
Polyphenols for Livestock Feed: Sustainable Perspectives for Animal Husbandry?

 ,
,  ,
,  and
and 
Abstract

1. Introduction
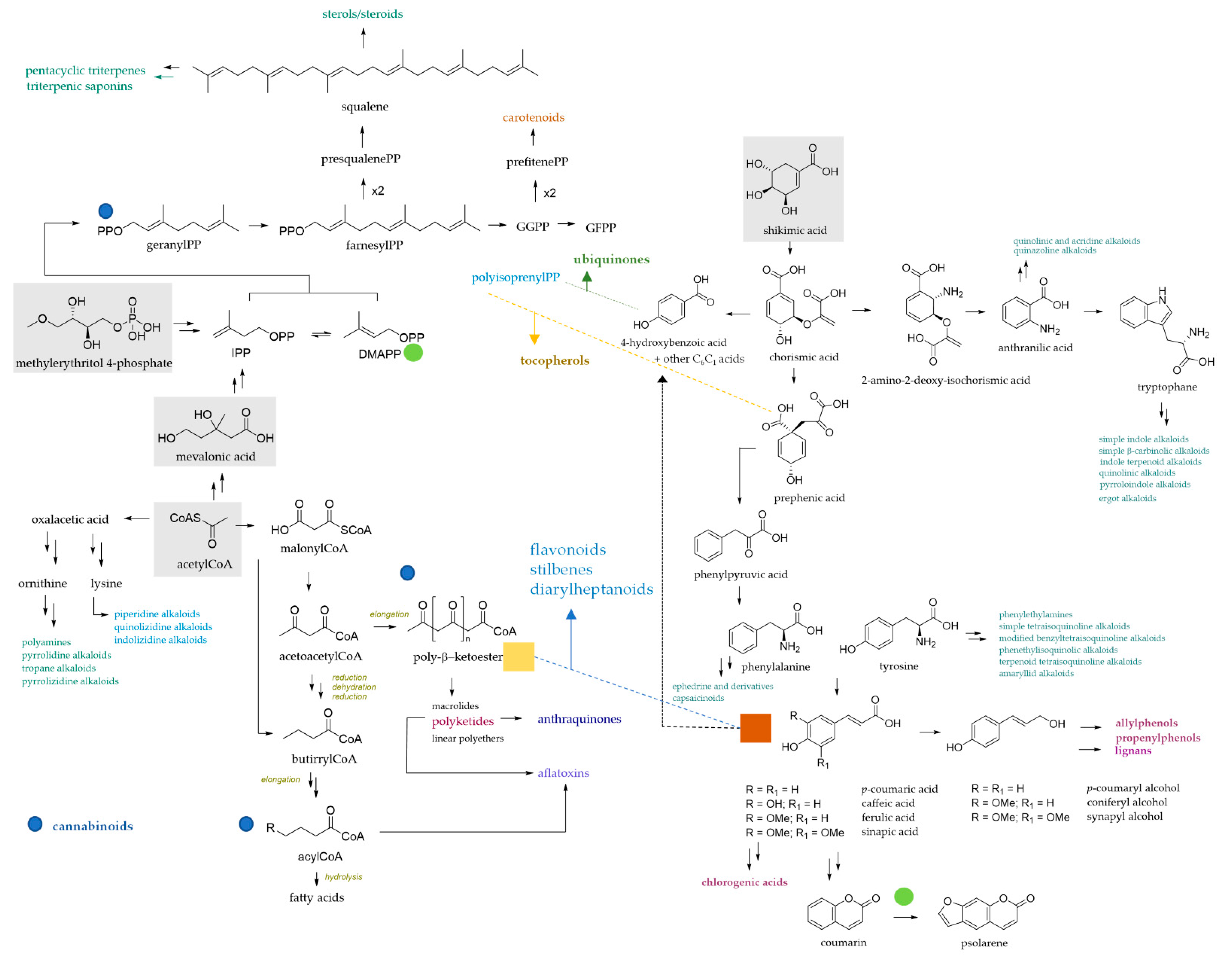
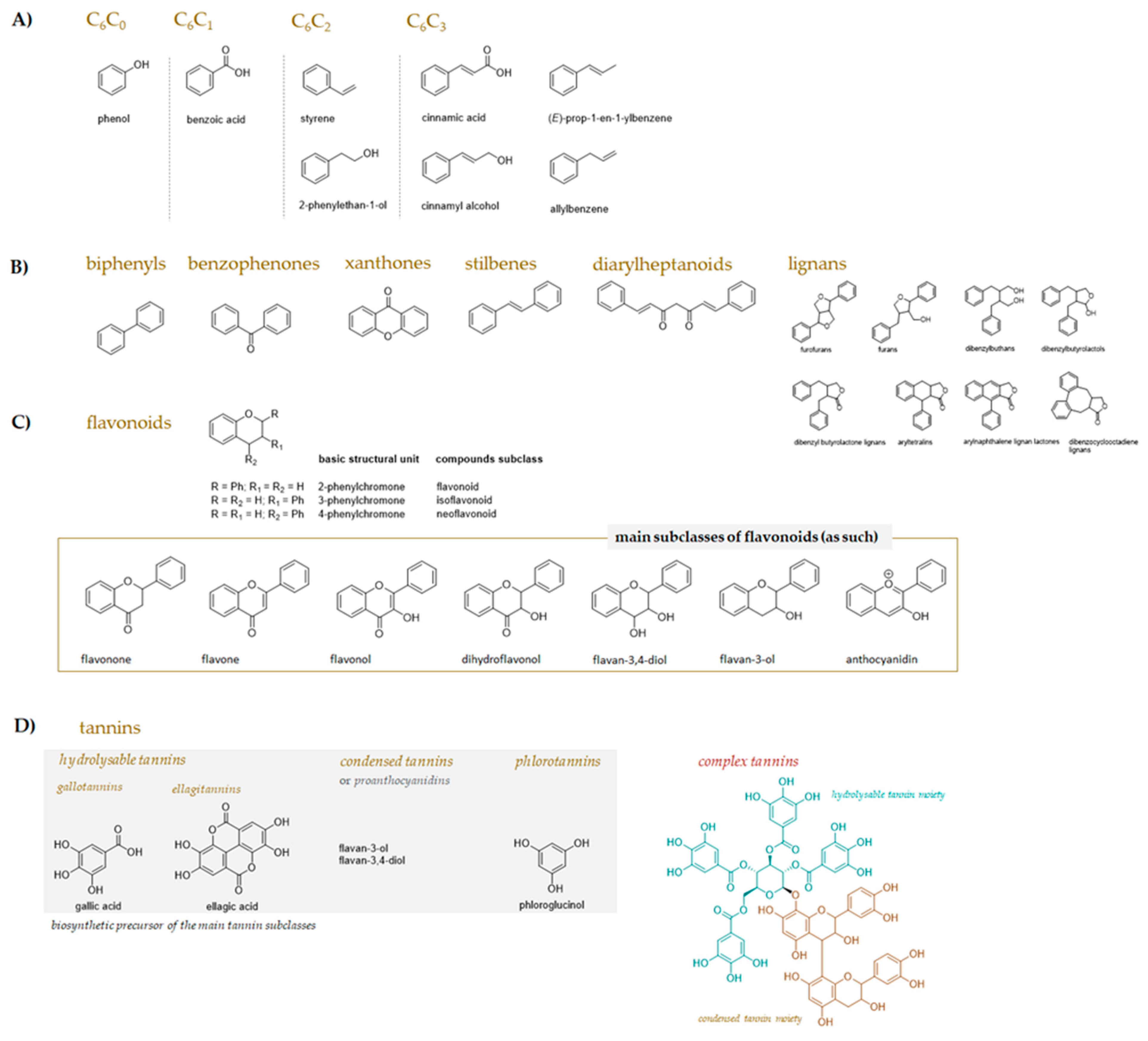
2. Chemical Complexity of Bioactive Plant Compounds Used as Livestock Feed
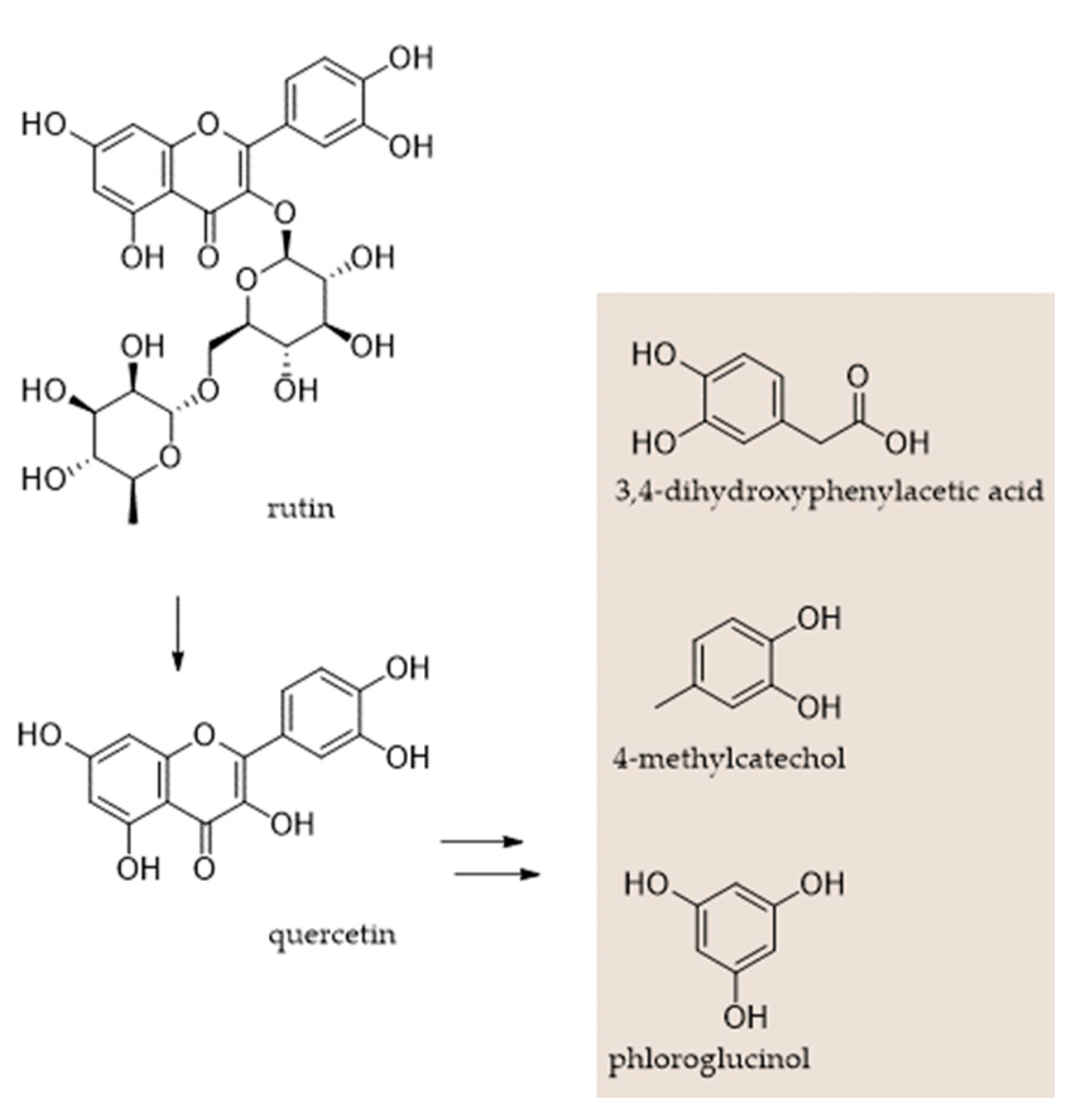
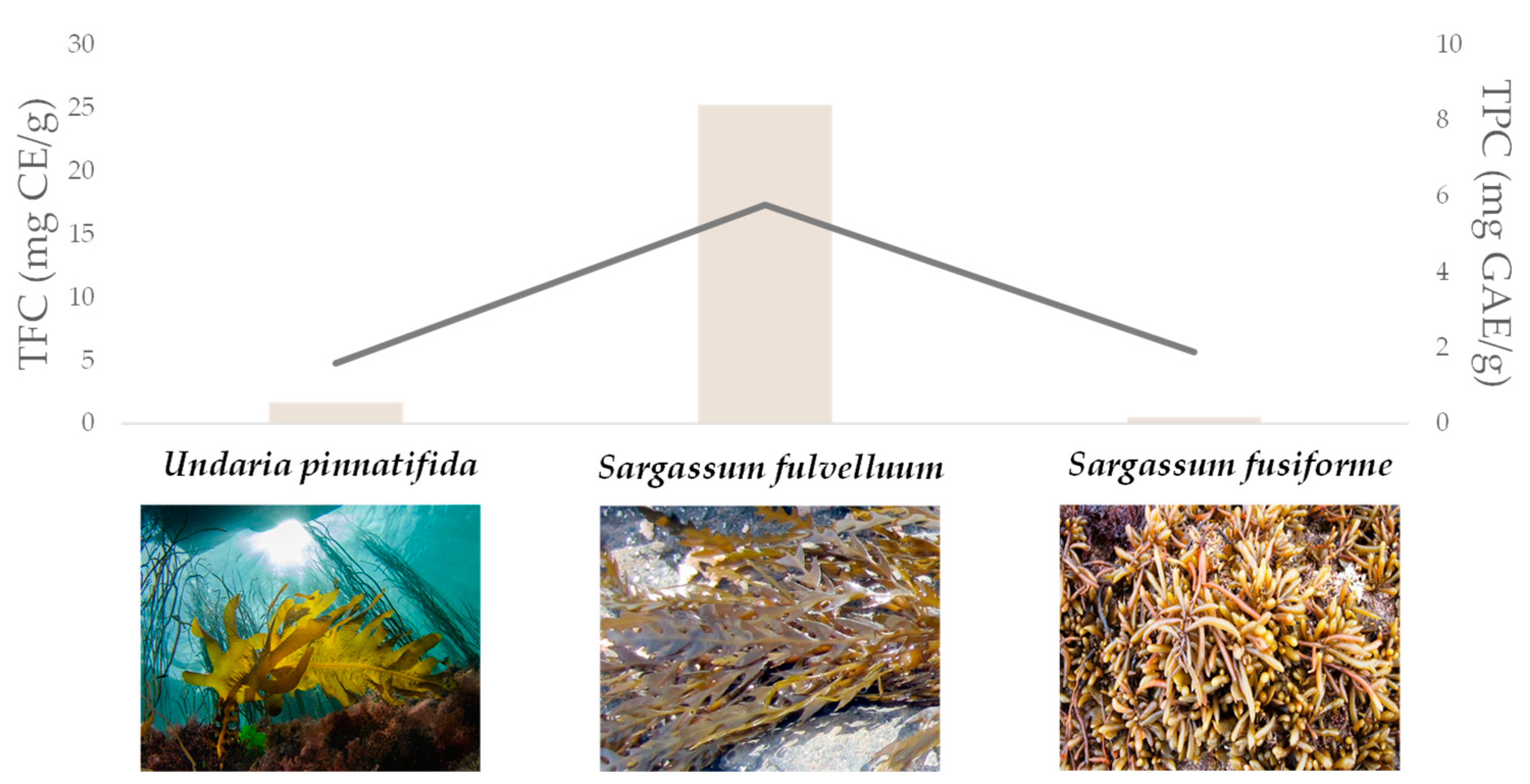
Multiple Approaches for (Poly)phenol Chemical Identification in Plant Extracts and Beyond
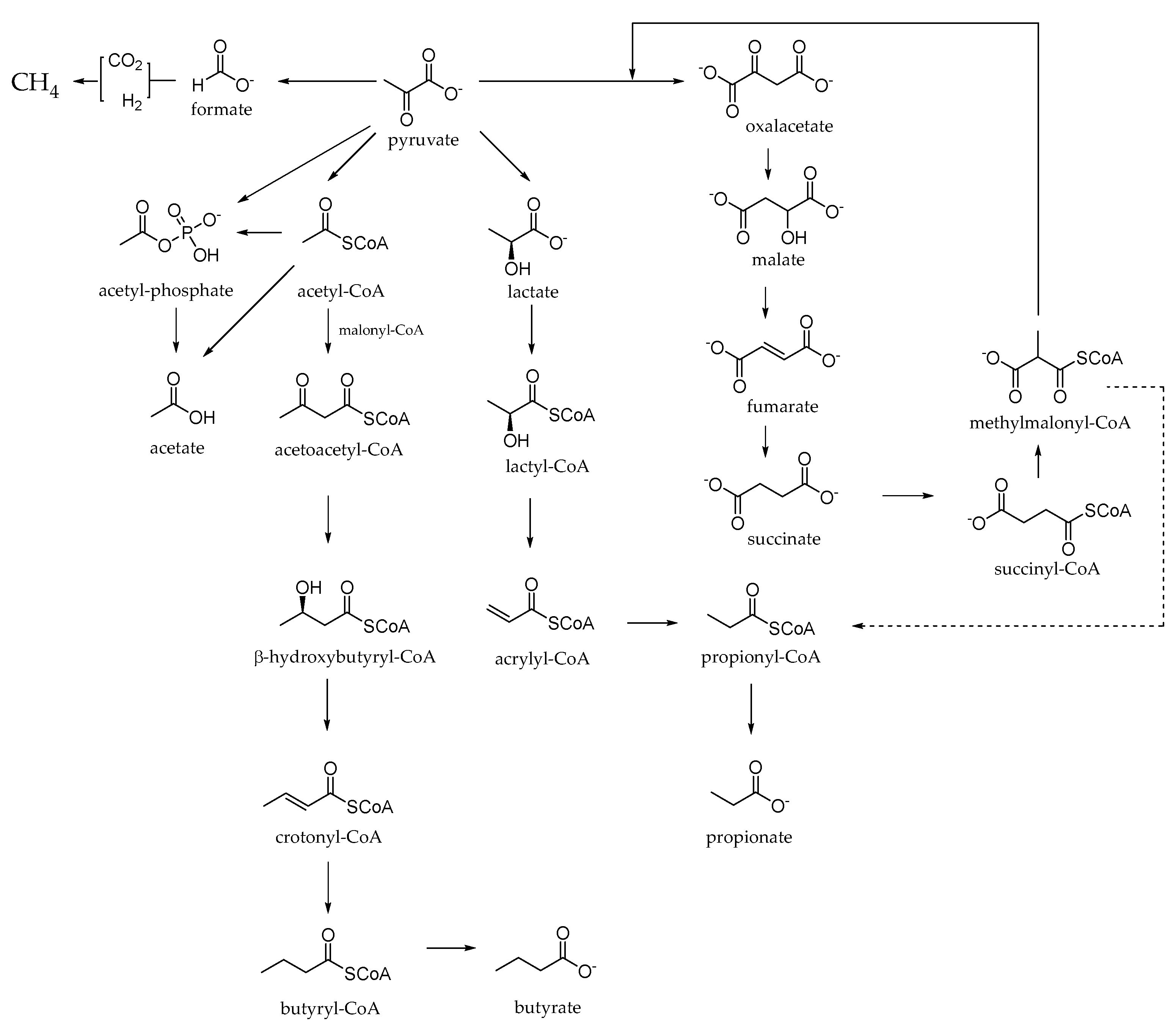
3. Focus on Rumen Fermentation: A Key Step to Unravel Polyphenol Efficacy
3.1. Rumen Microbiota Composition
3.2. Effect of Polyphenols on Rumen Fermentation Parameters
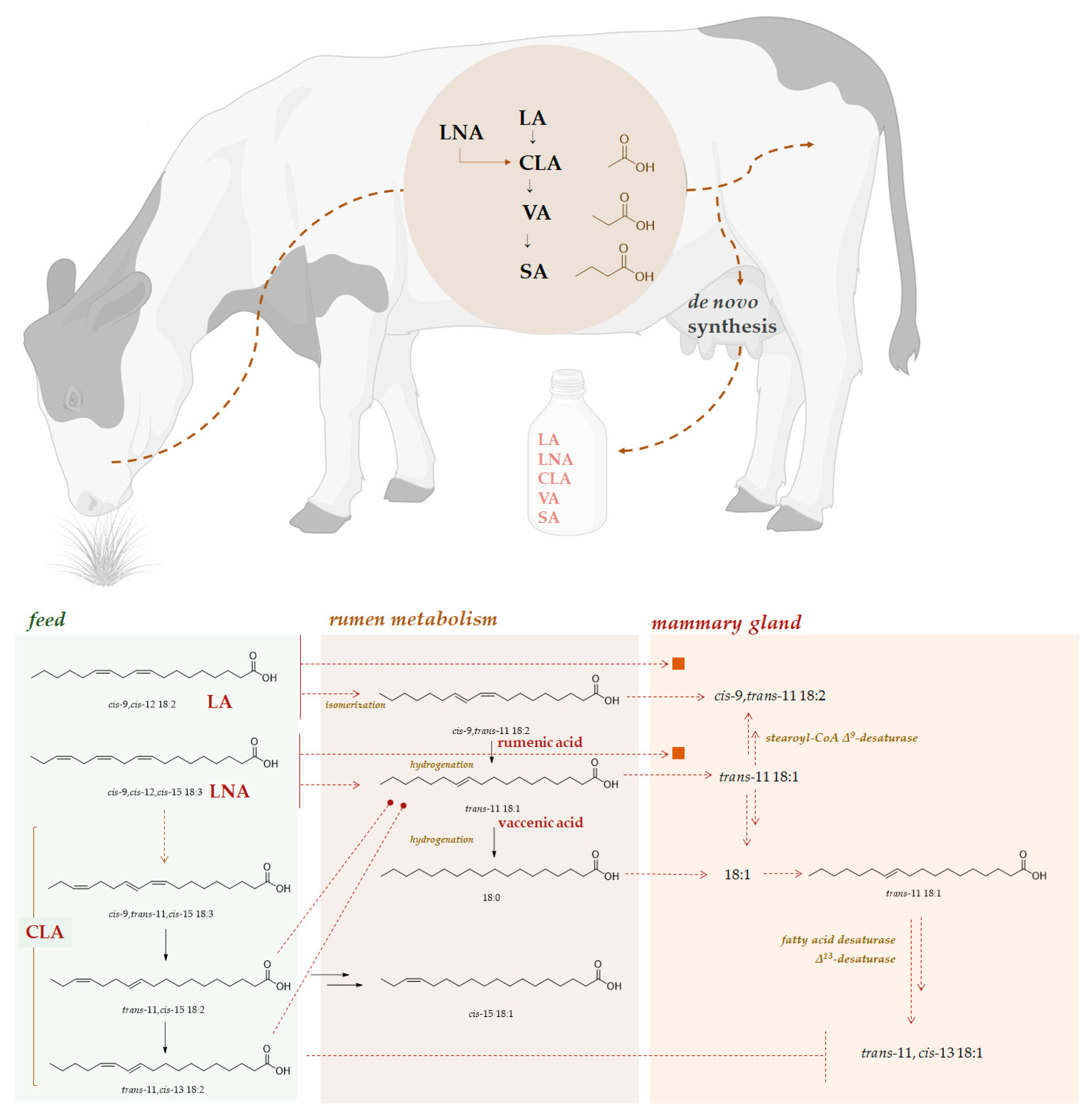
4. Biohydrogenation (BH) Process and Effects of Polyphenols on Lipidic Profile
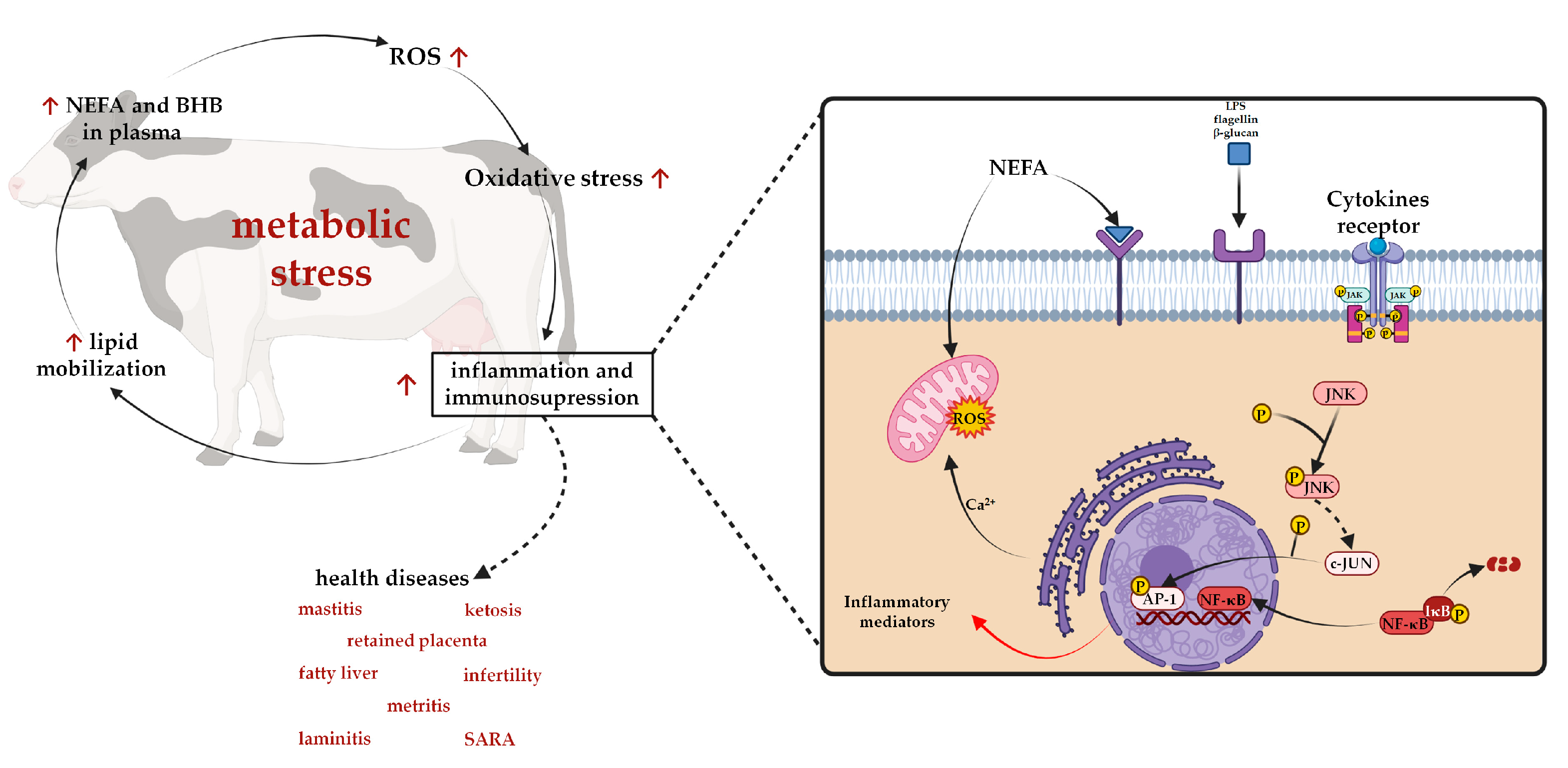
5. Effects of Polyphenol Supplementation on Milk Yield and Composition in Dairy Cows
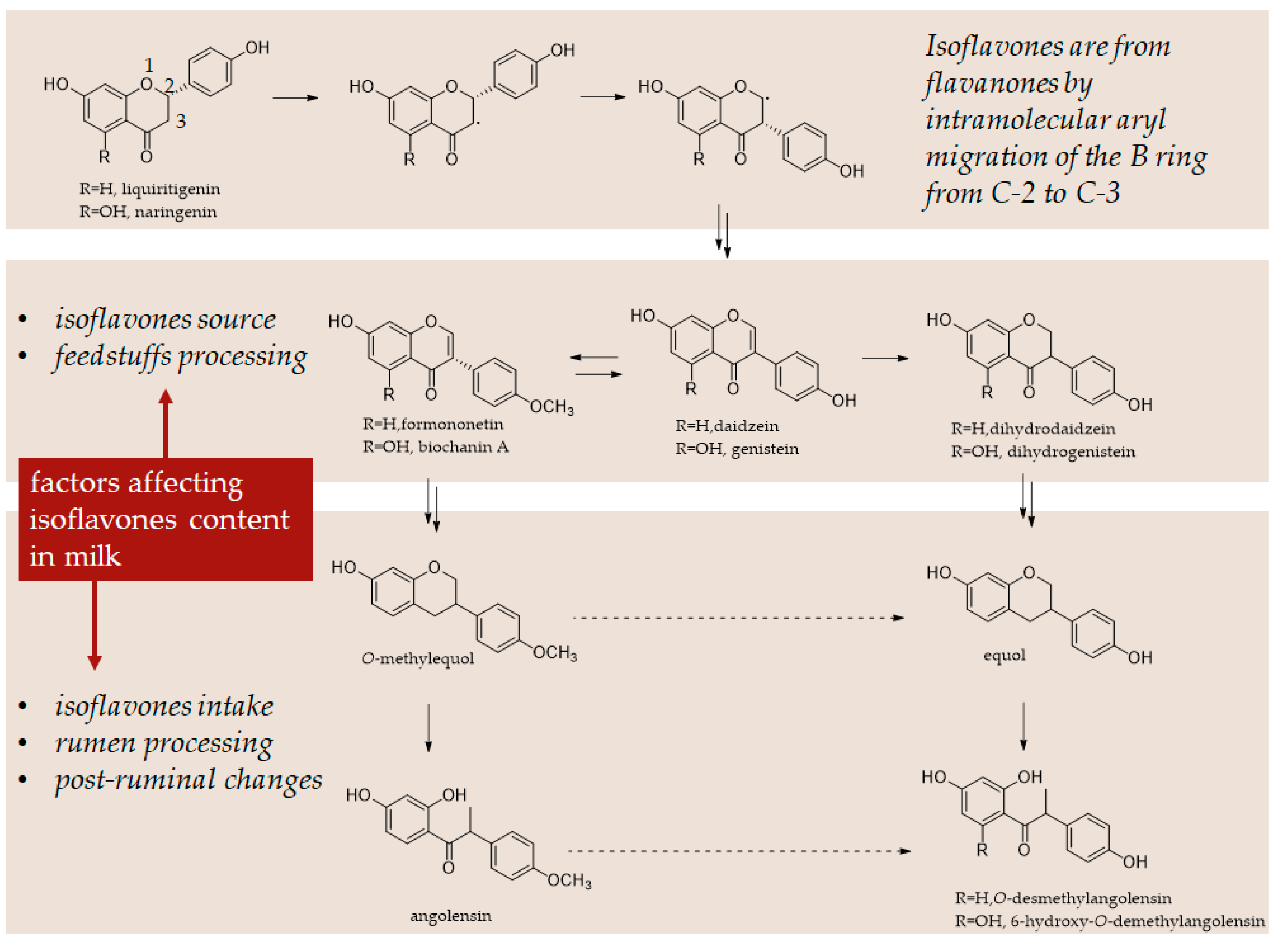
Milk Enriched in Polyphenols to Improve Human Health and Product Quality
6. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Castanon, J.I.R. History of the Use of Antibiotics as Growth Promoters in European Poultry Feeds. Poult. Sci. 2007, 86, 2466–2471. [Google Scholar] [CrossRef]
- Cheng, G.; Hao, H.; Xie, S.; Wang, X.; Dai, M.; Huang, L.; Yuan, Z. Antibiotic alternatives: The substitution of antibiotics in animal husbandry? Front. Microbiol. 2014, 5, 1–16. [Google Scholar] [CrossRef] [PubMed]
- Ballou, M.A.; Davis, E.M.; Kasl, B.A. Nutraceuticals an alternative strategy for the use of antimicrobials. Vet. Clin. N. Am. Food Anim. Pract. 2019, 35, 507–534. [Google Scholar] [CrossRef] [PubMed]
- Frankic, T.; Voljč, M.; Salobir, J.; Rezar, V. Use of herbs and spices and their extracts in animal nutrition. Acta Agric. Slov. 2009, 94, 95–102. [Google Scholar]
- Loy, D. By-product feed utilization by grazing cattle. Vet. Clin. North Am. Food Anim. Pract. 2007, 23, 41–52. [Google Scholar] [CrossRef]
- Halmemies-Beauchet-Filleau, A.; Rinne, M.; Lamminen, M.; Mapato, C.; Ampapon, T.; Wanapat, M.; Vanhatalo, A. Review: Alternative and novel feeds for ruminants: Nutritive value, product quality and environmental aspects. Animal 2018, 12, 295–309. [Google Scholar] [CrossRef]
- Correddu, F.; Lunesu, M.F.; Buffa, G.; Atzori, A.S.; Nudda, A.; Battacone, G.; Pulina, G. Can Agro-Industrial By-Products Rich in Polyphenols be Advantageously Used in the Feeding and Nutrition of Dairy Small Ruminants? Animals 2020, 10, 131. [Google Scholar] [CrossRef]
- Cencic, A.; Chingwaru, W. The Role of Functional Foods, Nutraceuticals, and Food Supplements in Intestinal Health. Nutrients 2010, 2, 611–625. [Google Scholar] [CrossRef]
- Cobellis, G.; Trabalza-Marinucci, M.; Yu, Z. Critical evaluation of essential oils as rumen modifiers in ruminant nutrition: A review. Sci. Total Environ. 2016, 545–546, 556–568. [Google Scholar] [CrossRef]
- Wang, J.-K.; Ye, J.-A.; Liu, J.-X. Effects of tea saponins on rumen microbiota, rumen fermentation, methane production and growth performance—A review. Trop. Anim. Health Prod. 2011, 44, 697–706. [Google Scholar] [CrossRef]
- Hartmann, T. From waste products to ecochemicals: Fifty years research of plant secondary metabolism. Phytochemistry 2007, 68, 2831–2846. [Google Scholar] [CrossRef] [PubMed]
- Dewick, P.M. Medicinal Natural Products: A Biosynthetic Approach, 3rd ed.; John Wiley & Sons: Hoboken, NJ, USA, 2009. [Google Scholar]
- Vasta, V.; Daghio, M.; Cappucci, A.; Buccioni, A.; Serra, A.; Viti, C.; Mele, M. Invited review: Plant polyphenols and rumen microbiota responsible for fatty acid biohydrogenation, fiber digestion, and methane emission: Experimental evidence and methodological approaches. J. Dairy Sci. 2019, 102, 3781–3804. [Google Scholar] [CrossRef]
- Jafari, S.; Ebrahimi, M.; Goh, Y.M.; Rajion, M.A.; Jahromi, M.F.; Al-Jumaili, W.S. Manipulation of rumen fermentation and methane gas production by plant secondary metabolites (saponin, tannin and essential oil) – A Review of Ten-Year Studie. Ann. Anim. Sci. 2019, 19, 3–29. [Google Scholar] [CrossRef]
- Khanbabaee, K.; Van Ree, T. Tannins: Classification and definition. Nat. Prod. Rep. 2001, 18, 641–649. [Google Scholar] [PubMed]
- Huang, Q.; Liu, X.; Zhao, G.; Hu, T.; Wang, Y. Potential and challenges of tannins as an alternative to in-feed antibiotics for farm animal production. Anim. Nutr. 2017, 4, 137–150. [Google Scholar] [CrossRef]
- Granato, D.; Santos, J.S.; Maciel, L.G.; Nunes, D.S. Chemical perspective and criticism on selected analytical methods used to estimate the total content of phenolic compounds in food matrices. Trends Analyt. Chem. 2016, 80, 266–279. [Google Scholar] [CrossRef]
- Folin, O.; Ciocalteu, V. On tyrosine and tryptophane determinations in proteins. J. Biol. Chem. 1927, 73, 627–650. [Google Scholar] [CrossRef]
- Singleton, V.L.; Rossi, J.A. Colorimetry of total phenolics with phosphomolybdic-phosphotungstic acid reagents. Am. J. Enol. Vitic. 1965, 16, 144–158. [Google Scholar]
- Munteanu, I.G.; Apetrei, C. Analytical Methods Used in Determining Antioxidant Activity: A Review. Int. J. Mol. Sci. 2021, 22, 3380. [Google Scholar] [CrossRef]
- Khoddami, A.; Wilkes, M.; Roberts, T. Techniques for Analysis of Plant Phenolic Compounds. Molecules 2013, 18, 2328–2375. [Google Scholar] [CrossRef]
- Pękal, A.; Pyrzynska, K. Evaluation of aluminium complexation reaction for flavonoid content assay. Food Anal. Methods 2014, 7, 1776–1782. [Google Scholar] [CrossRef]
- Herderich, M.J.; Smith, P.A. Analysis of grape and wine tannins: Methods, applications and challenges. Aust. J. Grape Wine Res. 2005, 11, 205–214. [Google Scholar] [CrossRef]
- Price, M.L.; Van Scoyoc, S.; Butler, L.G. A critical evaluation of the vanillin reaction as an assay for tannin in sorghum grain. J. Agric. Food Chem. 1978, 26, 1214–1218. [Google Scholar] [CrossRef]
- Hartzfeld, P.W.; Forkner, R.; Hunter, M.D.; Hagerman, A.E. Determination of hydrolysable tannins (gallotannins and ellagitannins) after reaction with potassium iodate. J. Agric. Food Chem. 2002, 50, 1785–1790. [Google Scholar] [CrossRef] [PubMed]
- Salminen, J.-P.; Karonen, M. Chemical ecology of tannins and other phenolics: We need a change in approach. Func. Ecol. 2011, 25, 325–338. [Google Scholar] [CrossRef]
- Costa, D.C.; Costa, H.S.; Albuquerque, T.G.; Ramos, F.; Castilho, M.C.; Sanches-Silva, A. Advances in phenolic compounds analysis of aromatic plants and their potential applications. Trends Food Sci. Technol. 2015, 45, 336–354. [Google Scholar] [CrossRef]
- Chiriac, E.R.; Chi¸tescu, C.L.; Gean ˘a, E.-I.; Gird, C.E.; Socoteanu, R.P.; Boscencu, R. Advanced Analytical Approaches for the Analysis of Polyphenols in Plants Matrices—A Review. Separations 2021, 8, 65. [Google Scholar] [CrossRef]
- Luis Aleixandre-Tudo, J.; du Toit, W. The Role of UV-Visible spectroscopy for phenolic compounds quantification in winemaking. In Frontiers and New Trends in the Science of Fermented Food and Beverages, 1st ed.; Rosa Lidia Solís-Oviedo; IntechOpen: London, UK, 2019. [Google Scholar]
- Lee, S.J.; Kim, H.S.; Eom, J.S.; Choi, Y.Y.; Jo, S.; Chu, G.M.; Lee, Y.; Seo, J.; Kim, K.H.; Lee, S.S. Effects of Olive (Olea europaea L.) leaves with antioxidant and antimicrobial activities on in Vitro ruminal fermentation and methane emission. Animals 2021, 11, 2008–2022. [Google Scholar]
- Pellikaan, W.F.; Stringano, E.; Leenaars, J.; Bongers, D.J.G.M.; van Laar-van Schuppen, S.; Plant, J.; Mueller-Harvey, I. Evaluating effects of tannins on extent and rate of in vitro gas and CH4 production using an automated pressure evaluation system (APES). Anim. Feed Sci. Technol. 2011, 166–167, 377–390. [Google Scholar] [CrossRef]
- Puchalska, J.; Szumacher-Strabel, M.; Patra, A.K.; Ślusarczyk, S.; Gao, M.; Petrič, D.; Nabzdyk, M.; Cieślak, A. The effect of different concentrations of total polyphenols from paulownia hybrid leaves on ruminal fermentation, methane production and microorganisms. Animals 2021, 11, 2843. [Google Scholar] [CrossRef]
- Formato, M.; Piccolella, S.; Zidorn, C.; Vastolo, A.; Calabrò, S.; Cutrignelli, M.I.; Pacifico, S. UHPLC-ESI-QqTOF Analysis and In vitro rumen fermentation for exploiting Fagus sylvatica leaf in ruminant diet. Molecules 2022, 27, 2217. [Google Scholar] [CrossRef] [PubMed]
- Saleem, F.; Bouatra, S.; Guo, A.C.; Psychogios, N.; Mandal, R.; Dunn, S.M.; Ametaj, N.B.; Wishart, D.S. The bovine ruminal fluid metabolome. Metabolomics 2013, 9, 360–378. [Google Scholar] [CrossRef]
- De Almeida, R.T.R.; Do Prado, R.M.; Porto, C.; Dos Santos, G.T.; Huws, S.A.; Pilau, E.J. Exploring the rumen fluid metabolome using liquid chromatography-high-resolution mass spectrometry and Molecular Networking. Sci. Rep. 2018, 8, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Foroutan, A.; Fitzsimmons, C.; Mandal, R.; Piri-Moghadam, H.; Zheng, J.; Guo, A.; Li, C.; Guan, L.L.; Wishart, D.S. The Bovine Metabolome. Metabolites 2020, 10, 233. [Google Scholar] [CrossRef]
- McDonald, P.; Greenhalgh, J.F.D.; Morgan, C.A. Digestion. In Animal Nutrion; Pearson Education Limited: London, UK, 2011; pp. 156–191. [Google Scholar]
- Vastolo, A.; Calabrò, S.; Pacifico, S.; Koura, B.I.; Cutrignelli, M.I. Chemical and nutritional characteristics of Cannabis sativa L. co-products. J. Anim. Physiol. Anim. Nutr. 2021, 105, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Vastolo, A.; Matera, R.; Serrapica, F.; Cutrignelli, M.I.; Neglia, G.; Kiatti, D.D.; Calabrò, S. Improvement of Rumen Fermentation Efficiency Using Different Energy Sources: In Vitro Comparison between Buffalo and Cow. Fermentation 2022, 8, 351. [Google Scholar] [CrossRef]
- Plaizier, J.C.; Khafipour, E.; Li, S.; Gozho, G.N.; Krause, D.O. Subacute ruminal acidosis (SARA), endotoxins and health consequences. Anim. Feed. Sci. Tech. 2012, 172, 9–21. [Google Scholar] [CrossRef]
- Moss, A.R.; Jouany, J.P.; Newbold, J. Methane production by ruminants: Its contribution to global warming. Ann. Zootech. 2000, 49, 231–253. [Google Scholar] [CrossRef]
- Firkins, J.L.; Yu, Z.; Morrison, M. Ruminal Nitrogen Metabolism: Perspectives for integration of microbiology and nutrition for dairy. J. Dairy Sci. 2007, 90, E1–E16. [Google Scholar] [CrossRef]
- Cholewińska, P.; Czyż, K.; Nowakowski, P.; Wyrostek, A. The microbiome of the digestive system of ruminants – a review. Anim. Health Res. Rev. 2020, 21, 3–14. [Google Scholar] [CrossRef]
- Matthews, C.; Crispie, F.; Lewis, E.; Reid, M.; O’Toole, P.W.; Cotter, P.D. The rumen microbiome: A crucial consideration when optimising milk and meat production and nitrogen utilisation efficiency. Gut Microbes 2018, 10, 115–132. [Google Scholar] [CrossRef] [PubMed]
- Ma, T.; Wu, W.; Tu, Y.; Zhang, N.; Diao, Q. Resveratrol affects in vitro rumen fermentation, methane production and prokaryotic community composition in a time- and diet-specific manner. Microb Biotechnol. 2020, 13, 1118–1131. [Google Scholar] [CrossRef]
- Khafipour, E.; Li, S.; Tun, H.M.; Derakhshani, H.; Moossavi, S.; Plaizier, J. Effects of grain feeding on microbiota in the digestive tract of cattle. Anim. Front. 2016, 6, 13–19. [Google Scholar] [CrossRef]
- Koike, S.; Kobayashi, Y. Fibrolytic Rumen Bacteria: Their Ecology and Functions. Anim. Biosci. 2009, 22, 131–138. [Google Scholar] [CrossRef]
- Duskova, D.; Marounek, M. Fermentation of pectin and glucose, and activity of pectin-degrading enzymes in the rumen bacterium Lachnospira multiparus. Lett. Appl. Microbiol. 2001, 33, 159–163. [Google Scholar] [CrossRef] [PubMed]
- Oren, A. The Family Methanobacteriaceae. In The Prokaryotes, 1st ed.; Springer: Berlin/Heidelberg, Germany, 2014; pp. 165–193. [Google Scholar]
- Patra, A.; Park, T.; Kim, M.; Yu, Z. Rumen methanogens and mitigation of methane emission by anti-methanogenic compounds and substances. J. Anim. Sci. Biotech. 2017, 8, 1–18. [Google Scholar] [CrossRef]
- Knapp, J.R.; Laur, G.L.; Vadas, P.A.; Weiss, W.P.; Tricarico, J.M. Invited review: Enteric methane in dairy cattle production: Quantifying the opportunities and impact of reducing emissions. Int. J. Dairy Sci. 2014, 97, 3231–3261. [Google Scholar] [CrossRef]
- Gruby, D.; Delafond, H.M.O. Recherches sur des animalcules se développant en grand nombre dans l’estomac et dans les intestins, pendant la digestion des animaux herbivores et carnivores. Compt. Rend. Acad. Sci. 1843, 17, 1304–1308. [Google Scholar]
- Hobson, P.N.; Stewart, C.S. The Rumen Microbial Ecosystem, 2nd ed.; Springer: Dordrecht, The Netherlands, 1997. [Google Scholar]
- Agarwal, N.; Kamra, D.N.; Chaudhary, L.C. Rumen microbial ecosystem of domesticated ruminants. In Rumen Microbiology: From Evolution to Revolution; Springer: New Delhi, India, 2015; pp. 17–30. [Google Scholar]
- Jafari, S.; Meng, G.Y.; Rajion, M.A.; Jahromi, M.F.; Ebrahimi, M. Manipulation of rumen microbial fermentation by polyphenol rich solvent fractions from papaya leaf to reduce green-house gas methane and biohydrogenation of c18 PUFA. J. Agric. Food Chem. 2016, 64, 4522–4530. [Google Scholar] [CrossRef]
- Buccioni, A.; Minieri, S.; Rapaccini, S.; Antongiovanni, M.; Mele, M. Effect of chestnut and quebracho tannins on fatty acid profile in rumen liquid- and solid-associated bacteria: An in vitro study. Animal 2011, 5, 1521–1530. [Google Scholar] [CrossRef]
- Getabalew, M.; Alemneh, T.; Akeberegn, D. Methane production in ruminant animals: Implication for their impact on climate change. Concepts Dairy Vet. Sci. 2019, 2, 204–211. [Google Scholar]
- McAllister, T.A.; Cheng, K.-J.; Okine, E.K.; Mathison, G.W. Dietary, environmental and microbiological aspects of methane production in ruminants. Can. J. Anim. Sci. 1996, 76, 231–243. [Google Scholar] [CrossRef]
- Finlay, B.J.; Esteban, G.; Clarke, K.J.; Williams, A.G.; Embley, T.M.; Hirt, R.P. Some rumen ciliates have endosymbiotic methanogens. FEMS Microbiol. Lett. 1994, 117, 157–161. [Google Scholar] [CrossRef] [PubMed]
- Morgavi, D.P.; Martin, C.; Jouany, J.-P.; Ranilla, M.J. Rumen protozoa and methanogenesis: Not a simple cause–effect relationship. Br. J. Nutr. 2011, 107, 388–397. [Google Scholar] [CrossRef]
- Cieslak, A.; Szumacher-Strabel, M.; Stochmal, A.; Oleszek, W. Plant components with specific activities against rumen methanogens. Animal 2013, 7, 253–265. [Google Scholar] [CrossRef]
- Chen, J.; Harstad, O.M.; McAllister, T.; Dörsch, P.; Holo, H. Propionic acid bacteria enhance ruminal feed degradation and reduce methane production in vitro. Acta Agric. Scand.-A Anim. Sci. 2020, 69, 1–7. [Google Scholar] [CrossRef]
- Xie, Y.; Yang, W.; Tang, F.; Chen, X.; Ren, L. Antibacterial activities of flavonoids: Structure-activity relationship and mechanism. Curr. Med. Chem. 2014, 22, 132–149. [Google Scholar] [CrossRef]
- Wang, D.; Huang, J.; Zhang, Z.; Tian, X.; Huang, H.; Yu, Y.; Zhang, G.; Ding, J.; Huang, R. Influences of Portulaca oleracea extracts on in vitro methane emissions and rumen fermentation of forage. J. Food Agric. Environ. 2013, 11, 483–488. [Google Scholar]
- Vera, N.; Gutierrez, C.; Williams, P.; Fuentealba, C.; Allende, R.; Avila-Stagno, J. Low concentrations of a polyphenolic extract from pine bark in high–concentrate diets decrease in vitro rumen ammonia nitrogen but not methane production. Res. J. Appl. Sci. 2021, 49, 413–422. [Google Scholar] [CrossRef]
- Sommai, S.; Cherdthong, A.; Suntara, C.; So, S.; Wanapat, M.; Polyorach, S. In vitro Fermentation Characteristics and Methane Mitigation Responded to Flavonoid Extract Levels from Alternanthera sissoo and Dietary Ratios. Fermentation 2021, 7, 109. [Google Scholar] [CrossRef]
- Oskoueian, E.; Abdullah, N.; Oskoueian, A. Effects of flavonoids on rumen fermentation activity, methane production, and microbial population. BioMed Res. Int. 2013, 2013, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Becker, P.M.; van Wikselaar, P.G.; Franssen, M.C.R.; de Vos, R.C.H.; Hall, R.D.; Beekwilder, J. Evidence for a hydrogen-sink mechanism of (+) catechin-mediated emission reduction of the ruminant greenhouse gas methane. Metabolomics 2014, 10, 179–189. [Google Scholar] [CrossRef]
- Stoldt, A.K.; Derno, M.; Das, G.; Weitzel, J.M.; Wolffram, S.; Metges, C.C. Effects of rutin and buckwheat seeds on energy metabolism and methane production in dairy cows. Int. J. Dairy Sci. 2016, 99, 2161–2168. [Google Scholar] [CrossRef]
- Berger, L.M.; Blank, R.; Zorn, F.; Wein, S.; Metges, C.C.; Wolffram, S. Ruminal degradation of quercetin and its influence on fermentation in ruminants. Int. J. Dairy Sci. 2015, 98, 5688–5698. [Google Scholar] [CrossRef]
- Cui, K.; Guo, X.D.; Tu, Y.; Zhang, N.F.; Ma, T.; Diao, Q.Y. Effect of dietary supplementation of rutin on lactation performance, ruminal fermentation and metabolism in dairy cows. J. Anim. Physiol. Anim. 2015, 99, 1065–1073. [Google Scholar] [CrossRef] [PubMed]
- Sinz, S.; Carmen, K.; Annette, L.; Ueli, B.; Svenja, M.; Carla, R.S.; Michael, K. In vitro bioactivity of various pure flavonoids in ruminal fermentation, with special reference to methane formation. Czech J. Anim. Sci. 2018, 63, 293–304. [Google Scholar]
- Seradj, A.R.; Abecia, L.; Crespo, J.; Villalba, D.; Fondevila, M.; Balcells, J. The effect of Bioflavex® and its pure flavonoid components on in vitro fermentation parameters and methane production in rumen fluid from steers given high concentrate diets. Anim. Feed Sci. Technol. 2014, 197, 85–91. [Google Scholar] [CrossRef]
- Kim, E.T.; Guan, L.L.; Lee, S.J.; Lee, S.M.; Lee, S.S.; Lee, I.D.; Lee, S.S. Effects of Flavonoid-rich Plant Extracts on In vitro Ruminal Methanogenesis, Microbial Populations and Fermentation Characteristics. Asian Australas J. Anim. Sci. 2015, 28, 530–537. [Google Scholar] [CrossRef]
- Boussaada, R.A.; Calabro, S.; Grazioli, R.; Ferrara, M.G.; Musco, N.; Cutrignelli, M.I.; Tidjane, M. Comparison of the Effect of Olives Leaves Extracts (Olea europaea) on in vitro Methane Production, Fermentation Efficiency and Protozoa Activity. Glob. Vet. 2017, 18, 445–453. [Google Scholar]
- de Paula, E.M.; Samensari, R.B.; Machado, E.; Pereira, L.M.; Maia, F.J.; Yoshimura, E.H.; Zeoula, L.M. Effects of phenolic compounds on ruminal protozoa population, ruminal fermentation, and digestion in water buffaloes. Livest. Sci. 2016, 185, 136–141. [Google Scholar] [CrossRef]
- Kong, L.; Yang, C.; Dong, L.; Diao, Q.; Si, B.; Ma, J.; Tu, Y. Rumen Fermentation Characteristics in Pre- and Post-Weaning Calves upon Feeding with Mulberry Leaf Flavonoids and Candida tropicalis Individually or in Combination as a Supplement. Animals 2019, 9, 990. [Google Scholar] [CrossRef] [PubMed]
- Bhatta, R.; Baruah, L.; Saravanan, M.; Suresh, K.P.; Sampath, K.T. Effect of medicinal and aromatic plants on rumen fermentation, protozoa population and methanogenesis in vitro. J. Anim. Physiol. Anim. 2012, 97, 446–456. [Google Scholar] [CrossRef] [PubMed]
- Bhatta, R.; Saravanan, M.; Baruah, L.; Prasad, C.S. Effects of graded levels of tannin-containing tropical tree leaves on in vitro rumen fermentation, total protozoa and methane production. J. App. Microbiol. 2015, 118, 557–564. [Google Scholar] [CrossRef]
- Jayanegara, A.; Goel, G.; Makkar, H.P.S.; Becker, K. Divergence between purified hydrolysable and condensed tannin effects on methane emission, rumen fermentation and microbial population in vitro. Anim. Feed Sci. Technol. 2015, 209, 60–68. [Google Scholar] [CrossRef]
- Dschaak, C.M.; Williams, C.M.; Holt, M.S.; Eun, J.-S.; Young, A.J.; Min, B.R. Effects of supplementing condensed tannin extract on intake, digestion, ruminal fermentation, and milk production of lactating dairy cows. J. Dairy Sci. 2011, 94, 2508–2519. [Google Scholar] [CrossRef]
- Wang, Y.; Yu, S.; Li, Y.; Zhang, S.; Qi, X.; Guo, K.; Guo, Y.; Fortina, R. Pilot Study of the Effects of Polyphenols from Chestnut Involucre on Methane Production, Volatile Fatty Acids, and Ammonia Concentration during in vitro Rumen Fermentation. Animals 2021, 11, 108. [Google Scholar] [CrossRef]
- Olennikov, D.; Kashchenko, N.; Chirikova, N. In Vitro Bioaccessibility, Human Gut Microbiota Metabolites and Hepatoprotective Potential of Chebulic Ellagitannins: A Case of Padma Hepaten® Formulation. Nutrients 2015, 7, 8456–8477. [Google Scholar] [CrossRef]
- Thanh, L.P.; Kha, P.T.T.; Loor, J.J.; Hang, T.T.T. Grape seed tannin extract and polyunsaturated fatty acids affect in vitro ruminal fermentation and methane production. J. Anim. Sci. 2022, 100, 1–38. [Google Scholar] [CrossRef]
- Moate, P.J.; Williams, S.R.O.; Torok, V.A.; Hannah, M.C.; Ribaux, B.E.; Tavendale, M.H.; Eckard, R.J.; Jacobs, J.L.; Auldist, M.J.; Wales, W.J. Grape marc reduces methane emissions when fed to dairy cows. J. Dairy S. 2014, 97, 5073–5087. [Google Scholar] [CrossRef]
- Guglielmelli, A.; Calabro`, S.; Primi, R.; Carone, F.; Cutrignelli, M.I.; Tudisco, R.; Piccolo, G.; Ronch, B.; Danieli, P.P. In vitro fermentation patterns and methane production of sainfoin (Onobrychis viciifolia Scop.) hay with different condensed tannin contents. Grass and Forage Sci. 2011, 66, 488–500. [Google Scholar] [CrossRef]
- Uushona, T.; Chikwanha, O.C.; Tayengwa, T.; Katiyatiya, C.L.F.; Strydom, P.E.; Mapiye, C. Nutraceutical and preservative potential of Acacia mearnsii and Acacia dealbata leaves for ruminant production and product quality enhancement. J. Agric. Sci. 2021, 159, 743–756. [Google Scholar] [CrossRef]
- Denninger, T.M.; Schwarm, A.; Birkinshaw, A.; Terranova, M.; Dohme-Meier, F.; Münger, A.; Eggerschwiler, L.; Bapst, B.; Wegmann, S.; Clauss, M.; et al. Immediate effect of Acacia mearnsii tannins on methane emissions and milk fatty acid profiles of dairy cows. Anim. Feed Sci. Technol. 2020, 261, 1–13. [Google Scholar] [CrossRef]
- Avila, A.S.; Zambom, M.A.; Faccenda, A.; Fischer, M.L.; Anschau, F.A.; Venturini, T.; Faciola, A.P. Effects of Black Wattle (Acacia mearnsii) condensed tannins on intake, protozoa population, ruminal fermentation, and nutrient digestibility in jersey steers. Animals 2020, 10, 1011. [Google Scholar] [CrossRef]
- Junior, F.P.; Cassiano, E.C.O.; Martins, M.F.; Romero, L.A.; Zapata, D.C.V.; Pinedo, L.A.; Marino, C.T.; Rodrigues, P.H.M. Effect of tannins-rich extract from Acacia mearnsii or monensin as feed additives on ruminal fermentation efficiency in cattle. Livest. Sci. 2017, 203, 21–29. [Google Scholar] [CrossRef]
- Junior, F.P.; Zapata Vásquez, D.C.; Gardinal, R.; Meyer, P.M.; Alexandre, B.; Friguetto, R.T.S.; de Abreu Demarchi, J.J.A.; Rodrigues, P.H.M. Short-term use of monensin and tannins as feed additives on digestibility and methanogenesis in cattle. R. Bras. Zootec. 2019, 49, 1–9. [Google Scholar]
- Singh, S.; Hundal, J.S.; Patra, A.K.; Sethi, R.S.; Sharma, A. A composite polyphenol-rich extract improved growth performance, ruminal fermentation and immunity, while decreasing methanogenesis and excretion of nitrogen and phosphorus in growing buffaloes. Environ. Sci. Pollut. Res. Int. 2021, 29, 24757–24773. [Google Scholar] [CrossRef] [PubMed]
- Cieslak, A.; Zmora, P.; Pers-Kamczyc, E.; Szumacher-Strabel, M. Effects of tannins source (Vaccinium vitis idaea L.) on rumen microbial fermentation in vivo. Anim Feed Sci. Technol. 2012, 176, 102–106. [Google Scholar] [CrossRef]
- Cieslak, A.; Zmora, P.; Pers-Kamczyc, E.; Stochmal, A.; Sadowinska, A.; Salem, A.Z.; Kowalczyk, D.; Zbonik, P.; Szumacher-Strabel, M. Effects of two sources of tannins (Quercus L. and Vaccinium vitis idaea L.) on rumen microbial fermentation: An in vitro study. Ital. J. Animal Sci. 2014, 13, 290–294. [Google Scholar] [CrossRef]
- Cieslak, A.; Zmora, P.; Matkowski, A.; Nawrot-Hadzik, I.; Pers- Kamczyc, E.; El-Sherbiny, M.; Bryszak, M.; Szumacher- Strabel, M. Tannins from Sanguisorba officinalis affect in vitro rumen methane production and fermentation. J. Anim. Plant Sci. 2016, 26, 54–62. [Google Scholar]
- Choi, Y.; Lee, S.J.; Kim, H.S.; Eom, J.S.; Jo, S.U.; Guan, L.L.; Seo, J.; Kim, H.; Lee, S.S.; Lee, S.S. Effects of seaweed extracts on in vitro rumen fermentation characteristics, methane production, and microbial abundance. Sci. Rep. 2021, 11, 24092–24104. [Google Scholar] [CrossRef]
- Kim, E.J.; Huws, S.A.; Lee, M.R.F.; Scollan, N.D. Dietary Transformation of Lipid in the Rumen Microbial Ecosystem. Asian-australas. J. Anim. Sci. 2009, 22, 1341–1350. [Google Scholar] [CrossRef]
- Lourenço, M.; Ramos-Morales, E.; Wallace, R.J. The role of microbes in rumen lipolysis and biohydrogenation and their manipulation. Animal 2010, 4, 1008–1023. [Google Scholar] [CrossRef] [PubMed]
- Dewanckele, L.; Toral, P.G.; Vlaeminck, B.; Fievez, V. Invited review: Role of rumen biohydrogenation intermediates and rumen microbes in diet-induced milk fat depression: An update. J. Dairy Sci. 2020, 103, 7655–7681. [Google Scholar] [CrossRef] [PubMed]
- Or-Rashid, M.M.; AlZahal, O.; McBride, B.W. Studies on the production of conjugated linoleic acid from linoleic and vaccenic acids by mixed rumen protozoa. Appl. Microbiol. Biotechnol. 2008, 81, 533–541. [Google Scholar] [CrossRef] [PubMed]
- Nam, I.S.; Garnsworthy, P.C. Biohydrogenation of linoleic acid by rumen fungi compared with rumen bacteria. J.Appl.Microbiol. 2007, 103, 551–556. [Google Scholar] [CrossRef] [PubMed]
- Chilliard, Y.; Glasser, F.; Ferlay, A.; Bernard, L.; Rouel, J.; Doreau, M. Diet, rumen biohydrogenation and nutritional quality of cow and goat milk fat. Eur. J. Lipid Sci. Tech. 2007, 109, 828–855. [Google Scholar] [CrossRef]
- Jayanegara, A.; Kreuzer, M.; Wina, E.; Leiber, F. Significance of phenolic compounds in tropical forages for the ruminal bypass of polyunsaturated fatty acids and the appearance of biohydrogenation intermediates as examined in vitro. Anim. Prod. Sci. 2011, 51, 1127–1136. [Google Scholar] [CrossRef]
- Khiaosa-Ard, R.; Bryner, S.F.; Scheeder, M.R.L.; Wettstein, H.R.; Leiber, F.; Kreuzer, M.; Soliva, C.R. Evidence for the inhibition of the terminal step of ruminal α-linolenic acid biohydrogenation by condensed tannins. J. Dairy Sci. 2009, 92, 177–188. [Google Scholar] [CrossRef]
- Jayanegara, A.; Kreuzer, M.; Leiber, F. Ruminal disappearance of polyunsaturated fatty acids and appearance of biohydrogenation products when incubating linseed oil with alpine forage plant species in vitro. Livest. Sci. 2012, 147, 104–112. [Google Scholar] [CrossRef]
- Lee, M.R.; Winters, A.L.; Scollan, N.D.; Dewhurst, R.J.; Theodorou, M.K.; Minchin, F.R. Plant-mediated lipolysis and proteolysis in red clover with different polyphenol oxidase activities. J. Sci. Food Agric. 2004, 84, 1639–1645. [Google Scholar] [CrossRef]
- Cabiddu, A.; Salis, L.; Tweed, J.K.; Molle, G.; Decandia, M.; Lee, M.R. The influence of plant polyphenols on lipolysis and biohydrogenation in dried forages at different phenological stages: In vitro study. J. Sci. Food Agric. 2010, 90, 829–835. [Google Scholar] [CrossRef] [PubMed]
- Halmemies-Beauchet-Filleau, A.; Vanhatalo, A.; Toivonen, V.; Heikkilä, T.; Lee, M.R.F.; Shingfield, K.J. Effect of replacing grass silage with red clover silage on ruminal lipid metabolism in lactating cows fed diets containing a 60:40 forage-to-concentrate ratio. J. Dairy Sci. 2013, 96, 5882–5900. [Google Scholar] [CrossRef]
- Vasta, V.; Makkar, H.P.S.; Mele, M.; Priolo, A. Ruminal biohydrogenation as affected by tannins in vitro. Br. J. Nutr. 2008, 102, 82–92. [Google Scholar] [CrossRef] [PubMed]
- Ishlak, A.; Günal, M.; AbuGhazaleh, A.A. The effects of cinnamaldehyde, monensin and quebracho condensed tannin on rumen fermentation, biohydrogenation and bacteria in continuous culture system. Anim. Feed Sci. Technol. 2015, 207, 31–40. [Google Scholar] [CrossRef]
- Kronberg, S.L.; Scholljegerdes, E.J.; Barceló-Coblijn, G.; Murphy, E.J. Flaxseed Treatments to Reduce Biohydrogenation of α-Linolenic Acid by Rumen Microbes in Cattle. Lipids 2007, 42, 1105–1111. [Google Scholar] [CrossRef] [PubMed]
- Szczechowiak, J.; Szumacher-Strabel, M.; El-Sherbiny, M.; Pers-Kamczyc, E.; Pawlak, P.; Cieslak, A. Rumen fermentation, methane concentration and fatty acid proportion in the rumen and milk of dairy cows fed condensed tannin and/or fish-soybean oils blend. Anim. Feed Sci. Technol. 2016, 216, 93–107. [Google Scholar] [CrossRef]
- Sato, Y.; Nakanishi, T.; Wang, L.; Oishi, K.; Hirooka, H.; Kumagai, H. In vitro and in vivo evaluations of wine lees as feeds for ruminants: Effects on ruminal fermentation characteristics, nutrient digestibility, blood metabolites and antioxidant status. Livest. Sci. 2020, 241, 104217. [Google Scholar] [CrossRef]
- Lourenço, M.; Cardozo, P.W.; Calsamiglia, S.; Fievez, V. Effects of saponins, quercetin, eugenol, and cinnamaldehyde on fatty acid biohydrogenation of forage polyunsaturated fatty acids in dual-flow continuous culture fermenters. J. Anim. Sci. 2008, 86, 3045–3053. [Google Scholar] [CrossRef]
- Keshri, A.; Bashir, Z.; Kumari, V.; Prasad, K.; Joysowal, M.; Singh, M.; Singh, D.; Tarun, A.; Shukla, S. Role of micronutrients during peri-parturient period of dairy animals—A review. Biol. Rhythm Res. 2019, 52, 1–13. [Google Scholar] [CrossRef]
- Xiao, J.; Khan, M.Z.; Ma, Y.; Alugongo, G.M.; Ma, J.; Chen, T.; Khan, A.; Cao, Z. The Antioxidant Properties of Selenium and Vitamin E; Their Role in Periparturient Dairy Cattle Health Regulation. Antioxidants 2021, 10, 1555. [Google Scholar] [CrossRef]
- Bradford, B.J.; Yuan, K.; Farney, J.K.; Mamedova, L.K.; Carpenter, A.J. Invited review: Inflammation during the transition to lactation: New adventures with an old flame. Int. J. Dairy Sci. 2015, 98, 6631–6650. [Google Scholar] [CrossRef] [PubMed]
- Abuelo, A.; Hernández, J.; Benedito, J.L.; Castillo, C. Redox biology in transition periods of dairy cattle: Role in the health of periparturient and neonatal animals. Antioxidants 2019, 8, 20. [Google Scholar] [CrossRef] [PubMed]
- Winkler, A.; Gessner, D.K.; Koch, C.; Romberg, F.J.; Dusel, G.; Herzog, E.; Most, E.; Eder, K. Effects of a plant product consisting of green tea and curcuma extract on milk production and the expression of hepatic genes involved in endoplasmic stress response and inflammation in dairy cows. Arch.Anim. Nutr. 2015, 69, 425–441. [Google Scholar] [CrossRef]
- Olagaray, K.E.; Brouk, M.J.; Mamedova, L.K.; Sivinski, S.E.; Liu, H.; Robert, F.; Dupuis, E.; Zachut, M.; Bradford, B.J.; Loor, J.J. Dietary supplementation of Scutellaria baicalensis extract during early lactation decreases milk somatic cells and increases whole lactation milk yield in dairy cattle. PLoS ONE 2019, 14, 1–23. [Google Scholar] [CrossRef]
- Hao, Y.; Huang, S.; Si, J.; Zhang, J.; Gaowa, N.; Sun, X.; Lv, J.; Liu, G.; He, Y.; Wang, W.; et al. Effects of Paper Mulberry Silage on the Milk Production, Apparent Digestibility, Antioxidant Capacity, and Fecal Bacteria Composition in Holstein Dairy Cows. Animals 2020, 10, 1152. [Google Scholar] [CrossRef] [PubMed]
- Costa, A.; Neglia, G.; Campanile, G.; De Marchi, M. Milk somatic cell count and its relationship with milk yield and quality traits in Italian water buffaloes. J. Diary Sci. 2020, 103, 5485–5494. [Google Scholar] [CrossRef]
- Bonanno, A.; Di Grigoli, A.; Todaro, M.; Alabiso, M.; Vitale, F.; Di Trana, A.; Giorgio, D.; Settanni, L.; Gaglio, R.; Laddomada, B.; et al. Improvement of Oxidative Status, Milk and Cheese Production, and Food Sustainability Indexes by Addition of Durum Wheat Bran to Dairy Cows’ Diet. Animals 2019, 9, 698. [Google Scholar] [CrossRef]
- Chedea, V.S.; Pelmus, R.S.; Lazar, C.; Pistol, G.C.; Calin, L.G.; Toma, S.M.; Dragomir, C.; Taranu, I. Effects of a diet containing dried grape pomace on blood metabolites and milk composition of dairy cows. J. Sci. Food Agric. 2017, 97, 2516–2523. [Google Scholar] [CrossRef] [PubMed]
- Ianni, A.; Di Maio, G.; Pittia, P.; Grotta, L.; Perpetuini, G.; Tofalo, R.; Cichelli, A.; Martino, G. Chemical-nutritional quality and oxidative stability of milk and dairy products obtained from Friesian cows fed with a dietary supplementation of dried grape pomace. J. Sci. Food Agric. 2019, 99, 3635–3643. [Google Scholar] [CrossRef]
- Lee, S.H.; Humphries, D.J.; Cockman, D.A.; Givens, D.I.; Spencer, J.P. Accumulation of Citrus Flavanones in Bovine Milk Following Citrus Pulp Incorporation into the Diet of Dairy Cows. EC Nutr. 2017, 7, 143–154. [Google Scholar]
- Ma, Y.; Feng, Y.; Song, L.; Li, M.; Dai, H.; Bao, H.; Zhang, G.; Zhao, L.; Zhang, C.; Yi, J.; et al. Green tea polyphenols supplementation alters immunometabolism and oxidative stress in dairy cows with hyperketonemia. Animal Nutr. 2021, 7, 206–215. [Google Scholar] [CrossRef] [PubMed]
- Pauletto, M.; Elgendy, R.; Ianni, A.; Marone, E.; Giantin, M.; Grotta, L.; Ramazzotti, S.; Bennato, F.; Dacasto, M.; Martino, G. Nutrigenomic Effects of Long-Term Grape Pomace Supplementation in Dairy Cows. Animals 2020, 10, 714. [Google Scholar] [CrossRef] [PubMed]
- Cohen-Zinder, M.; Weinberg, Z.; Leibovich, H.; Chen, Y.; Rosen, M.; Sagi, G.; Orlov, A.; Agmon, R.; Yishay, M.; Miron, J.; et al. Ensiled Moringa Oleifera: An Antioxidant-Rich Feed That Improves Dairy Cattle Performance. J. Agric. Sci. 2017, 155, 1174–1186. [Google Scholar] [CrossRef]
- Gunun, P.; Gunun, N.; Khejornsart, P.; Ouppamong, T.; Cherdthong, A.; Wanapat, M.; Sirilaophaisan, S.U.; Yuangklang, C.; Polyorach, S.; Kenchaiwong, W.; et al. Effects of Antidesma thwaitesianum Muell. Arg. pomace as a source of plant secondary compounds on digestibility, rumen environment, hematology, and milk production in dairy cows. J. Anim. Sci. 2018, 90, 372–381. [Google Scholar] [CrossRef]
- Grazziotin, R.C.B.; Halfen, J.; Rosa, F.; Schmitt, E.; Anderson, J.L.; Ballard, V.; Osorio, J.S. Altered rumen fermentation patterns in lactating dairy cows supplemented with phytochemicals improve milk production and efficiency. J. Dairy Sci. 2020, 103, 301–312. [Google Scholar] [CrossRef] [PubMed]
- Davidović, V.; Jovetić, B.; Joksimović-Todorović, M.; Stojanović, B.; Lazarević, M.; Perisić, P.; Radivojević, M.; Maletić, M.; Miletić, A. The effect of tannin supplementation of mid-lactation dairy cows diets on metabolic profile parameters and production characteristics. Slov. Vet. Res. 2019, 56, 143–151. [Google Scholar] [CrossRef]
- Estrada-Flores, J.G.; Pedraza-Beltrán, P.E.; Yong-Ángel, G.; Avilés-Nova, F.; Rayas-Amor, A.A.; Solís-Méndez, A.D.; González-Ronquillo, M.; Vázquez-Carrillo, M.F.; Castelán-Ortega, O.A. Effect of Increasing Supplementation Levels of Coffee Pulp on Milk Yield and Food Intake in Dual-Purpose Cows: An Alternative Feed Byproduct for Smallholder Dairy Systems of Tropical Climate Regions. Agriculture 2021, 11, 416. [Google Scholar] [CrossRef]
- Focant, M.; Froidmont, E.; Archambeau, Q.; Dang Van, Q.C.; Larondelle, Y. The effect of oak tannin (Quercus robur) and hops (Humulus lupulus) on dietary nitrogen efficiency, methane emission, and milk fatty acid composition of dairy cows fed a low-protein diet including linseed. J. Dairy Sci. 2018, 102, 1144–1159. [Google Scholar] [CrossRef]
- Mapato, C.; Viennasay, B.; Cherdthong, A.; Wanapat, M. Milk production and composition efficiency as influenced by feeding Pennisetum purpureum cv. Mahasarakham with Tiliacora triandra, Diels pellet supplementation. Trop. Anim. Health Prod. 2021, 53, 64–73. [Google Scholar] [CrossRef]
- Aguerre, M.J.; Duval, B.; Powell, J.M.; Vadas, P.A.; Wattiaux, M.A. Effects of feeding a quebracho–chestnut tannin extract on lactating cow performance and nitrogen utilization efficiency. Int. J. Dairy Sci. 2019, 103, 2264–2271. [Google Scholar] [CrossRef]
- Herremans, S.; Decruyenaere, V.; Cantalapiedra-Hijar, G.; Beckers, Y.; Froidmont, E. Effects of hydrolysable tannin-treated grass silage on milk yield and composition, nitrogen partitioning and nitrogen isotopic discrimination in lactating dairy cows. Animal 2020, 14, 771–779. [Google Scholar] [CrossRef] [PubMed]
- Santos, F.S.; Zeoula, L.M.; De Lima, L.S.; De Marchi, F.E.; Ítavo, L.C.V.; Santos, N.W.; Pintro, P.M.; Damasceno, J.C.; dos Santos, G.T. Effect of supplementation with Yerba Mate (Ilex paraguariensis) and vitamin E on milk lipoperoxidation in cows receiving diets containing ground soybean seeds. J. Dairy Res. 2019, 86, 279–282. [Google Scholar] [CrossRef] [PubMed]
- Avila, A.S.; Zambom, M.A.; Faccenda, A.; Werle, C.H.; Almeida, A.R.E.; Schneider, C.R.; Grunevald, D.G.; Faciola, A.P. Black Wattle (Acacia mearnsii) Condensed Tannins as Feed Additives to Lactating Dairy Cows. Animals 2020, 10, 662. [Google Scholar] [CrossRef] [PubMed]
- Gerlach, K.; Pries, M.; Tholen, E.; Schmithausen, A.J.; Büscher, W.; Südekum, K.H. Effect of condensed tannins in rations of lactating dairy cows on production variables and nitrogen use efficiency. Animal 2018, 12, 1847–1855. [Google Scholar] [CrossRef] [PubMed]
- Alves, T.P.; Dall-Orsoletta, A.C.; Ribeiro-Filho, H.M.N. The effects of supplementing Acacia mearnsii tannin extract on dairy cow dry matter intake, milk production, and methane emission in a tropical pasture. Trop. Anim. Health Prod. 2017, 49, 1663–1668. [Google Scholar] [CrossRef]
- Alves, T.P.; Dias, K.M.; Dallastra, L.J.H.; Fonseca, B.L.; Ribeiro-Filho, H.M.N. Energy and tannin extract supplementation for dairy cows on annual winter pastures. Semin. Ciênc. Agrár. 2017, 38, 1017–1026. [Google Scholar] [CrossRef][Green Version]
- Orlandi, T.; Pozo, C.A.; Schiavo, J.; Oliveira, L.; Kozloski, G.V. Effect of using Acacia mearnsii tannin extract as a feed additive on nutritional variables and productive performance in dairy cows grazing a temperate pasture. J. Anim. Sci. 2020, 91, 1–7. [Google Scholar] [CrossRef]
- Williams, S.R.O.; Hannah, M.C.; Eckard, R.J.; Wales, W.J.; Moate, P.J. Supplementing the diet of dairy cows with fat or tannin reduces methane yield, and additively when fed in combination. Animal 2020, 14, 464–472. [Google Scholar] [CrossRef]
- Henke, A.; Dickhoefer, U.; Westreicher-Kristen, E.; Knappstein, K.; Molkentin, J.; Hasler, M.; Susenbeth, A. Effect of dietary Quebracho tannin extract on feed intake, digestibility, excretion of urinary purine derivatives and milk production in dairy cows. Arch Anim. Nutr. 2017, 71, 37–53. [Google Scholar] [CrossRef]
- Sallam, S.M.A.; Attia, M.F.A.; Nour El-Din, A.N.M.; El-Zarkouny, S.Z.; Saber, A.M.; El-Zaiat, H.M.; Zeitoun, M.M. Involvement of Quebracho tannins in diet alters productive and reproductive efficiency of postpartum buffalo cows. Anim. Nutr. 2018, 5, 80–86. [Google Scholar] [CrossRef]
- Aguerre, M.J.; Capozzolo, M.C.; Lencioni, P.; Cabral, C.; Wattiaux, M.A. Effect of quebracho-chestnut tannin extracts at 2 dietary crude protein levels on performance, rumen fermentation, and nitrogen partitioning in dairy cows. Int. J. Dairy Sci. 2016, 99, 4476–4486. [Google Scholar] [CrossRef] [PubMed]
- Menci, R.; Natalello, A.; Luciano, G.; Priolo, A.; Valenti, B.; Farina, G.; Caccamo, M.; Niderkorn, V.; Coppa, M. Effect of dietary tannin supplementation on cow milk quality in two different grazing seasons. Sci. Rep. 2021, 11, 19654. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Xu, X.; Cao, Z.; Wang, Y.; Yang, H.; Azarfar, A.; Li, S. Effect of Different Tannin Sources on Nutrient Intake, Digestibility, Performance, Nitrogen Utilization, and Blood Parameters in Dairy Cows. Animals 2019, 9, 507. [Google Scholar] [CrossRef] [PubMed]
- Kapp-Bitter, A.N.; Dickhoefer, U.; Suglo, E.; Baumgartner, L.; Kreuzer, M.; Leiber, F. Graded supplementation of chestnut tannins to dairy cows fed protein-rich spring pasture: Effects on indicators of protein utilization. J. Anim. Feed Sci. 2020, 29, 97–104. [Google Scholar] [CrossRef]
- Terranova, M.; Eggerschwiler, L.; Ortmann, S.; Clauss, M.; Kreuzer, M.; Schwarm, A. Increasing the proportion of hazel leaves in the diet of dairy cows reduced methane yield and excretion of nitrogen in volatile form, but not milk yield. Anim. Feed Sci. Technol. 2021, 276, 114790–114803. [Google Scholar] [CrossRef]
- Renna, M.; Lussiana, C.; Malfatto, V.; Gerbelle, M.; Turille, G.; Medana, C.; Ghirardello, D.; Mimosi, A.; Cornale, P. Evaluating the Suitability of Hazelnut Skin as a Feed Ingredient in the Diet of Dairy Cows. Animals 2020, 10, 1653. [Google Scholar] [CrossRef]
- Santos, N.W.; Yoshimura, E.H.; Machado, E.; Matumoto-Pintro, P.T.; Montanher, P.F.; Visentainer, J.V.; dos Santos, G.T.; Zeoula, L.M. Antioxidant effects of a propolis extract and vitamin E in blood and milk of dairy cows fed diet containing flaxseed oil. Livest. Sci. 2016, 191, 132–138. [Google Scholar] [CrossRef]
- Ineichen, S.; Kuenzler, A.D.; Kreuzer, M.; Marquardt, S.; Reidy, B. Digestibility, nitrogen utilization and milk fatty acid profile of dairy cows fed hay from species rich mountainous grasslands with elevated herbal and phenolic contents. Anim. Feed Sci. Technol. 2018, 247, 210–221. [Google Scholar] [CrossRef]
- Bryszak, M.; Szumacher-Strabel, M.; El-Sherbiny, M.; Stochmal, A.; Oleszek, W.; Roj, E.; Patra, A.K.; Cieslak, A. Effects of berry seed residues on ruminal fermentation, methane concentration, milk production, and fatty acid proportions in the rumen and milk of dairy cows. Int. J. Dairy Sci. 2019, 102, 1257–1273. [Google Scholar] [CrossRef]
- Grosse Brinkhaus, A.; Bee, G.; Silacci, P.; Kreuzer, M.; Dohme-Meier, F. Effect of exchanging Onobrychis viciifolia and Lotus corniculatus for Medicago sativa on ruminal fermentation and nitrogen turnover in dairy cows. J. Dairy Sci. 2016, 99, 4384–4397. [Google Scholar] [CrossRef]
- Girard, M.; Dohme-Meier, F.; Wechsler, D.; Goy, D.; Kreuzer, M.; Bee, G. Ability of 3 tanniferous forage legumes to modify quality of milk and Gruyère-type cheese. J. Dairy Sci. 2016, 99, 205–220. [Google Scholar] [CrossRef] [PubMed]
- Kapp-Bitter, A.N.; Dickhoefer, U.; Kaptijn, G.; Pedan, V.; Perler, E.; Kreuzer, M.; Leiber, F. On-farm examination of sainfoin supplementation effects in dairy cows in a roughage-based feeding system: Indicators of protein utilisation. Livest. Sci. 2021, 248, 104509–104516. [Google Scholar] [CrossRef]
- Moats, J.; Mutsvangwa, T.; Refat, B.; Christensen, D.A. Evaluation of whole flaxseed and the use of tannin-containing fava beans as an alternative to peas in a co-extruded flaxseed product on ruminal fermentation, selected milk fatty acids, and production in dairy cows. Prof. Anim. Sci. 2018, 34, 435–446. [Google Scholar] [CrossRef]
- Broderick, G.A.; Grabber, J.H.; Muck, R.E.; Hymes-Fecht, U.C. Replacing alfalfa silage with tannin-containing birdsfoot trefoil silage in total mixed rations for lactating dairy cows. J. Dairy Sci. 2017, 100, 3548–3562. [Google Scholar] [CrossRef] [PubMed]
- Lopreiato, V.; Mezzetti, M.; Cattaneo, L.; Ferronato, G.; Minuti, A.; Trevisi, E. Role of nutraceuticals during the transition period of dairy cows: A review. J. Anim. Sci. Biotechnol. 2020, 11, 96–113. [Google Scholar] [CrossRef] [PubMed]
- Kasparovska, J.; Pecinkova, M.; Dadakova, K.; Krizova, L.; Hadrova, S.; Lexa, M.; Lochman, J.; Kasparovsky, T. Effects of Isoflavone-Enriched Feed on the Rumen Microbiota in Dairy Cows. PLoS ONE 2016, 11, e0154642–e0154658. [Google Scholar] [CrossRef]
- Třináctý, J.; Křížová, L.; Schulzová, V.; Hajšlová, J.; Hanuš, O. The effect of feeding soybean-derived phytoestogens on their concentration in plasma and milk of lactating dairy cows. Arch. Anim. Nutr. 2009, 63, 219–229. [Google Scholar] [CrossRef]
- Flachowsky, G.; Hünerberg, M.; Meyer, U.; Kammerer, D.R.; Carle, R.; Goerke, M.; Eklund, M. Isoflavone concentration of soybean meal from various origins and transfer of isoflavones into milk of dairy cows. J. Verbrauch. Lebensm. 2011, 6, 449–456. [Google Scholar] [CrossRef]
- Andersen, C.; Nielsen, T.S.; Purup, S.; Kristensen, T.; Eriksen, J.; Søegaard, K.; Sørensen, J.; Fretté, X.C. Phyto-oestrogens in herbage and milk from cows grazing white clover, red clover, lucerne or chicory-rich pastures. Animal 2009, 3, 1189–1195. [Google Scholar] [CrossRef]
- Tsen, S.Y.; Siew, J.; Lau, E.K.L.; Afiqah bte Roslee, F.; Chan, H.M.; Loke, W.M. Cow’s milk as a dietary source of equol and phenolic antioxidants: Differential distribution in the milk aqueous and lipid fractions. Dairy Sci. Technol. 2014, 94, 625–632. [Google Scholar] [CrossRef]
- Steinshamn, H.; Purup, S.; Thuen, E.; Hansen-Møller, J. Effects of Clover-Grass Silages and Concentrate Supplementation on the Content of Phytoestrogens in Dairy Cow Milk. J. Dairy Sci. 2008, 91, 2715–2725. [Google Scholar] [CrossRef]
- Gagnon, N.; Côrtes, C.; Da Silva, D.; Kazama, R.; Benchaar, C.; dos Santos, G.; Zeoula, L.; Petit, H.V. Ruminal metabolism of flaxseed (Linum usitatissimum) lignans to the mammalian lignan enterolactone and its concentration in ruminal fluid, plasma, urine and milk of dairy cows. Br. J. Nutr. 2009, 102, 1015–1023. [Google Scholar] [CrossRef] [PubMed]
- Antignac, J.P.; Cariou, R.; Le Bizec, B.; André, F. New data regarding phytoestrogens content in bovine milk. Food Chem. 2004, 87, 275–281. [Google Scholar] [CrossRef]
- Jansson, T.; Rauh, V.; Danielsen, B.P.; Poojary, M.M.; Waehrens, S.S.; Bredie, W.L.P.; Sørensen, J.; Petersen, M.A.; Ray, C.A.; Lund, M.N. Green Tea Polyphenols Decrease Strecker Aldehydes and Bind to Proteins in Lactose-Hydrolyzed UHT Milk. J. Agric. Food Chem. 2017, 65, 10550–10561. [Google Scholar] [CrossRef] [PubMed]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Milk Composition | |||||
|---|---|---|---|---|---|
| Plant/Extract | %Milk Yield vs. CD | Fat % | Protein % | Lactose % | References |
| Acacia mearnsii TE | - | - | - | [88] | |
| - | - | - | |||
| - | - | - | |||
| Acacia mearnsii TE | 2.56 | 3.03 | 4.46 | [143] | |
| Acacia mearnsii TE | 2.94 | 3.37 | - | [142] | |
| Acacia mearnsii TE | 3.59 | 3.35 | - | [141] | |
| Acacia mearnsii TE | - | - | - | [139] | |
| - | - | - | |||
| - | - | - | |||
| - | - | - | |||
| Acacia mearnsii TE | 3.88 | 3.35 | - | [140] | |
| 3.80 | 3.22 | - | |||
| Acacia mangium TE | 3.33 | 3.13 | 4.80 | [149] | |
| Myrica sp. TE | 3.39 | 3.11 | 4.81 | [149] | |
| Castanea sativa TE | 3.67 | 3.32 | 4.88 | [150] | |
| 3.57 | 3.24 | 4.77 | |||
| Schinopsis spp./C. sativa TE | 3.6 | 2.92 | 4.86 | [147] | |
| 3.51 | 2.86 | 4.87 | |||
| 3.57 | 2.83 | 4.90 | |||
| Schinopsis spp./C. sativa TE | 1.89 | 1.50 | 2.27 | [136] | |
| 1.85 | 1.41 | 2.24 | |||
| Schinopsis spp./C. sativa TE | 3.97 | 3.92 | 4.6 | [148] | |
| Schinopsis spp. TE | 3.33 | 3.29 | 4.69 | [145] | |
| 3.43 | 3.29 | 4.67 | |||
| Schinopsis spp. TE | 5.93 | 4.30 | 5.23 | [146] | |
| 6.08 | 3.94 | 4.87 | |||
| Quercus robur TE | 4.01 | 3.26 | - | [134] | |
| Quercus robur TE | - | - | - | [137] | |
| Onobrychis vicifolia TE | 1.47 | 1.12 | 1.97 | [156] | |
| Onobrychis vicifolia TE | 4.10 | 3.20 | 5.00 | [157] | |
| Quercus aegylops TE | 3.42 | 3.01 | 4.74 | [149] | |
| Tannins/Capsicum spp. | 3.96 | 3.03 | 4.90 | [131] | |
| Tanimil SCC | 3.73 | 3.20 | 4.63 | [132] | |
| Scutellaria baicalensis FE (trited based on baicalein) | 3.84 | 3.10 | 4.89 | [120] | |
| 3.08 | 2.89 | 4.95 | |||
| 3.84 | 3.12 | 4.95 | |||
| 3.29 | 2.97 | 4.97 | |||
| Propolis extract | - | - | - | [153] | |
| Propolis extract + Vit. E | - | - | - | ||
| Citrus pulp | - | - | - | [126] | |
| Birdsfoot trefoil | 3.93 | 3.14 | 4.76 | [160] | |
| 3.96 | 3.10 | 4.76 | |||
| 3.78 | 3.10 | 4.77 | |||
| Coffee pulp | - | - | - | [133] | |
| - | - | - | |||
| - | - | - | |||
| Dried grape pomace | 3.83 | 3.32 | 4.84 | [125] | |
| Durum wheat bran | 4.05 | 3.5 | 4.98 | [123] | |
| 4.01 | 3.6 | 4.92 | |||
| Hazelnut skin | - | - | - | [152] | |
| Mao pomace meal | 4.6 | 3.1 | 4.5 | [130] | |
| 4.4 | 3.1 | 4.6 | |||
| 4.5 | 3.2 | 4.5 | |||
| Onobrychis vicifolia | 4.33 | 3.69 | 4.53 | [158] | |
| 4.38 | 3.88 | 4.57 | |||
| 4.20 | 3.52 | 4.69 | |||
| 4.29 | 3.75 | 4.69 | |||
| Paper bulberry silage | 4.3 | 3.7 | 5.1 | [121] | |
| 4.2 | 3.6 | 5.0 | |||
| 3.9 | 3.6 | 5.1 | |||
| 4.1 | 3.6 | 5.1 | |||
| Rice straw/bamboo grass pellet | 3.5 | 3.1 | 4.7 | [135] | |
| Sweet grass/bamboo grass pellet | 4.0 | 3.3 | 4.6 | [135] | |
| Yerba mate | - | - | - | [138] | |
| Yerba mate + Vit. E | - | - | - | ||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Formato, M.; Cimmino, G.; Brahmi-Chendouh, N.; Piccolella, S.; Pacifico, S. Polyphenols for Livestock Feed: Sustainable Perspectives for Animal Husbandry? Molecules 2022, 27, 7752. https://doi.org/10.3390/molecules27227752
Formato M, Cimmino G, Brahmi-Chendouh N, Piccolella S, Pacifico S. Polyphenols for Livestock Feed: Sustainable Perspectives for Animal Husbandry? Molecules. 2022; 27(22):7752. https://doi.org/10.3390/molecules27227752
Chicago/Turabian StyleFormato, Marialuisa, Giovanna Cimmino, Nabila Brahmi-Chendouh, Simona Piccolella, and Severina Pacifico. 2022. "Polyphenols for Livestock Feed: Sustainable Perspectives for Animal Husbandry?" Molecules 27, no. 22: 7752. https://doi.org/10.3390/molecules27227752
APA StyleFormato, M., Cimmino, G., Brahmi-Chendouh, N., Piccolella, S., & Pacifico, S. (2022). Polyphenols for Livestock Feed: Sustainable Perspectives for Animal Husbandry? Molecules, 27(22), 7752. https://doi.org/10.3390/molecules27227752