
Purification, Identification and Molecular Docking of Novel Antioxidant Peptides from Walnut (Juglans regia L.) Protein Hydrolysates


Abstract

1. Introduction
2. Materials and Methods
2.1. Materials
2.2. Extraction of WPI
2.3. Preparation of Walnut Protein Hydrolysates (WPH)
2.4. Determination of Degree of Hydrolysis (DH)
2.5. Hydrolysis Kinetic
2.5.1. Enzymatic Hydrolysis
2.5.2. Hydrolysis Model
2.6. Amino Acid Composition
2.7. Structural Characterization of WPHs
2.7.1. Ultraviolet Visible (UV) Absorption Spectroscopy
2.7.2. Intrinsic Fluorescence Spectroscopy
2.7.3. Fourier-Transform Infrared Spectroscopy
2.8. Determination of Antioxidant Activities
2.8.1. DPPH Radical Scavenging Activity
2.8.2. ABTS+• Scavenging Activity
2.8.3. Hydroxyl Radical Scavenging Activity
2.8.4. Reducing Power
2.9. Determination of Molecular Weight Distribution
2.10. Separation and Purification of WPH
2.11. Identification of Peptides of WPH by LC-MS/MS
2.12. Bioinformatics Determination of Physicochemical Properties of Antioxidant Peptides
2.13. Molecular Docking
2.14. Statistical Analysis
3. Results and Discussion
3.1. Screening of Enzymes
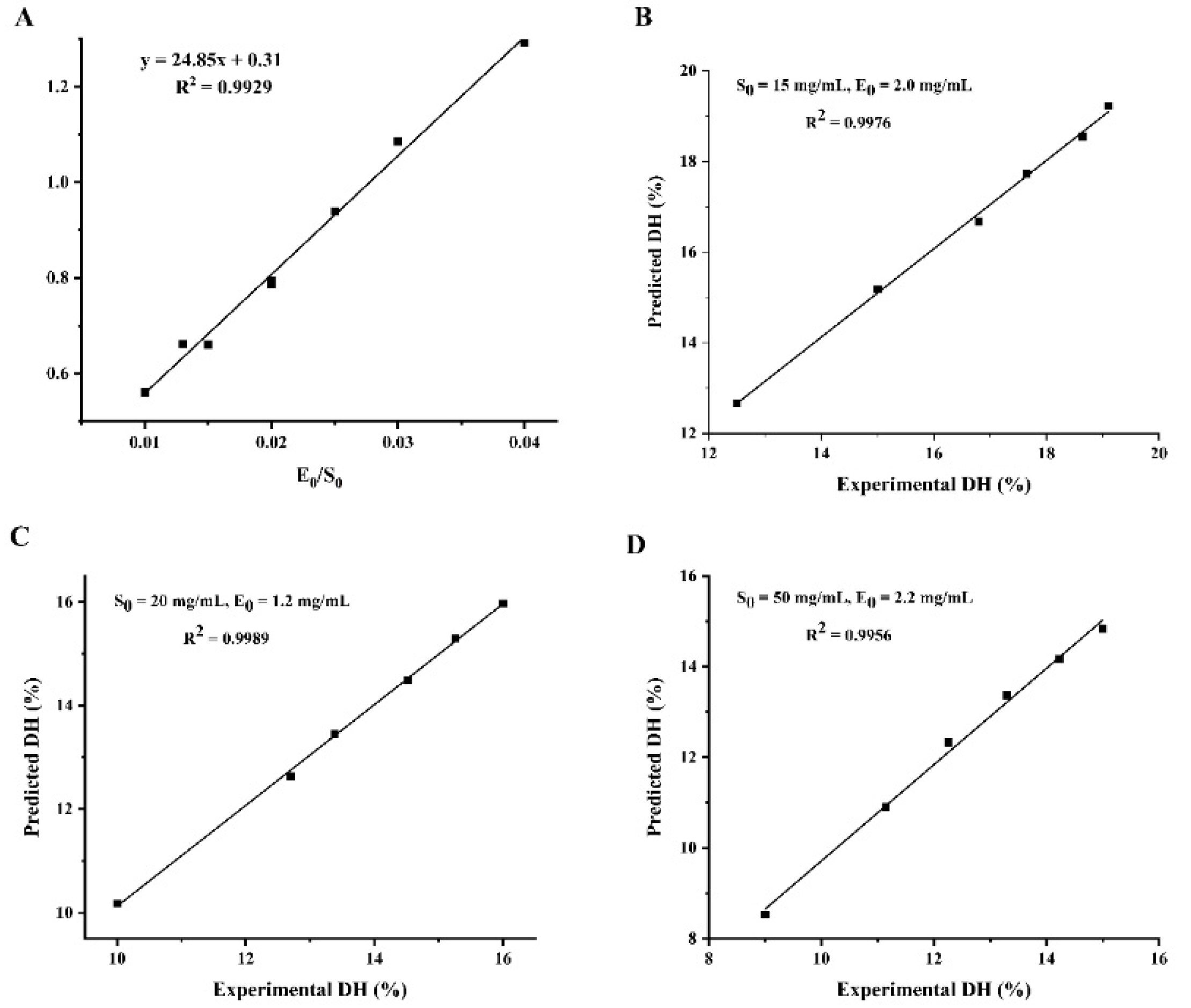
3.2. The Enzymolysis Kinetic Model of WPI
3.3. Amino Acid Composition Analysis
3.4. Structural Characteristics by Spectroscopy
3.4.1. UV-Visible Absorption Spectroscopy
3.4.2. Fluorescence Spectra Analysis
3.4.3. FTIR Spectra Analysis
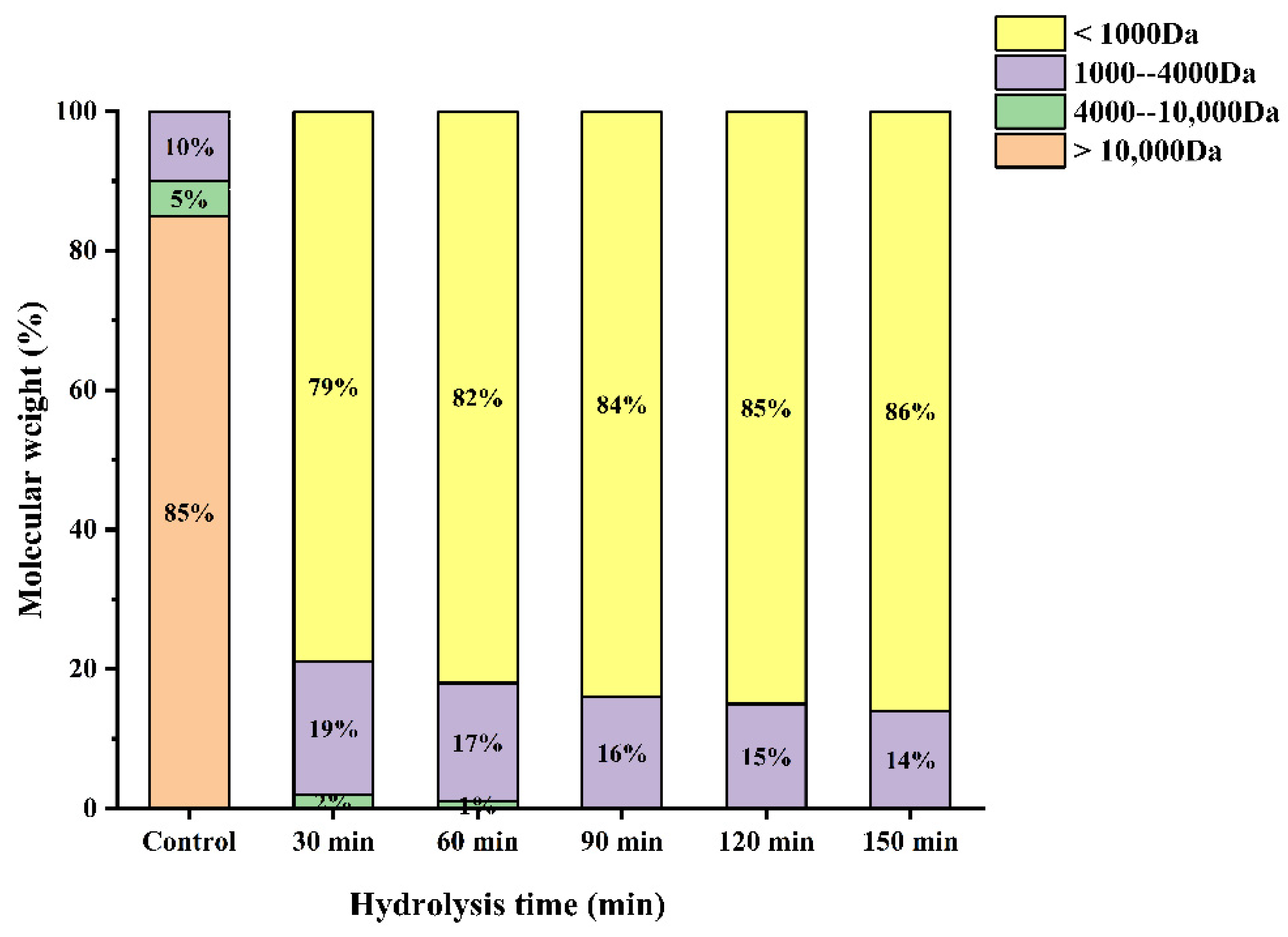
3.5. Molecular Weight Distribution
3.6. In Vitro Antioxidant Activities
3.6.1. DPPH Radical Scavenging Activity
3.6.2. ABTS Radical Scavenging Activity
3.6.3. Hydroxyl Radical Scavenging Activity
3.6.4. Reducing Power
3.7. Separation and Purification of WPH
3.8. Identification of Antioxidant Peptides by LC-MS/MS
3.9. Molecular Docking Analysis
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Valko, M.; Leibfritz, D.; Moncol, J.; Cronin, M.T.D.; Mazur, M.; Telser, J. Free radicals and antioxidants in normal physiological functions and human disease. Int. J. Biochem. Cell Biol. 2007, 39, 44–84. [Google Scholar] [CrossRef]
- Giacco, F.; Brownlee, M. Oxidative Stress and Diabetic Complications. Circ. Res. 2010, 107, 1058–1070. [Google Scholar] [CrossRef]
- Lobo, V.; Patil, A.; Phatak, A.; Chandra, N. Free radicals, antioxidants and functional foods: Impact on human health. Pharmacogn. Rev. 2010, 4, 118–126. [Google Scholar] [CrossRef]
- Zhan, J.; Li, G.; Dang, Y.; Pan, D. Purification and identification of a novel hypotensive and antioxidant peptide from porcine plasma. J. Sci. Food Agric. 2022, 102, 4933–4941. [Google Scholar] [CrossRef]
- Qian, Z.-J.; Jung, W.-K.; Kim, S.-K. Free radical scavenging activity of a novel antioxidative peptide purified from hydrolysate of bullfrog skin, Rana catesbeiana Shaw. Bioresour. Technol. 2008, 99, 1690–1698. [Google Scholar] [CrossRef]
- Noman, A.; Wang, Y.; Zhang, C.; Abed, S.M. Antioxidant Activity of Hybrid Sturgeon (Huso dauricus × Acipenser schrenckii) Protein Hydrolysate Prepared Using Bromelain, Its Fractions and Purified Peptides. Pol. J. Food Nutr. Sci. 2022, 72, 79–89. [Google Scholar] [CrossRef]
- Zhang, F.; Qu, J.; Thakur, K.; Zhang, J.-G.; Mocan, A.; Wei, Z.-J. Purification and identification of an antioxidative peptide from peony (Paeonia suffruticosa Andr.) seed dreg. Food Chem. 2019, 285, 266–274. [Google Scholar] [CrossRef]
- Luo, X.; Wu, S.; Xue, J.; Hu, H.; He, Z.; Liu, X.; Wu, F. The bioactive peptide screening from Torreya grandis meal protein hydrolysates. Food Biosci. 2021, 44, 101419. [Google Scholar] [CrossRef]
- Zhang, X.; Dai, Z.; Zhang, Y.; Dong, Y.; Hu, X. Structural characteristics and stability of salmon skin protein hydrolysates obtained with different proteases. Lwt-Food Sci. Technol. 2022, 153, 112460. [Google Scholar] [CrossRef]
- Feng, L.; Peng, F.; Wang, X.; Li, M.; Lei, H.; Xu, H. Identification and characterization of antioxidative peptides derived from simulated in vitro gastrointestinal digestion of walnut meal proteins. Food Res. Int. 2019, 116, 518–526. [Google Scholar] [CrossRef] [PubMed]
- Hu, F.; Ci, A.-T.; Wang, H.; Zhang, Y.-Y.; Zhang, J.-G.; Thakur, K.; Wei, Z.-J. Identification and hydrolysis kinetic of a novel antioxidant peptide from pecan meal using Alcalase. Food Chem. 2018, 261, 301–310. [Google Scholar] [CrossRef] [PubMed]
- Zou, T.-B.; He, T.-P.; Li, H.-B.; Tang, H.-W.; Xia, E.-Q. The Structure-Activity Relationship of the Antioxidant Peptides from Natural Proteins. Molecules 2016, 21, 72. [Google Scholar] [CrossRef] [PubMed]
- Adlernissen, J. Enzymic hydrolysis of food proteins. Can. Med. Assoc. J. 1986, 172, 1783–1785. [Google Scholar]
- Dong, X.-H.; Li, J.; Jiang, G.-X.; Li, H.-Y.; Zhao, M.-M.; Jiang, Y.-M. Effects of combined high pressure and enzymatic treatments on physicochemical and antioxidant properties of peanut proteins. Food Sci. Nutr. 2019, 7, 1417–1425. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Yu, G.; Sun, A.; Cai, X.; Yue, C. Effects of enzymolysis on the functional ingredient contents and antioxidant activities of aqueous enzymatic extracts from Rice Bran and correlation analysis. Food Sci. Biotechnol. 2015, 24, 1609–1616. [Google Scholar] [CrossRef]
- Song, R.; Liang, T.; Shen, Q.; Liu, J.; Lu, Y.; Tang, C.; Chen, X.; Hou, T.; Chen, Y. The optimization of production and characterization of antioxidant peptides from protein hydrolysates of Agrocybe aegerita. Lwt-Food Sci. Technol. 2020, 134, 109987. [Google Scholar] [CrossRef]
- An, L.; Wang, G.; Jia, H.; Liu, C.; Sui, W.; Si, C. Fractionation of enzymatic hydrolysis lignin by sequential extraction for enhancing antioxidant performance. Int. J. Biol. Macromol. 2017, 99, 674–681. [Google Scholar] [CrossRef] [PubMed]
- Cui, Q.; Sun, Y.; Cheng, J.; Guo, M. Effect of two-step enzymatic hydrolysis on the antioxidant properties and proteomics of hydrolysates of milk protein concentrate. Food Chem. 2022, 366, 130711. [Google Scholar] [CrossRef] [PubMed]
- Kong, X.Z.; Zhang, L.N.; Song, W.G.; Zhang, C.M.; Hua, Y.F.; Chen, Y.M.; Li, X.F. Separation, identification and molecular binding mechanism of dipeptidyl peptidase IV inhibitory peptides derived from walnut (Juglans regia L.) protein. Food Chem. 2021, 347, 129062. [Google Scholar] [CrossRef] [PubMed]
- Famuwagun, A.A.; Alashi, A.M.; Gbadamosi, S.O.; Taiwo, K.A.; Oyedele, D.; Adebooye, O.C.; Aluko, R.E. Effect of Protease Type and Peptide Size on the in vitro Antioxidant, Antihypertensive and Anti-Diabetic Activities of Eggplant Leaf Protein Hydrolysates. Foods 2021, 10, 1112. [Google Scholar] [CrossRef]
- Zheng, Z.; Li, J.; Liu, Y. Effects of partial hydrolysis on the structural, functional and antioxidant properties of oat protein isolate. Food Funct. 2020, 11, 3144–3155. [Google Scholar] [CrossRef]
- Ghribi, A.M.; Gafsi, I.M.; Sila, A.; Blecker, C.; Danthine, S.; Attia, H.; Bougatef, A.; Besbes, S. Effects of enzymatic hydrolysis on conformational and functional properties of chickpea protein isolate. Food Chem. 2015, 187, 322–330. [Google Scholar] [CrossRef] [PubMed]
- Atmaca, G. Antioxidant effects of sulfur-containing amino acids. Yonsei Med. J. 2004, 45, 776–788. [Google Scholar] [CrossRef] [PubMed]
- Wade, A.M.; Tucker, H.N. Antioxidant characteristics of L-histidine. J. Nutr. Biochem. 1998, 9, 308–315. [Google Scholar] [CrossRef]
- Moghadam, M.; Salami, M.; Mohammadian, M.; Emam-Djomeh, Z.; Jahanbani, R.; Moosavi-Movahedi, A.A. Physicochemical and bio-functional properties of walnut proteins as affected by trypsin-mediated hydrolysis. Food Biosci. 2020, 36, 100611. [Google Scholar] [CrossRef]
- Zhou, C.; Hu, J.; Yu, X.; Yagoub, A.E.A.; Zhang, Y.; Ma, H.; Gao, X.; Otu, P.N.Y. Heat and/or ultrasound pretreatments motivated enzymolysis of corn gluten meal: Hydrolysis kinetics and protein structure. Lwt-Food Sci. Technol. 2017, 77, 488–496. [Google Scholar] [CrossRef]
- Wang, H.; Wang, J.; Lv, Z.; Liu, Y.; Lu, F. Preparing oligopeptides from broken rice protein by ultrafiltration-coupled enzymatic hydrolysis. Eur. Food Res. Technol. 2013, 236, 419–424. [Google Scholar] [CrossRef]
- Oussaief, O.; Jrad, Z.; Adt, I.; Dbara, M.; Khorchani, T.; El-Hatmi, H. Antioxidant activities of enzymatic-hydrolysed proteins of dromedary (Camelus dromedarius) colostrum. Int. J. Dairy Technol. 2020, 73, 333–340. [Google Scholar] [CrossRef]
- Pownall, T.L.; Udenigwe, C.C.; Aluko, R.E. Amino Acid Composition and Antioxidant Properties of Pea Seed (Pisum sativum L.) Enzymatic Protein Hydrolysate Fractions. J. Agric. Food Chem. 2010, 58, 4712–4718. [Google Scholar] [CrossRef]
- Wu, D.; Sun, N.; Ding, J.; Zhu, B.; Lin, S. Evaluation and structure-activity relationship analysis of antioxidant shrimp peptides. Food Funct. 2019, 10, 5605–5615. [Google Scholar] [CrossRef]
- Re, R.; Pellegrini, N.; Proteggente, A.; Pannala, A.; Yang, M.; Rice-Evans, C. Antioxidant activity applying an improved ABTS radical cation decolorization assay. Free Radic. Biol. Med. 1999, 26, 1231–1237. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Wang, G.; Chen, N.; An, F.; Zhang, R.; Zhang, Y.; Rahman, M.U.; Zhang, Y. Characterization of structural, functional and antioxidant properties and amino acid composition of pepsin-derived glutelin-1 hydrolysate from walnut processing by-products. Rsc Adv. 2021, 11, 19158–19168. [Google Scholar] [CrossRef]
- Tang, C.-H.; Wang, X.-S.; Yang, X.-Q. Enzymatic hydrolysis of hemp (Cannabis sativa L.) protein isolate by various proteases and antioxidant properties of the resulting hydrolysates. Food Chem. 2009, 114, 1484–1490. [Google Scholar] [CrossRef]
- Zhu, K.; Zhou, H.; Qian, H. Antioxidant and free radical-scavenging activities of wheat germ protein hydrolysates (WGPH) prepared with alcalase. Process Biochem. 2006, 41, 1296–1302. [Google Scholar] [CrossRef]
- Ghassem, M.; Arihara, K.; Mohammadi, S.; Sani, N.A.; Babji, A.S. Identification of two novel antioxidant peptides from edible bird’s nest (Aerodramus fuciphagus) protein hydrolysates. Food Funct. 2017, 8, 2046–2052. [Google Scholar] [CrossRef] [PubMed]
- Habinshuti, I.; Mu, T.-H.; Zhang, M. Ultrasound microwave-assisted enzymatic production and characterisation of antioxidant peptides from sweet potato protein. Ultrason. Sonochem. 2020, 69, 105262. [Google Scholar] [CrossRef]
- Chen, N.; Yang, H.; Sun, Y.; Niu, J.; Liu, S. Purification and identification of antioxidant peptides from walnut (Juglans regia L.) protein hydrolysates. Peptides 2012, 38, 344–349. [Google Scholar] [CrossRef]
- Safitri, N.M.; Hsu, J.-L.; Violando, W.A. Antioxidant Activity from the Enzymatic Hydrolysates of Chlorella sorokiniana and Its Potential Peptides Identification in Combination with Molecular Docking Analysis. Turk. J. Fish. Aquat. Sci. 2022, 22, TRJFAS20316. [Google Scholar] [CrossRef]
- Umayaparvathi, S.; Meenakshi, S.; Vimalraj, V.; Arumugam, M.; Balasubramanian, T. Isolation and Structural Elucidation of Antioxidant Peptides from Oyster (Saccostrea cucullata) Protein Hydrolysate. Protein Pept. Lett. 2014, 21, 1073–1083. [Google Scholar] [CrossRef]
- Wen, C.; Zhang, J.; Zhang, H.; Duan, Y.; Ma, H. Study on the structure-activity relationship of watermelon seed antioxidant peptides by using molecular simulations. Food Chem. 2021, 364, 130432. [Google Scholar] [CrossRef]












{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Type of Enzyme | pH | Temperature (°C) |
|---|---|---|
| Alcalase | 9.0 | 55 |
| Flavourzyme | 7.5 | 45 |
| Neutrase | 7.0 | 50 |
| Pepsin | 2.0 | 37 |
| Trypsin | 7.0 | 37 |
| S0 (mg/mL) | E0 (mg/mL) | E0/S0 | a (min−1) | b | R2 |
|---|---|---|---|---|---|
| 40 | 0.6 | 0.015 | 0.660 | 0.303 | 0.997 |
| 40 | 0.8 | 0.020 | 0.794 | 0.289 | 0.998 |
| 40 | 1.0 | 0.025 | 0.939 | 0.299 | 0.997 |
| 40 | 1.2 | 0.030 | 1.085 | 0.302 | 0.995 |
| 20 | 0.8 | 0.040 | 1.293 | 0.273 | 0.998 |
| 40 | 0.8 | 0.020 | 0.787 | 0.235 | 0.998 |
| 60 | 0.8 | 0.013 | 0.662 | 0.229 | 0.997 |
| 80 | 0.8 | 0.010 | 0.561 | 0.218 | 0.998 |
| AA | Control | 30 min | 60 min | 90 min | 120 min | 150 min |
|---|---|---|---|---|---|---|
| Asp | 10.03 | 10.58 | 10.27 | 10.22 | 10.09 | 10.10 |
| Thr | 3.66 | 3.49 | 3.54 | 3.62 | 3.43 | 3.44 |
| Ser | 5.28 | 4.85 | 5.33 | 5.53 | 5.27 | 5.35 |
| Glu | 23.46 | 21.71 | 22.19 | 22.26 | 22.44 | 22.6 |
| Gly | 4.46 | 4.45 | 4.46 | 4.38 | 4.4 | 4.39 |
| Ala | 4.01 | 4.63 | 4.46 | 4.36 | 4.34 | 4.32 |
| Cys | 0.55 | 0.23 | 0.21 | 0.34 | 0.23 | 0.25 |
| Val | 4.39 | 5.07 | 4.83 | 4.98 | 4.74 | 4.70 |
| Met | 0.15 | 1.34 | 1.38 | 1.24 | 1.57 | 1.46 |
| Ile | 3.65 | 3.95 | 3.67 | 3.59 | 3.75 | 3.59 |
| Leu | 6.12 | 7.15 | 6.96 | 6.64 | 6.96 | 6.89 |
| Tyr | 1.72 | 2.72 | 2.81 | 2.5 | 2.76 | 2.86 |
| Phe | 4.48 | 4.57 | 4.43 | 4.5 | 4.49 | 4.49 |
| Lys | 3.56 | 2.7 | 2.61 | 2.6 | 2.55 | 2.53 |
| His | 2.33 | 2.82 | 2.75 | 2.69 | 2.74 | 2.85 |
| Arg | 14.95 | 14.56 | 15.01 | 14.64 | 14.97 | 14.98 |
| Pro | 7.19 | 5.17 | 5.10 | 5.91 | 5.27 | 5.20 |
| EAA | 26.01 | 28.27 | 27.42 | 27.17 | 27.48 | 27.1 |
| SCAA | 0.70 | 1.57 | 1.59 | 1.58 | 1.80 | 1.71 |
| BCAA | 14.16 | 16.17 | 15.46 | 15.21 | 15.45 | 15.18 |
| DEAA | 23.61 | 23.05 | 23.57 | 23.5 | 24.01 | 24.06 |
| PCAA | 20.84 | 20.08 | 20.37 | 19.93 | 20.26 | 20.36 |
| HAA | 31.56 | 33.27 | 32.26 | 32.42 | 32.49 | 32.05 |
| AAA | 6.20 | 7.30 | 7.25 | 6.94 | 7.43 | 7.35 |
| Hydrolysis Time (min) | α-Helical (%) | β-Sheet (%) | β-Turn (%) | Random Coil (%) |
|---|---|---|---|---|
| Control | 21.08% | 32.92% | 45.99% | 0.01% |
| 30 | 3.88% | 60.38% | 10.83% | 24.92% |
| 60 | 5.61% | 62.07% | 13.04% | 19.28% |
| 90 | 5.02% | 57.18% | 21.82% | 15.98% |
| 120 | 0.01% | 14.93% | 63.28% | 21.78% |
| 150 | 12.97% | 21.35% | 59.02% | 6.66% |
| Peptide Sequence | Mass (Da) | Origin Protein | Location |
|---|---|---|---|
| GFEWVAF | 855.406 | 11S globulin-like | 422–428 |
| LFPGCPE | 819.372 | 11S globulin-like | 108–114 |
| QAGQLLPL | 839.498 | glutaredoxin-like | 104–111 |
| HADMVFY | 883.384 | vicilin-like seed storage protein At2g18540 | 90–96 |
| VPQNF | 604.312 | type IV inositol polyphosphate 5-phosphatase 9 | 170–174 |
| PSYQPTP | 789.379 | 11S globulin-like | 85–91 |
| NHCQYYL | 997.424 | 2S sulfur-rich seed storage protein 2 | 57–63 |
| NLFHKRP | 911.519 | Haloacid dehalogenase-like hydrolase domain-containing protein At2g33255 | 143–148 |
| LFPGCPET | 920.420 | 11S globulin-like | 110–117 |
| SNAPQL | 629.326 | 11S globulin-like | 92–97 |
| Peptide Sequence | Toxicity | Steric Hindrance | Amphipathicity | Net Hydrogen | Hydrophobicity (kcal/mol) |
|---|---|---|---|---|---|
| PSYQPTP | Non-toxin | 0.50 | 0.18 | 0.71 | +9.09 |
| NLFHKRP | Non-toxin | 0.53 | 1.08 | 1.29 | +12.87 |
| NHCQYYL | Non-toxin | 0.57 | 0.39 | 1.00 | +9.16 |
| HADMVFY | Non-toxin | 0.59 | 0.21 | 0.43 | +10.82 |
| GFEWVAF | Non-toxin | 0.64 | 0.18 | 0.29 | +7.21 |
| QAGQLLPL | Non-toxin | 0.56 | 0.31 | 0.50 | +7.48 |
| LFPGCPE | Non-toxin | 0.56 | 0.18 | 0.14 | +9.98 |
| LFPGCPET | Non-toxin | 0.56 | 0.16 | 0.25 | +10.23 |
| VPQNF | Non-toxin | 0.64 | 0.25 | 0.80 | +7.49 |
| SNAPQL | Non-toxin | 0.56 | 0.21 | 0.83 | +9.37 |
| Number | Peptide Sequence | Length | Binding Energy |
|---|---|---|---|
| 1 | PSYQPTP | 7 | −4.423 |
| 2 | NLFHKRP | 7 | −4.401 |
| 3 | NHCQYYL | 7 | −4.379 |
| 4 | HADMVFY | 7 | −4.263 |
| 5 | GFEWVAF | 7 | −4.181 |
| 6 | QAGQLLPL | 8 | −4.143 |
| 7 | LFPGCPE | 7 | −4.094 |
| 8 | LFPGCPET | 8 | −4.08 |
| 9 | VPQNF | 5 | −3.99 |
| 10 | SNAPQL | 6 | −3.592 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fan, L.; Mao, X.; Wu, Q. Purification, Identification and Molecular Docking of Novel Antioxidant Peptides from Walnut (Juglans regia L.) Protein Hydrolysates. Molecules 2022, 27, 8423. https://doi.org/10.3390/molecules27238423
Fan L, Mao X, Wu Q. Purification, Identification and Molecular Docking of Novel Antioxidant Peptides from Walnut (Juglans regia L.) Protein Hydrolysates. Molecules. 2022; 27(23):8423. https://doi.org/10.3390/molecules27238423
Chicago/Turabian StyleFan, Luhao, Xiaoying Mao, and Qingzhi Wu. 2022. "Purification, Identification and Molecular Docking of Novel Antioxidant Peptides from Walnut (Juglans regia L.) Protein Hydrolysates" Molecules 27, no. 23: 8423. https://doi.org/10.3390/molecules27238423
APA StyleFan, L., Mao, X., & Wu, Q. (2022). Purification, Identification and Molecular Docking of Novel Antioxidant Peptides from Walnut (Juglans regia L.) Protein Hydrolysates. Molecules, 27(23), 8423. https://doi.org/10.3390/molecules27238423