
Biological Activity of an Epilobium angustifolium L. (Fireweed) Infusion after In Vitro Digestion


 , ,
, ,  , and
, and 
Abstract
:1. Introduction
2. Results
2.1. Chemical Characterization
2.2. Experiments on Animal Cell Lines
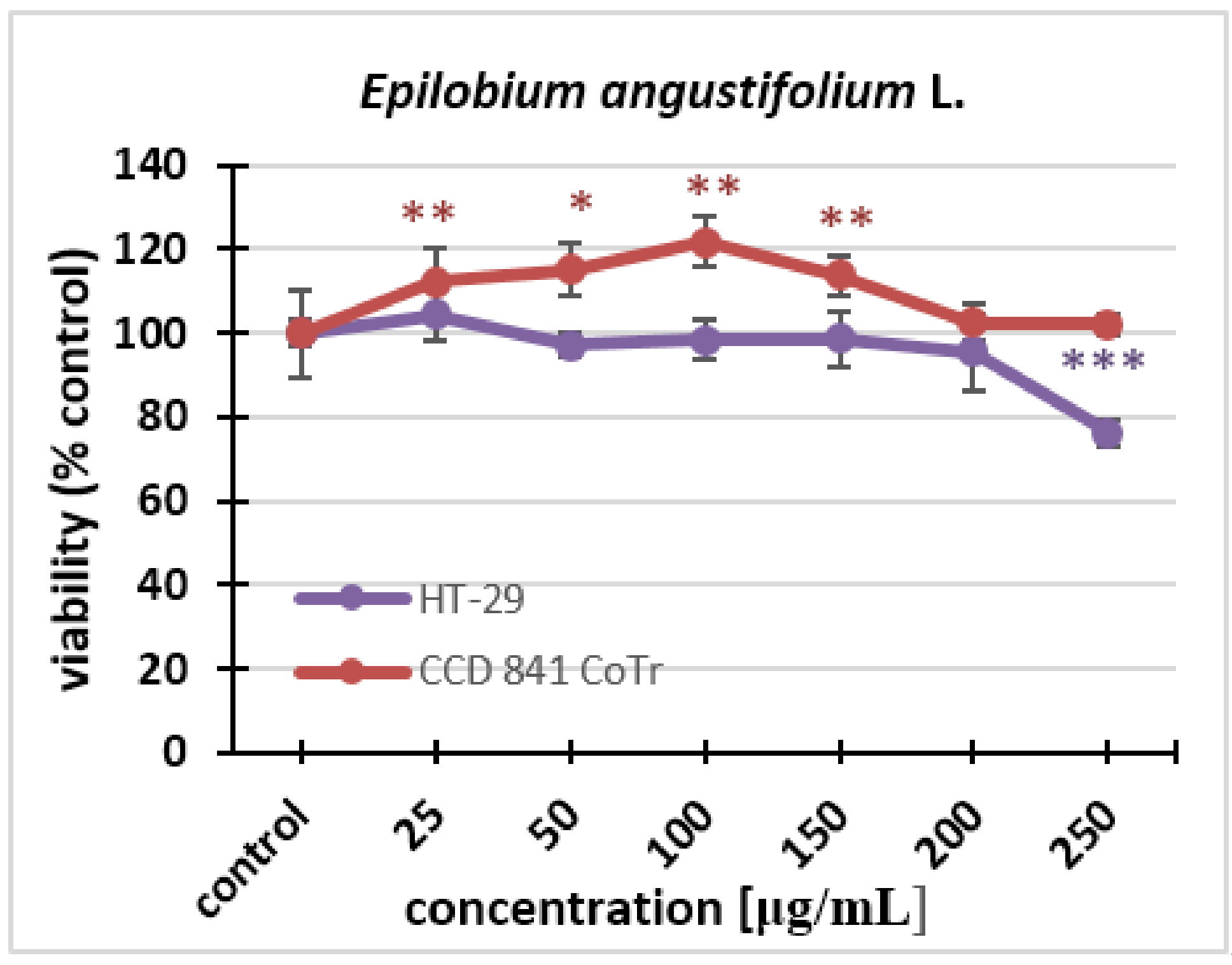
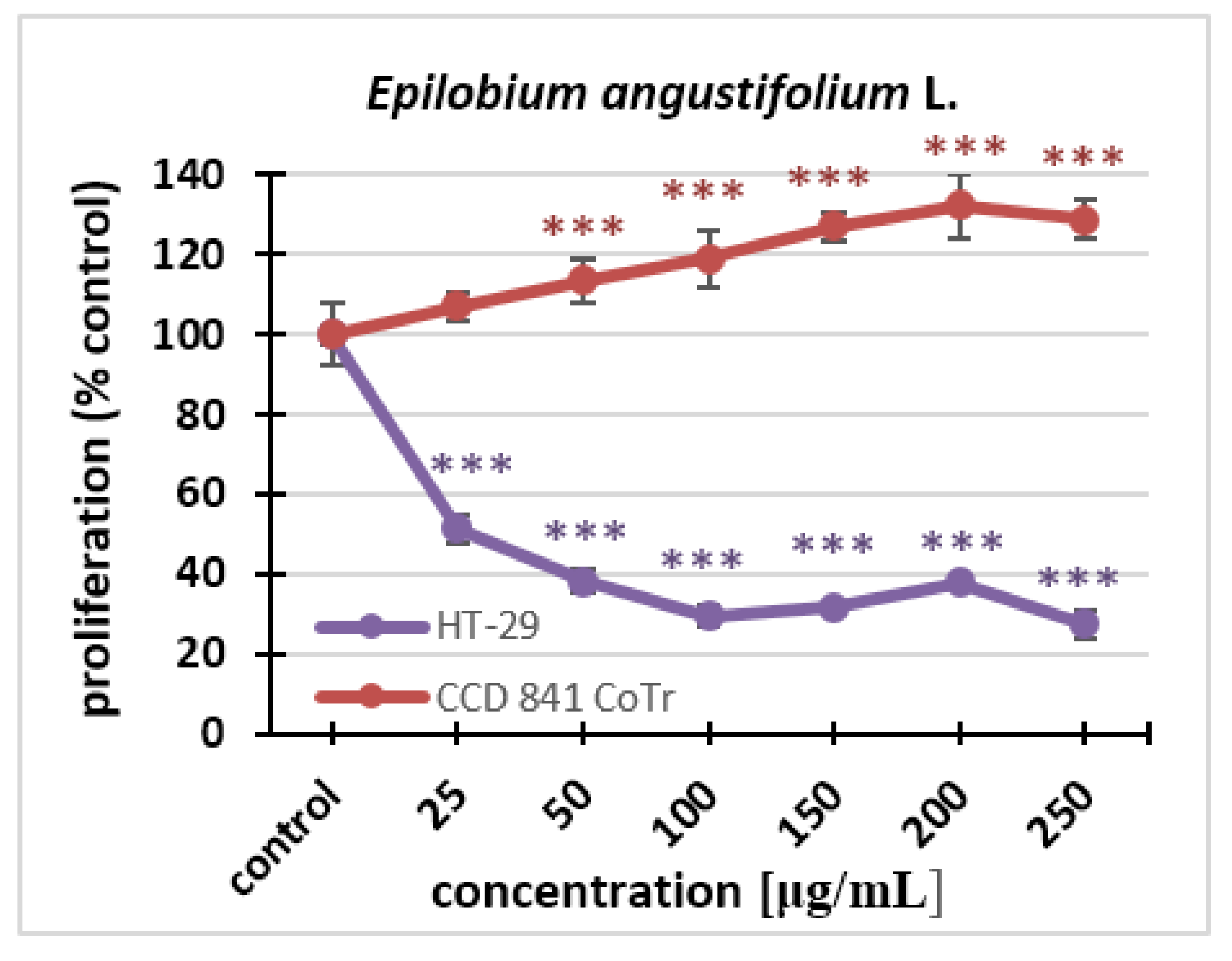
2.2.1. MTT Assay
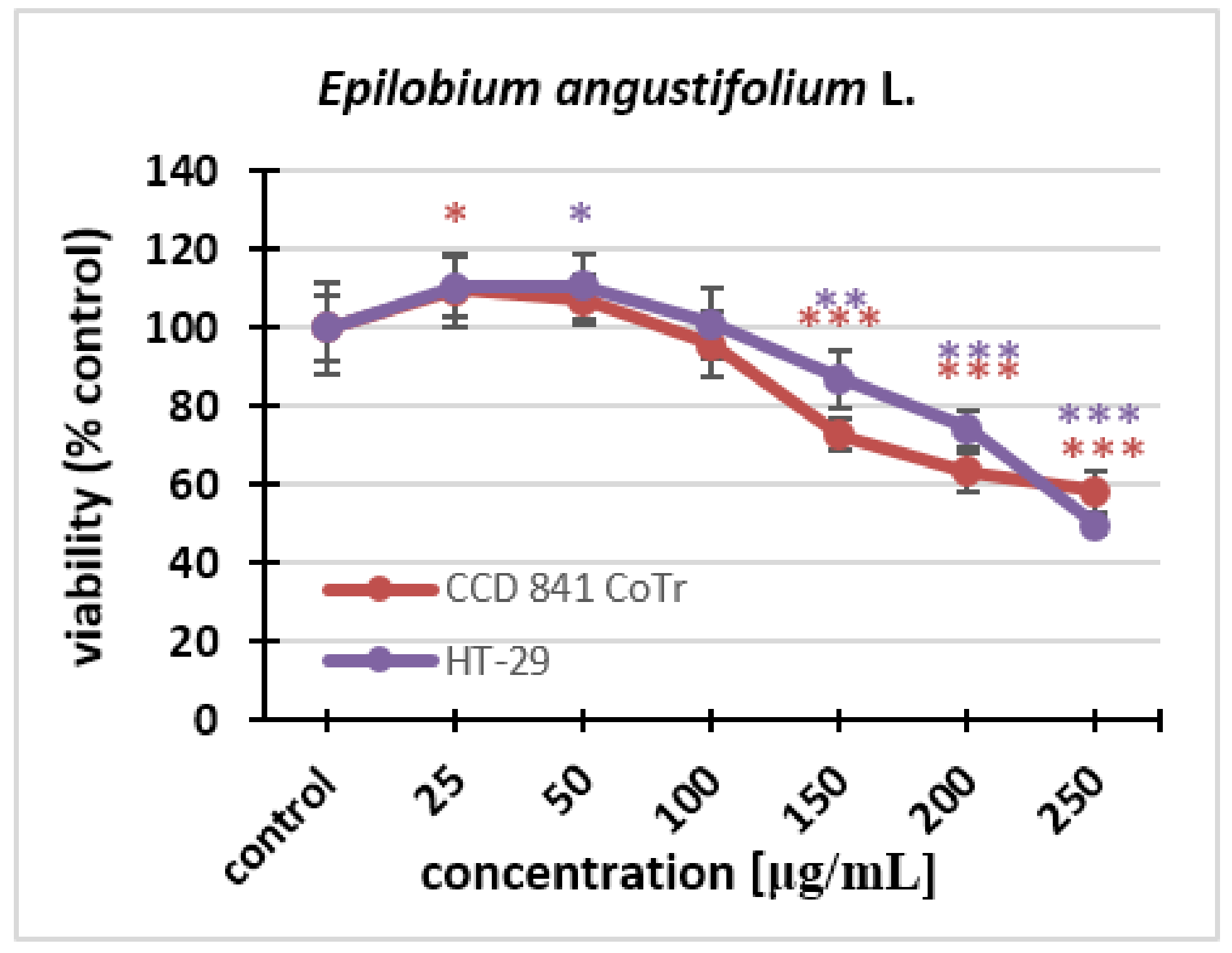
2.2.2. Neutral Red (NR) Uptake Assay
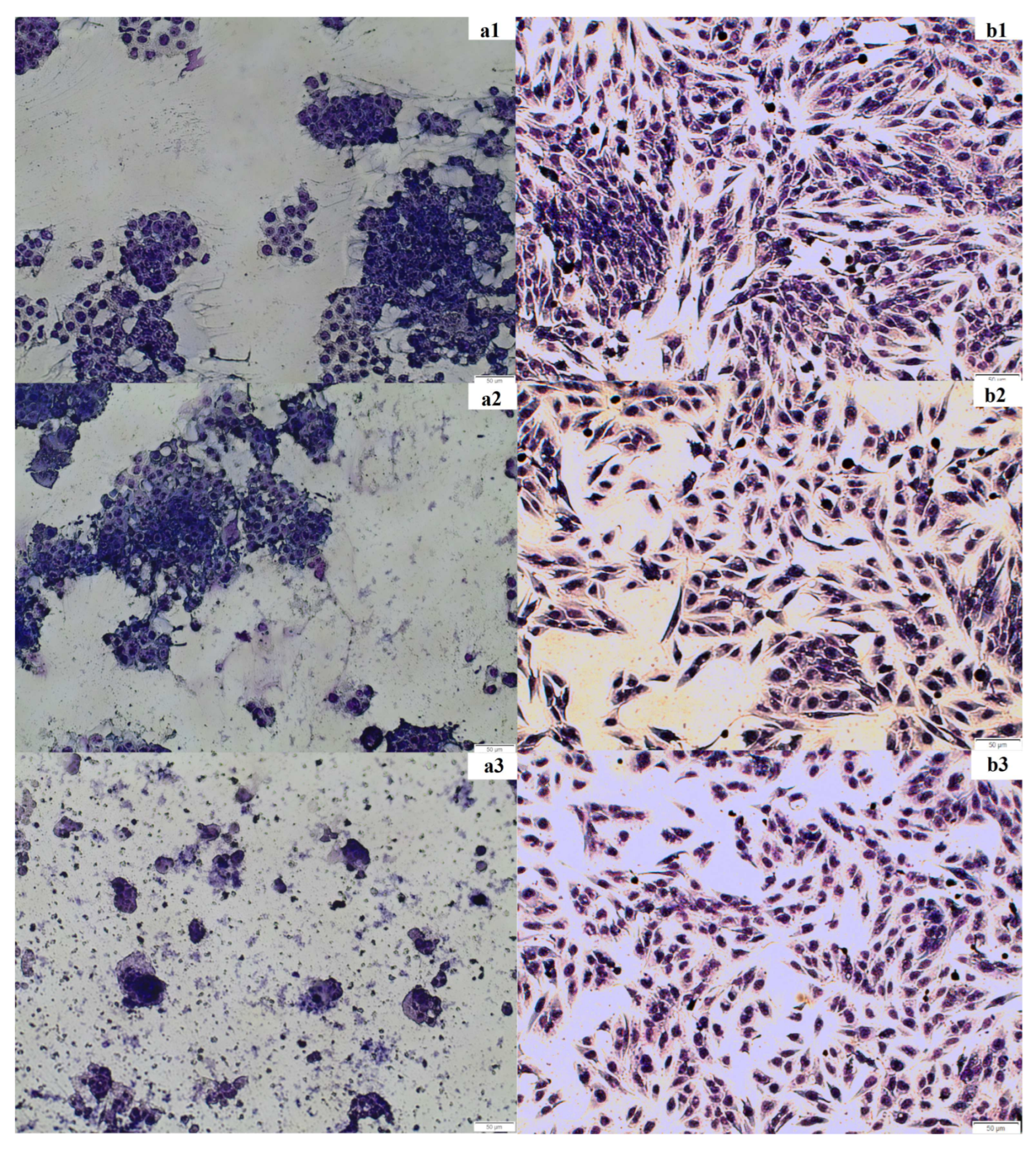
2.2.3. May–Grünwald–Giemsa Staining
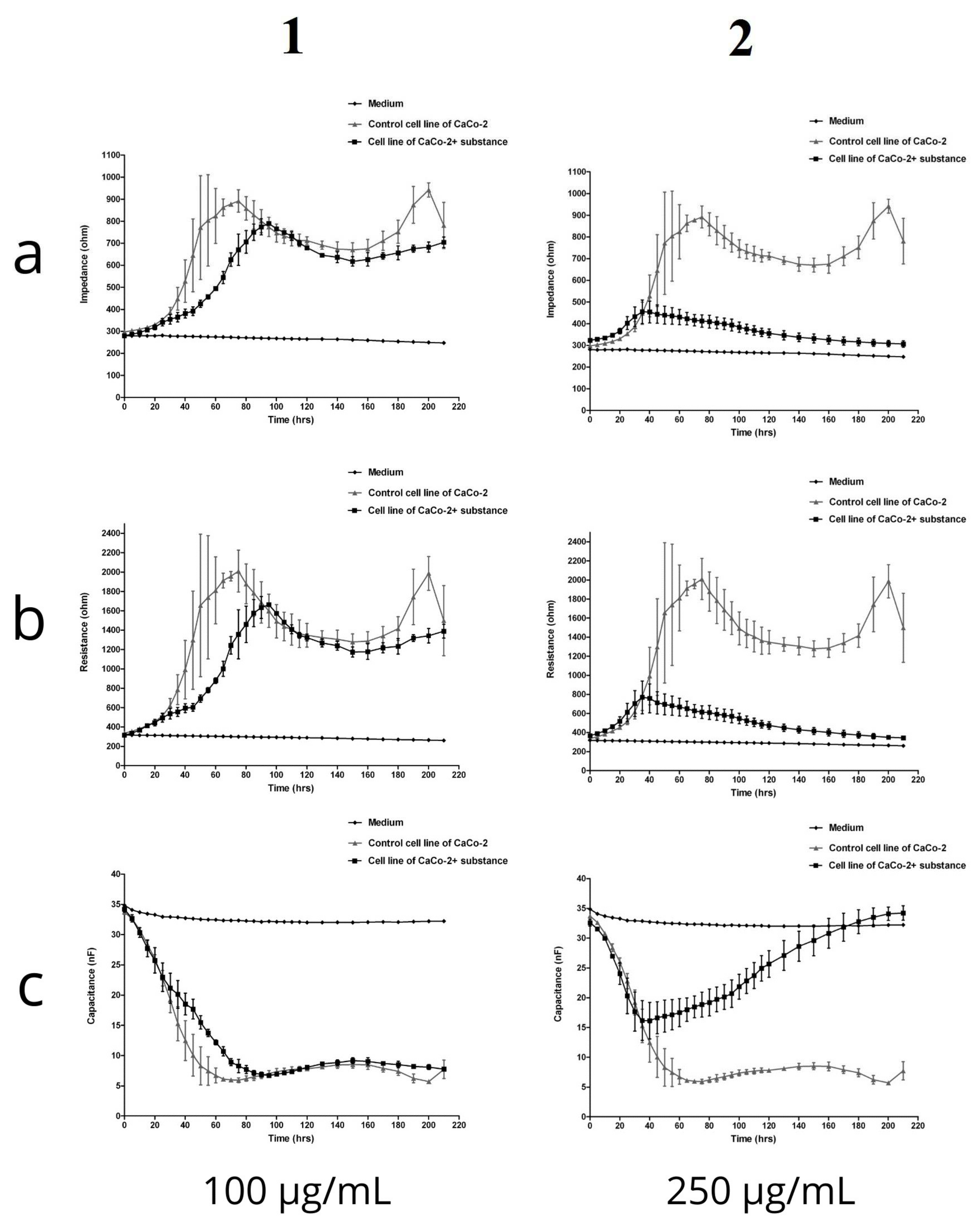
2.2.4. Electric Cell-Substrate Impedance Sensing (ECIS)
2.2.5. Antimicrobial Properties
3. Discussion
4. Materials and Methods
4.1. Materials
Plant Material
4.2. Qualitative and Quantitative Analysis of Samples by HPLC-ESI-QTOF-MS/MS
4.3. In Vitro Gastrointestinal Digestion of Fireweed Infusion
4.4. Cell Cultures
4.5. MTT Cytotoxicity Assay
4.6. MTT Proliferation Assay
4.7. Neutral Red (NR) Uptake Assay
4.8. May-Grünwald-Giemsa Staining
4.9. Electric Cell-Substrate Impedance Sensing (ECIS)
4.10. Bacterial Strains and Culture Method
Effect on Bacterial Growth
4.11. Statiscical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Sample Availability
References
- Battinelli, L.; Tita, B.; Evandri, M.G.; Mazzanti, G. Antimicrobial activity of Epilobium spp. extracts. Il Farm. 2001, 56, 345–348. [Google Scholar] [CrossRef]
- Ostrovska, H.; Oleshchuk, O.; Vannini, S.; Cataldi, S. Epilobium angustifolium L.: A medicinal plant with therapeutic properties. EuroBiotech J. 2017, 1, 126–131. [Google Scholar] [CrossRef] [Green Version]
- Bartfay, W.J.; Bartfay, E.; Johnson, J.G. Gram-negative and gram-positive antibacterial properties of the whole plant extract of willow herb (Epilobium angustifolium). Biol. Res. Nurs. 2012, 14, 85–89. [Google Scholar] [CrossRef] [PubMed]
- Kosalec, I.; Kopjar, N.; Kremer, D. Antimicrobial activity of willowherb (Epilobium angustifolium L.) leaves and flowers. Curr. Drug Targets 2013, 14, 986–991. [Google Scholar] [CrossRef] [PubMed]
- Deng, L.; Zong, W.; Tao, X.; Liu, S.; Feng, Z.; Lin, Y.; Liao, Z.; Chen, M. Evaluation of the therapeutic effect against benign prostatic hyperplasia and the active constituents from Epilobium angustifolium L. J. Ethnopharmacol. 2019, 232, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Hevesi Tóth, B.; Blazics, B.; Kéry, Á. Polyphenol composition and antioxidant capacity of Epilobium species. J. Pharm. Biomed. Anal. 2009, 49, 26–31. [Google Scholar] [CrossRef] [PubMed]
- Vitalone, A.; Allkanjari, O. Epilobium spp: Pharmacology and phytochemistry. Phyther. Res. 2018, 32, 1229–1240. [Google Scholar] [CrossRef] [PubMed]
- Shikov, A.N.; Poltanov, E.A.; Dorman, H.J.D.; Makarov, V.G.; Tikhonov, V.P.; Hiltunen, R. Chemical composition and in vitro antioxidant evaluation of commercial water-soluble willow herb (Epilobium angustifolium L.) extracts. J. Agric. Food Chem. 2006, 54, 3617–3624. [Google Scholar] [CrossRef]
- Schepetkin, I.A.; Ramstead, A.G.; Kirpotina, L.N.; Voyich, J.M.; Jutila, M.A.; Quinn, M.T. Therapeutic potential of polyphenols from Epilobium angustifolium (Fireweed). Phyther. Res. 2016, 30, 1287–1297. [Google Scholar] [CrossRef] [Green Version]
- Schepetkin, I.A.; Kirpotina, L.N.; Jakiw, L.; Khlebnikov, A.I.; Blaskovich, C.L.; Jutila, M.A.; Quinn, M.T. Immunomodulatory activity of oenothein B isolated from Epilobium angustifolium. J. Immunol. 2009, 183, 6754. [Google Scholar] [CrossRef] [Green Version]
- Sõukand, R.; Mattalia, G.; Kolosova, V.; Stryamets, N.; Prakofjewa, J.; Belichenko, O.; Kuznetsova, N.; Minuzzi, S.; Keedus, L.; Prūse, B.; et al. Inventing a herbal tradition: The complex roots of the current popularity of Epilobium angustifolium in Eastern Europe. J. Ethnopharmacol. 2020, 247, 112254. [Google Scholar] [CrossRef]
- Zunino, V.; Meineri, G.; Bellone, G.; Vizio, B.; Prati, A.; Grandi, M.; Radice, E.; Dal Bello, F.; Medana, C. Alcoholic extracts of Epilobium, Urtica dioica and Evernia prunastri with 5-fluorouracil in controlling murine colon carcinoma cell growth in vitro. Orient. Pharm. Exp. Med. 2017, 17, 325–336. [Google Scholar] [CrossRef]
- Soleymani, J.; Hasanzadeh, M.; Somi, M.H.; Shadjou, N.; Jouyban, A. Highly sensitive and specific cytosensing of HT 29 colorectal cancer cells using folic acid functionalized-KCC-1 nanoparticles. Biosens. Bioelectron. 2019, 132, 122–131. [Google Scholar] [CrossRef] [PubMed]
- Tariq, K.; Ghias, K. Colorectal cancer carcinogenesis: A review of mechanisms. Cancer Biol. Med. 2016, 13, 120–135. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- O’Keefe, S.J.D. Diet, microorganisms and their metabolites, and colon cancer. Nat. Rev. Gastroenterol. Hepatol. 2016, 13, 691–706. [Google Scholar] [CrossRef]
- Huang, M.; Wang, Y.; Xu, L.; You, M. Anti-tumor properties of Prunella vulgaris. Curr. Pharmacol. Rep. 2015, 1, 401–419. [Google Scholar] [CrossRef] [Green Version]
- Xavier, C.P.R.; Lima, C.F.; Fernandes-Ferreira, M.; Pereira-Wilson, C. Salvia fruticosa, Salvia officinalis, and rosmarinic acid induce apoptosis and inhibit proliferation of human colorectal cell lines: The role in MAPK/ERK pathway. Nutr. Cancer 2009, 61, 564–571. [Google Scholar] [CrossRef] [Green Version]
- Guo, Y.; Wu, R.; Gaspar, J.M.; Sargsyan, D.; Su, Z.Y.; Zhang, C.; Gao, L.; Cheng, D.; Li, W.; Wang, C.; et al. DNA methylome and transcriptome alterations and cancer prevention by curcumin in colitis-accelerated colon cancer in mice. Carcinogenesis 2018, 39, 669–680. [Google Scholar] [CrossRef]
- Szwajgier, D.; Baranowska-Wójcik, E.; Kukula-Koch, W.; Kowalik, K.; Polak-Berecka, M.; Waśko, A. Evolution of the anticholinesterase, antioxidant, and anti-inflammatory activity of Epilobium angustifolium L. infusion during in vitro digestion. J. Funct. Foods 2021, 85, 104645. [Google Scholar] [CrossRef]
- Baert, N.; Kim, J.; Karonen, M.; Salminen, J.P. Inter-population and inter-organ distribution of the main polyphenolic compounds of Epilobium angustifolium. Phytochemistry 2017, 134, 54–63. [Google Scholar] [CrossRef]
- Dacrema, M.; Sommella, E.; Santarcangelo, C.; Bruno, B.; Marano, M.G.; Insolia, V.; Saviano, A.; Campiglia, P.; Stornaiuolo, M.; Daglia, M. Metabolic profiling, in vitro bioaccessibility and in vivo bioavailability of a commercial bioactive Epilobium angustifolium L. extract. Biomed. Pharmacother. 2020, 131, 110670. [Google Scholar] [CrossRef] [PubMed]
- Vitalone, A.; McColl, J.; Thome, D.; Costa, L.G.; Tita, B. Characterization of the effect of Epilobium extracts on human cell proliferation. Pharmacology 2003, 69, 79–87. [Google Scholar] [CrossRef] [PubMed]
- Kia, B.H.; Noureini, S.K.; Kakhki, M.R.V. The extracts of Epilobium parviflorum inhibit MCF-7 breast cancer cells. Iran. J. Toxicol. 2021, 15, 65–72. [Google Scholar] [CrossRef]
- Kiss, A.; Stolarczyk, M.; Piwowarski, J.P.; Granica, S. Effects of standardized extracts from Epilobium sp. herbs on of hormone-dependent prostate cancer cells (LNCaP)-searching for active compounds. Pharmacol. Rep. 2013, 65, 13. [Google Scholar] [CrossRef]
- Granica, S.; Piwowarski, J.P.; Czerwińska, M.E.; Kiss, A.K. Phytochemistry, pharmacology and traditional uses of different Epilobium species (Onagraceae): A review. J. Ethnopharmacol. 2014, 156, 316–346. [Google Scholar] [CrossRef] [PubMed]
- Tita, B.; Abdel-Haq, H.; Vitalone, A.; Mazzanti, G.; Saso, L. Analgesic properties of Epilobium angustifolium, evaluated by the hot plate test and the writhing test. Il Farm. 2001, 56, 341–343. [Google Scholar] [CrossRef]
- Nowak, A.; Cybulska, K.; Makuch, E.; Kucharski, Ł.; Różewicka-Czabańska, M.; Prowans, P.; Czapla, N.; Bargiel, P.; Petriczko, J.; Klimowicz, A. In vitro human skin penetration, antioxidant and antimicrobial activity of ethanol-water extract of fireweed (Epilobium angustifolium L.). Molecules 2021, 26, 329. [Google Scholar] [CrossRef] [PubMed]
- Allen-Vercoe, E.; Jobin, C. Fusobacterium and Enterobacteriaceae: Important players for CRC? Immunol. Lett. 2014, 162, 54–61. [Google Scholar] [CrossRef] [Green Version]
- Wang, T.; Cai, G.; Qiu, Y.; Fei, N.; Zhang, M.; Pang, X.; Jia, W.; Cai, S.; Zhao, L. Structural segregation of gut microbiota between colorectal cancer patients and healthy volunteers. ISME J. 2012, 6, 320–329. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jobin, C. Colorectal cancer: Looking for answers in the microbiota. Cancer Discov. 2013, 3, 384–387. [Google Scholar] [CrossRef] [Green Version]
- Kiss, A.K.; Bazylko, A.; Filipek, A.; Granica, S.; Jaszewska, E.; Kiarszys, U.; Kośmider, A.; Piwowarski, J. Oenothein B’s contribution to the anti-inflammatory and antioxidant activity of Epilobium sp. Phytomedicine 2011, 18, 557–560. [Google Scholar] [CrossRef]
- Szwajgier, D.; Paduch, R.; Kukuła-Koch, W.; Polak-Berecka, M.; Waśko, A. Study on biological activity of bread enriched with natural polyphenols in terms of growth inhibition of tumor intestine cells. J. Med. Food 2020, 23, 181–190. [Google Scholar] [CrossRef] [PubMed]
- Kumar, P.; Nagarajan, A.; Uchil, P.D. Analysis of cell viability by the MTT Assay. Cold Spring Harb. Protoc. 2018, 2018, 469–471. [Google Scholar] [CrossRef]
- Ates, G.; Vanhaecke, T.; Rogiers, V.; Rodrigues, R.M. Assaying cellular viability using the neutral red uptake assay. Methods Mol. Biol. 2017, 1601, 19–26. [Google Scholar] [CrossRef] [PubMed]
- Hoeflinger, J.L.; Hoeflinger, D.E.; Miller, M.J. A dynamic regression analysis tool for quantitative assessment of bacterial growth written in Python. J. Microbiol. Methods 2017, 132, 83–85. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Name of Substance | Content in μg/mL after Digestion |
|---|---|
| Quercetine glucuronide | 593.0 |
| Giaverin | 212.7 |
| Afzelin | 148.2 |
| Neochlorogenic acid | 37.3 |
| Kaempferol glucoside | 24.9 |
| Chlorogenic acid | 20.8 |
| Isoquercetin | 15.6 |
| Hyperin gallate | 15.5 |
| Name of Substance | Before Digestion | SD | RSD | After Digestion | SD | RSD |
|---|---|---|---|---|---|---|
| Neochlorogenic acid | 0.28796 | 0.00434 | 1.5 | 0.00370 | 0.00012 | 3.4 |
| Chlorogenic acid | 0.09889 | 0.00279 | 2.8 | 0.00239 | 0.00009 | 3.8 |
| Isoquercetin | 0.44318 | 0.01478 | 3.3 | 0.00149 | 0.00013 | 8.8 |
| Kaempferol glucoside | 0.59884 | 0.06533 | 10.9 | 0.00252 | 0.00007 | 2.8 |
| Hyperin gallate | 1.48623 | 0.02287 | 1.5 | 0.00236 | 0.00008 | 3.6 |
| Quercetine glucuronide | 4.75316 | 0.09163 | 1.9 | 0.06274 | 0.00058 | 0.9 |
| Giaverin | 0.71688 | 0.01206 | 1.7 | 0.02349 | 0.00004 | 0.2 |
| Afzelin | 1.11672 | 0.03015 | 2.7 | 0.02849 | 0.00062 | 2.2 |
| Strain | Concentration of Fireweed Extract [μg/mL] | Lag Time (h) | Doubling Time (h) | Min OD | Max OD | Effect on Bacterial Growth |
|---|---|---|---|---|---|---|
| Lactobacillus rhamnosus | Control * | 12.12 | 3.24 | 0.43 | 1.99 | |
| 16 | 14.97 | 3.58 | 0.38 | 1.91 | no differences in growth | |
| 32 | 14.78 | 3.50 | 0.40 | 1.94 | ||
| 64 | 14.86 | 3.51 | 0.40 | 1.95 | ||
| 128 | 13.96 | 3.88 | 0.39 | 1.95 | ||
| 250 | 13.14 | 3.75 | 0.41 | 1.98 | ||
| Enterococcus faecalis | Control * | 7.46 | 10.20 | 0.19 | 0.61 | |
| 16 | 7.26 | 10.74 | 0.19 | 0.60 | ||
| 32 | 6.47 | 10.38 | 0.18 | 0.76 | stimulation of growth | |
| 64 | 6.47 | 11.75 | 0.18 | 0.77 | ||
| 128 | 5.30 | 8.97 | 0.19 | 1.07 | ||
| 250 | 7.28 | 6.18 | 0.18 | 1.35 | ||
| Enterobacter cloacae | Control * | 0.52 | 5.47 | 0.19 | 1.29 | |
| 16 | 0 | 6.25 | 0.27 | 1.20 | ||
| 32 | 0 | 5.92 | 0.19 | 1.17 | no differences in growth | |
| 64 | 0 | 5,67 | 0.14 | 1.17 | ||
| 128 | 0 | 6.42 | 0.19 | 1.20 | ||
| 250 | 0 | 6.05 | 0.22 | 1.32 | ||
| Escherichia coli | Control * | 0 | 14.85 | 0.20 | 1.41 | |
| 16 | 0 | 16.10 | 0.22 | 1.05 | ||
| 32 | 0 | 15.96 | 0.24 | 1.07 | MIC-64 μg/mL | |
| 64 | 0 | 20.21 | 0.22 | 0.84 | ||
| 128 | 0 | 16.74 | 0.19 | 0.83 | ||
| 250 | 0 | 25.12 | 0.25 | 0.87 | ||
| Bifidobacterium adolescentis | Control * | 4.53 | 33.25 | 0.30 | 0.78 | |
| 16 | 6.98 | 38.41 | 0.28 | 0.67 | ||
| 32 | 2.77 | 40.73 | 0.24 | 0.69 | no differences in growth | |
| 64 | 9.15 | 42.12 | 0.25 | 0.70 | ||
| 128 | 3.77 | 39.43 | 0.28 | 0.74 | ||
| 250 | 0 | 93.21 | 0.39 | 0.69 | ||
| Bifidobacterium longum | Control * | 4.66 | 34.13 | 0.27 | 0.75 | |
| 16 | 5.72 | 46.14 | 0.25 | 0.64 | ||
| 32 | 8 | 52.48 | 0.26 | 0.63 | no differences in growth | |
| 64 | 6.72 | 41.14 | 0.25 | 0.63 | ||
| 128 | 0 | 48.85 | 0.26 | 0.61 | ||
| 250 | 0 | 73.18 | 0.27 | 0.53 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kowalik, K.; Polak-Berecka, M.; Prendecka-Wróbel, M.; Pigoń-Zając, D.; Niedźwiedź, I.; Szwajgier, D.; Baranowska-Wójcik, E.; Waśko, A. Biological Activity of an Epilobium angustifolium L. (Fireweed) Infusion after In Vitro Digestion. Molecules 2022, 27, 1006. https://doi.org/10.3390/molecules27031006
Kowalik K, Polak-Berecka M, Prendecka-Wróbel M, Pigoń-Zając D, Niedźwiedź I, Szwajgier D, Baranowska-Wójcik E, Waśko A. Biological Activity of an Epilobium angustifolium L. (Fireweed) Infusion after In Vitro Digestion. Molecules. 2022; 27(3):1006. https://doi.org/10.3390/molecules27031006
Chicago/Turabian StyleKowalik, Klaudia, Magdalena Polak-Berecka, Monika Prendecka-Wróbel, Dominika Pigoń-Zając, Iwona Niedźwiedź, Dominik Szwajgier, Ewa Baranowska-Wójcik, and Adam Waśko. 2022. "Biological Activity of an Epilobium angustifolium L. (Fireweed) Infusion after In Vitro Digestion" Molecules 27, no. 3: 1006. https://doi.org/10.3390/molecules27031006
APA StyleKowalik, K., Polak-Berecka, M., Prendecka-Wróbel, M., Pigoń-Zając, D., Niedźwiedź, I., Szwajgier, D., Baranowska-Wójcik, E., & Waśko, A. (2022). Biological Activity of an Epilobium angustifolium L. (Fireweed) Infusion after In Vitro Digestion. Molecules, 27(3), 1006. https://doi.org/10.3390/molecules27031006