
Effects of Different Opioid Drugs on Oxidative Status and Proteasome Activity in SH-SY5Y Cells

 , , , , ,
, , , , ,  , and
, and 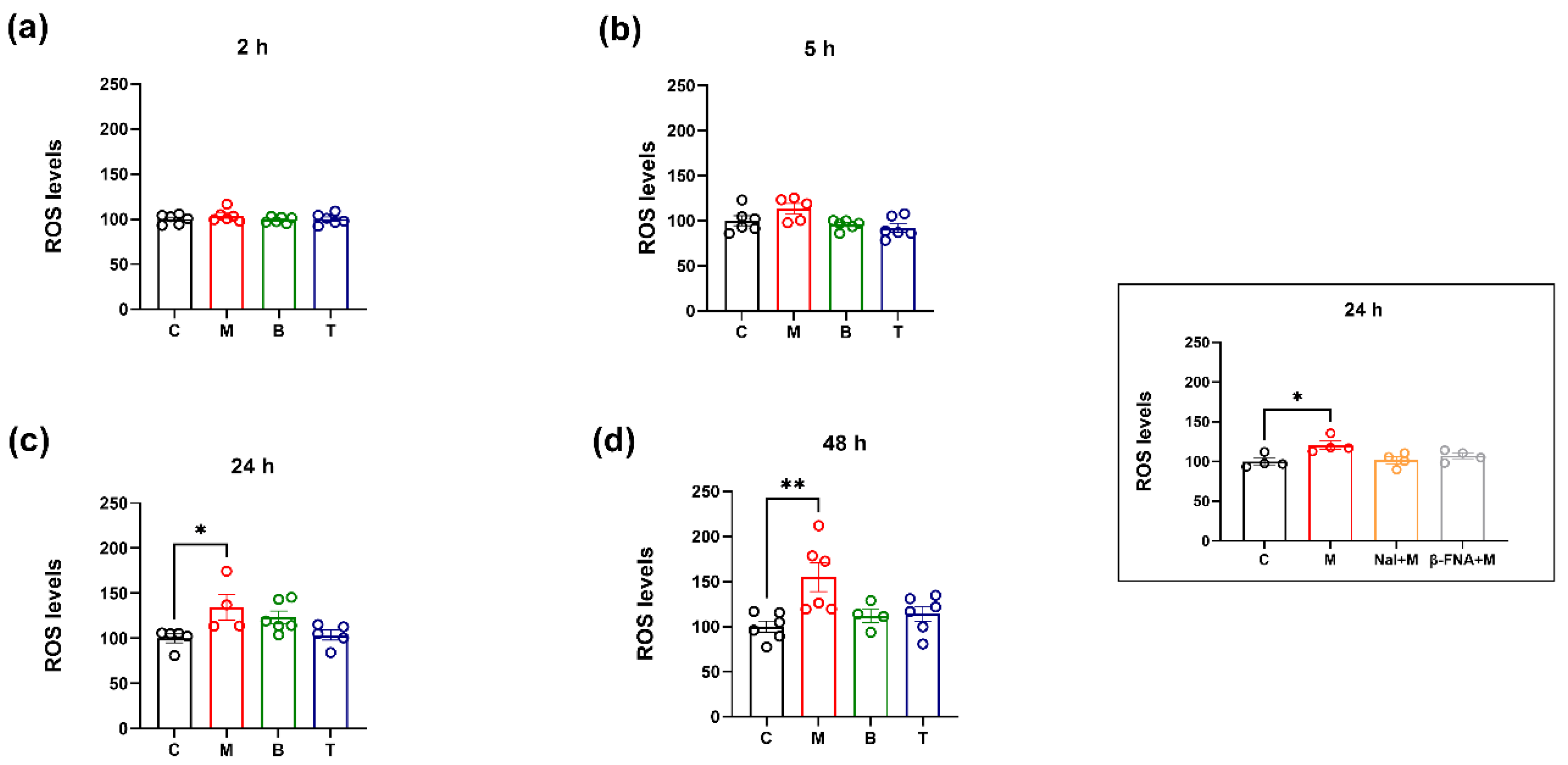{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Results
2.1. Intracellular ROS Levels
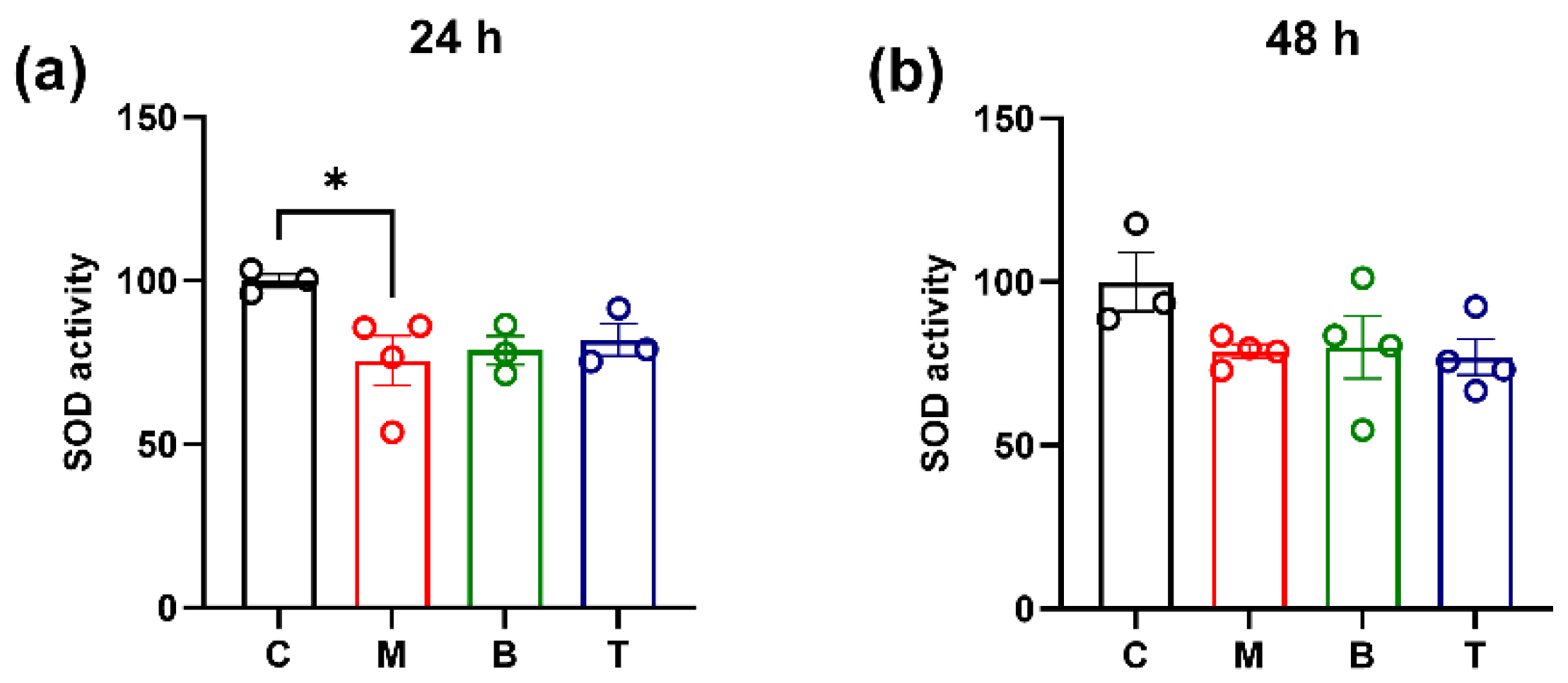
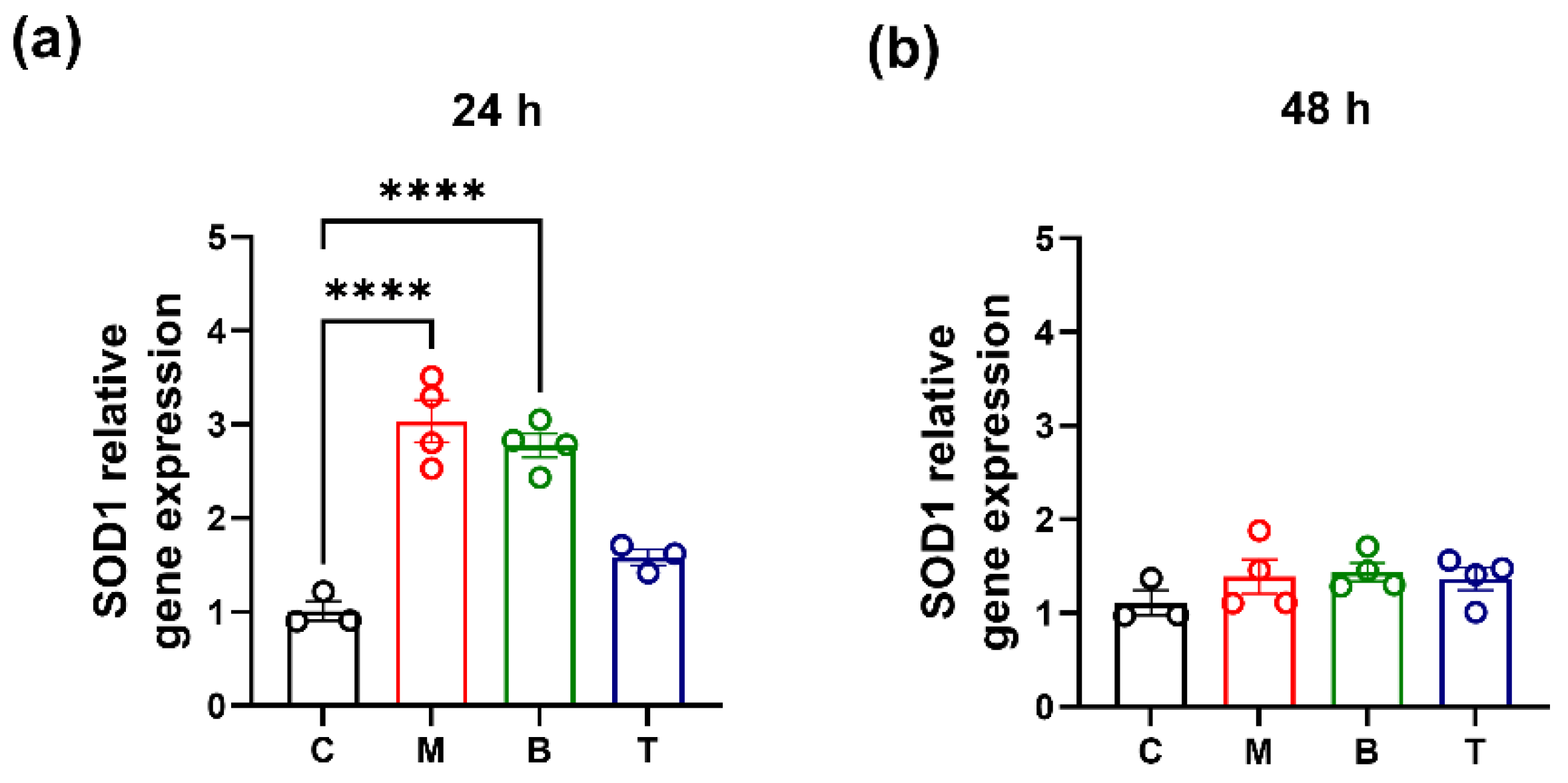
2.2. SOD Activity and Gene Expression
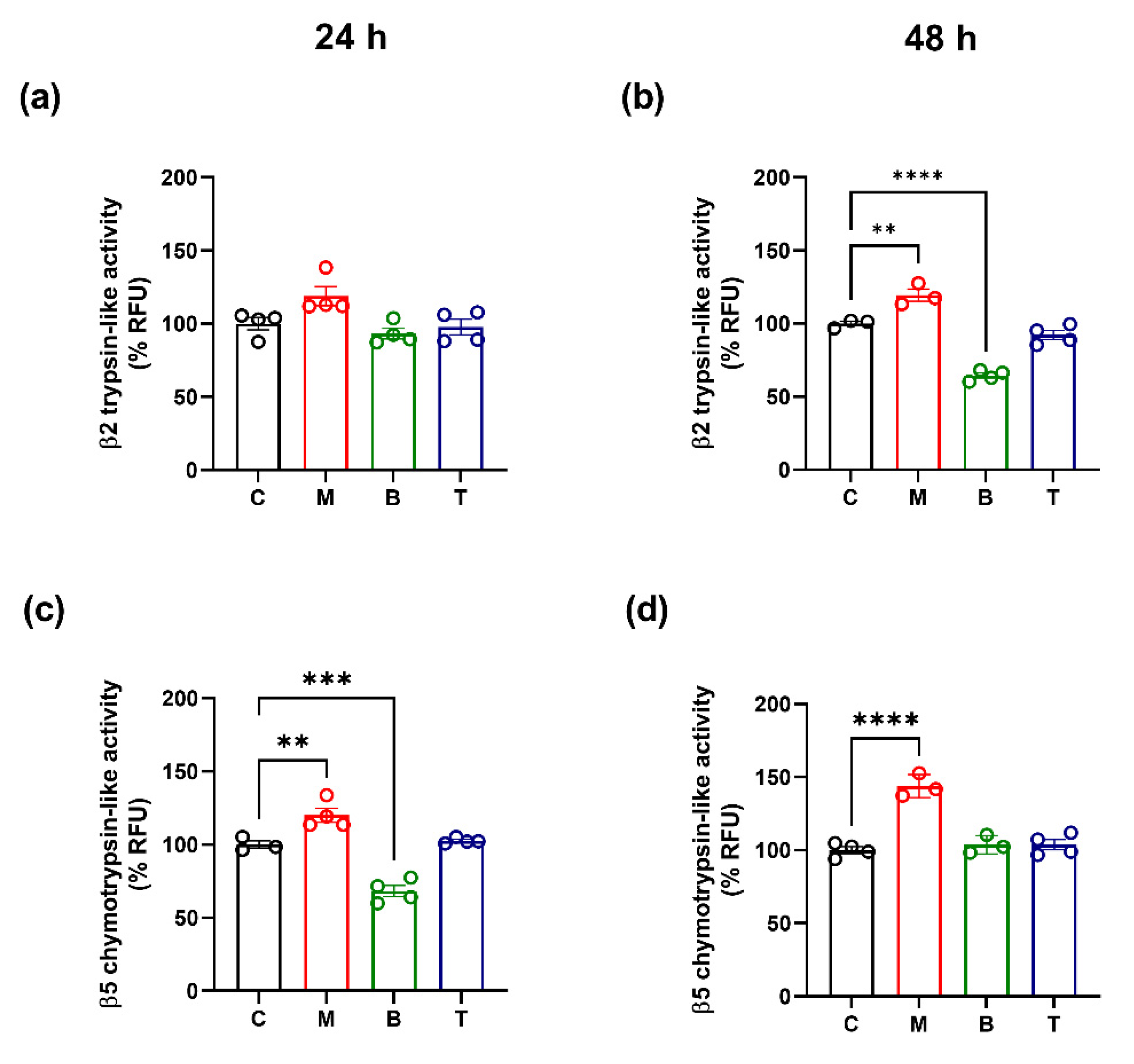
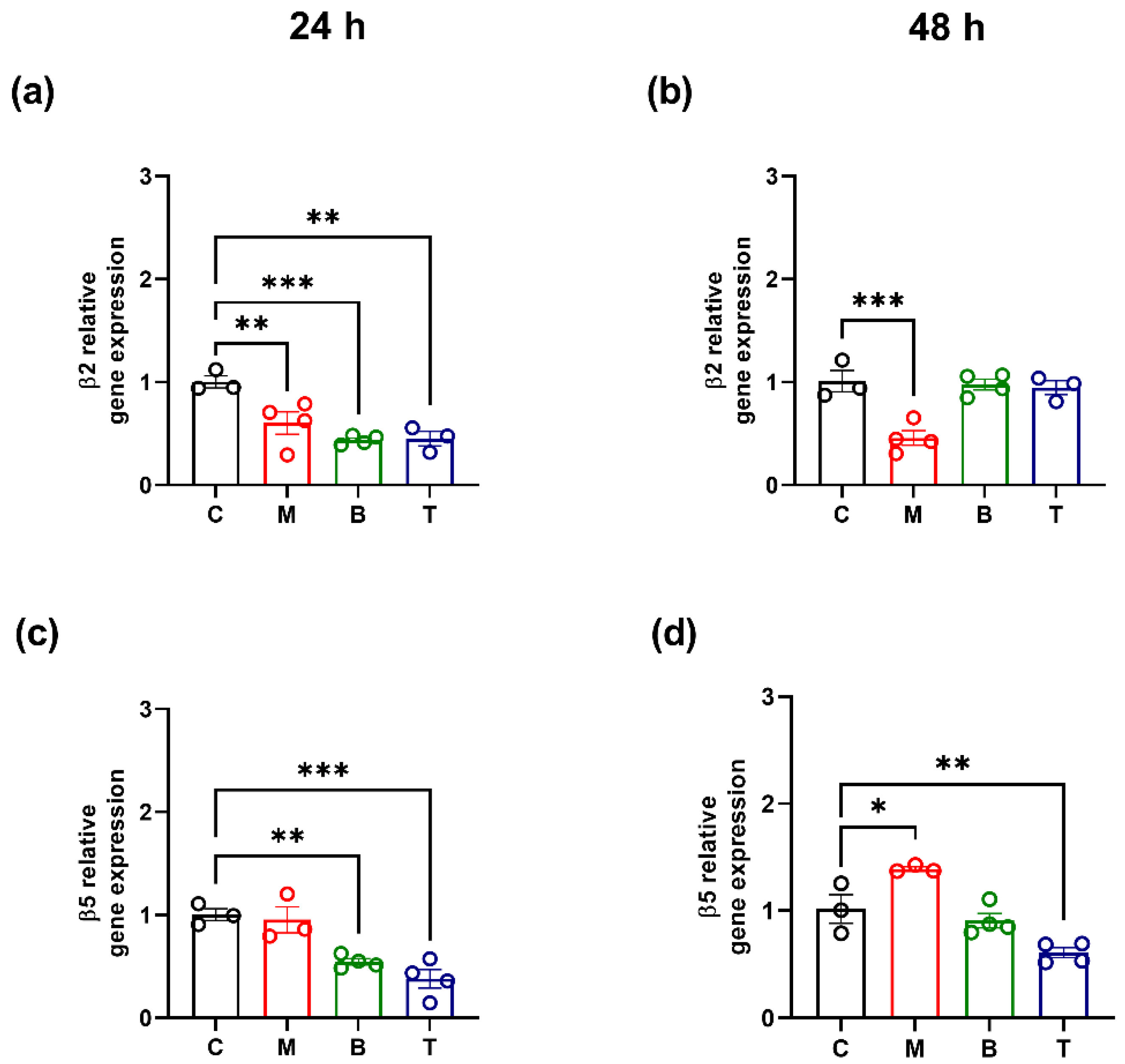
2.3. Proteasome Activity and β2/β5 Subunit Biosynthesis
3. Discussion
4. Materials and Methods
4.1. Cell Culture and Treatments
4.2. Intracellular Reactive Oxygen Species Production
4.3. Protein Extraction
4.4. SOD Activity Assay
4.5. Proteasome Activity Assay
4.6. RNA Isolation and qRT-PCR
4.7. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Sample Availability
References
- Treede, R.D.; Rief, W.; Barke, A.; Aziz, Q.; Bennett, M.I.; Benoliel, R.; Cohen, M.; Evers, S.; Finnerup, N.B.; First, M.B.; et al. Chronic pain as a symptom or a disease: The IASP classification of chronic pain for the international classification of diseases (ICD-11). Pain 2019, 160, 19–27. [Google Scholar] [CrossRef]
- Coluzzi, F.; Rullo, L.; Scerpa, M.S.; Losapio, L.M.; Rocco, M.; Billeci, D.; Candeletti, S.; Romualdi, P. Current and future therapeutic options in pain management: Multi-mechanistic opioids involving both MOR and NOP receptor activation. CNS Drugs 2022, 36, 617–632. [Google Scholar] [CrossRef]
- Chu, L.F.; Clark, D.J.; Angst, M.S. Opioid tolerance and hyperalgesia in chronic pain patients after one month of oral morphine therapy: A preliminary prospective study. J. Pain 2006, 7, 43–48. [Google Scholar] [CrossRef]
- Hutchinson, M.R.; Shavit, Y.; Grace, P.M.; Rice, K.C.; Maier, S.F.; Watkins, L.R. Exploring the neuroimmunopharmacology of opioids: An integrative review of mechanisms of central immune signaling and their implications for opioid analgesia. Pharmacol. Rev. 2011, 63, 772–810. [Google Scholar] [CrossRef]
- Lee, M.; Silverman, S.M.; Hansen, H.; Patel, V.B.; Manchikanti, L. A comprehensive review of opioid-induced hyperalgesia. Pain Physician 2011, 14, 145–161. [Google Scholar] [CrossRef]
- Micheli, L.; Lucarini, E.; Corti, F.; Ciccocioppo, R.; Calò, G.; Rizzi, A.; Ghelardini, C.; Di Cesare Mannelli, L. Involvement of the N/OFQ-NOP system in rat morphine antinociceptive tolerance: Are astrocytes the crossroad? Eur. J. Pharmacol. 2018, 823, 79–86. [Google Scholar] [CrossRef]
- Roeckel, L.-A.; Le Coz, G.-M.; Gavériaux-Ruff, C.; Simonin, F. Opioid-induced hyperalgesia: Cellular and molecular mechanism. Neuroscience 2016, 338, 160–182. [Google Scholar] [CrossRef]
- Cahill, C.M.; Taylor, A.M. Neuroinflammation-a co-occurring phenomenon linking chronic pain and opioid dependence. Curr. Opin. Behav. Sci. 2017, 13, 171–177. [Google Scholar] [CrossRef]
- Reiss, D.; Maduna, T.; Maurin, H.; Audouard, E.; Gaveriaux-Ruff, C. Mu opioid receptor in microglia contributes to morphine analgesic tolerance, hyperalgesia, and withdrawal in mice. J. Neurosci. Res. 2022, 100, 203–219. [Google Scholar] [CrossRef] [PubMed]
- Eidson, L.N.; Murphy, A.Z. Inflammatory mediators of opioid tolerance: Implications for dependency and addiction. Peptides 2019, 115, 51–58. [Google Scholar] [CrossRef]
- Little, J.W.; Doyle, T.; Salvemini, D. Reactive nitroxidative species and nociceptive processing: Determining the roles for nitric oxide, superoxide, and peroxynitrite in pain. Amino Acids 2012, 42, 75–94. [Google Scholar] [CrossRef]
- Skrabalova, J.; Drastichova, Z.; Novotny, J. Morphine as a potential oxidative stress-causing agent. Mini Rev. Org. Chem. 2013, 10, 367–372. [Google Scholar] [CrossRef] [PubMed]
- Reymond, S.; Vujić, T.; Schvartz, D.; Sanchez, J.C. Morphine-induced modulation of Nrf2-antioxidant response element signaling pathway in primary human brain microvascular endothelial cells. Sci. Rep. 2022, 12, 4588. [Google Scholar] [CrossRef] [PubMed]
- Muscoli, C.; Cuzzocrea, S.; Ndengele, M.M.; Mollace, V.; Porreca, F.; Fabrizi, F.; Esposito, E.; Masini, E.; Matuschak, G.M.; Salvemini, D. Therapeutic manipulation of peroxynitrite attenuates the development of opiate-induced antinociceptive tolerance in mice. J. Clin. Investig. 2007, 117, 3530–3539. [Google Scholar] [CrossRef] [PubMed]
- Salvemini, D.; Neumann, W.L. Peroxynitrite: A strategic linchpin of opioid analgesic tolerance. Trends Pharmacol. Sci. 2009, 30, 194–202. [Google Scholar] [CrossRef]
- Tobore, T.O. Towards a comprehensive theory of non-cancer acute and chronic pain management: The critical role of reactive oxygen and nitrogen species in pain, and opioid dependence, addiction, hyperalgesia, and tolerance. Adv. Redox Res. 2021, 2, 100003. [Google Scholar] [CrossRef]
- Schieber, M.; Chandel, N.S. ROS function in redox signaling and oxidative stress. Curr. Biol. 2014, 24, R453–R462. [Google Scholar] [CrossRef]
- Lefaki, M.; Papaevgeniou, N.; Chondrogianni, N. Redox regulation of proteasome function. Redox Biol. 2017, 13, 452–458. [Google Scholar] [CrossRef]
- Kiffin, R.; Christian, C.; Knecht, E.; Cuervo, A.M. Activation of chaperone-mediated autophagy during oxidative stress. Mol. Biol. Cell 2004, 15, 4829–4840. [Google Scholar] [CrossRef]
- Ding, Q.; Dimayuga, E.; Martin, S.; Bruce-Keller, A.J.; Nukala, V.; Cuervo, A.M.; Keller, J.N. Characterization of chronic low-level proteasome inhibition on neural homeostasis. J. Neurochem. 2003, 86, 489–497. [Google Scholar] [CrossRef]
- Grune, T.; Jung, T.; Merker, K.; Davies, K.J. Decreased proteolysis caused by protein aggregates, inclusion bodies, plaques, lipofuscin, ceroid, and ‘aggresomes’ during oxidative stress, aging, and disease. Int. J. Biochem. Cell Biol. 2004, 36, 2519–2530. [Google Scholar] [CrossRef] [PubMed]
- Aiken, C.T.; Kaake, R.M.; Wang, X.; Huang, L. Oxidative stress-mediated regulation of proteasome complexes. Mol. Cell Proteom. 2011, 10, R110.006924. [Google Scholar] [CrossRef] [PubMed]
- Tanaka, K. The proteasome: Overview of structure and functions. Proc. Jpn. Acad. Ser. B Phys. Biol. Sci. 2009, 85, 12–36. [Google Scholar] [CrossRef]
- Caputi, F.F.; Rullo, L.; Stamatakos, S.; Candeletti, S.; Romualdi, P. Interplay between the endogenous opioid system and proteasome complex: Beyond signaling. Int. J. Mol. Sci. 2019, 20, 1441. [Google Scholar] [CrossRef]
- Dahlmann, B. Role of proteasomes in disease. BMC Biochem. 2007, 8 (Suppl. 1), S3. [Google Scholar] [CrossRef] [PubMed]
- Coux, O.; Zieba, B.A.; Meiners, S. The proteasome system in health and disease. Adv. Exp. Med. Biol. 2020, 1233, 55–100. [Google Scholar] [CrossRef] [PubMed]
- Massaly, N.; Francès, B.; Moulédous, L. Roles of the ubiquitin proteasome system in the effects of drugs of abuse. Front. Mol. Neurosci. 2015, 7, 99. [Google Scholar] [CrossRef]
- Moss, A.; Blackburn-Munro, G.; Garry, E.M.; Blakemore, J.A.; Dickinson, T.; Rosie, R.; Mitchell, R.; Fleetwood-Walker, S.M. A role of the ubiquitin-proteasome system in neuropathic pain. J. Neurosci. 2002, 22, 1363–1372. [Google Scholar] [CrossRef] [PubMed]
- Ossipov, M.H.; Bazov, I.; Gardell, L.R.; Kowal, J.; Yakovleva, T.; Usynin, I.; Ekström, T.J.; Porreca, F.; Bakalkin, G. Control of chronic pain by the ubiquitin proteasome system in the spinal cord. J. Neurosci. 2007, 27, 8226–8237. [Google Scholar] [CrossRef]
- Massaly, N.; Dahan, L.; Baudonnat, M.; Hovnanian, C.; Rekik, K.; Solinas, M.; David, V.; Pech, S.; Zajac, J.M.; Roullet, P.; et al. Involvement of protein degradation by the ubiquitin proteasome system in opiate addictive behaviors. Neuropsychopharmacology 2013, 38, 596–604. [Google Scholar] [CrossRef]
- Yang, L.; Wang, S.; Lim, G.; Sung, B.; Zeng, Q.; Mao, J. Inhibition of the ubiquitin-proteasome activity prevents glutamate transporter degradation and morphine tolerance. Pain 2008, 140, 472–478. [Google Scholar] [CrossRef]
- Wang, Y.; Branicky, R.; Noë, A.; Hekimi, S. Superoxide dismutases: Dual roles in controlling ROS damage and regulating ROS signaling. J. Cell Biol. 2018, 217, 1915–1928. [Google Scholar] [CrossRef]
- Ma, J.; Yuan, X.; Qu, H.; Zhang, J.; Wang, D.; Sun, X.; Zheng, Q. The role of reactive oxygen species in morphine addiction of SH-SY5Y cells. Life Sci. 2015, 124, 128–135. [Google Scholar] [CrossRef]
- Halcrow, P.W.; Kumar, N.; Hao, E.; Khan, N.; Meucci, O.; Geiger, J.D. Mu opioid receptor-mediated release of endolysosome iron increases levels of mitochondrial iron, reactive oxygen species, and cell death. Neuroim. Pharmacol. Ther. 2022, published online 14 September. [Google Scholar] [CrossRef]
- Dickinson, B.C.; Chang, C.J. Chemistry and biology of reactive oxygen species in signaling or stress responses. Nat. Chem. Biol. 2011, 7, 504–511. [Google Scholar] [CrossRef]
- Schattauer, S.S.; Land, B.B.; Reichard, K.L.; Abraham, A.D.; Burgeno, L.M.; Kuhar, J.R.; Phillips, P.E.M.; Ong, S.E.; Chavkin, C. Peroxiredoxin 6 mediates Gαi protein-coupled receptor inactivation by cJun kinase. Nat. Commun. 2017, 8, 743. [Google Scholar] [CrossRef]
- Caputi, F.F.; Acquas, E.; Kasture, S.; Ruiu, S.; Candeletti, S.; Romualdi, P. The standardized Withania somnifera Dunal root extract alters basal and morphine-induced opioid receptor gene expression changes in neuroblastoma cells. BMC Complement Altern. Med. 2018, 18, 9. [Google Scholar] [CrossRef]
- Doyle, T.; Bryant, L.; Muscoli, C.; Cuzzocrea, S.; Esposito, E.; Chen, Z.; Salvemini, D. Spinal NADPH oxidase is a source of superoxide in the development of morphine-induced hyperalgesia and antinociceptive tolerance. Neurosci. Lett. 2010, 483, 85–89. [Google Scholar] [CrossRef]
- Doyle, T.; Esposito, E.; Bryant, L.; Cuzzocrea, S.; Salvemini, D. NADPH-oxidase 2 activation promotes opioid-induced antinociceptive tolerance in mice. Neuroscience 2013, 241, 1–9. [Google Scholar] [CrossRef]
- Bu, Q.; Yang, Y.; Yan, G.; Hu, Z.; Hu, C.; Duan, J.; Lv, L.; Zhou, J.; Zhao, J.; Shao, X.; et al. Proteomic analysis of the nucleus accumbens in rhesus monkeys of morphine dependence and withdrawal intervention. J. Proteom. 2012, 75, 1330–1342. [Google Scholar] [CrossRef]
- Saify, K.; Saadat, I.; Saadat, M. Down-regulation of antioxidant genes in human SH-SY5Y cells after treatment with morphine. Life Sci. 2016, 144, 26–29. [Google Scholar] [CrossRef] [PubMed]
- Zhou, J.; Li, Y.; Yan, G.; Bu, Q.; Lv, L.; Yang, Y.; Zhao, J.; Shao, X.; Deng, Y.; Zhu, R.; et al. Protective role of taurine against morphine-induced neurotoxicity in C6 cells via inhibition of oxidative stress. Neurotox. Res. 2011, 20, 334–342. [Google Scholar] [CrossRef] [PubMed]
- Caputi, F.F.; Rullo, L.; Acquas, E.; Ciccocioppo, R.; Candeletti, S.; Romualdi, P. Evidence of a PPARγ-mediated mechanism in the ability of Withania somnifera to attenuate tolerance to the antinociceptive effects of morphine. Pharmacol. Res. 2019, 139, 422–430. [Google Scholar] [CrossRef] [PubMed]
- Ghavimi, H.; Charkhpour, M.; Ghasemi, S.; Mesgari, M.; Hamishehkar, H.; Hassanzadeh, K.; Arami, S.; Hassanzadeh, K. Pioglitazone prevents morphine antinociceptive tolerance via ameliorating neuroinflammation in rat cerebral cortex. Pharmacol. Rep. 2015, 67, 78–84. [Google Scholar] [CrossRef]
- Janes, K.; Neumann, W.L.; Salvemini, D. Anti-superoxide and anti-peroxynitrite strategies in pain suppression. Biochim. Biophys. Acta 2012, 1822, 815–821. [Google Scholar] [CrossRef]
- Zeng, X.S.; Geng, W.S.; Wang, Z.Q.; Jia, J.J. Morphine Addiction and Oxidative Stress: The Potential Effects of Thioredoxin-1. Front. Pharmacol. 2020, 11, 82. [Google Scholar] [CrossRef]
- Yang, L.; Wang, S.; Sung, B.; Lim, G.; Mao, J. Morphine induces ubiquitin-proteasome activity and glutamate transporter degradation. J. Biol. Chem. 2008, 283, 21703–21713. [Google Scholar] [CrossRef]
- Chondrogianni, N.; Tzavelas, C.; Pemberton, A.J.; Nezis, I.P.; Rivett, A.J.; Gonos, E.S. Overexpression of proteasome beta5 assembled subunit increases the amount of proteasome and confers ameliorated response to oxidative stress and higher survival rates. J. Biol. Chem. 2005, 280, 11840–11850. [Google Scholar] [CrossRef]
- Manandhar, P.; Connor, M.; Santiago, M. Tapentadol shows lower intrinsic efficacy at µ receptor than morphine and oxycodone. Pharmacol. Res. Perspect. 2022, 10, e00921. [Google Scholar] [CrossRef]
- Morgan, D.; Cook, C.D.; Smith, M.A.; Picker, M.J. An examination of the interactions between the antinociceptive effects of morphine and various mu-opioids: The role of intrinsic efficacy and stimulus intensity. Anesth. Analg. 1999, 88, 407–413. [Google Scholar] [CrossRef]
- Chaturvedi, K.; Bandari, P.; Chinen, N.; Howells, R.D. Proteasome involvement in agonist-induced down-regulation of mu and delta opioid receptors. J. Biol. Chem. 2001, 276, 12345–12355. [Google Scholar] [CrossRef]
- Moulédous, L.; Neasta, J.; Uttenweiler-Joseph, S.; Stella, A.; Matondo, M.; Corbani, M.; Monsarrat, B.; Meunier, J.C. Long-term morphine treatment enhances proteasome-dependent degradation of G beta in human neuroblastoma SH-SY5Y cells: Correlation with onset of adenylate cyclase sensitization. Mol. Pharmacol. 2005, 68, 467–476. [Google Scholar] [CrossRef]
- Almeida, M.B.; Costa-Malaquias, A.; Nascimento, J.L.; Oliveira, K.R.; Herculano, A.M.; Crespo-López, M.E. Therapeutic concentration of morphine reduces oxidative stress in glioma cell line. Braz. J. Med. Biol. Res. 2014, 47, 398–402. [Google Scholar] [CrossRef]
- Elyasi, L.; Eftekhar-Vaghefi, S.H.; Asadi-Shekaari, M.; Esmaeili-Mahani, S. Induction of cross-tolerance between protective effect of morphine and nicotine in 6-hydroxydopamine-induce neurotoxicity in SH-SY5Y human dopaminergic neuroblastoma cells. Int. J. Neurosci. 2019, 129, 129–138. [Google Scholar] [CrossRef]
- Gülçin, I.; Beydemir, S.; Alici, H.A.; Elmastaş, M.; Büyükokuroğlu, M.E. In vitro antioxidant properties of morphine. Pharmacol. Res. 2004, 49, 59–66. [Google Scholar] [CrossRef] [PubMed]
- Bhat, R.S.; Bhaskaran, M.; Mongia, A.; Hitosugi, N.; Singhal, P.C. Morphine-induced macrophage apoptosis: Oxidative stress and strategies for modulation. J. Leukoc. Biol. 2004, 75, 1131–1138. [Google Scholar] [CrossRef]
- Hemati, K.; Pourhanifeh, M.H.; Dehdashtian, E.; Fatemi, I.; Mehrzadi, S.; Reiter, R.J.; Hosseinzadeh, A. Melatonin and morphine: Potential beneficial effects of co-use. Fundam. Clin. Pharmacol. 2021, 35, 25–39. [Google Scholar] [CrossRef]
- Lauro, F.; Giancotti, L.A.; Ilari, S.; Dagostino, C.; Gliozzi, M.; Morabito, C.; Malafoglia, V.; Raffaeli, W.; Muraca, M.; Goffredo, B.M.; et al. Inhibition of Spinal Oxidative Stress by Bergamot Polyphenolic Fraction Attenuates the Development of Morphine Induced Tolerance and Hyperalgesia in Mice. PLoS ONE 2016, 11, e0156039. [Google Scholar] [CrossRef]
- Ndengele, M.M.; Cuzzocrea, S.; Masini, E.; Vinci, M.C.; Esposito, E.; Muscoli, C.; Petrusca, D.N.; Mollace, V.; Mazzon, E.; Li, D.; et al. Spinal ceramide modulates the development of morphine antinociceptive tolerance via peroxynitrite-mediated nitroxidative stress and neuroimmune activation. J. Pharmacol. Exp. Ther. 2009, 329, 64–75. [Google Scholar] [CrossRef]
- Raghavendra, V.; Kulkarni, S.K. Possible mechanisms of action in melatonin reversal of morphine tolerance and dependence in mice. Eur. J. Pharmacol. 2000, 409, 279–289. [Google Scholar] [CrossRef]
- Caputi, F.F.; Carretta, D.; Tzschentke, T.M.; Candeletti, S.; Romualdi, P. Opioid receptor gene expression in human neuroblastoma SH-SY5Y cells following tapentadol exposure. J. Mol. Neurosci. 2014, 53, 669–676. [Google Scholar] [CrossRef]
- Caputi, F.F.; Lattanzio, F.; Carretta, D.; Mercatelli, D.; Candeletti, S.; Romualdi, P. Morphine and fentanyl differently affect MOP and NOP gene expression in human neuroblastoma SH-SY5Y cells. J. Mol. Neurosci. 2013, 51, 532–538. [Google Scholar] [CrossRef] [PubMed]
- Khanna, I.K.; Pillarisetti, S. Buprenorphine—An attractive opioid with underutilized potential in treatment of chronic pain. J. Pain Res. 2015, 8, 859–870. [Google Scholar] [CrossRef] [PubMed]
- Vivarelli, F.; Canistro, D.; Cirillo, S.; Papi, A.; Spisni, E.; Vornoli, A.; Croce, C.M.D.; Longo, V.; Franchi, P.; Filippi, S.; et al. Co-carcinogenic effects of vitamin E in prostate. Sci. Rep. 2019, 9, 11636. [Google Scholar] [CrossRef] [PubMed]
- Caputi, F.F.; Carboni, L.; Mazza, D.; Candeletti, S.; Romualdi, P. Cocaine and ethanol target 26S proteasome activity and gene expression in neuroblastoma cells. Drug Alcohol Depend. 2016, 161, 265–275. [Google Scholar] [CrossRef] [PubMed]
- Costa, G.; Caputi, F.F.; Serra, M.; Simola, N.; Rullo, L.; Stamatakos, S.; Sanna, F.; Germain, M.; Martinoli, M.G.; Candeletti, S.; et al. Activation of Antioxidant and Proteolytic Pathways in the Nigrostriatal Dopaminergic System After 3,4-Methylenedioxymethamphetamine Administration: Sex-Related Differences. Front. Pharmacol. 2021, 12, 713486. [Google Scholar] [CrossRef] [PubMed]
- Caputi, F.F.; Di Cesare Mannelli, L.; Rullo, L.; Micheli, L.; Stamatakos, S.; Posa, L.; Ghelardini, C.; Romualdi, P.; Candeletti, S. The active second-generation proteasome inhibitor oprozomib reverts the oxaliplatin-induced neuropathy symptoms. Biochem. Pharmacol. 2020, 182, 114255. [Google Scholar] [CrossRef]
- Chomczynski, P.; Sacchi, N. The single-step method of RNA isolation by acid guanidinium thiocyanate-phenol-chloroform extraction: Twenty-something years on. Nat. Protoc. 2006, 1, 581–585. [Google Scholar] [CrossRef]
- Caputi, F.F.; Di Benedetto, M.; Carretta, D.; Bastias del Carmen Candia, S.; D’Addario, C.; Cavina, C.; Candeletti, S.; Romualdi, P. Dynorphin/KOP and nociceptin/NOP gene expression and epigenetic changes by cocaine in rat striatum and nucleus accumbens. Prog. Neuropsychopharmacol. Biol. Psychiatry 2014, 49, 36–46. [Google Scholar] [CrossRef]
- Caputi, F.F.; Stopponi, S.; Rullo, L.; Palmisano, M.; Ubaldi, M.; Candeletti, S.; Ciccocioppo, R.; Romualdi, P. Dysregulation of Nociceptin/Orphanin FQ and Dynorphin Systems in the Extended Amygdala of Alcohol Preferring Marchigian Sardinian (msP) Rats. Int. J. Mol. Sci. 2021, 22, 2448. [Google Scholar] [CrossRef]





Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rullo, L.; Caputi, F.F.; Losapio, L.M.; Morosini, C.; Posa, L.; Canistro, D.; Vivarelli, F.; Romualdi, P.; Candeletti, S. Effects of Different Opioid Drugs on Oxidative Status and Proteasome Activity in SH-SY5Y Cells. Molecules 2022, 27, 8321. https://doi.org/10.3390/molecules27238321
Rullo L, Caputi FF, Losapio LM, Morosini C, Posa L, Canistro D, Vivarelli F, Romualdi P, Candeletti S. Effects of Different Opioid Drugs on Oxidative Status and Proteasome Activity in SH-SY5Y Cells. Molecules. 2022; 27(23):8321. https://doi.org/10.3390/molecules27238321
Chicago/Turabian StyleRullo, Laura, Francesca Felicia Caputi, Loredana Maria Losapio, Camilla Morosini, Luca Posa, Donatella Canistro, Fabio Vivarelli, Patrizia Romualdi, and Sanzio Candeletti. 2022. "Effects of Different Opioid Drugs on Oxidative Status and Proteasome Activity in SH-SY5Y Cells" Molecules 27, no. 23: 8321. https://doi.org/10.3390/molecules27238321
APA StyleRullo, L., Caputi, F. F., Losapio, L. M., Morosini, C., Posa, L., Canistro, D., Vivarelli, F., Romualdi, P., & Candeletti, S. (2022). Effects of Different Opioid Drugs on Oxidative Status and Proteasome Activity in SH-SY5Y Cells. Molecules, 27(23), 8321. https://doi.org/10.3390/molecules27238321