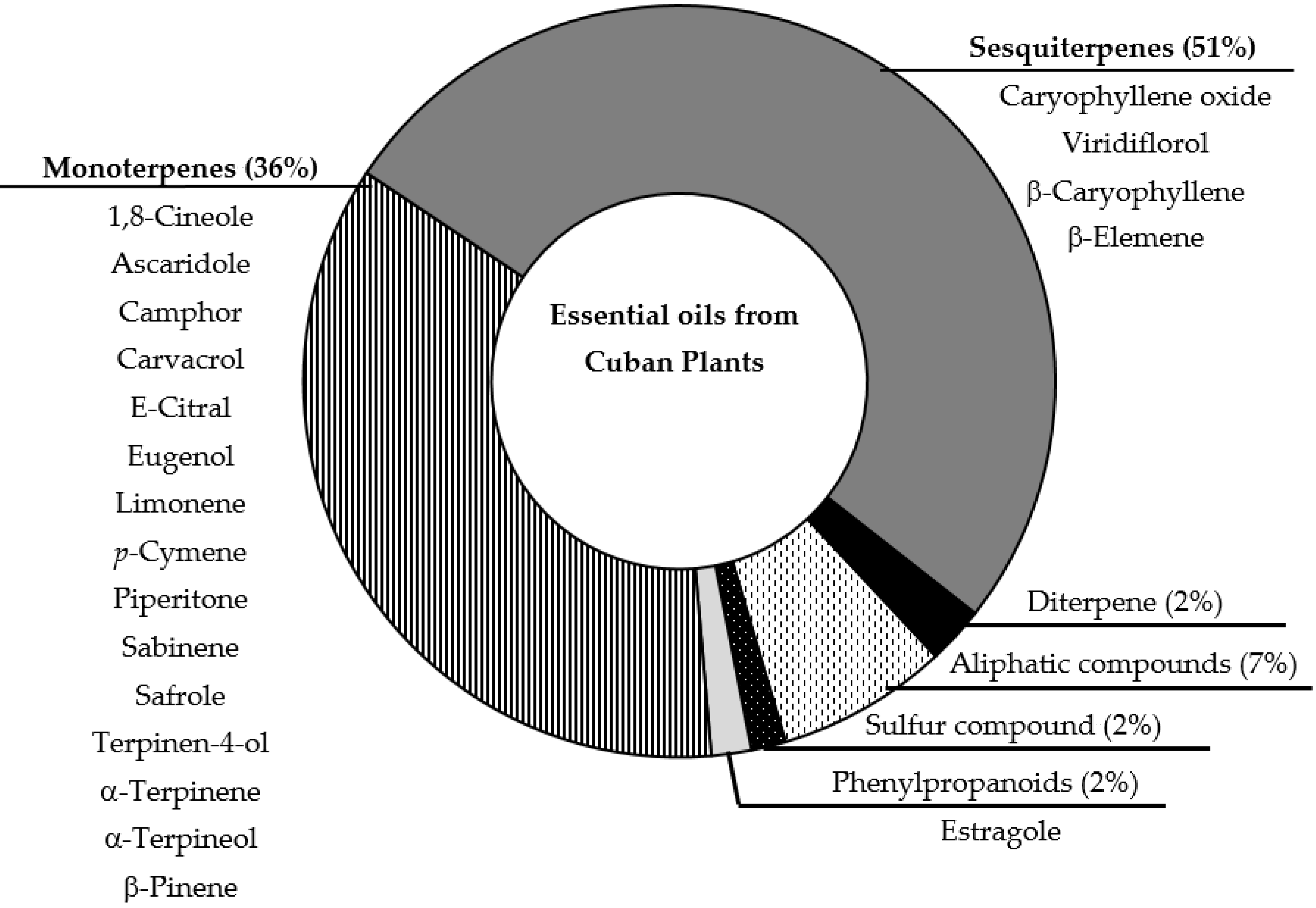{kind=link}
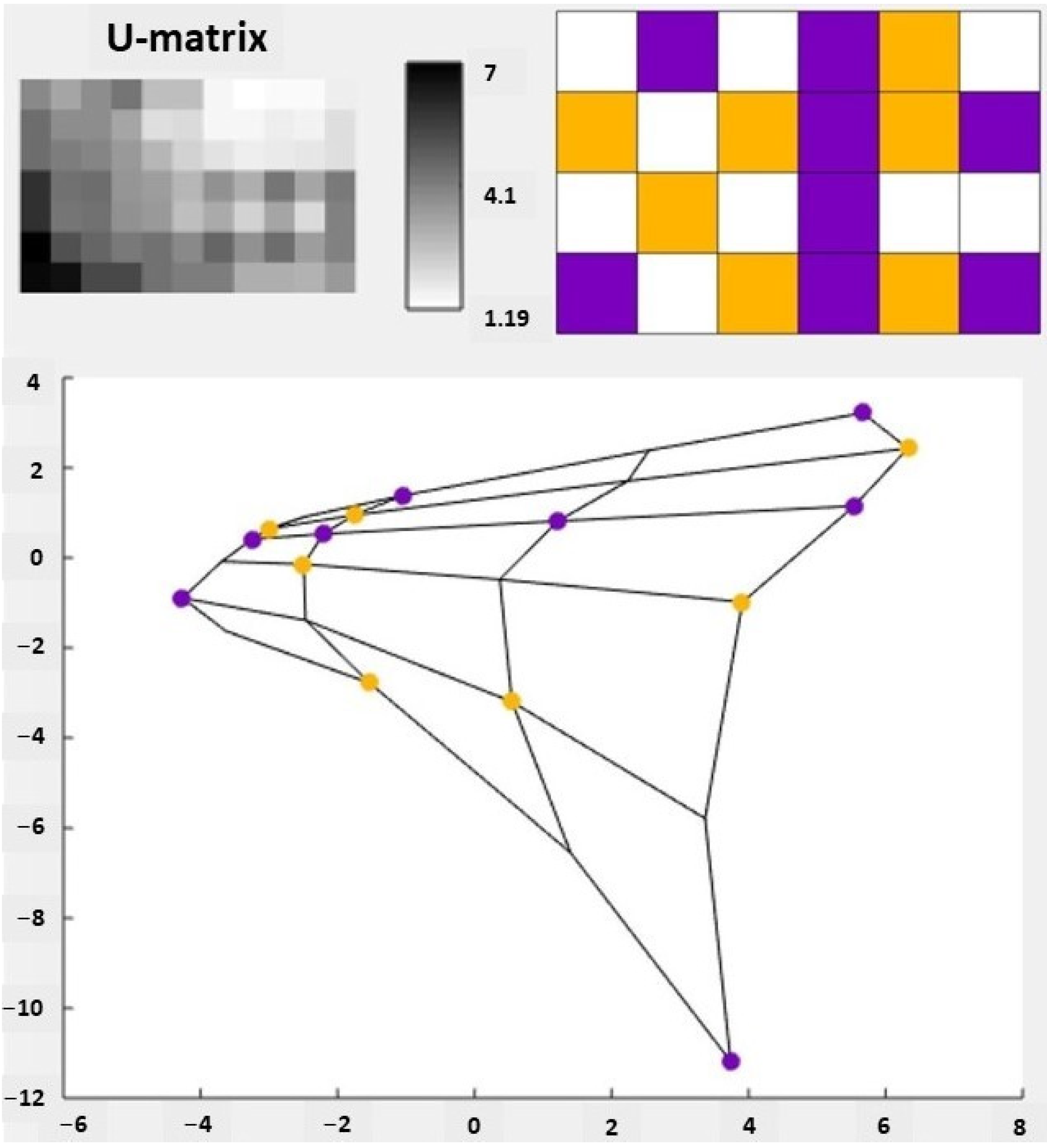{kind=link}
{kind=link}
| Compound | Country | Plant | Compound % | Targeted Protozoa (Result) | Ref. |
|---|---|---|---|---|---|
| (E)-β-Ocimene | Brazil | Annona vepretorum Mart. | 6.8% | Trypanosoma cruzi (IC50 = 32 μg/mL) | [51] |
| Syzygium cumini (L.) Skeels. | 11.7% | Leishmania amazonensis (IC50 = 60 mg/L) | [52] | ||
| Xylopia frutescens Aubl. | 6.8% | Trypanosoma cruzi (IC50 = 15 to 30 μg/mL) | [53] | ||
| Cuba * | Bursera graveolens Triana & Planch | 13% | Leishmania amazonensis (IC50 = 36.7 mg/L) | [54] | |
| Piper auritum Kunt | 0.49% | Leishmania major (IC50 = 29.1 μg/mL) Leishmania mexicana (IC50 = 63.3 μg/mL) Leishmania braziliensis (IC50 = 52.1 μg/mL) Leishmania donovani (IC50 = 12.8 μg/mL) | [55] | ||
| (Z)-β-Ocimene | Brazil | Syzygium cumini (L.) Skeels. | 29% | Leishmania amazonensis (60 mg/L) | [53] |
| Cuba * | Bursera graveolens Triana & Planch | 0.9% | Leishmania amazonensis (IC50 = 36.7 mg/L) | [54] | |
| Piper ossanum (C.DC.) Trel | 0.14% | Plasmodium falciparum (IC50 = 1.5 µg/mL) Trypanosoma brucei (IC50 = 8.1 µg/mL) Trypanosoma cruzi (IC50 = 8.0 µg/mL) Leishmania amazonensis (IC50 = 19.3 µg/mL) | [47] | ||
| β-Phellandrene | Cameroon | Ocimum gratissimum L. | 21.1% | Plasmodium berghei (at 200, 300 and 500 mg/kg caused a suppression of parasitaemia of 55.0%, 75.2% and 77.8%, respectively) | [56] |
| Cuba * | Bixa orellana L. | 0.2% | Leishmania amazonensis (8.5 mg/L) | [57] | |
| Piper ossanum (C.DC.) Trel | 2.1% | Plasmodium falciparum (IC50 = 2.8 µg/mL) Trypanosoma brucei (IC50 = 8.4 µg/mL) Trypanosoma cruzi (IC50 = 8.6 µg/mL) | [47] | ||
| Camphor | Brazil | Alpinia speciosa K. Schum | 17.1% | Trypanosoma cruzi (IC50 = 92 μg/mL) Leishmania brasiliensis (IC50 = 67 μg/mL) | [58] |
| Ocotea odorifera (Vell) Rohwer | 6.5% | Leishmania amazonensis (IC50 = 11.7 μg/mL) | [59] | ||
| Piper cubeba L | 5.6% | Trypanosoma cruzi (IC50 = 87.9 µg/mL) | [40] | ||
| Cuba * | Alpinia zerumbet (Pers.) B.L.Burtt & R.M.Smith | 0.1% | Plasmodium falciparum (IC50 = 66.2 µg/mL) | [45] | |
| Piper aduncum L. | 17.1% | Leishmania amazonensis (IC50 = 23.8 μg/mL) Leishmania donovani (IC50 = 7.7 μg/mL) Leishmania infantum (IC50 = 8.1 μg/mL) | [46] | ||
| Piper ossanum (C.DC.) Trel | 13.8 and 9.4% | Plasmodium falciparum (IC50 = 1.5 and 2.8 µg/mL) Trypanosoma brucei (IC50 = 8.1 and 8.4 µg/mL) Trypanosoma cruzi (IC50 = 8.0 and 8.6 µg/mL) | [47] | ||
| Ethiopia | Artemisia absinthium L. | 27.4% | Leishmania aethiopica (IC50 = 8 μg/mL) Leishmania donovani (IC50 = 42 μg/mL) | [60] | |
| Morocco | Rosmarinus officinalis L. | 18.7% | Leishmania major (IC50 = 1.2 μg/mL) | [61] | |
| Spain | Artemisia absinthium L. | 4.5% | Trypanosoma cruzi (84% of inhibition at 200 μg/mL) | [62] | |
| Artemisia pedemontana subsp. assoana (Willk.) Rivas Mart. | 7.7% | Trypanosoma cruzi (20 to 70% of inhibition at 200 μg/mL) | [62] | ||
| Turkey | Salvia recognita Fisch. & Meyer | 42% | Plasmodium falciparum (IC50 = 17 to 12 µg/mL) | [63] | |
| Tunisia | Thymus hirtus sp. algeriensis Boiss. et Reut | 13.8% | Leishmania major (IC50 = 0.43 μg/mL) Leishmania infantum (IC50 = 0.25 μg/mL) | [39] | |
| Germacrene D | Brazil | Casearia sylvestris Sw. | 19.6% | Leishmania amazonensis (IC50 = 24.2 µg/mL) | [64] |
| Eugenia gracillima Kiaersk. | 16.1% | Leishmania braziliensis (IC50 = 74.6 µg/mL) Leishmania infantum (IC50 = 80.4 µg/mL) | [65] | ||
| Guatteria australis A. St.-Hil. | 22.2% | Leishmania infantum (IC50 = 30.7 μg/mL) | [66] | ||
| Lantana camara L. | 11.7% | Trypanosoma cruzi (IC50 = 201.94 μg/mL) Leishmania braziliensis (IC50 = 72.31 μg/mL) | [67] | ||
| Melampodium divaricatum (Rich. ex Rich.) DC. | 12.7% | Leishmania amazonensis (IC50 = 10.7 µg/mL) | [64] | ||
| Piper cernuum Vell. | 12.7% | Leishmania amazonensis (Infection index of 115 at 10 μg/mL) | [68] | ||
| Piper duckei C. DC. | 14.7% | Leishmania amazonensis (IC50 = 42–46 μg/mL) Leishmania guyanensis (IC50 = 15.2 μg/mL) | [69] | ||
| Vernonia polyanthes (Spreng.) Vega & M. Dematteis | 4.3% | Leishmania infantum (IC50 = 19.4 µg/mL) | [70] | ||
| Xylopia frutescens Aubl. | 17.8% | Trypanosoma cruzi (IC50 = 15 to 30 μg/mL) | [53] | ||
| Cuba * | Tagetes lucida Cav. | 0.3% | Plasmodium berghei (IC50 = 72 μg/mL) | [50] | |
| Methyl eugenol | Brazil | Aniba canelilla (H.B.K.) Mez | 14.8% | Trypanosoma evansi (Growth inhibition at 0.5%, 1.0% and 2.0%) | [71] |
| Hypenia salzmannii (Benth.) Harley. | 5.6% | Trypanosoma cruzi (IC50 = 35–42 µg/mL) | [72] | ||
| Cuba * | Piper auritum Kunt | 0.6% | Leishmania major (IC50 = 29.1 μg/mL) Leishmania mexicana (IC50 = 63.3 μg/mL) Leishmania braziliensis (IC50 = 52.1 μg/mL) Leishmania donovani (IC50 = 12.8 μg/mL) | [55] | |
| Bhutan | Pleurospermum amabile W.W.Smith, | 3.8% | Plasmodium falciparum (IC50 = 79 μg/mL) | [73] | |
| Piperitone | Benin | Cymbopogon schoenantus Spreng. | 60.3% | Trypanosoma brucei (IC50 = 2.1 µg/mL) Plasmodium falciparum (IC50 = 43.1 µg/mL) | [49] |
| Cuba * | Alpinia zerumbet (Pers.)B.L.Burtt&R.M.Smith | 0.1% | Plasmodium falciparum (IC50 = 71.4 µg/mL) | [45] | |
| Bursera graveolens Triana & Planch | 0.1% | Leishmania amazonensis (IC50 = 36.7 mg/L) | [54] | ||
| Piper aduncum L. | 23.7% | Leishmania amazonensis (IC50 = 23.8 μg/mL) Leishmania donovani (IC50 = 7.7 μg/mL) Leishmania infantum (IC50 = 8.1 μg/mL) | [46] | ||
| Piper ossanum (C.DC.) Trel | 20.1 and 19.0% | Plasmodium falciparum (IC50 = 1.5 and 2.8 µg/mL) Trypanosoma brucei (IC50 = 8.1 and 8.4 µg/mL) Trypanosoma cruzi (IC50 = 8.0 and 8.6 µg/mL) | [47] | ||
| Safrole | Brazil | Myroxylon peruiferum L.f. | 8.3% | Leishmania amazonensis (IC50 = 54–162 μg/mL) | [74] |
| Ocotea odorifera (Vell) Rohwer | 6.5% | Leishmania amazonensis (IC50 = 11.7 μg/mL) | [59] | ||
| Cuba * | Piper auritum Kunt | 71.8% | Leishmania major (IC50 = 29.1 μg/mL) Leishmania mexicana (IC50 = 63.3 μg/mL) Leishmania braziliensis (IC50 = 52.1 μg/mL) Leishmania donovani (IC50 = 12.8 μg/mL) | [75] |
3. Materials and Methods
3.1. Essential Oils Database
3.2. Machine Learning Analysis
3.2.1. Self-Organizing Maps (SOMs)
3.2.2. Principal Component Analysis (PCA)
3.2.3. Random Forest Model
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Sample Availability
References
- Yang, S.-K.; Tan, N.-P.; Chong, C.-W.; Abushelaibi, A.; Lim, S.-H.; Lai, K.-S. The missing piece: Recent approaches investigating the antimicrobial mode of action of essential oils. Evol. Bioinform. 2021, 17, 1176934320938391. [Google Scholar] [CrossRef] [PubMed]
- Galvan, D.; Effting, L.; Neto, L.T.; Conte-Junior, C.A. An overview of research of essential oils by self-organizing maps: A novel approach for meta-analysis study. Compr. Rev. Food Sci. Food Saf. 2021, 20, 3136–3163. [Google Scholar] [CrossRef]
- Sabatino, M.; Fabiani, M.; Božović, M.; Garzoli, S.; Antonini, L.; Marcocci, M.E.; Palamara, A.T.; De Chiara, G.; Ragno, R. Experimental data based machine learning classification models with predictive ability to select in vitro active antiviral and non-toxic essential oils. Molecules 2020, 25, 2452. [Google Scholar] [CrossRef] [PubMed]
- Bakkali, F.; Averbeck, S.; Averbeck, D.; Idaomar, M. Biological effects of essential oils—A review. Food Chem. Toxicol. 2008, 46, 446–475. [Google Scholar] [CrossRef] [PubMed]
- Falleh, H.; Ben Jemaa, M.; Saada, M.; Ksouri, R. Essential oils: A promising eco-friendly food preservative. Food Chem. 2020, 330, 127268. [Google Scholar] [CrossRef] [PubMed]
- Lucia, A.; Guzmán, E. Emulsions containing essential oils, their components or volatile semiochemicals as promising tools for insect pest and pathogen management. Adv. Colloid Interface Sci. 2020, 287, 102330. [Google Scholar] [CrossRef] [PubMed]
- Agreles, M.A.A.; Cavalcanti, I.D.L.; Cavalcanti, I.M.F. The role of essential oils in the inhibition of efflux pumps and reversion of bacterial resistance to antimicrobials. Curr. Microbiol. 2021, 78, 3609–3619. [Google Scholar] [CrossRef]
- Wińska, K.; Mączka, W.; Łyczko, J.; Grabarczyk, M.; Czubaszek, A.; Szumny, A. Essential oils as antimicrobial agents-myth or real alternative? Molecules 2019, 24, 2130. [Google Scholar] [CrossRef]
- Cai, J.; Yan, R.; Shi, J.; Chen, J.; Long, M.; Wu, W.; Kuca, K. Antifungal and mycotoxin detoxification ability of essential oils: A review. Phytother. Res. 2021, 36, 62–72. [Google Scholar] [CrossRef]
- Luna, E.C.; Luna, I.S.; Scotti, L.; Monteiro, A.F.M.; Scotti, M.T.; De Moura, R.O.; De Araújo, R.S.A.; Monteiro, K.L.C.; De Aquino, T.M.; Ribeiro, F.F.; et al. Active essential oils and their components in use against neglected diseases and arboviruses. Oxidative Med. Cell. Longev. 2019, 45, 6587150. [Google Scholar] [CrossRef]
- Reichling, J. Antiviral and virucidal properties of essential oils and isolated compounds – A scientific approach. Planta Med. 2021. [Google Scholar] [CrossRef] [PubMed]
- Jankowska, M.; Rogalska, J.; Wyszkowska, J.; Stankiewicz, M. Molecular targets for components of essential oils in the insect nervous system—A review. Molecules 2018, 23, 34. [Google Scholar] [CrossRef] [PubMed]
- Andrade, M.; Braga, M.A.; Cesar, P.H.S.; Trento, M.V.C.; Espósito, M.A.; Silva, L.F.; Marcussi, S. Anticancer properties of essential oils: An overview. Curr. Cancer Drug Targets 2018, 18, 957–966. [Google Scholar] [CrossRef] [PubMed]
- Spisni, E.; Petrocelli, G.; Imbesi, V.; Spigarelli, R.; Azzinnari, D.; Sarti, M.D.; Campieri, M.; Valerii, M.C. Antioxidant, anti-inflammatory, and microbial-modulating activities of essential oils: Implications in colonic pathophysiology. Int. J. Mol. Sci. 2020, 21, 4152. [Google Scholar] [CrossRef]
- Poma, P.; Labbozzetta, M.; Ramarosandratana, A.; Rosselli, S.; Tutone, M.; Sajeva, M.; Notarbartolo, M. In vitro modulation of P-glycoprotein activity by Euphorbia intisy essential oil on acute myeloid leukemia cell line HL-60R. Pharmaceuticals 2021, 14, 111. [Google Scholar] [CrossRef] [PubMed]
- Costa, R.G.A.; da Anunciação, T.A.; Araujo, M.D.S.; Souza, C.A.; Dias, R.B.; Sales, C.B.S.; Rocha, C.A.G.; Soares, M.B.P.; da Silva, F.M.A.; Koolen, H.H.F.; et al. In vitro and in vivo growth inhibition of human acute promyelocytic leukemia HL-60 cells by Guatteria megalophylla Diels (Annonaceae) leaf essential oil. Biomed. Pharmacother. 2020, 122, 109713. [Google Scholar] [CrossRef]
- Sandner, G.; Heckmann, M.; Weghuber, J. Immunomodulatory activities of selected essential oils. Biomolecules 2020, 10, 1139. [Google Scholar] [CrossRef]
- Patsilinakos, A.; Artini, M.; Papa, R.; Sabatino, M.; Božović, M.; Garzoli, S.; Vrenna, G.; Buzzi, R.; Manfredini, S.; Selan, L.; et al. Machine learning analyses on data including essential oil chemical composition and in vitro experimental antibiofilm activities against Staphylococcus species. Molecules 2019, 24, 890. [Google Scholar] [CrossRef]
- Monzote, L.; García, J.; González, R.; Scotti, M.T.; Setzer, W.N. Bioactive essential oils from Cuban plants: An inspiration to drug development. Plants 2021, 10, 2515. [Google Scholar] [CrossRef]
- Kohonen, T. Self-organized formation of topologically correct feature maps. Biol. Cybern. 1982, 43, 59–69. [Google Scholar] [CrossRef]
- Kohonen, T.; Kaski, S.; Lagus, K.; Salojärvi, J.; Honkela, J.; Paatero, V.; Saarela, A. Self organization of a massive document collection. IEEE Trans. Neural Netw. 2000, 11, 574–585. [Google Scholar] [CrossRef] [PubMed]
- Livingstone, D. Data Analysis for Chemists; Oxford University Press, Ed.; Oxford Science Publications: Oxford, UK, 1995; ISBN 0-19-855728-0. [Google Scholar]
- Menezes, R.; Sessions, Z.; Muratov, E.; Scotti, L.; Scotti, M. Secondary metabolites extracted from Annonaceae and chemotaxonomy study of terpenoids. J. Braz. Chem. Soc. 2021, 32, 2061–2070. [Google Scholar] [CrossRef]
- Vesanto, J.; Himberg, J.; Alhoniemi, E.; Parhankangas, J. Self−organizing map in matlab: The SOM toolbox. Proc. Matlab DSP Conf. 1999, 99, 35–40. [Google Scholar]
- Salzberg, S.L. Book Review: C4.5: Programs for Machine Learning by J. Ross Quinlan. Morgan Kaufmann Publishers, Inc., 1993. Mach. Learn. 1994, 16, 235–240. [Google Scholar] [CrossRef]
- Mao, J.; Akhtar, J.; Zhang, X.; Sun, L.; Guan, S.; Li, X.; Chen, G.; Liu, J.; Jeon, H.-N.; Kim, M.S.; et al. Comprehensive strategies of machine-learning-based quantitative structure-activity relationship models. Iscience 2021, 24, 103052. [Google Scholar] [CrossRef]
- Barros, R.P.C.; Da Cunha, E.V.L.; Catão, R.M.R.; Scotti, L.; Souza, M.S.R.; Brás, A.A.Q.; Scotti, M.T. Virtual screening of secondary metabolites of the genus Solanum with potential antimicrobial activity. Rev. Bras. Farmacogn. 2018, 28, 686–691. [Google Scholar] [CrossRef]
- Kovdienko, N.A.; Polishchuk, P.G.; Muratov, E.N.; Artemenko, A.G.; Kuz’Min, V.E.; Gorb, L.; Hill, F.; Leszczynski, J. Application of random forest and multiple linear regression techniques to QSPR prediction of an aqueous solubility for military compounds. Mol. Inform. 2010, 29, 394–406. [Google Scholar] [CrossRef]
- Papa, R.; Garzoli, S.; Vrenna, G.; Sabatino, M.; Sapienza, F.; Relucenti, M.; Donfrancesco, O.; Fiscarelli, E.V.; Artini, M.; Selan, L.; et al. Essential oils biofilm modulation activity, chemical and machine learning analysis. Application on Staphylococcus aureus isolates from cystic fibrosis patients. Int. J. Mol. Sci. 2020, 21, 9258. [Google Scholar] [CrossRef]
- Artini, M.; Papa, R.; Vrenna, G.; Lauro, C.; Ricciardelli, A.; Casillo, A.; Corsaro, M.M.; Tutino, M.L.; Parrilli, E.; Selan, L. Cold-adapted bacterial extracts as a source of anti-infective and antimicrobial compounds against Staphylococcus aureus. Future Microbiol. 2019, 14, 1369–1382. [Google Scholar] [CrossRef]
- El-Attar, N.E.; Hassan, M.K.; Alghamdi, O.A.; Awad, W.A. Deep learning model for classification and bioactivity prediction of essential oil-producing plants from Egypt. Sci. Rep. 2020, 10, 21349. [Google Scholar] [CrossRef]
- Fourches, D.; Pu, D.; Tassa, C.; Weissleder, R.; Shaw, S.Y.; Mumper, R.J.; Tropsha, A. Quantitative nanostructure−activity relationship modeling. ACS Nano 2010, 4, 5703–5712. [Google Scholar] [CrossRef]
- Cherkasov, A.; Muratov, E.N.; Fourches, D.; Varnek, A.; Baskin, I.I.; Cronin, M.; Dearden, J.C.; Gramatica, P.; Martin, Y.C.; Todeschini, R.; et al. QSAR modeling: Where have you been? Where are you going to? J. Med. Chem. 2014, 57, 4977–5010. [Google Scholar] [CrossRef]
- Arcoverde, G.F.B.; Almeida, C.M.; Ximenes, A.D.C.; Maeda, E.E.; Araújo, L.S. Identification of priority areas for forest restoration using self-organizing maps neural network. Bol. Ciênc. Geod. 2011, 1, 379–400. [Google Scholar] [CrossRef]
- Isah, M.; Ibrahim, M.A.; Mohammed, A.; Aliyu, A.B.; Masola, B.; Coetzer, T. A systematic review of pentacyclic triterpenes and their derivatives as chemotherapeutic agents against tropical parasitic diseases. Parasitology 2016, 143, 1219–1231. [Google Scholar] [CrossRef] [PubMed]
- Scotti, M.T.; Scotti, L.; Ishiki, H.; Ribeiro, F.F.; Da Cruz, R.M.D.; De Oliveira, M.P.; Mendonça, F.J.B. Natural products as a source for antileishmanial and antitrypanosomal agents. Comb. Chem. High Throughput Screen. 2016, 19, 537–553. [Google Scholar] [CrossRef] [PubMed]
- Nweze, J.A.; Mbaoji, F.N.; Li, Y.-M.; Yang, L.-Y.; Huang, S.-S.; Chigor, V.N.; Eze, E.A.; Pan, L.-X.; Zhang, T.; Yang, D.-F. Potentials of marine natural products against malaria, leishmaniasis, and trypanosomiasis parasites: A review of recent articles. Infect. Dis. Poverty 2021, 10, 1–19. [Google Scholar] [CrossRef] [PubMed]
- Pandey, A.K.; Singh, P. The genus Artemisia: A 2012–2017 literature review on chemical composition, antimicrobial, insecticidal and antioxidant activities of essential oils. Medicines 2017, 4, 68. [Google Scholar] [CrossRef] [PubMed]
- Fampa, P.; Florencio, M.; Santana, R.C.; Rosa, D.; Soares, D.C.; Guedes, H.L.D.M.; da Silva, A.C.; Chaves, D.S.A.; Pinto-Da-Silva, L.H. Anti-Leishmania effects of volatile oils and their isolates. Rev. Bras. Farmacogn. 2021, 1–18. [Google Scholar] [CrossRef]
- de Morais, M.C.; de Souza, J.V.; da Silva Maia Bezerra Filho, C.; Dolabella, S.S.; de Sousa, D.P. Trypanocidal essential oils: A review. Molecules 2020, 25, 4568. [Google Scholar] [CrossRef]
- Peixoto, J.F.; Ramos, Y.J.; Moreira, D.D.L.; Alves, C.R.; Gonçalves-Oliveira, L.F. Potential of Piper spp. as a source of new compounds for the leishmaniases treatment. Parasitol. Res. 2021, 120, 2731–2747. [Google Scholar] [CrossRef]
- Kamte, S.L.N.; Ranjbarian, F.; Cianfaglione, K.; Sut, S.; Dall’Acqua, S.; Bruno, M.; Afshar, F.H.; Iannarelli, R.; Benelli, G.; Cappellacci, L.; et al. Identification of highly effective antitrypanosomal compounds in essential oils from the Apiaceae family. Ecotoxicol. Environ. Saf. 2018, 156, 154–165. [Google Scholar] [CrossRef] [PubMed]
- Nibret, E.; Wink, M. Trypanocidal and antileukaemic effects of the essential oils of Hagenia abyssinica, Leonotis ocymifolia, Moringa stenopetala, and their main individual constituents. Phytomedicine 2010, 17, 911–920. [Google Scholar] [CrossRef] [PubMed]
- Greve, H.L.; Kaiser, M.; Brun, R.; Schmidt, T.J. Terpenoids from the oleo-gum-resin of Boswellia serrata and their antiplasmodial effects in vitro. Planta Med. 2017, 83, 1214–1226. [Google Scholar] [CrossRef] [PubMed]
- Mendiola, J.; Pino, J.A.; Fernández-Calienes, A.; Mendoza, D.; Herrera, P. Chemical composition and in vitro antiplasmodial activity of essential oils of leaves and flowers of Alpinia zerumbet grown in Cuba. PharmacologyOnline 2015, 2, 1–5. [Google Scholar]
- Monzote, L.; Scull, R.; Cos, P.; Setzer, W.N. Essential oil from Piper aduncum: Chemical analysis, antimicrobial assessment, and literature review. Medicines 2017, 4, 49. [Google Scholar] [CrossRef]
- Gutiérrez, Y.; Montes, R.; Scull, R.; Sánchez, A.; Cos, P.; Monzote, L.; Setzer, W.N. Chemodiversity associated with cytotoxicity and antimicrobial activity of Piper aduncum var. ossanum. Chem. Biodivers. 2016, 13, 1715–1719. [Google Scholar] [CrossRef]
- Esperandim, V.R.; Ferreira, D.D.S.; Rezende, K.C.S.; Magalhães, L.G.; Souza, J.M.; Pauletti, P.M.; Januário, A.H.; Laurentz, R.D.S.D.; Bastos, J.K.; Símaro, G.V.; et al. In vitro antiparasitic activity and chemical composition of the essential oil obtained from the fruits of Piper cubeba. Planta Med. 2013, 79, 1653–1655. [Google Scholar] [CrossRef]
- Kpoviessi, S.; Bero, J.; Agbani, P.; Gbaguidi, F.; Kpadonou-Kpoviessi, B.; Sinsin, B.; Accrombessi, G.; Frédérich, M.; Moudachirou, M.; Quetin-Leclercq, J. Chemical composition, cytotoxicity and in vitro antitrypanosomal and antiplasmodial activity of the essential oils of four Cymbopogon species from Benin. J. Ethnopharmacol. 2014, 151, 652–659. [Google Scholar] [CrossRef]
- Regalado, E.L.; Fernández, M.D.; Pino, J.A.; Mendiola, J.; Echemendia, O.A. Chemical composition and biological properties of the leaf essential oil of Tagetes lucida Cav. from Cuba. J. Essent. Oil Res. 2011, 23, 63–67. [Google Scholar] [CrossRef]
- Costa, E.V.; Dutra, L.; Nogueira, P.C.D.L.; Moraes, V.R.D.S.; Salvador, M.; Ribeiro, L.H.G.; Gadelha, F.R. Essential oil from the leaves of Annona vepretorum: Chemical composition and bioactivity. Nat. Prod. Commun. 2012, 7, 265–266. [Google Scholar] [CrossRef]
- Dias, C.N.; Rodrigues, K.A.F.; Carvalho, F.A.A.; Carneiro, S.M.P.; Maia, J.G.S.; Andrade, E.H.A.; Moraes, D.F.C. Molluscicidal and leishmanicidal activity of the leaf essential oil of Syzygium cumini (L.) SKEELS from Brazil. Chem. Biodivers. 2013, 10, 1133–1141. [Google Scholar] [CrossRef] [PubMed]
- Da Silva, T.B.; Menezes, L.; Sampaio, M.F.C.; Meira, C.S.; Guimarães, E.T.; Soares, M.; Prata, A.P.D.N.; Nogueira, P.C.D.L.; Costa, E. Chemical composition and anti-Trypanosoma cruzi activity of essential oils obtained from leaves of Xylopia frutescens and X. laevigata (Annonaceae). Nat. Prod. Commun. 2013, 8, 403–406. [Google Scholar] [CrossRef] [PubMed]
- Monzote, L.; Hill, G.M.; Cuellar, A.; Scull, R.; Setzer, W.N. Chemical composition and anti-proliferative properties of Bursera graveolens essential oil. Nat. Prod. Commun. 2012, 7, 1531–1534. [Google Scholar] [CrossRef] [PubMed]
- Rodríguez, E.J.; Saucedo-Hernández, Y.; Heyden, Y.V.; Simó-Alfonso, E.F.; Ramis-Ramos, G.; Lerma-García, M.J.; Monteagudo, U.; Bravo, L.; Medinilla, M.; De Armas, Y.; et al. Chemical analysis and antioxidant activity of the essential oils of three Piperaceae species growing in the central region of Cuba. Nat. Prod. Commun. 2013, 8, 1325–1328. [Google Scholar] [CrossRef] [PubMed]
- Tchoumbougnang, F.; Amvam Zollo, P.H.; Dagne, E.; Mekonnen, Y. In vivo antimalarial activity of essential oils from Cymbopogon citratus and Ocimum gratissimum on mice infected with Plasmodium berghei. Planta Med. 2005, 71, 20–23. [Google Scholar] [CrossRef]
- Monzote, L.; García, M.; Scull, R.; Cuellar, A.; Setzer, W.N. Antileishmanial activity of the essential oil from Bixa orellana. Phytother. Res. 2013, 28, 753–758. [Google Scholar] [CrossRef]
- Pereira, P.S.; Maia, A.J.; Duarte, A.E.; Oliveira-Tintino, C.D.M.; Tintino, S.R.; Barros, L.M.; Vega-Gomez, M.C.; Rolón, M.; Coronel, C.; Coutinho, H.D.; et al. Cytotoxic and anti-kinetoplastid potential of the essential oil of Alpinia speciosa K. Schum. Food Chem. Toxicol. 2018, 119, 387–391. [Google Scholar] [CrossRef]
- Alcoba, A.E.T.; De Melo, D.C.; De Andrade, P.M.; Dias, H.J.; Pagotti, M.C.; Magalhães, L.G.; Júnior, W.G.F.; Crotti, A.; Miranda, M.L.D. Chemical composition and in vitro antileishmanial and cytotoxic activities of the essential oils of Ocotea dispersa (Nees) Mez and Ocotea odorifera (Vell) Rohwer (Lauraceae). Nat. Prod. Res. 2017, 32, 2865–2868. [Google Scholar] [CrossRef]
- Tariku, Y.; Hymete, A.; Hailu, A.; Rohloff, J. In vitro evaluation of antileishmanial activity and toxicity of essential oils of Artemisia absinthium and Echinops kebericho. Chem. Biodivers. 2011, 8, 614–623. [Google Scholar] [CrossRef]
- Bouyahya, A.; Et-Touys, A.; Bakri, Y.; Talbaui, A.; Fellah, H.; Abrini, J.; Dakka, N. Chemical composition of Mentha pulegium and Rosmarinus officinalis essential oils and their antileishmanial, antibacterial and antioxidant activities. Microb. Pathog. 2017, 111, 41–49. [Google Scholar] [CrossRef]
- Sainz, P.; Andrés, M.F.; Díaz, M.; Bailén, M.; Rocha, J.N.; Martínez-Díaz, R.A.; González-Coloma, A. Chemical composition and biological activities of Artemisia pedemontana subsp. assoana essential oils and hydrolate. Biomolecules 2019, 9, 558. [Google Scholar] [CrossRef] [PubMed]
- Tabanca, N.; Demirci, B.; Baser, K.H.C.; Aytac, Z.; Ekici, M.; Khan, S.I.; Jacob, M.R.; Wedge, D.E. Chemical composition and antifungal activity of Salvia macrochlamys and Salvia recognita essential oils. J. Agric. Food Chem. 2006, 54, 6593–6597. [Google Scholar] [CrossRef] [PubMed]
- Moreira, R.R.D.; Dos Santos, A.G.; Carvalho, F.A.; Perego, C.H.; Crevelin, E.J.; Crotti, A.E.M.; Cogo, J.; Cardoso, M.L.C.; Nakamura, C.V. Antileishmanial activity of Melampodium divaricatum and Casearia sylvestris essential oils on Leishmania amazonensis. Rev. Inst. Med. Trop. 2019, 61, e33. [Google Scholar] [CrossRef] [PubMed]
- Sampaio, M.G.V.; Dos Santos, C.R.B.; Vandesmet, L.C.S.; Dos Santos, B.S.; Santos, I.B.D.S.; Correia, M.T.D.S.; Martins, A.L.D.B.; Da Silva, L.C.N.; Menezes, I.R.D.A.; Gomez, M.C.V.; et al. Chemical composition, antioxidant and antiprotozoal activity of Eugenia gracillima Kiaersk. leaves essential oil. Nat. Prod. Res. 2019, 35, 1914–1918. [Google Scholar] [CrossRef]
- Siqueira, C.A.T.; Serain, A.F.; Pascoal, A.C.R.F.; Andreazza, N.L.; De Lourenço, C.C.; Góis Ruiz, A.L.T.; De carvalho, J.E.; De Souza, A.C.O.; Tonini Mesquita, J.; Tempone, A.G.; et al. Bioactivity and chemical composition of the essential oil from the leaves of Guatteria australis A.St.-Hil. Nat. Prod. Res. 2015, 29, 1966–1969. [Google Scholar] [CrossRef]
- Barros, L.M.; Duarte, A.E.; Morais-Braga, M.F.B.; Waczuk, E.P.; Vega, C.; Leite, N.F.; De Menezes, I.R.A.; Coutinho, H.D.M.; Rocha, J.B.T.; Kamdem, J.P. Chemical characterization and trypanocidal, leishmanicidal and cytotoxicity potential of Lantana camara L. (Verbenaceae) essential oil. Molecules 2016, 21, 209. [Google Scholar] [CrossRef]
- Capello, T.M.; A Martins, E.G.; de Farias, C.F.; Figueiredo, C.R.; Matsuo, A.L.; Felipe Passero, L.D.; Oliveira-Silva, D.; Sartorelli, P.; Henrique Lago, J.G. Chemical composition and in vitro cytotoxic and antileishmanial activities of extract and essential oil from leaves of Piper cernuum. Nat. Prod. Commun. 2015, 10, 285–288. [Google Scholar] [CrossRef]
- Carmo, D.F.M.D.; Amaral, A.C.F.; Machado, G.M.C.; Leon, L.; Silva, J.R.D.A. Chemical and biological analyses of the essential oils and main constituents of Piper species. Molecules 2012, 17, 1819–1829. [Google Scholar] [CrossRef]
- Moreira, R.R.D.; Martins, G.Z.; Varandas, R.; Cogo, J.; Perego, C.H.; Roncoli, G.; Sousa, M.D.C.R.; Nakamura, C.V.; Salgueiro, L.; Cavaleiro, C. Composition and leishmanicidal activity of the essential oil of Vernonia polyanthes Less (Asteraceae). Nat. Prod. Res. 2017, 31, 2905–2908. [Google Scholar] [CrossRef]
- Giongo, J.L.; Vaucher, R.A.; Da Silva, A.S.; Oliveira, C.B.; de Mattos, C.B.; Baldissera, M.D.; Sagrillo, M.R.; Monteiro, S.G.; Custódio, D.L.; de Matos, M.S.; et al. Trypanocidal activity of the compounds present in Aniba canelilla oil against Trypanosoma evansi and its effects on viability of lymphocytes. Microb. Pathog. 2017, 103, 13–18. [Google Scholar] [CrossRef]
- De Souza, L.I.O.; Bezzera-Silva, P.C.; do Amaral Ferraz Navarro, D.M.; da Silva, A.G.; dos Santos Correia, M.T.; da Silva, M.V.; de Figueiredo, R.C.B.Q. The chemical composition and trypanocidal activity of volatile oils from Brazilian Caatinga plants. Biomed. Pharmacother. 2017, 96, 1055–1064. [Google Scholar] [CrossRef] [PubMed]
- Wangchuk, P.; Keller, P.; Pyne, S.; Taweechotipatr, M.; Kamchonwongpaisan, S. GC/GC-MS analysis, isolation and identification of bioactive essential oil components from the Bhutanese medicinal plant, Pleurospermum amabile. Nat. Prod. Commun. 2013, 8, 1305–1308. [Google Scholar] [CrossRef] [PubMed]
- Andrade, M.; Azevedo, C.D.S.; Motta, F.N.; dos Santos, M.L.; Silva, C.L.; De Santana, J.M.; Bastos, I.M.D. Essential oils: In vitro activity against Leishmania amazonensis, cytotoxicity and chemical composition. BMC Complement. Altern. Med. 2016, 16, 1–8. [Google Scholar] [CrossRef]
- Monzote, L.; García, M.; Montalvo, A.M.; Scull, R.; Miranda, M. Chemistry, cytotoxicity and antileishmanial activity of the essential oil from Piper auritum. Mem. Inst. Oswaldo Cruz. 2010, 105, 168–173. [Google Scholar] [CrossRef] [PubMed]
- Berthold, M.R.; Cebron, N.; Dill, F.; Di Fatta, G.; Gabriel, T.R.; Georg, F.; Meinl, T.; Ohl, P.; Sieb, C.; Wiswedel, B. KNIME: The Konstanz Information Miner. ACM SIGKDD Explor. Newsl. 2006, 11, 58–61. [Google Scholar]
- Berthold, M.R.; Cebron, N.; Dill, F.; Gabriel, T.R.; Kötter, T.; Meinl, T.; Ohl, P.; Thiel, K.; Wiswedel, B. KNIME-the Konstanz information miner: Version 2.0 and beyond. AcM SIGKDD Explor. Newsl. 2009, 11, 26–31. [Google Scholar] [CrossRef]
- Gramatica, P. Principles of QSAR Modeling. Int. J. Quant. Struct. Relatsh. 2020, 5, 61–97. [Google Scholar] [CrossRef]
- Roy, K.; Kar, S.; Das, R.N. Understanding the Basics of QSAR for Applications in Pharmaceutical Sciences and Risk Assessment, 1st ed.; Academic Press: San Diego, CA, USA, 2015; ISBN 9780128016336. [Google Scholar]
- Kar, S.; Roy, K.; Leszczynski, J. Applicability aomain: A step toward confident predictions and decidability for QSAR Modeling. Methods Mol. Biol. 2018, 1800, 141–169. [Google Scholar] [CrossRef]
- Scotti, M.; Speck-Planche, A.; Tavares, J.; Sobral da Silva, M.; Cordeiro, N.D.; Scotti, L. Virtual screening of alkaloids from Apocynaceae with potential antitrypanosomal activity. Curr. Bioinform. 2015, 10, 509–519. [Google Scholar] [CrossRef]



| Classification of EOs | Training | Average | ||||
| 1 | 2 | 3 | 4 | 5 | ||
| True positive rate | 0.90 | 0.95 | 0.99 | 0.90 | 0.80 | 0.91 |
| True negative rate | 0.70 | 0.65 | 0.68 | 0.70 | 0.75 | 0.71 |
| Accuracy | 0.81 | 0.80 | 0.83 | 0.81 | 0.78 | 0.81 |
| Classification of EOs | Test | Average | ||||
| 1 | 2 | 3 | 4 | 5 | ||
| True positive rate | 0.60 | 0.60 | 0.95 | 0.60 | 0.99 | 0.75 |
| True negative rate | 0.75 | 0.75 | 0.70 | 0.95 | 0.80 | 0.80 |
| Accuracy | 0.67 | 0.67 | 0.83 | 0.77 | 0.90 | 0.78 |
| Model | Specificity | Sensitivity | Accuracy | PPV | NPV |
|---|---|---|---|---|---|
| RF | 0.83 | 0.65 | 0.71 | 0.75 | 0.70 |
| Classification of EOs | Average | |
|---|---|---|
| SOM (Unsupervised) | RF (Supervised) | |
| Active | 0.85 | 0.75 |
| Not reported | 0.83 | 0.70 |
| Accuracy | 0.84 | 0.71 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Barros de Menezes, R.P.; Scotti, L.; Scotti, M.T.; García, J.; González, R.; Monzote, L.; Setzer, W.N. Machine Learning Analysis of Essential Oils from Cuban Plants: Potential Activity against Protozoa Parasites. Molecules 2022, 27, 1366. https://doi.org/10.3390/molecules27041366
Barros de Menezes RP, Scotti L, Scotti MT, García J, González R, Monzote L, Setzer WN. Machine Learning Analysis of Essential Oils from Cuban Plants: Potential Activity against Protozoa Parasites. Molecules. 2022; 27(4):1366. https://doi.org/10.3390/molecules27041366
Chicago/Turabian StyleBarros de Menezes, Renata Priscila, Luciana Scotti, Marcus Tullius Scotti, Jesús García, Rosalia González, Lianet Monzote, and William N. Setzer. 2022. "Machine Learning Analysis of Essential Oils from Cuban Plants: Potential Activity against Protozoa Parasites" Molecules 27, no. 4: 1366. https://doi.org/10.3390/molecules27041366
APA StyleBarros de Menezes, R. P., Scotti, L., Scotti, M. T., García, J., González, R., Monzote, L., & Setzer, W. N. (2022). Machine Learning Analysis of Essential Oils from Cuban Plants: Potential Activity against Protozoa Parasites. Molecules, 27(4), 1366. https://doi.org/10.3390/molecules27041366