
Genistein—Opportunities Related to an Interesting Molecule of Natural Origin

 ,
,  , , and
, , and 
Abstract
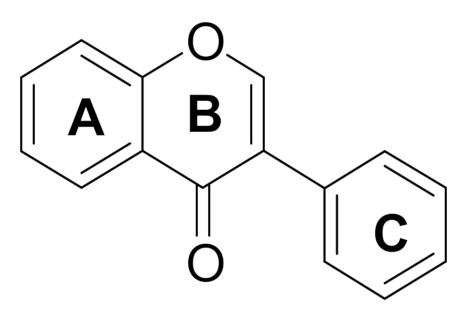
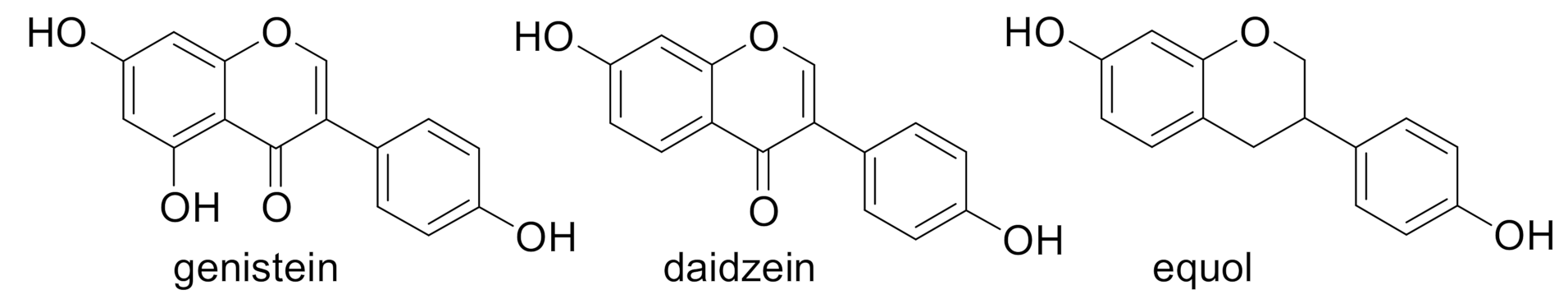
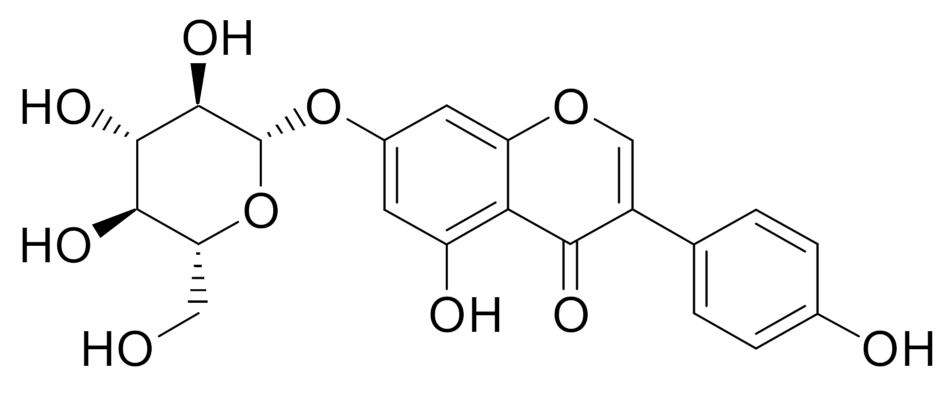
1. Introduction
2. Fermentation as a Way to Increase Genistein Content
3. Genistein as a Biologically Active Plant Compound
3.1. Psoriasis
3.2. Cataracts
3.3. Cystic Fibrosis
3.4. Non-Alcoholic Fatty Liver Disease
3.5. Type 2 Diabetes
4. Limitations
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Conflicts of Interest
References
- Zagórska-Dziok, M.; Kleczkowska, P.; Olędzka, E.; Figat, R.; Sobczak, M. Poly(chitosan-ester-ether-urethane) Hydrogels as Highly Controlled Genistein Release Systems. Int. J. Mol. Sci. 2021, 22, 3339. [Google Scholar] [CrossRef]
- Hou, W.; Liu, C.; Xia, J.; Niu, H.; Li, S. Rapid screening and purification of potential inhibitors from Medicago sativa by ultrafiltration-liquid chromatography combined with stepwise flow rate counter-current chromatography. Phytochem. Anal. 2021, 32, 382–394. [Google Scholar] [CrossRef] [PubMed]
- Tirdiľová, I.; Vollmannová, A.; Siekel, P.; Zetochová, E.; Čéryová, S.; Trebichalský, P. Selected legumes as a source of valuable substances in human nutrition. J. Food Nutr. Res. 2020, 59, 193–201. [Google Scholar]
- Neacsu, M.; Vaughan, N.J.; Perri, V.; Duncan, G.J.; Walker, R.; Coleman, M.; Russell, W.R. Nutritional and chemical profiling of UK-grown potato bean (Apios americana Medik) reveal its potential for diet biodiversification and revalorisation. J. Food Compos. Anal. 2021, 98, 103821. [Google Scholar] [CrossRef]
- Tuli, H.S.; Tuorkey, M.J.; Thakral, F.; Sak, K.; Kumar, M.; Sharma, A.K.; Sharma, U.; Jain, A.; Aggarwal, V.; Bishayee, A. Molecular Mechanisms of Action of Genistein in Cancer: Recent Advances. Front. Pharmacol. 2019, 10, 1336. [Google Scholar] [CrossRef]
- Naveen, J.; Baskaran, R.; Baskaran, V. Profiling of bioactives and in vitro evaluation of antioxidant and antidiabetic property of polyphenols of marine algae Padina tetrastromatica. Algal Res. 2021, 55, 102250. [Google Scholar] [CrossRef]
- Dixon, R.A.; Ferreira, D. Genistein. Phytochemistry 2002, 60, 205–211. [Google Scholar] [CrossRef]
- Thangavel, P.; Puga-Olguín, A.; Rodríguez-Landa, J.F.; Zepeda, R.C. Genistein as Potential Therapeutic Candidate for Menopausal Symptoms and Other Related Diseases. Molecules 2019, 24, 3892. [Google Scholar] [CrossRef]
- Matthies, A.; Blaut, M.; Braune, A. Isolation of a human intestinal bacterium capable of daidzein and genistein conversion. Appl. Env. Microbiol. 2009, 75, 1740–1744. [Google Scholar] [CrossRef]
- Iino, C.; Shimoyama, T.; Iino, K.; Yokoyama, Y.; Chinda, D.; Sakuraba, H.; Fukuda, S.; Nakaji, S. Daidzein Intake Is Associated with Equol Producing Status through an Increase in the Intestinal Bacteria Responsible for Equol Production. Nutrients 2019, 11, 433. [Google Scholar] [CrossRef]
- Liggins, J.; Bluck, L.J.; Runswick, S.; Atkinson, C.; Coward, W.A.; Bingham, S.A. Daidzein and genistein contents of vegetables. Br. J. Nutr. 2000, 84, 717–725. [Google Scholar] [CrossRef]
- Zhou, L.; Xiao, X.; Zhang, Q.; Zheng, J.; Li, M.; Deng, M. A Possible Mechanism: Genistein Improves Metabolism and Induces White Fat Browning Through Modulating Hypothalamic Expression of Ucn3, Depp, and Stc1. Front Endocrinol. 2019, 10, 478. [Google Scholar] [CrossRef] [PubMed]
- Cercato, L.M.; Oliveira, J.P.; Souza, M.T.S.; Andrade, N.; Martel, F.; Camargo, E.A. Effect of flavonoids in preclinical models of experimental obesity. PharmaNutrition 2021, 16, 100260. [Google Scholar] [CrossRef]
- Abron, J.D.; Singh, N.P.; Price, R.L.; Nagarkatti, M.; Nagarkatti, P.S.; Singh, U.P. Genistein induces macrophage polarization and systemic cytokine to ameliorate experimental colitis. PLoS ONE 2018, 13, e0199631. [Google Scholar]
- Skolmowska, D.; Głąbska, D.; Guzek, D.; Lech, G. Association between Dietary Isoflavone Intake and Ulcerative Colitis Symptoms in Polish Caucasian Individuals. Nutrients 2019, 11, 1936. [Google Scholar] [CrossRef]
- Ren, B.; Liu, Y.; Zhang, Y.; Cai, Y.; Gong, X.; Chang, Y.; Xu, L.; Zheng, J. Genistein: A Dual Inhibitor of Both Amyloid β and Human Islet Amylin Peptides. ACS Chem. Neurosci. 2018, 9, 1215–1224. [Google Scholar] [CrossRef]
- Gorzkiewicz, J.; Bartosz, G.; Sadowska-Bartosz, I. The Potential Effects of Phytoestrogens: The Role in Neuroprotection. Molecules 2021, 26, 2954. [Google Scholar] [CrossRef]
- Jang, C.H.; Oh, J.; Lim, J.S.; Kim, H.J.; Kim, J.-S. Fermented Soy Products: Beneficial Potential in Neurodegenerative Diseases. Foods 2021, 10, 636. [Google Scholar] [CrossRef]
- Han, S.; Wu, H.; Li, W.; Gao, P. Protective effects of genistein in homocysteine-induced endothelial cell inflammatory injury. Mol. Cell. Biochem. 2015, 403, 43–49. [Google Scholar] [CrossRef]
- Amerizadeh, A.; Asgary, S.; Vaseghi, G.; Farajzadegan, Z. Effect of Genistein Intake on Some Cardiovascular Risk Factors: An Updated Systematic Review and Meta-analysis. Curr. Probl. Cardiol. 2021, 100902. [Google Scholar] [CrossRef]
- Palanisamy, N.; Venkataraman, A.C. Beneficial effect of genistein on lowering blood pressure and kidney toxicity in fructose-fed hypertensive rats. Br. J. Nutr. 2013, 109, 1806–1812. [Google Scholar] [CrossRef] [PubMed]
- Jha, A.K.; Gairola, S.; Kundu, S.; Doye, P.; Syed, A.M.; Ram, C.; Murty, U.S.; Naidu, V.G.M.; Sahu, B.D. Toll-like receptor 4: An attractive therapeutic target for acute kidney injury. Life Sci. 2021, 271, 119155. [Google Scholar] [CrossRef] [PubMed]
- Yang, L.; Chen, Y.; Liu, Y.; Xing, Y.; Miao, C.; Zhao, Y.; Chang, X.; Zhang, Q. The Role of Oxidative Stress and Natural Antioxidants in Ovarian Aging. Front. Pharmacol. 2021, 11, 2364. [Google Scholar] [CrossRef] [PubMed]
- Čoma, M.; Lachová, V.; Mitrengová, P.; Gál, P. Molecular Changes Underlying Genistein Treatment of Wound Healing: A Review. Curr. Issues Mol. Biol. 2021, 43, 127–141. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Kong, D.; Bao, B.; Ahmad, A.; Sarkar, F.H. Induction of Cancer Cell Death by Isoflavone: The Role of Multiple Signaling Pathways. Nutrients 2011, 3, 877–896. [Google Scholar] [CrossRef]
- Ziaei, S.; Halaby, R. Dietary Isoflavones and Breast Cancer Risk. Medicines 2017, 4, 18. [Google Scholar] [CrossRef]
- Shah, R.D.; Tang, Z.-Z.; Chen, G.; Huang, S.; Ferguson, J.F. Soy food intake associates with changes in the metabolome and reduced blood pressure in a gut microbiota dependent manner. Nutr. Metab. Cardiovasc. Dis. 2020, 30, 1500–1511. [Google Scholar] [CrossRef]
- Shreiner, A.B.; Kao, J.Y.; Young, V.B. The gut microbiome in health and in disease. Curr. Opin. Gastroenterol. 2015, 31, 69–75. [Google Scholar] [CrossRef]
- World Health Organization. Global Report on Psoriasis; World Health Organization: Geneva, Switzerland, 2016. [Google Scholar]
- Younossi, Z.M.; Koenig, A.B.; Abdelatif, D.; Fazel, Y.; Henry, L.; Wymer, M. Global epidemiology of nonalcoholic fatty liver disease—Meta-analytic assessment of prevalence, incidence, and outcomes. Hepatology 2016, 64, 73–84. [Google Scholar] [CrossRef]
- Hashemi, H.; Pakzad, R.; Yekta, A.; Aghamirsalim, M.; Pakbin, M.; Ramin, S.; Khabazkhoob, M. Global and regional prevalence of age-related cataract: A comprehensive systematic review and meta-analysis. Eye 2020, 34, 1357–1370. [Google Scholar] [CrossRef]
- Scotet, V.; L’Hostis, C.; Férec, C. The Changing Epidemiology of Cystic Fibrosis: Incidence, Survival and Impact of the CFTR Gene Discovery. Genes 2020, 11, 589. [Google Scholar] [CrossRef] [PubMed]
- Sanders, D.B.; Fink, A.K. Background and Epidemiology. Pediatr. Clin. N. Am. 2016, 63, 567–584. [Google Scholar] [CrossRef] [PubMed]
- Lin, X.; Xu, Y.; Pan, X.; Xu, J.; Ding, Y.; Sun, X.; Song, X.; Ren, Y.; Shan, P.-F. Global, regional, and national burden and trend of diabetes in 195 countries and territories: An analysis from 1990 to 2025. Sci. Rep. 2020, 10, 14790. [Google Scholar] [CrossRef] [PubMed]
- Kim, I.-S.; Kim, C.-H.; Yang, W.-S. Physiologically Active Molecules and Functional Properties of Soybeans in Human Health—A Current Perspective. Int. J. Mol. Sci. 2021, 22, 4054. [Google Scholar] [CrossRef]
- Kim, I.-S. Current Perspectives on the Beneficial Effects of Soybean Isoflavones and Their Metabolites for Humans. Antioxidants 2021, 10, 1064. [Google Scholar] [CrossRef] [PubMed]
- Mureșan, L.; Clapa, D.; Borsai, O.; Rusu, T.; Wang, T.T.Y.; Park, J.B. Potential Impacts of Soil Tillage System on Isoflavone Concentration of Soybean as Functional Food Ingredients. Land 2020, 9, 386. [Google Scholar] [CrossRef]
- Nile, S.H.; Nile, A.; Oh, J.-W.; Kai, G. Soybean processing waste: Potential antioxidant, cytotoxic and enzyme inhibitory activities. Food Biosci. 2020, 38, 100778. [Google Scholar] [CrossRef]
- Gupta, S.; Chen, W.N. A metabolomics approach to evaluate post-fermentation enhancement of daidzein and genistein in a green okara extract. J. Sci. Food Agric. 2021, 101, 5124–5131. [Google Scholar] [CrossRef]
- Xiao, Y.; Huang, Y.; Chen, Y.; Fan, Z.; Chen, R.; He, C.; Li, Z.; Wang, Y. Effects of Solid-State Fermentation with Eurotium cristatum YL-1 on the Nutritional Value, Total Phenolics, Isoflavones, Antioxidant Activity, and Volatile Organic Compounds of Black Soybeans. Agronomy 2021, 11, 1029. [Google Scholar] [CrossRef]
- Khosravi, A.; Razavi, S.H. Therapeutic effects of polyphenols in fermented soybean and black soybean products. J. Funct. Foods 2021, 81, 104467. [Google Scholar] [CrossRef]
- Kim, I.-S.; Hwang, C.-W.; Yang, W.-S.; Kim, C.-H. Current Perspectives on the Physiological Activities of Fermented Soybean-Derived Cheonggukjang. Int. J. Mol. Sci. 2021, 22, 5746. [Google Scholar] [CrossRef] [PubMed]
- Zheng, Y.; Li, L.; Jin, Z.; An, P.; Yang, S.-T.; Fei, Y.; Liu, G. Characterization of fermented soymilk by Schleiferilactobacillus harbinensis M1, based on the whole-genome sequence and corresponding phenotypes. LWT 2021, 144, 111237. [Google Scholar] [CrossRef]
- Feng, J.-Y.; Thakur, K.; Ni, Z.-J.; Zhu, Y.-Y.; Hu, F.; Zhang, J.-G.; Wei, Z.-J. Effects of okara and vitamin B2 bioenrichment on the functional properties and in vitro digestion of fermented soy milk. Food Res. Int. 2021, 145, 110419. [Google Scholar] [CrossRef] [PubMed]
- Gao, Y.X.; Xu, B.; Fan, H.R.; Zhang, M.R.; Zhang, L.J.; Lu, C.; Zhang, N.N.; Fan, B.; Wang, F.Z.; Li, S. 1H NMR-based chemometric metabolomics characterization of soymilk fermented by Bacillus subtilis BSNK-5. Food Res. Int. 2020, 138, 109686. [Google Scholar] [CrossRef]
- Undhad, T.; Hati, S.; Makwana, S. Significance of storage study on ACE inhibitory, antioxidative, antimicrobial activities, and biotransformation of isoflavones of functional fermented soy-based beverage. J. Food Processing Preserv. 2021, 45, e15062. [Google Scholar] [CrossRef]
- Rendon, A.; Schäkel, K. Psoriasis Pathogenesis and Treatment. Int. J. Mol. Sci. 2019, 20, 1475. [Google Scholar] [CrossRef]
- Kim, W.B.; Jerome, D.; Yeung, J. Diagnosis and management of psoriasis. Can Fam. Physician 2017, 63, 278–285. [Google Scholar]
- Prignano, F.; Ricceri, F.; Pescitelli, L.; Lotti, T. Itch in psoriasis: Epidemiology, clinical aspects and treatment options. Clin. Cosmet. Investig. Derm. 2009, 2, 9–13. [Google Scholar] [CrossRef]
- Horn, E.J.; Fox, K.M.; Patel, V.; Chiou, C.F.; Dann, F.; Lebwohl, M. Are patients with psoriasis undertreated? Results of National Psoriasis Foundation survey. J. Am. Acad. Derm. 2007, 57, 957–962. [Google Scholar] [CrossRef]
- Fan, Y.; Yang, R.; He, J.; Gou, X.; Zhuang, Z.; Guo, X.; Li, X. Explore the rule and mechanism of psoriasis herbal treatment based on data mining and network pharmacology. Asian Toxicol. Res. 2021, 3, 6–10. [Google Scholar]
- Parisi, R.; Iskandar, I.Y.K.; Kontopantelis, E.; Augustin, M.; Griffiths, C.E.M.; Ashcroft, D.M.; Global Psoriasis, A. National, regional, and worldwide epidemiology of psoriasis: Systematic analysis and modelling study. BMJ 2020, 369, m1590. [Google Scholar] [CrossRef] [PubMed]
- Nestle, F.O.; Kaplan, D.H.; Barker, J. Psoriasis. N. Engl. J. Med. 2009, 361, 496–509. [Google Scholar] [CrossRef] [PubMed]
- Ogawa, E.; Sato, Y.; Minagawa, A.; Okuyama, R. Pathogenesis of psoriasis and development of treatment. J. Derm. 2018, 45, 264–272. [Google Scholar] [CrossRef] [PubMed]
- Lowes, M.A.; Suárez-Fariñas, M.; Krueger, J.G. Immunology of psoriasis. Annu Rev. Immunol. 2014, 32, 227–255. [Google Scholar] [CrossRef]
- Baliwag, J.; Barnes, D.H.; Johnston, A. Cytokines in psoriasis. Cytokine 2015, 73, 342–350. [Google Scholar] [CrossRef]
- Pal, S.; Sen, S.; Nath, I.; Kumar, A.; Biswas, U. Psoriasis, An inflammatory condition associated with oxidative stress. Asian J. Med. Sci. 2021, 12, 24–30. [Google Scholar] [CrossRef]
- Pleńkowska, J.; Gabig-Cimińska, M.; Mozolewski, P. Oxidative Stress as an Important Contributor to the Pathogenesis of Psoriasis. Int. J. Mol. Sci. 2020, 21, 6206. [Google Scholar] [CrossRef]
- Kanda, N.; Hoashi, T.; Saeki, H. Nutrition and Psoriasis. Int. J. Mol. Sci. 2020, 21, 5405. [Google Scholar] [CrossRef]
- Pazyar, N.; Yaghoobi, R. Soybean: A potential antipsoriasis agent. Jundishapur J. Nat. Pharm. Prod. 2015, 10, e20924. [Google Scholar] [CrossRef]
- Kładna, A.; Berczyński, P.; Kruk, I.; Piechowska, T.; Aboul-Enein, H.Y. Studies on the antioxidant properties of some phytoestrogens. Luminescence 2016, 31, 1201–1206. [Google Scholar] [CrossRef]
- Smolińska, E.; Moskot, M.; Jakóbkiewicz-Banecka, J.; Węgrzyn, G.; Banecki, B.; Szczerkowska-Dobosz, A.; Purzycka-Bohdan, D.; Gabig-Cimińska, M. Molecular action of isoflavone genistein in the human epithelial cell line HaCaT. PLoS ONE 2018, 13, e0192297. [Google Scholar] [CrossRef] [PubMed]
- Smolińska, E.; Węgrzyn, G.; Gabig-Cimińska, M. Genistein modulates gene activity in psoriatic patients. Acta Biochim. Pol. 2019, 66, 101–110. [Google Scholar] [CrossRef] [PubMed]
- Li, H.-J.; Wu, N.-L.; Lee, G.-A.; Hung, C.-F. The Therapeutic Potential and Molecular Mechanism of Isoflavone Extract against Psoriasis. Sci. Rep. 2018, 8, 6335. [Google Scholar] [CrossRef] [PubMed]
- Wang, A.; Wei, J.; Lu, C.; Chen, H.; Zhong, X.; Lu, Y.; Li, L.; Huang, H.; Dai, Z.; Han, L. Genistein suppresses psoriasis-related inflammation through a STAT3–NF-κB-dependent mechanism in keratinocytes. Int. Immunopharmacol. 2019, 69, 270–278. [Google Scholar] [CrossRef]
- Gupta, V.B.; Rajagopala, M.; Ravishankar, B. Etiopathogenesis of cataract: An appraisal. Indian J. Ophthalmol 2014, 62, 103–110. [Google Scholar] [CrossRef]
- Nartey, A. The Pathophysiology of Cataract and Major Interventions to Retarding Its Progression: A Mini Review. Adv. Ophthalmol. Vis. Syst. 2017, 6, 00178. [Google Scholar] [CrossRef][Green Version]
- Tewari, D.; Samoilă, O.; Gocan, D.; Mocan, A.; Moldovan, C.; Devkota, H.P.; Atanasov, A.G.; Zengin, G.; Echeverría, J.; Vodnar, D.; et al. Medicinal Plants and Natural Products Used in Cataract Management. Front. Pharmacol. 2019, 10, 466. [Google Scholar] [CrossRef]
- Cao, J.; Wang, T.; Wang, M. Investigation of the anti-cataractogenic mechanisms of curcumin through in vivo and in vitro studies. BMC Ophthalmol. 2018, 18, 48. [Google Scholar] [CrossRef]
- Soni, P.; Choudhary, R.; Bodakhe, S.H. Effects of a novel isoflavonoid from the stem bark of Alstonia scholaris against fructose-induced experimental cataract. J. Integr. Med. 2019, 17, 374–382. [Google Scholar] [CrossRef]
- Pollreisz, A.; Schmidt-Erfurth, U. Diabetic Cataract—Pathogenesis, Epidemiology and Treatment. J. Ophthalmol. 2010, 2010, 608751. [Google Scholar] [CrossRef]
- Hashim, Z.; Zarina, S. Osmotic stress induced oxidative damage: Possible mechanism of cataract formation in diabetes. J. Diabetes Complicat. 2012, 26, 275–279. [Google Scholar] [CrossRef] [PubMed]
- Lam, D.; Rao, S.K.; Ratra, V.; Liu, Y.; Mitchell, P.; King, J.; Tassignon, M.J.; Jonas, J.; Pang, C.P.; Chang, D.F. Cataract. Nat. Rev. Dis. Primers 2015, 1, 15014. [Google Scholar] [CrossRef] [PubMed]
- Davis, G. The Evolution of Cataract Surgery. Mo. Med. 2016, 113, 58–62. [Google Scholar] [PubMed]
- Chan, E.; Mahroo, O.A.; Spalton, D.J. Complications of cataract surgery. Clin. Exp. Optom. 2010, 93, 379–389. [Google Scholar] [CrossRef]
- Ma, Y.; Gao, W.; Wu, K.; Bao, Y. Flavonoid intake and the risk of age-related cataract in China’s Heilongjiang Province. Food Nutr. Res. 2015, 59, 29564. [Google Scholar] [CrossRef]
- Patil, K.K.; Meshram, R.J.; Dhole, N.A.; Gacche, R.N. Role of dietary flavonoids in amelioration of sugar induced cataractogenesis. Arch. Biochem. Biophys. 2016, 593, 1–11. [Google Scholar] [CrossRef]
- Kim, Y.S.; Kim, N.H.; Jung, D.H.; Jang, D.S.; Lee, Y.M.; Kim, J.M.; Kim, J.S. Genistein inhibits aldose reductase activity and high glucose-induced TGF-β2 expression in human lens epithelial cells. Eur. J. Pharmacol. 2008, 594, 18–25. [Google Scholar] [CrossRef]
- Huang, R.; Shi, F.; Lei, T.; Song, Y.; Hughes, C.L.; Liu, G. Effect of the Isoflavone Genistein Against Galactose-Induced Cataracts in Rats. Exp. Biol. Med. 2007, 232, 118–125. [Google Scholar]
- Floyd, K.A.; Stella, D.R.; Wang, C.-C.; Laurentz, S.; McCabe, G.P.; Srivastava, O.P.; Barnes, S. Genistein and genistein-containing dietary supplements accelerate the early stages of cataractogenesis in the male ICR/f rat. Exp. Eye Res. 2011, 92, 120–127. [Google Scholar] [CrossRef]
- Sinha, R.; Shekhar, H.; Sharma, N.; Titiyal, J.S.; Vajpayee, R.B. Posterior capsular opacification: A review. Indian J. Ophthalmol. 2013, 61, 371–376. [Google Scholar] [CrossRef]
- Awasthi, N.; Guo, S.; Wagner, B.J. Posterior Capsular Opacification: A Problem Reduced but Not Yet Eradicated. Arch. Ophthalmol. 2009, 127, 555–562. [Google Scholar] [CrossRef] [PubMed]
- Zhang, W.; Li, X.; Ye, T.; Chen, F.; Sun, X.; Kong, J.; Yang, X.; Pan, W.; Li, S. Design, characterization, and in vitro cellular inhibition and uptake of optimized genistein-loaded NLC for the prevention of posterior capsular opacification using response surface methodology. Int. J. Pharm. 2013, 454, 354–366. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.L.; Zhang, W.J.; Li, X.D.; Yang, N.; Pan, W.S.; Kong, J.; Zhang, J.S. Sustained-release genistein from nanostructured lipid carrier suppresses human lens epithelial cell growth. Int. J. Ophthalmol. 2016, 9, 643–649. [Google Scholar] [PubMed]
- Zhang, W.; Liu, J.; Zhang, Q.; Li, X.; Yu, S.; Yang, X.; Kong, J.; Pan, W. Enhanced cellular uptake and anti-proliferating effect of chitosan hydrochlorides modified genistein loaded NLC on human lens epithelial cells. Int. J. Pharm. 2014, 471, 118–126. [Google Scholar] [CrossRef]
- Yan, T.; Ma, Z.; Liu, J.; Yin, N.; Lei, S.; Zhang, X.; Li, X.; Zhang, Y.; Kong, J. Thermoresponsive GenisteinNLC-dexamethasone-moxifloxacin multi drug delivery system in lens capsule bag to prevent complications after cataract surgery. Sci. Rep. 2021, 11, 181. [Google Scholar] [CrossRef]
- Ratjen, F.; Bell, S.C.; Rowe, S.M.; Goss, C.H.; Quittner, A.L.; Bush, A. Cystic fibrosis. Nat. Rev. Dis. Primers 2015, 1, 15010. [Google Scholar] [CrossRef]
- Martin, C.; Hamard, C.; Kanaan, R.; Boussaud, V.; Grenet, D.; Abély, M.; Hubert, D.; Munck, A.; Lemonnier, L.; Burgel, P.-R. Causes of death in French cystic fibrosis patients: The need for improvement in transplantation referral strategies! J. Cyst. Fibros. 2016, 15, 204–212. [Google Scholar] [CrossRef]
- Bell, S.C.; Mall, M.A.; Gutierrez, H.; Macek, M.; Madge, S.; Davies, J.C.; Burgel, P.-R.; Tullis, E.; Castaños, C.; Castellani, C.; et al. The future of cystic fibrosis care: A global perspective. Lancet Respir. Med. 2020, 8, 65–124. [Google Scholar] [CrossRef]
- Castellani, C.; Duff, A.J.A.; Bell, S.C.; Heijerman, H.G.M.; Munck, A.; Ratjen, F.; Sermet-Gaudelus, I.; Southern, K.W.; Barben, J.; Flume, P.A.; et al. ECFS best practice guidelines: The 2018 revision. J. Cyst. Fibros. 2018, 17, 153–178. [Google Scholar] [CrossRef]
- Węgrzyn, G.; Pierzynowska, K.; Podlacha, M.; Brokowska, J.; Gaffke, L.; Mantej, J.; Cyske, Z.; Rintz, E.; Osiadły, M.; Bartkowski, M.; et al. Molecular mechanisms of genistein action in the light of therapies for genetic and immunological diseases. Postepy Biochem. 2018, 64, 262–276. [Google Scholar] [CrossRef]
- Arora, K.; Yarlagadda, S.; Zhang, W.; Moon, C.; Bouquet, E.; Srinivasan, S.; Li, C.; Stokes, D.C.; Naren, A.P. Personalized medicine in cystic fibrosis: Genistein supplementation as a treatment option for patients with a rare S1045Y-CFTR mutation. Am. J. Physiol.—Lung Cell. Mol. Physiol. 2016, 311, L364–L374. [Google Scholar] [CrossRef] [PubMed]
- Southern, K.W.; Patel, S.; Sinha, I.P.; Nevitt, S.J. Correctors (specific therapies for class II CFTR mutations) for cystic fibrosis. Cochrane Database Syst. Rev. 2018, 8, Cd010966. [Google Scholar] [CrossRef] [PubMed]
- Lord, R.; Fairbourn, N.; Mylavarapu, C.; Dbeis, A.; Bowman, T.; Chandrashekar, A.; Banayat, T.; Hodges, C.A.; Al-Nakkash, L. Consuming Genistein Improves Survival Rates in the Absence of Laxative in ΔF508-CF Female Mice. Nutrients 2018, 10, 1418. [Google Scholar] [CrossRef] [PubMed]
- Dekkers, J.F.; Van Mourik, P.; Vonk, A.M.; Kruisselbrink, E.; Berkers, G.; de Winter-de Groot, K.M.; Janssens, H.M.; Bronsveld, I.; van der Ent, C.K.; de Jonge, H.R.; et al. Potentiator synergy in rectal organoids carrying S1251N, G551D, or F508del CFTR mutations. J. Cyst. Fibros. 2016, 15, 568–578. [Google Scholar] [CrossRef] [PubMed]
- Berkers, G.; van der Meer, R.; van Mourik, P.; Vonk, A.M.; Kruisselbrink, E.; Suen, S.W.F.; Heijerman, H.G.M.; Majoor, C.J.; Koppelman, G.H.; Roukema, J.; et al. Clinical effects of the three CFTR potentiator treatments curcumin, genistein and ivacaftor in patients with the CFTR-S1251N gating mutation. J. Cyst. Fibros. 2020, 19, 955–961. [Google Scholar] [CrossRef]
- Yang, Z.; Kulkarni, K.; Zhu, W.; Hu, M. Bioavailability and pharmacokinetics of genistein: Mechanistic studies on its ADME. Anticancer Agents Med. Chem. 2012, 12, 1264–1280. [Google Scholar] [CrossRef]
- Chandrasekharan, A. Pharmacokinetics of Dietary Isoflavones. J. Steroids Horm. Sci. 2013, s12, 1–8. [Google Scholar] [CrossRef]
- Lee, Y.H.; Cho, Y.; Lee, B.W.; Park, C.Y.; Lee, D.H.; Cha, B.S.; Rhee, E.J. Nonalcoholic Fatty Liver Disease in Diabetes. Part I: Epidemiology and Diagnosis. Diabetes Metab. J. 2019, 43, 31–45. [Google Scholar] [CrossRef]
- Smith, B.W.; Adams, L.A. Non-alcoholic fatty liver disease. Crit. Rev. Clin. Lab. Sci. 2011, 48, 97–113. [Google Scholar] [CrossRef]
- Milić, S.; Stimac, D. Nonalcoholic fatty liver disease/steatohepatitis: Epidemiology, pathogenesis, clinical presentation and treatment. Dig Dis. 2012, 30, 158–162. [Google Scholar] [CrossRef]
- Benedict, M.; Zhang, X. Non-alcoholic fatty liver disease: An expanded review. World J. Hepatol. 2017, 9, 715–732. [Google Scholar] [CrossRef] [PubMed]
- Hsu, W.-F.; Sheen, L.-Y.; Lin, H.-J.; Chang, H.-H. A Review of Western and Traditional Chinese Medical Approaches to Managing Nonalcoholic Fatty Liver Disease. Evid Based Complement Altern. Med. 2016, 2016, 6491420. [Google Scholar] [CrossRef] [PubMed]
- Sayiner, M.; Koenig, A.; Henry, L.; Younossi, Z.M. Epidemiology of Nonalcoholic Fatty Liver Disease and Nonalcoholic Steatohepatitis in the United States and the Rest of the World. Clin. Liver Dis. 2016, 20, 205–214. [Google Scholar] [CrossRef] [PubMed]
- Wong, S.-W.; Chan, W.-K. Epidemiology of non-alcoholic fatty liver disease in Asia. Indian J. Gastroenterol. 2020, 39, 1–8. [Google Scholar] [CrossRef]
- Abdelmalek, M.F.; Suzuki, A.; Guy, C.; Unalp-Arida, A.; Colvin, R.; Johnson, R.J.; Diehl, A.M.; Nonalcoholic Steatohepatitis Clinical Research Network. Increased fructose consumption is associated with fibrosis severity in patients with nonalcoholic fatty liver disease. Hepatology 2010, 51, 1961–1971. [Google Scholar] [CrossRef]
- Kalia, H.S.; Gaglio, P.J. The Prevalence and Pathobiology of Nonalcoholic Fatty Liver Disease in Patients of Different Races or Ethnicities. Clin. Liver Dis. 2016, 20, 215–224. [Google Scholar] [CrossRef]
- Fan, J.-G.; Kim, S.-U.; Wong, V.W.-S. New trends on obesity and NAFLD in Asia. J. Hepatol. 2017, 67, 862–873. [Google Scholar] [CrossRef]
- Mohanty, S.R.; Troy, T.N.; Huo, D.; O’Brien, B.L.; Jensen, D.M.; Hart, J. Influence of ethnicity on histological differences in non-alcoholic fatty liver disease. J. Hepatol. 2009, 50, 797–804. [Google Scholar] [CrossRef]
- Nasr, P.; Iredahl, F.; Dahlström, N.; Rådholm, K.; Henriksson, P.; Cedersund, G.; Dahlqvist Leinhard, O.; Ebbers, T.; Alfredsson, J.; Carlhäll, C.-J.; et al. Evaluating the prevalence and severity of NAFLD in primary care: The EPSONIP study protocol. BMC Gastroenterol. 2021, 21, 180. [Google Scholar] [CrossRef]
- Leoni, S.; Tovoli, F.; Napoli, L.; Serio, I.; Ferri, S.; Bolondi, L. Current guidelines for the management of non-alcoholic fatty liver disease: A systematic review with comparative analysis. World J. Gastroenterol. 2018, 24, 3361–3373. [Google Scholar] [CrossRef]
- Xin, X.; Chen, C.; Hu, Y.Y.; Feng, Q. Protective effect of genistein on nonalcoholic fatty liver disease (NAFLD). BioMed. Pharm. 2019, 117, 109047. [Google Scholar] [CrossRef] [PubMed]
- Yin, Y.; Liu, H.; Zheng, Z.; Lu, R.; Jiang, Z. Genistein can ameliorate hepatic inflammatory reaction in nonalcoholic steatohepatitis rats. Biomed. Pharmacother. 2019, 111, 1290–1296. [Google Scholar] [CrossRef] [PubMed]
- Gao, X.; Liu, S.; Ding, C.; Miao, Y.; Gao, Z.; Li, M.; Fan, W.; Tang, Z.; Mhlambi, N.H.; Yan, L.; et al. Comparative effects of genistein and bisphenol A on non-alcoholic fatty liver disease in laying hens. Environ. Pollut. 2021, 288, 117795. [Google Scholar] [CrossRef] [PubMed]
- Zamani-Garmsiri, F.; Hashemnia, S.M.R.; Shabani, M.; Bagherieh, M.; Emamgholipour, S.; Meshkani, R. Combination of metformin and genistein alleviates non-alcoholic fatty liver disease in high-fat diet-fed mice. J. Nutr. Biochem. 2021, 87, 108505. [Google Scholar] [CrossRef] [PubMed]
- Farruggio, S.; Cocomazzi, G.; Marotta, P.; Romito, R.; Surico, D.; Calamita, G.; Bellan, M.; Pirisi, M.; Grossini, E. Genistein and 17β-Estradiol Protect Hepatocytes from Fatty Degeneration by Mechanisms Involving Mitochondria, Inflammasome and Kinases Activation. Cell Physiol. Biochem. 2020, 54, 401–416. [Google Scholar]
- Seidemann, L.; Krüger, A.; Kegel-Hübner, V.; Seehofer, D.; Damm, G. Influence of Genistein on Hepatic Lipid Metabolism in an In Vitro Model of Hepatic Steatosis. Molecules 2021, 26, 1156. [Google Scholar] [CrossRef]
- Pummoung, S.; Werawatganon, D.; Chayanupatkul, M.; Klaikeaw, N.; Siriviriyakul, P. Genistein Modulated Lipid Metabolism, Hepatic PPARγ, and Adiponectin Expression in Bilateral Ovariectomized Rats with Nonalcoholic Steatohepatitis (NASH). Antioxidants 2020, 10, 24. [Google Scholar] [CrossRef]
- Witayavanitkul, N.; Werawatganon, D.; Chayanupatkul, M.; Klaikeaw, N.; Siriviriyakul, P. Genistein and exercise treatment reduced NASH related HDAC3, IL-13 and MMP-12 expressions in ovariectomized rats fed with high fat high fructose diet. J. Tradit. Complementary Med. 2021, 11, 503–512. [Google Scholar] [CrossRef]
- Gan, M.; Shen, L.; Fan, Y.; Tan, Y.; Zheng, T.; Tang, G.; Niu, L.; Zhao, Y.; Chen, L.; Jiang, D.; et al. MicroRNA-451 and Genistein Ameliorate Nonalcoholic Steatohepatitis in Mice. Int. J. Mol. Sci. 2019, 20, 6084. [Google Scholar] [CrossRef]
- Larkin, T.; Price, W.E.; Astheimer, L. The Key Importance of Soy Isoflavone Bioavailability to Understanding Health Benefits. Crit. Rev. Food Sci. Nutr. 2008, 48, 538–552. [Google Scholar] [CrossRef]
- Zheng, X.; Lee, S.-K.; Chun, O.K. Soy Isoflavones and Osteoporotic Bone Loss: A Review with an Emphasis on Modulation of Bone Remodeling. J. Med. Food 2016, 19, 1–14. [Google Scholar] [CrossRef]
- Erdman, J.W. Soy Protein and Cardiovascular Disease. Circulation 2000, 102, 2555–2559. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Lv, J.; Yu, C.; Li, L.; Hu, Y.; Qin, L.-Q.; Dong, J.-Y. Dietary Soy Consumption and Cardiovascular Mortality among Chinese People with Type 2 Diabetes. Nutrients 2021, 13, 2513. [Google Scholar] [CrossRef] [PubMed]
- Kimura, T.; Egawa, S. Epidemiology of prostate cancer in Asian countries. Int. J. Urol. 2018, 25, 524–531. [Google Scholar] [CrossRef] [PubMed]
- Morey, B.N.; Gee, G.C.; von Ehrenstein, O.S.; Shariff-Marco, S.; Canchola, A.J.; Yang, J.; Allen, L.; Lee, S.S.J.; Bautista, R.; La Chica, T.; et al. Higher Breast Cancer Risk Among Immigrant Asian American Women Than Among US-Born Asian American Women. PRev. Chronic Dis. 2019, 16, 180221. [Google Scholar] [CrossRef]
- Ma, R.C.W.; Chan, J.C.N. Type 2 diabetes in East Asians: Similarities and differences with populations in Europe and the United States. Ann. N. Y. Acad. Sci. 2013, 1281, 64–91. [Google Scholar] [CrossRef]
- Li, H.; Khor, C.-C.; Fan, J.; Lv, J.; Yu, C.; Guo, Y.; Bian, Z.; Yang, L.; Millwood, I.Y.; Walters, R.G.; et al. Genetic risk, adherence to a healthy lifestyle, and type 2 diabetes risk among 550,000 Chinese adults: Results from 2 independent Asian cohorts. Am. J. Clin. Nutr. 2020, 111, 698–707. [Google Scholar] [CrossRef]
- Li, Y.; Teng, D.; Shi, X.; Qin, G.; Qin, Y.; Quan, H.; Shi, B.; Sun, H.; Ba, J.; Chen, B.; et al. Prevalence of diabetes recorded in mainland China using 2018 diagnostic criteria from the American Diabetes Association: National cross sectional study. BMJ 2020, 369, m997. [Google Scholar] [CrossRef]
- Yokokawa, H.; Fukuda, H.; Saita, M.; Goto, K.; Kaku, T.; Miyagami, T.; Takahashi, Y.; Hamada, C.; Hisaoka, T.; Naito, T. An association between visceral or subcutaneous fat accumulation and diabetes mellitus among Japanese subjects. Diabetol. Metab. Syndr. 2021, 13, 44. [Google Scholar] [CrossRef]
- Rhee, E.J. Diabetes in Asians. Endocrinol. Metab. 2015, 30, 263–269. [Google Scholar] [CrossRef]
- Tajdar, D.; Lühmann, D.; Fertmann, R.; Steinberg, T.; van den Bussche, H.; Scherer, M.; Schäfer, I. Low health literacy is associated with higher risk of type 2 diabetes: A cross-sectional study in Germany. BMC Public Health 2021, 21, 510. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.-H.; Lee, E.-S.; Yoo, J.; Kim, Y. Predicting risk of type 2 diabetes mellitus in Korean adults aged 40–69 by integrating clinical and genetic factors. Prim. Care Diabetes 2019, 13, 3–10. [Google Scholar] [CrossRef] [PubMed]
- Nogueira-de-Almeida, C.A.; Ferraz, I.S.; Ued, F.V.; Almeida, A.C.F.; Del Ciampo, L.A. Impact of soy consumption on human health: Integrative review. Braz. J. Food Technol. 2020, 23, e2019129. [Google Scholar] [CrossRef]
- Xiao, C.W. Health Effects of Soy Protein and Isoflavones in Humans. J. Nutr. 2008, 138, 1244S–1249S. [Google Scholar] [CrossRef] [PubMed]
- Villegas, R.; Shu, X.O.; Gao, Y.-T.; Yang, G.; Elasy, T.; Li, H.; Zheng, W. Vegetable but not fruit consumption reduces the risk of type 2 diabetes in Chinese women. J. Nutr. 2008, 138, 574–580. [Google Scholar] [CrossRef]
- Zamora-Ros, R.; Forouhi, N.G.; Sharp, S.J.; González, C.A.; Buijsse, B.; Guevara, M.; van der Schouw, Y.T.; Amiano, P.; Boeing, H.; Bredsdorff, L.; et al. The association between dietary flavonoid and lignan intakes and incident type 2 diabetes in European populations: The EPIC-InterAct study. Diabetes Care 2013, 36, 3961–3970. [Google Scholar] [CrossRef]
- Nanri, A.; Mizoue, T.; Takahashi, Y.; Kirii, K.; Inoue, M.; Noda, M.; Tsugane, S. Soy Product and Isoflavone Intakes Are Associated with a Lower Risk of Type 2 Diabetes in Overweight Japanese Women. J. Nutr. 2010, 140, 580–586. [Google Scholar] [CrossRef]
- Ko, K.-P.; Kim, C.-S.; Ahn, Y.; Park, S.-J.; Kim, Y.-J.; Park, J.K.; Lim, Y.-K.; Yoo, K.-Y.; Kim, S.S. Plasma isoflavone concentration is associated with decreased risk of type 2 diabetes in Korean women but not men: Results from the Korean Genome and Epidemiology Study. Diabetologia 2015, 58, 726–735. [Google Scholar] [CrossRef]
- Mayo, B.; Vázquez, L.; Flórez, A.B. Equol: A Bacterial Metabolite from The Daidzein Isoflavone and Its PresuMed. Beneficial Health Effects. Nutrients 2019, 11, 2231. [Google Scholar] [CrossRef]
- Atkinson, C.; Frankenfeld, C.L.; Lampe, J.W. Gut Bacterial Metabolism of the Soy Isoflavone Daidzein: Exploring the Relevance to Human Health. Exp. Biol. Med. 2005, 230, 155–170. [Google Scholar] [CrossRef]
- Haron, H.; Ismail, A.; Azlan, A.; Shahar, S.; Peng, L.S. Daidzein and genestein contents in tempeh and selected soy products. Food Chem. 2009, 115, 1350–1356. [Google Scholar] [CrossRef]
- Dong, H.-L.; Tang, X.-Y.; Deng, Y.-Y.; Zhong, Q.-W.; Wang, C.; Zhang, Z.-Q.; Chen, Y.-M. Urinary equol, but not daidzein and genistein, was inversely associated with the risk of type 2 diabetes in Chinese adults. Eur. J. Nutr. 2020, 59, 719–728. [Google Scholar] [CrossRef] [PubMed]
- Talaei, M.; Lee, B.L.; Ong, C.N.; van Dam, R.M.; Yuan, J.M.; Koh, W.P.; Pan, A. Urine phyto-oestrogen metabolites are not significantly associated with risk of type 2 diabetes: The Singapore Chinese health study. Br. J. Nutr. 2016, 115, 1607–1615. [Google Scholar] [CrossRef] [PubMed]
- Ye, Y.-B.; Chen, A.-L.; Lu, W.; Zhuo, S.-Y.; Liu, J.; Guan, J.-H.; Deng, W.-P.; Fang, S.; Li, Y.-B.; Chen, Y.-M. Daidzein and genistein fail to improve glycemic control and insulin sensitivity in Chinese women with impaired glucose regulation: A double-blind, randomized, placebo-controlled trial. Mol. Nutr. Food Res. 2015, 59, 240–249. [Google Scholar] [CrossRef] [PubMed]
- Squadrito, F.; Marini, H.; Bitto, A.; Altavilla, D.; Polito, F.; Adamo, E.B.; D’Anna, R.; Arcoraci, V.; Burnett, B.P.; Minutoli, L.; et al. Genistein in the Metabolic Syndrome: Results of a Randomized Clinical Trial. J. Clin. Endocrinol. Metab. 2013, 98, 3366–3374. [Google Scholar] [CrossRef]
- Ding, M.; Franke, A.; Rosner, B.; Giovannucci, E.; Van Dam, R.; Tworoger, S.; Hu, F.B.; Sun, Q. Urinary isoflavonoids and risk of type 2 diabetes: A prospective investigation in US women. Br. J. Nutr. 2015, 114, 1694–1701. [Google Scholar] [CrossRef][Green Version]
- Ding, M.; Pan, A.; Manson, J.E.; Willett, W.C.; Malik, V.; Rosner, B.; Giovannucci, E.; Hu, F.B.; Sun, Q. Consumption of soy foods and isoflavones and risk of type 2 diabetes: A pooled analysis of three US cohorts. Eur. J. Clin. Nutr. 2016, 70, 1381–1387. [Google Scholar] [CrossRef]
- Jayagopal, V.; Albertazzi, P.; Kilpatrick, E.S.; Howarth, E.M.; Jennings, P.E.; Hepburn, D.A.; Atkin, S.L. Beneficial Effects of Soy Phytoestrogen Intake in Postmenopausal Women With Type 2 Diabetes. Diabetes Care 2002, 25, 1709. [Google Scholar] [CrossRef]
- Zhou, L.; Xiao, X.; Zhang, Q.; Zheng, J.; Li, M.; Yu, M.; Wang, X.; Deng, M.; Zhai, X.; Li, R.; et al. Dietary Genistein Could Modulate Hypothalamic Circadian Entrainment, Reduce Body Weight, and Improve Glucose and Lipid Metabolism in Female Mice. Int. J. Endocrinol. 2019, 2019, 2163838. [Google Scholar] [CrossRef]
- Makena, W.; Hambolu, J.O.; Timbuak, J.A.; Umana, U.E.; Iliya, A.I.; Dibal, N.I. Mormodica charantia L. fruit and Genistein ameliorates type 2 diabetes in rats by preventing lipid accumulation, insulin resistance and enhancing beta cell function. J. Diabetes Metab. Disord. 2020, 19, 1303–1310. [Google Scholar] [CrossRef]
- Fu, Z.; Gilbert, E.R.; Pfeiffer, L.; Zhang, Y.; Fu, Y.; Liu, D. Genistein ameliorates hyperglycemia in a mouse model of nongenetic type 2 diabetes. Appl. Physiol. Nutr. Metab. 2012, 37, 480–488. [Google Scholar] [CrossRef] [PubMed]
- Zhang, P.; Li, T.; Wu, X.; Nice, E.C.; Huang, C.; Zhang, Y. Oxidative stress and diabetes: Antioxidative strategies. Front. Med. 2020, 14, 583–600. [Google Scholar] [CrossRef] [PubMed]
- López, P.; Sánchez, M.; Perez-Cruz, C.; Velázquez-Villegas, L.A.; Syeda, T.; Aguilar-López, M.; Rocha-Viggiano, A.K.; del Carmen Silva-Lucero, M.; Torre-Villalvazo, I.; Noriega, L.G.; et al. Long-Term Genistein Consumption Modifies Gut Microbiota, Improving Glucose Metabolism, Metabolic Endotoxemia, and Cognitive Function in Mice Fed a High-Fat Diet. Mol. Nutr. Food Res. 2018, 62, 1800313. [Google Scholar] [CrossRef] [PubMed]
- de Kort, S.; Keszthelyi, D.; Masclee, A.A.M. Leaky gut and diabetes mellitus: What is the link? Obes. Rev. 2011, 12, 449–458. [Google Scholar] [CrossRef] [PubMed]
- Zhou, L.; Xiao, X.; Zhang, Q.; Zheng, J.; Li, M.; Yu, M.; Wang, X.; Deng, M.; Zhai, X.; Li, R. Improved Glucose and Lipid Metabolism in the Early Life of Female Offspring by Maternal Dietary Genistein Is Associated With Alterations in the Gut Microbiota. Front. Endocrinol. 2018, 9, 516. [Google Scholar] [CrossRef]
- Valsecchi, A.E.; Franchi, S.; Panerai, A.E.; Rossi, A.; Sacerdote, P.; Colleoni, M. The soy isoflavone genistein reverses oxidative and inflammatory state, neuropathic pain, neurotrophic and vasculature deficits in diabetes mouse model. Eur. J. Pharmacol. 2011, 650, 694–702. [Google Scholar] [CrossRef]
- Nakajima, M.; Cooney, M.J.; Tu, A.H.; Chang, K.Y.; Cao, J.; Ando, A.; An, G.-J.; Melia, M.; de Juan, E., Jr. Normalization of Retinal Vascular Permeability in Experimental Diabetes with Genistein. Investig. Ophthalmol. Vis. Sci. 2001, 42, 2110–2114. [Google Scholar]
- Eo, H.; Lee, H.-J.; Lim, Y. Ameliorative effect of dietary genistein on diabetes induced hyper-inflammation and oxidative stress during early stage of wound healing in alloxan induced diabetic mice. Biochem. Biophys. Res. Commun. 2016, 478, 1021–1027. [Google Scholar] [CrossRef]
- Xiao, Y.; Ho, C.-T.; Chen, Y.; Wang, Y.; Wei, Z.; Dong, M.; Huang, Q. Synthesis, Characterization, and Evaluation of Genistein-Loaded Zein/Carboxymethyl Chitosan Nanoparticles with Improved Water Dispersibility, Enhanced Antioxidant Activity, and Controlled Release Property. Foods 2020, 9, 1604. [Google Scholar] [CrossRef]
- Messina, M. Soy and Health Update: Evaluation of the Clinical and Epidemiologic Literature. Nutrients 2016, 8, 754. [Google Scholar] [CrossRef]
- Morito, K.; Hirose, T.; Kinjo, J.; Hirakawa, T.; Okawa, M.; Nohara, T.; Ogawa, S.; Inoue, S.; Muramatsu, M.; Masamune, Y. Interaction of phytoestrogens with estrogen receptors alpha and beta. Biol. Pharm. Bull. 2001, 24, 351–356. [Google Scholar] [CrossRef] [PubMed]
- Szkudelska, K.; Nogowski, L. Genistein—A dietary compound inducing hormonal and metabolic changes. J. Steroid Biochem. Mol. Biol. 2007, 105, 37–45. [Google Scholar] [CrossRef] [PubMed]
- Messina, M. Soybean isoflavone exposure does not have feminizing effects on men: A critical examination of the clinical evidence. Fertil. Steril. 2010, 93, 2095–2104. [Google Scholar] [CrossRef]
- Messina, M.J.; Wood, C.E. Soy isoflavones, estrogen therapy, and breast cancer risk: Analysis and commentary. Nutr. J. 2008, 7, 17. [Google Scholar] [CrossRef]
- Travis, R.C.; Key, T.J. Oestrogen exposure and breast cancer risk. Breast Cancer Res. 2003, 5, 239–247. [Google Scholar] [CrossRef] [PubMed]
- Russo, J.; Russo, I.H. The role of estrogen in the initiation of breast cancer. J. Steroid Biochem. Mol. Biol. 2006, 102, 89–96. [Google Scholar] [CrossRef]
- Setchell, K.D. Soy isoflavones-benefits and risks from nature’s selective estrogen receptor modulators (SERMs). J. Am. Coll Nutr. 2001, 20, 354S–362S. [Google Scholar] [CrossRef]
- Messina, M. Impact of Soy Foods on the Development of Breast Cancer and the Prognosis of Breast Cancer Patients. KomplementMed 2016, 23, 75–80. [Google Scholar] [CrossRef]
- Hilakivi-Clarke, L.; Andrade, J.E.; Helferich, W. Is Soy Consumption Good or Bad for the Breast? J. Nutr. 2010, 140, 2326S–2334S. [Google Scholar] [CrossRef]
- Maskarinec, G.; Ju, D.; Morimoto, Y.; Franke, A.A.; Stanczyk, F.Z. Soy Food Intake and Biomarkers of Breast Cancer Risk: Possible Difference in Asian Women? Nutr. Cancer 2017, 69, 146–153. [Google Scholar] [CrossRef]
- Divi, R.L.; Chang, H.C.; Doerge, D.R. Anti-Thyroid Isoflavones from Soybean: Isolation, Characterization, and Mechanisms of Action. Biochem. Pharmacol. 1997, 54, 1087–1096. [Google Scholar] [CrossRef]
- Chang, H.C.; Doerge, D.R. Dietary genistein inactivates rat thyroid peroxidase in vivo without an apparent hypothyroid effect. Toxicol. Appl. Pharm. 2000, 168, 244–252. [Google Scholar] [CrossRef] [PubMed]
- Messina, M.; Redmond, G. Effects of soy protein and soybean isoflavones on thyroid function in healthy adults and hypothyroid patients: A review of the relevant literature. Thyroid 2006, 16, 249–258. [Google Scholar] [CrossRef]
- Hüser, S.; Guth, S.; Joost, H.G.; Soukup, S.T.; Köhrle, J.; Kreienbrock, L.; Diel, P.; Lachenmeier, D.W.; Eisenbrand, G.; Vollmer, G.; et al. Effects of isoflavones on breast tissue and the thyroid hormone system in humans: A comprehensive safety evaluation. Arch. Toxicol. 2018, 92, 2703–2748. [Google Scholar] [CrossRef] [PubMed]
- Testa, I.; Salvatori, C.; Di Cara, G.; Latini, A.; Frati, F.; Troiani, S.; Principi, N.; Esposito, S. Soy-Based Infant Formula: Are Phyto-Oestrogens Still in Doubt? Front. Nutr. 2018, 5, 110. [Google Scholar] [CrossRef]
- Doerge, D.R.; Sheehan, D.M. Goitrogenic and estrogenic activity of soy isoflavones. Env. Health Perspect. 2002, 110, 349–353. [Google Scholar] [CrossRef]
- Conrad, S.C.; Chiu, H.; Silverman, B.L. Soy formula complicates management of congenital hypothyroidism. Arch. Dis. Child. 2004, 89, 37–40. [Google Scholar] [CrossRef]
- Soukup, S.T.; Helppi, J.; Müller, D.R.; Zierau, O.; Watzl, B.; Vollmer, G.; Diel, P.; Bub, A.; Kulling, S.E. Phase II metabolism of the soy isoflavones genistein and daidzein in humans, rats and mice: A cross-species and sex comparison. Arch. Toxicol. 2016, 90, 1335–1347. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Disease | Models | Period | Genistein Dosage and Administration | Plasma Concentration of Genistein | Main Results | Year Published | Reference |
|---|---|---|---|---|---|---|---|
| Cataracts | Cataract induced by galactose-rich diet in male Long–Evans rats | 4 weeks | 15 mg/kg body weight oral gavage | N/A | 1. Genistein treatment did not prevent cataract formation 2. Score and extent of cataracts↓ 3. Serum glucose↓ 4. Serum testosterone↑ | 2007 | [79] |
| Cataracts | Male hereditary cataractous ICR/f rat | 85 days | 0.018% and 0.2% in the diet (as an aglycone or β-glucoside) | 0.018% Genistein: 113.4 ± 63.2 nmol/L 0.20% Genistein: 2138.1 ± 720.8 nmol/L NovaSoy®200: 120.9 ± 107.3 nmol/L SPI: 72.5 ± 38.9 nmol/L | 1. Acceleration of early stages of cataractogenesis | 2011 | [80] |
| Cystic fibrosis | F508del-CF male and female mouse | 45 days | 600 mg genistein/kg in the diet | N/A | 1. Female survival rate↑ 2. Male body weight↑ 3. SGLT-1 expression↑ | 2018 | [94] |
| NAFLD | High-fat high-sucrose diet-fed SPF male Sprague-Dawley rats | 12 weeks | 0.1% and 0.2% in the diet | N/A | 1. NAFLD activity score↓ 2. Body weight↑, liver index↓ 3. Liver and serum TG↓, TC↓; LDL-C, HDL-C had no difference 4. ALT↓, AST↓ 5. Blood glucose did not change significantly; insulin↓, HOMA-IR↓ 6. Liver TNFα↓, serum endotoxin↓, TLR4↓ | 2019 | [113] |
| NAFLD | High-energy and low-protein diet-fed Hy-Line Brown laying hens | 90 days | 40 mg/kg, 200 mg/kg, 400 mg/kg in the diet | N/A | 1. Hepatic steatosis↓, NAFLD activity score↓ 2. ALT↓, AST↓ 3. Serum TG↓, TC↓, LDL-C↓, HDL-C↑ 4. Expression of AMPKα↑, CPT-1↑, PPARα↑ mRNA levels; ACC↓, FAS↓, SCD-1↓, SREBP-1↓, LPL↓ and Cd36↓ 5. Restored insulin sensitivity 6. mRNA and protein levels of NLRP3↓, caspase-1↓, IL-18↓, IL-1β↓ 7. Up-regulation of ERα in dose-dependent effect, no changes in ERβ expression | 2021 | [114] |
| NAFLD | High-fat diet-fed male C57BL/6 mice | 3 months | 0.2% in the diet (alone and in combination with metformin) | N/A | 1. Body and liver weight↓ 2. Blood glucose↓, plasma insulin↓, HOMA-IR↓; ameliorated glucose tolerance 3. ALT↓, AST↓, liver and plasma TG↓; reduced steatosis 4. Expression of FAS↓, CPT1↑, SREBP-1c↓ (combination of metformin and genistein) 5. NAFLD activity score↓ 6. Liver phospho-AMPK protein level↑, TNF-α↓, IL-1β↓, IL-6↓, NF-κB p65 protein level↓ 7. Hepatic expression level of PEPCK↓ and G6Pase↓; hepatic GSK-3β phosphorylation↑ | 2020 | [115] |
| NAFLD | High-fat high-fructose diet-fed ovariectomized female Sprague-Dawley rats | 5 weeks | 16 mg/kg body weight dissolved in 0.1% dimethyl sulfoxide oral gavage | N/A | 1. Body weight did not change significantly 2. Hepatic MMP-12 expression↓, level of HDAC3 expression↓, IL-13↓ 3. NASH activity score↓ | 2021 | [119] |
| NAFLD | High-fat high-fructose diet-fed ovariectomized female Sprague-Dawley rats | 8 weeks | 16 mg/kg body weight dissolved in 1 mL 0.1% dimethyl sulfoxide oral gavage | N/A | 1. Improved liver steatosis 2. Hepatic MDA level↓ 3. Hepatic TG↓, FFA↓ 4. PPARγ↓, adiponectin↑ | 2021 | [118] |
| NAFLD | High-fat diet-fed lipopolysaccharide-injected female ICR mice | 1 month | 100 mg/kg in the diet | N/A | 1. Liver weight index↓ 2. Hepatic TNFα↓, IL6↓, IL1β↓ 3. Expression of miR-451↑, Cab39↓ 4. NASH activity score↓ | 2019 | [120] |
| Diabetes | High-fat diet-fed C57BL/6J female mice | 8 weeks | 2 g/kg diet | N/A | 1. Body weight↓ 2. Blood glucose levels↓, serum insulin level did not change significantly, HOMA-IR↓ 3. Serum TG↓, LDL-C↓, FFA↓; TC and HDL-C did not change significantly 4. Up-regulated genes: Per1, c-Fos, Calm1, Gng5 5. Down-regulated genes: Grin1, Cacna1g, Kir3.1, Adcy4, Gucy1a2 | 2019 | [150] |
| Diabetes | High-fat diet-fed streptozotocin-injected female Wistar rats | 4 weeks | 10 mg/kg and 20 mg/kg in the diet | N/A | 1. Blood glucose↓, serum insulin level↑, HOMA-IR↓ 2. Cholesterol↓, triglyceride↓, LDL↓, VLDL↓, HDL↑ 3. Regeneration of β-cells | 2020 | [151] |
| Diabetes | High-fat diet-fed streptozotocin-injected male C57BL/6 mice | 8 weeks | 250 mg/kg diet | N/A | 1. Body weight, major organ weight did not change significantly 2. Fasting blood glucose↓ 3. TC, TG, HDL-C did not change significantly 4. β-cell mass improved | 2012 | [152] |
| Disease | Participants/ Type | Period | Genistein Dosage | Concentration of Genistein | Main Results | Year Published | Reference |
|---|---|---|---|---|---|---|---|
| Psoriasis | 34/randomized, double-blind, placebo-controlled trial | 56 days | 75 mg and 150 mg | N/A | 1. Up-regulated genes: IL1RN 2. Down-regulated genes: CCL4, NFKB1, STAT3, CXCL10, IL6, IL8 | 2019 | [63] |
| Cystic fibrosis | Study 1: 13 Study 3: 14/multicenter clinical trials | 8 weeks | Study 1. 3.3 and 5.0 mg/kg/day + curcumin in 3–4 doses Study 2. ivacaftor Study 3. 5.0 and 10.0 mg/kg/day + ivacaftor | Study 1. 3 µg/L (0.01 µM) Study 3. 14 µg/L (0.05 µM) | 1. Study 1, 3: no statistically significant clinical effect | 2020 | [96] |
| Diabetes | 120 postmenopausal women/randomized, double-blind, placebo-controlled trial | 1 year | 54 mg | 790 ± 95 nmol/L (serum) | 1. HOMA-IR↓, blood glucose↓, insulin↓ 2. TC↓, TG↓, HDL-C↑ 3. Serum adiponectin↑, visfatin↓, homocysteine↓ | 2013 | [146] |
| Diabetes | 165 women/randomized, double-blind, placebo-controlled trial | 24 weeks | 10 g of soy protein plus (i) no addition, (ii) 50 mg of daidzein, (iii) 50 mg of genistein | 16.6 mmol/24 h (urine) | 1. Glycemic control, insulin resistance-no significant improvement | 2014 | [145] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Garbiec, E.; Cielecka-Piontek, J.; Kowalówka, M.; Hołubiec, M.; Zalewski, P. Genistein—Opportunities Related to an Interesting Molecule of Natural Origin. Molecules 2022, 27, 815. https://doi.org/10.3390/molecules27030815
Garbiec E, Cielecka-Piontek J, Kowalówka M, Hołubiec M, Zalewski P. Genistein—Opportunities Related to an Interesting Molecule of Natural Origin. Molecules. 2022; 27(3):815. https://doi.org/10.3390/molecules27030815
Chicago/Turabian StyleGarbiec, Ewa, Judyta Cielecka-Piontek, Magdalena Kowalówka, Magdalena Hołubiec, and Przemysław Zalewski. 2022. "Genistein—Opportunities Related to an Interesting Molecule of Natural Origin" Molecules 27, no. 3: 815. https://doi.org/10.3390/molecules27030815
APA StyleGarbiec, E., Cielecka-Piontek, J., Kowalówka, M., Hołubiec, M., & Zalewski, P. (2022). Genistein—Opportunities Related to an Interesting Molecule of Natural Origin. Molecules, 27(3), 815. https://doi.org/10.3390/molecules27030815