
Structural Investigation of Betulinic Acid Plasma Metabolites by Tandem Mass Spectrometry

.jpg)
 , , , ,
, , , ,  , ,
, , 
Abstract
1. Introduction
2. Results and Discussion
2.1. Screening of BA and BA Metabolites by Negative Ion Mode ESI Orbitrap MS in Plasma Samples
2.1.1. Phase I Metabolic Products
2.1.2. Phase II Metabolic Products
2.2. Structural Analysis of BA and BA Metabolites by Negative Ion Mode ESI Orbitratp MS/MS in Plasma Samples
2.2.1. Structural Analysis of BA Isolated from Plasma Samples
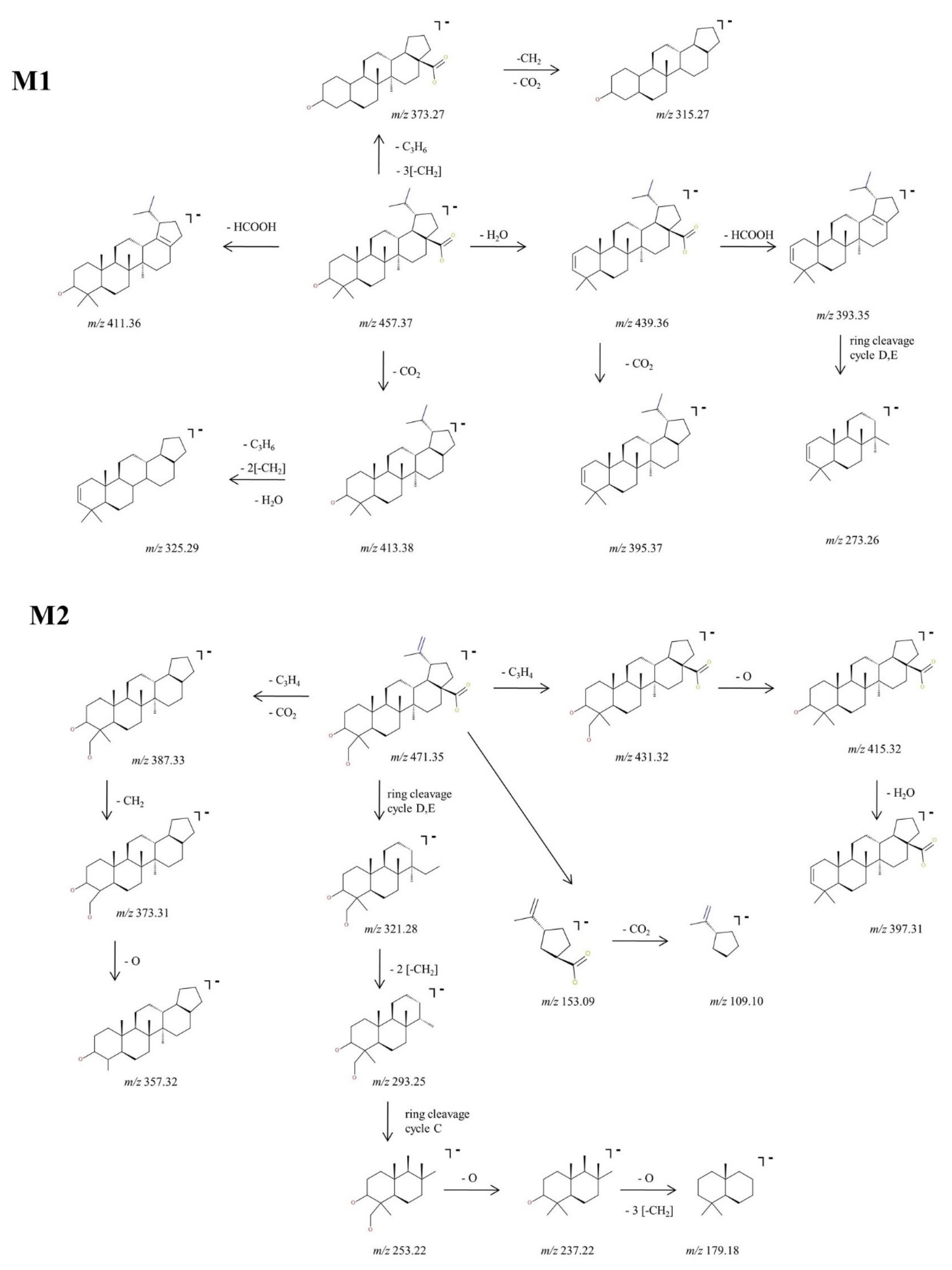
2.2.2. Structural Analysis of Phase I Metabolites
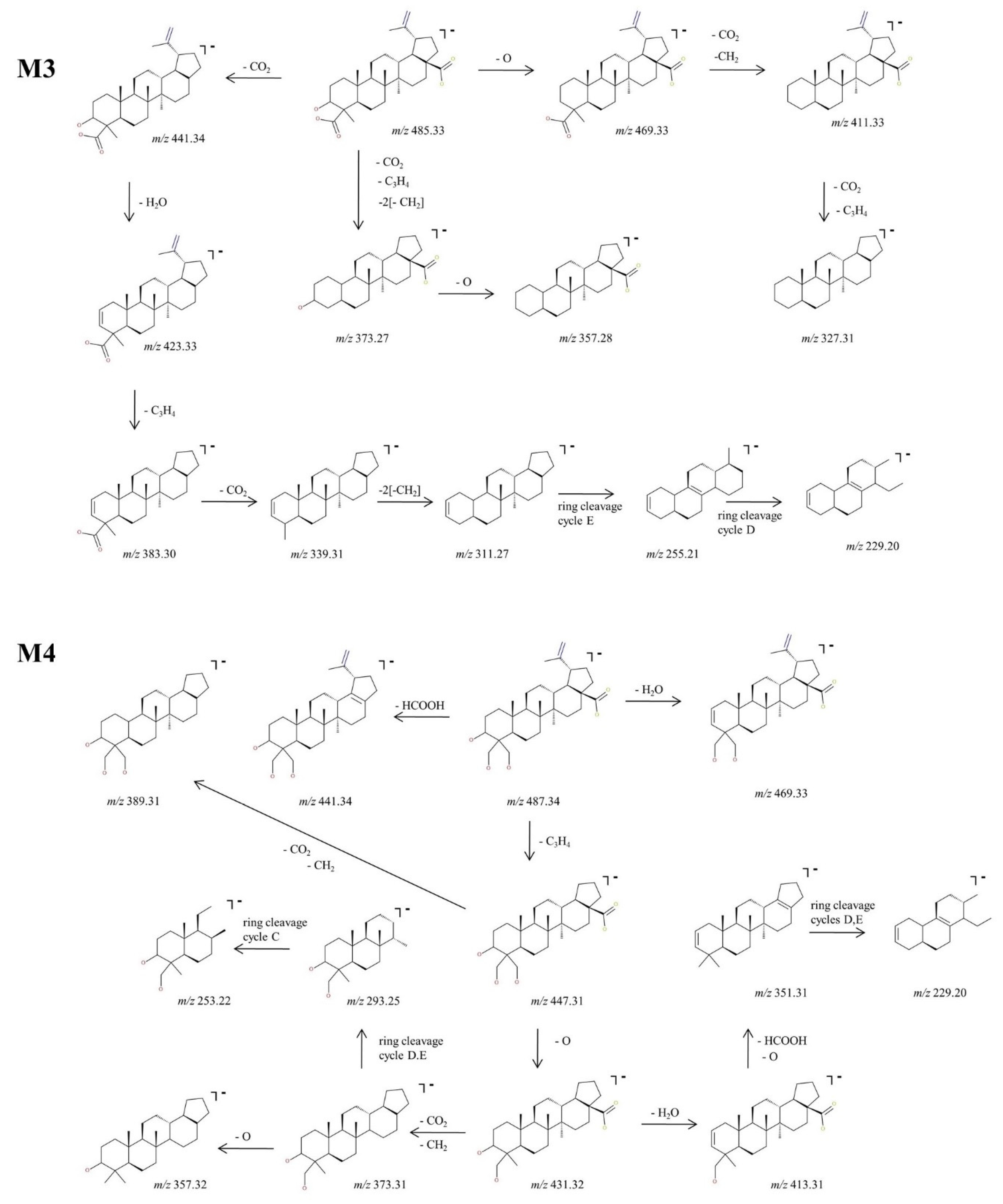
2.2.3. Structural Analysis of Phase II Metabolites
Structural Analysis of Sulphated Metabolites
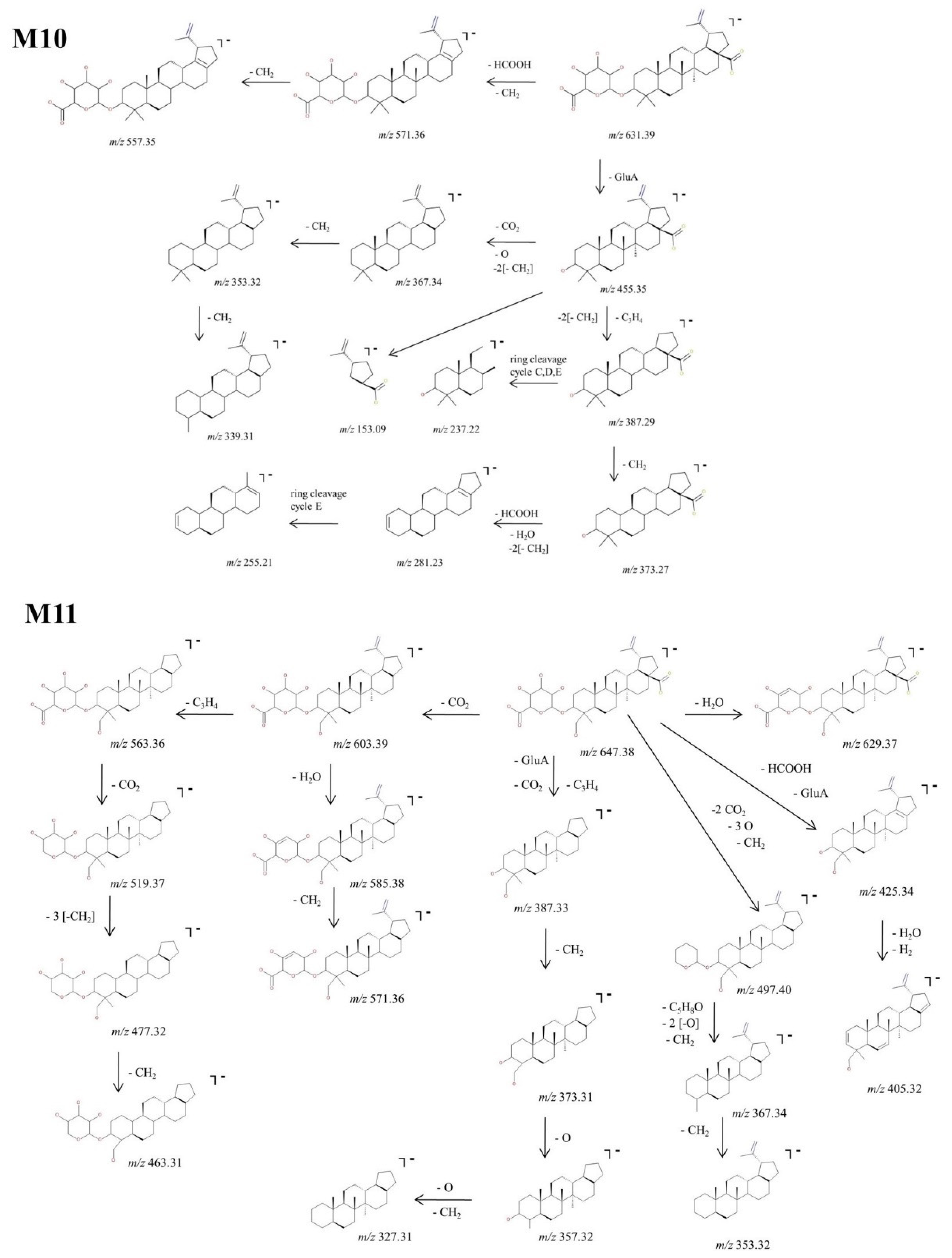
Structural Analysis of Glucuronated Metabolites
2.3. Proposed Metabolic Pathway for BA in SKH1 Female Mice
3. Materials and Methods
3.1. Reagents
3.1.1. Chemicals
3.1.2. BA Nanoemulsion
3.2. In Vivo Experiment
3.3. Plasma Samples
3.4. Orbitrap Mass Spectrometry
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Fulda, S. Betulinic acid for cancer treatment and prevention. Int. J. Mol. Sci. 2008, 9, 1096–1107. [Google Scholar] [CrossRef]
- Guo, S.; Duan, J.A.; Tang, Y.P.; Yang, N.Y.; Qian, D.W.; Su, S.L.; Shang, E.X. Characterization of triterpenic acids in fruits of ziziphus species by HPLC-ELSD-MS. J. Agric. Food Chem. 2010, 58, 6285–6289. [Google Scholar] [CrossRef] [PubMed]
- Karan, B.N.; Maity, T.K.; Pal, B.C.; Singha, T.; Jana, S. Betulinic Acid, the first lupane-type triterpenoid isolated via bioactivity-guided fractionation, and identified by spectroscopic analysis from leaves of Nyctanthes arbor-tristis: Its potential biological activities in vitro assays. Nat. Prod. Res. 2019, 33, 3287–3292. [Google Scholar] [CrossRef] [PubMed]
- Mioc, M.; Prodea, A.; Racoviceanu, R.; Mioc, A.; Ghiulai, R.; Milan, A.; Voicu, M.; Mardale, G.; Șoica, C. Recent Advances Regarding the Molecular Mechanisms of Triterpenic Acids: A Review (Part II). Int. J. Mol. Sci. 2022, 23, 8896. [Google Scholar] [CrossRef] [PubMed]
- Hordyjewska, A.; Ostapiuk, A.; Horecka, A.; Kurzepa, J. Betulin and betulinic acid: Triterpenoids derivatives with a powerful biological potential. Phytochem. Rev. 2019, 18, 929–951. [Google Scholar] [CrossRef]
- Coricovac, D.; Dehelean, C.A.; Pinzaru, I.; Mioc, A.; Aburel, O.M.; Macasoi, I.; Draghici, G.A.; Petean, C.; Soica, C.; Boruga, M.; et al. Assessment of Betulinic Acid Cytotoxicity and Mitochondrial Metabolism Impairment in a Human Melanoma Cell Line. Int. J. Mol. Sci. 2021, 22, 4870. [Google Scholar] [CrossRef]
- Zeng, A.; Hua, H.; Liu, L.; Zhao, J. Betulinic acid induces apoptosis and inhibits metastasis of human colorectal cancer cells in vitro and in vivo. Bioorganic Med. Chem. 2019, 27, 2546–2552. [Google Scholar] [CrossRef] [PubMed]
- Lee, D.; Lee, S.R.; Kang, K.S.; Ko, Y.; Pang, C.; Yamabe, N.; Kim, K.H. Betulinic Acid Suppresses Ovarian Cancer Cell Proliferation through Induction of Apoptosis. Biomolecules 2019, 9, 257. [Google Scholar] [CrossRef] [PubMed]
- Xu, T.; Pang, Q.; Wang, Y.; Yan, X. Betulinic acid induces apoptosis by regulating PI3K/Akt signaling and mitochondrial pathways in human cervical cancer cells. Int. J. Mol. Med. 2017, 40, 1669–1678. [Google Scholar] [CrossRef] [PubMed]
- Zeng, A.Q.; Yu, Y.; Yao, Y.Q.; Yang, F.F.; Liao, M.; Song, L.J.; Li, Y.L.; Yu, Y.; Li, Y.J.; Deng, Y.L.; et al. Betulinic acid impairs metastasis and reduces immunosuppressive cells in breast cancer models. Oncotarget 2018, 9, 3794–3804. [Google Scholar] [CrossRef]
- Oliveira-Costa, J.F.; Meira, C.S.; Neves, M.V.G.D.; Dos Reis, B.P.Z.C.; Soares, M.B.P. Anti-Inflammatory Activities of Betulinic Acid: A Review. Front. Pharmacol. 2022, 13, 1866. [Google Scholar] [CrossRef] [PubMed]
- Bildziukevich, U.; Özdemir, Z.; Wimmer, Z. Recent achievements in medicinal and supramolecular chemistry of betulinic acid and its derivatives. Molecules 2019, 24, 3546. [Google Scholar] [CrossRef] [PubMed]
- Ríos, J.L.; Máñez, S. New Pharmacological Opportunities for Betulinic Acid. Planta Med. 2018, 84, 8–19. [Google Scholar] [CrossRef] [PubMed]
- Ghiulai, R.; Roşca, O.J.; Antal, D.S.; Mioc, M.; Mioc, A.; Racoviceanu, R.; Macaşoi, I.; Olariu, T.; Dehelean, C.; Creţu, O.M.; et al. Tetracyclic and pentacyclic triterpenes with high therapeutic efficiency in wound healing approaches. Molecules 2020, 25, 5557. [Google Scholar] [CrossRef] [PubMed]
- Soica, C.; Danciu, C.; Savoiu-Balint, G.; Borcan, F.; Ambrus, R.; Zupko, I.; Bojin, F.; Coricovac, D.; Ciurlea, S.; Avram, S.; et al. Betulinic Acid in Complex with a Gamma-Cyclodextrin Derivative Decreases Proliferation and in vivo Tumor Development of Non-Metastatic and Metastatic B164A5 Cells. Int. J. Mol. Sci. 2014, 15, 8235–8255. [Google Scholar] [CrossRef] [PubMed]
- Saneja, A.; Arora, D.; Kumar, R.; Dubey, R.D.; Panda, A.K.; Gupta, P.N. Therapeutic applications of betulinic acid nanoformulations. Ann. N. Y. Acad. Sci. 2018, 1421, 5–18. [Google Scholar] [CrossRef] [PubMed]
- Roberts, S.A. Drug metabolism and pharmacokinetics in drug discovery. Curr. Opin. Drug Discov. Dev. 2003, 6, 66–80. [Google Scholar]
- Bachmann, K. Drug Metabolism, 1st ed.; Elsevier Inc.: Amsterdam, The Netherlands, 2009; ISBN 9780123695215. [Google Scholar]
- Scott Obach, R. Pharmacologically active drug metabolites: Impact on drug discovery and pharmacotherapy. Pharmacol. Rev. 2013, 65, 578–640. [Google Scholar] [CrossRef]
- Udeani, G.O.; Zhao, G.M.; Shin, Y.G.; Cooke, B.P.; Graham, J.; Beecher, C.W.W.; Kinghorn, A.D.; Pezzuto, J.M. Pharmacokinetics and tissue distribution of betulinic acid in CD-1 mice. Biopharm. Drug Dispos. 1999, 20, 379–383. [Google Scholar] [CrossRef]
- Cheng, X.; Shin, Y.G.; Levine, B.S.; Smith, A.C.; Tomaszewski, J.E.; Van Breemen, R.B. Quantitative analysis of betulinic acid in mouse, rat and dog plasma using electrospray liquid chromatography/mass spectrometry. Rapid Commun. Mass Spectrom. 2003, 17, 2089–2092. [Google Scholar] [CrossRef] [PubMed]
- Zhang, W.D.; Jin, M.M.; Jiang, H.H.; Yang, J.X.; Wang, Q.; Du, Y.F.; Cao, L.; Xu, H.J. Study on the metabolites of betulinic acid in vivo and in vitro by ultra high performance liquid chromatography with time-of-flight mass spectrometry. J. Sep. Sci. 2019, 42, 628–635. [Google Scholar] [CrossRef] [PubMed]
- Li, M.; Zhang, F.X.; Wei, Z.C.; Li, Z.T.; Zhang, G.X.; Li, H.J. Systematically characterization of in vivo substances of Ziziphi Spinosae Semen in rats by ultra-high-performance liquid chromatography coupled with quadrupole time-of-flight tandem mass spectrometry analysis. J. Pharm. Biomed. Anal. 2021, 193, 113756. [Google Scholar] [CrossRef] [PubMed]
- Nagana Gowda, G.A.; Djukovic, D. Overview of mass spectrometry-based metabolomics: Opportunities and challenges. Methods Mol. Biol. 2014, 1198, 3–12. [Google Scholar] [CrossRef]
- Gouveia, M.J.; Brindley, P.J.; Santos, L.L.; Correia Da Costa, J.M.; Gomes, P.; Vale, N. Mass spectrometry techniques in the survey of steroid metabolites as potential disease biomarkers: A review. Metabolism 2013, 62, 1206–1217. [Google Scholar] [CrossRef] [PubMed]
- Segers, K.; Declerck, S.; Mangelings, D.; Heyden, Y.V.; Eeckhaut, A.V. Analytical techniques for metabolomic studies: A review. Bioanalysis 2019, 11, 2297–2318. [Google Scholar] [CrossRef] [PubMed]
- Dinda, B.; Chakraborty, A. Pharmacokinetics and Metabolism of Phytochemicals Having Anti-obesity and Antidiabetic Activity. In Natural Products in Obesity and Diabetes: Therapeutic Potential and Role in Prevention and Treatment; Dinda, B., Ed.; Springer International Publishing: Cham, Switzerland, 2022; pp. 469–505. ISBN 978-3-030-92196-5. [Google Scholar]
- Song, Y.; Li, C.; Liu, G.; Liu, R.; Chen, Y.; Li, W.; Cao, Z.; Zhao, B.; Lu, C.; Liu, Y. Drug-Metabolizing Cytochrome P450 Enzymes Have Multifarious Influences on Treatment Outcomes. Clin. Pharmacokinet. 2021, 60, 585–601. [Google Scholar] [CrossRef] [PubMed]
- Strathmann, J. Bioavailability and metabolism of curcuminoids. In Natural Compounds as Inducers of Cell Death; Springer: Dordrecht, The Netherlands, 2012; Volume 47, pp. 95–124. [Google Scholar] [CrossRef]
- García, C.J.; Beltrán, D.; Tomás-Barberán, F.A. Human Gut Microbiota Metabolism of Dietary Sesquiterpene Lactones: Untargeted Metabolomics Study of Lactucopicrin and Lactucin Conversion In Vitro and In Vivo. Mol. Nutr. Food Res. 2020, 64, 2000619. [Google Scholar] [CrossRef]
- Zhang, Y.F.; Liu, L.J.; Xu, F.; Shang, M.Y.; Liu, G.X.; Cai, S.Q. Investigation of the in vivo metabolism of sibirioside A and Angoroside C in Rats by HPLC-ESI-IT-TOF-MSn. Molecules 2018, 23, 2702. [Google Scholar] [CrossRef] [PubMed]
- El-Haj, B.M.; Ahmed, S.B.M. Metabolic-hydroxy and carboxy functionalization of alkyl moieties in drug molecules: Prediction of structure influence and pharmacologic activity. Molecules 2020, 25, 1937. [Google Scholar] [CrossRef] [PubMed]
- Xia, B.; Bai, L.; Li, X.; Xiong, J.; Xu, P.; Xue, M. Structural analysis of metabolites of asiatic acid and its analogue madecassic acid in zebrafish using LC/IT-MSn. Molecules 2015, 20, 3001–3019. [Google Scholar] [CrossRef]
- Lozano-Mena, G.; Sánchez-González, M.; Parra, A.; Juan, M.E.; Planas, J.M. Identification of gut-derived metabolites of maslinic acid, a bioactive compound from Olea europaea L. Mol. Nutr. Food Res. 2016, 60, 2053–2064. [Google Scholar] [CrossRef] [PubMed]
- Chatterjee, P.; Kouzi, S.A.; Pezzuto, J.M.; Hamann, M.T. Biotransformation of the antimelanoma agent betulinic acid by Bacillus megaterium ATCC 13368. Appl. Environ. Microbiol. 2000, 66, 3850–3855. [Google Scholar] [CrossRef] [PubMed]
- De Sousa, I.P.; Sousa Teixeira, M.V.; Jacometti Cardoso Furtado, N.A. An overview of biotransformation and toxicity of diterpenes. Molecules 2018, 23, 1387. [Google Scholar] [CrossRef]
- Guan, H.; Li, K.; Wang, X.; Luo, X.; Su, M.; Tan, W.; Chang, X.; Shi, Y. Identification of metabolites of the cardioprotective alkaloid dehydrocorydaline in rat plasma & bile by liquid chromatography coupled with triple quadrupole linear ion trap mass spectrometry. Molecules 2017, 22, 1686. [Google Scholar] [CrossRef]
- Janov, P.; Šiller, M. Phase II Drug Metabolism. Top. Drug Metab. 2012, 35–60, 29241. [Google Scholar] [CrossRef]
- Li, M.; Si, D.; Fu, Z.; Sang, M.; Zhang, Z.; Liu, E.; Yang, W.; Gao, X.; Han, L. Enhanced identification of the in vivo metabolites of Ecliptae Herba in rat plasma by integrating untargeted data-dependent MS 2 and predictive multiple reaction monitoring-information dependent acquisition-enhanced product ion scan. J. Chromatogr. B Anal. Technol. Biomed. Life Sci. 2019, 1109, 99–111. [Google Scholar] [CrossRef]
- Gamage, N.; Barnett, A.; Hempel, N.; Duggleby, R.G.; Windmill, K.F.; Martin, J.L.; McManus, M.E. Human sulfotransferases and their role in chemical metabolism. Toxicol. Sci. 2006, 90, 5–22. [Google Scholar] [CrossRef]
- Zhang, W.; Jiang, H.; Jin, M.; Wang, Q.; Sun, Q.; Du, Y.; Cao, L.; Xu, H. UHPLC-Q-TOF-MS/MS based screening and identification of the metabolites in vivo after oral administration of betulin. Fitoterapia 2018, 127, 29–41. [Google Scholar] [CrossRef]
- Wang, L.; Li, S.; Li, J.; Cheng, Z.; Feng, Y.; Ouyang, H.; Du, Z.; Jiang, H. Comprehensive metabolic profiling of Alismatis Rhizoma triterpenes in rats based on characteristic ions and a triterpene database. J. Pharm. Anal. 2021, 11, 96–107. [Google Scholar] [CrossRef]
- Kutsuno, Y.; Itoh, T.; Tukey, R.H.; Fujiwara, R. Glucuronidation of drugs and drug-induced toxicity in humanized UDP-glucuronosyltransferase 1 mice. Drug Metab. Dispos. 2014, 42, 1146–1152. [Google Scholar] [CrossRef]
- Hu, X.; Shen, Y.; Yang, S.; Lei, W.; Luo, C.; Hou, Y.; Bai, G. Metabolite identification of ursolic acid in mouse plasma and urine after oral administration by ultra-high performance liquid chromatography/quadrupole time-of-flight mass spectrometry. RSC Adv. 2018, 8, 6532–6539. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Zuo, Z.; Lin, G. Intestinal and hepatic glucuronidation of flavonoids. Mol. Pharm. 2007, 4, 833–845. [Google Scholar] [CrossRef] [PubMed]
- Hu, Z.; Wang, Z.; Luo, S.; James, M.O.; Wang, Y. Phase II metabolism of betulin by rat and human UDP-glucuronosyltransferases and sulfotransferases. Chem. Biol. Interact. 2019, 302, 190–195. [Google Scholar] [CrossRef] [PubMed]
- Cao, F.R.; Feng, L.; Ye, L.H.; Wang, L.S.; Xiao, B.X.; Tao, X.; Chang, Q. Ganoderic acid A metabolites and their metabolic kinetics. Front. Pharmacol. 2017, 8, 101. [Google Scholar] [CrossRef] [PubMed]
- Sut, S.; Poloniato, G.; Malagoli, M.; Dall’acqua, S. Fragmentation of the main triterpene acids of apple by LC- APCI-MSn. J. Mass Spectrom. 2018, 53, 882–892. [Google Scholar] [CrossRef]
- Shao, S.Y.; Ting, Y.; Wang, J.; Sun, J.; Guo, X.F. Characterization and Identification of the Major Flavonoids in Phyllostachys edulis Leaf Extract by UPLC-QTOF-MS/MS. Acta Chromatogr. 2020, 32, 228–237. [Google Scholar] [CrossRef]
- Li, Y.; Meng, W.; Yuan, L.; Jiang, L.; Zhou, Z.; Chi, M.; Gong, Z.; Ma, X.; Huang, Y.; Zheng, L. Identification of Protosappanoside D from Caesalpinia decapetala and Evaluation of Its Pharmacokinetic, Metabolism and Pharmacological Activity. Molecules 2022, 27, 6090. [Google Scholar] [CrossRef]
- Fitzgerald, C.C.J.; Hedman, R.; Uduwela, D.R.; Paszerbovics, B.; Carroll, A.J.; Neeman, T.; Cawley, A.; Brooker, L.; McLeod, M.D. Profiling Urinary Sulfate Metabolites with Mass Spectrometry. Front. Mol. Biosci. 2022, 9, 829511. [Google Scholar] [CrossRef]
- Rincon Nigro, M.E.; Du, T.; Gao, S.; Kaur, M.; Xie, H.; Olaleye, O.A.; Liang, D. Metabolite Identification of a Novel Anti-Leishmanial Agent OJT007 in Rat Liver Microsomes Using LC-MS/MS. Molecules 2022, 27, 2854. [Google Scholar] [CrossRef]
- Dehelean, C.A.; Feflea, S.; Ganta, S.; Amiji, M. Anti-angiogenic effects of betulinic acid administered in nanoemulsion formulation using chorioallantoic membrane assay. J. Biomed. Nanotechnol. 2011, 7, 317–324. [Google Scholar] [CrossRef]
- Parasuraman, S.; Raveendran, R.; Kesavan, R. Blood sample collection in small laboratory animals. J. Pharmacol. Pharmacother. 2010, 1, 87–93. [Google Scholar] [CrossRef] [PubMed]
- Liigand, P.; Kaupmees, K.; Haav, K.; Liigand, J.; Leito, I.; Girod, M.; Antoine, R.; Kruve, A. Think Negative: Finding the Best Electrospray Ionization/MS Mode for Your Analyte. Anal. Chem. 2017, 89, 5665–5668. [Google Scholar] [CrossRef] [PubMed]












{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
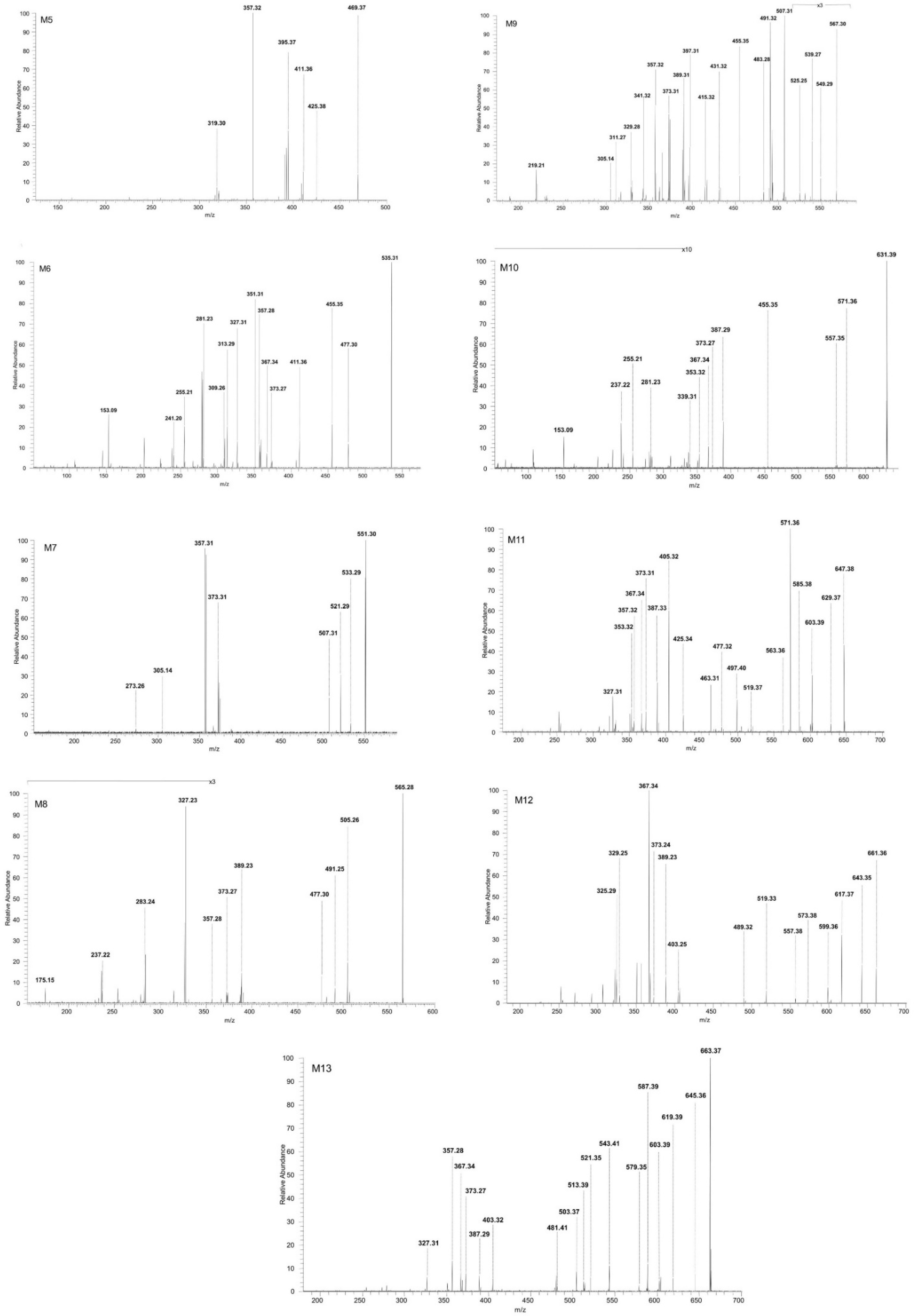
| Metabolite | Molecular Formula | m/z | Molecular Ion | Metabolic Reaction | Molecular Alteration | Mass Change |
|---|---|---|---|---|---|---|
| M 1 | C30H50O3 | 457.37 | [M-H+]− | -hydrogenation | +2H | +2 Da |
| M 2 | C30H48O4 | 471.35 | [M-H+]− | -monohydroxylation | +O | +16 Da |
| M 3 | C30H46O5 | 485.33 | [M-H+]− | -oxydation | +2O −2H | +30 Da |
| M 4 | C30H48O5 | 487.34 | [M-H+]− | -dihydroxylation | +2O | +32 Da |
| M 5 | C31H50O3 | 469.37 | [M-H+]− | -methylation | +CH2 | +14 Da |
| M 6 | C30H48O6S | 535.31 | [M-H+]− | -sulfoconjugation | +SO3 | +80 Da |
| M 7 | C30H48O7S | 551.30 | [M-H+]− | -hydroxylation -sulfoconjugation | +O +SO3 | +96 Da |
| M 8 | C30H46O8S | 565.28 | [M-H+]− | -oxidation -sulfoconjugation | +2O −2H +SO3 | +110 Da |
| M 9 | C30H48O8S | 567.30 | [M-H+]− | -dihydroxylation -sulfoconjugation | +2O +SO3 | +112 Da |
| M 10 | C36H56O9 | 631.39 | [M-H+]− | -glucuronidation | +GluA | +176 Da |
| M 11 | C36H56O10 | 647.38 | [M-H+]− | -hydroxylation -glucuronidation | +O +GluA | +192 Da |
| M 12 | C36H54O11 | 661.36 | [M-H+]− | -oxydation -glucuronidation | +2O −2H +GluA | +206 Da |
| M 13 | C36H56O11 | 663.37 | [M-H+]− | -dihydroxylation -glucuronidation | +2O +GluA | +208 Da |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ghiulai, R.; Mioc, M.; Racoviceanu, R.; Prodea, A.; Milan, A.; Coricovac, D.; Dehelean, C.; Avram, Ș.; Zamfir, A.D.; Munteanu, C.V.A.; et al. Structural Investigation of Betulinic Acid Plasma Metabolites by Tandem Mass Spectrometry. Molecules 2022, 27, 7359. https://doi.org/10.3390/molecules27217359
Ghiulai R, Mioc M, Racoviceanu R, Prodea A, Milan A, Coricovac D, Dehelean C, Avram Ș, Zamfir AD, Munteanu CVA, et al. Structural Investigation of Betulinic Acid Plasma Metabolites by Tandem Mass Spectrometry. Molecules. 2022; 27(21):7359. https://doi.org/10.3390/molecules27217359
Chicago/Turabian StyleGhiulai, Roxana, Marius Mioc, Roxana Racoviceanu, Alexandra Prodea, Andreea Milan, Dorina Coricovac, Cristina Dehelean, Ștefana Avram, Alina D. Zamfir, Cristian V. A. Munteanu, and et al. 2022. "Structural Investigation of Betulinic Acid Plasma Metabolites by Tandem Mass Spectrometry" Molecules 27, no. 21: 7359. https://doi.org/10.3390/molecules27217359
APA StyleGhiulai, R., Mioc, M., Racoviceanu, R., Prodea, A., Milan, A., Coricovac, D., Dehelean, C., Avram, Ș., Zamfir, A. D., Munteanu, C. V. A., Ivan, V., & Şoica, C. (2022). Structural Investigation of Betulinic Acid Plasma Metabolites by Tandem Mass Spectrometry. Molecules, 27(21), 7359. https://doi.org/10.3390/molecules27217359