
A Hydrazine–Hydrazone Adamantine Compound Shows Antimycobacterial Activity and Is a Probable Inhibitor of MmpL3
 , ,
, ,
Abstract
1. Introduction
2. Results and Discussion
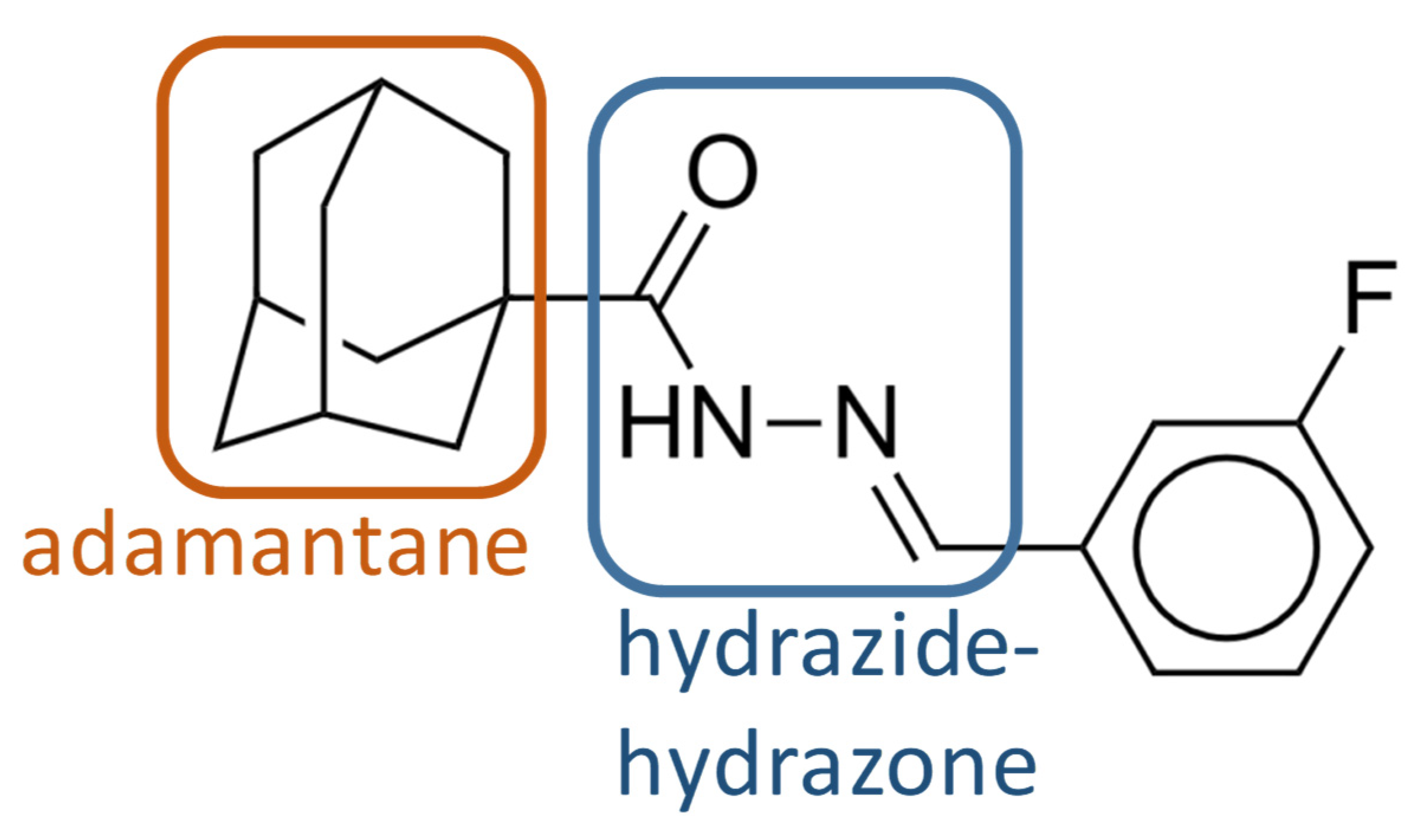
2.1. Screening of the Chemical Library and Identification of Compound 1
2.2. Compound 1 Shows Strong Activity against Clinical Isolates of M. tuberculosis
2.3. Combination of Compound 1 with Linezolid In Vitro
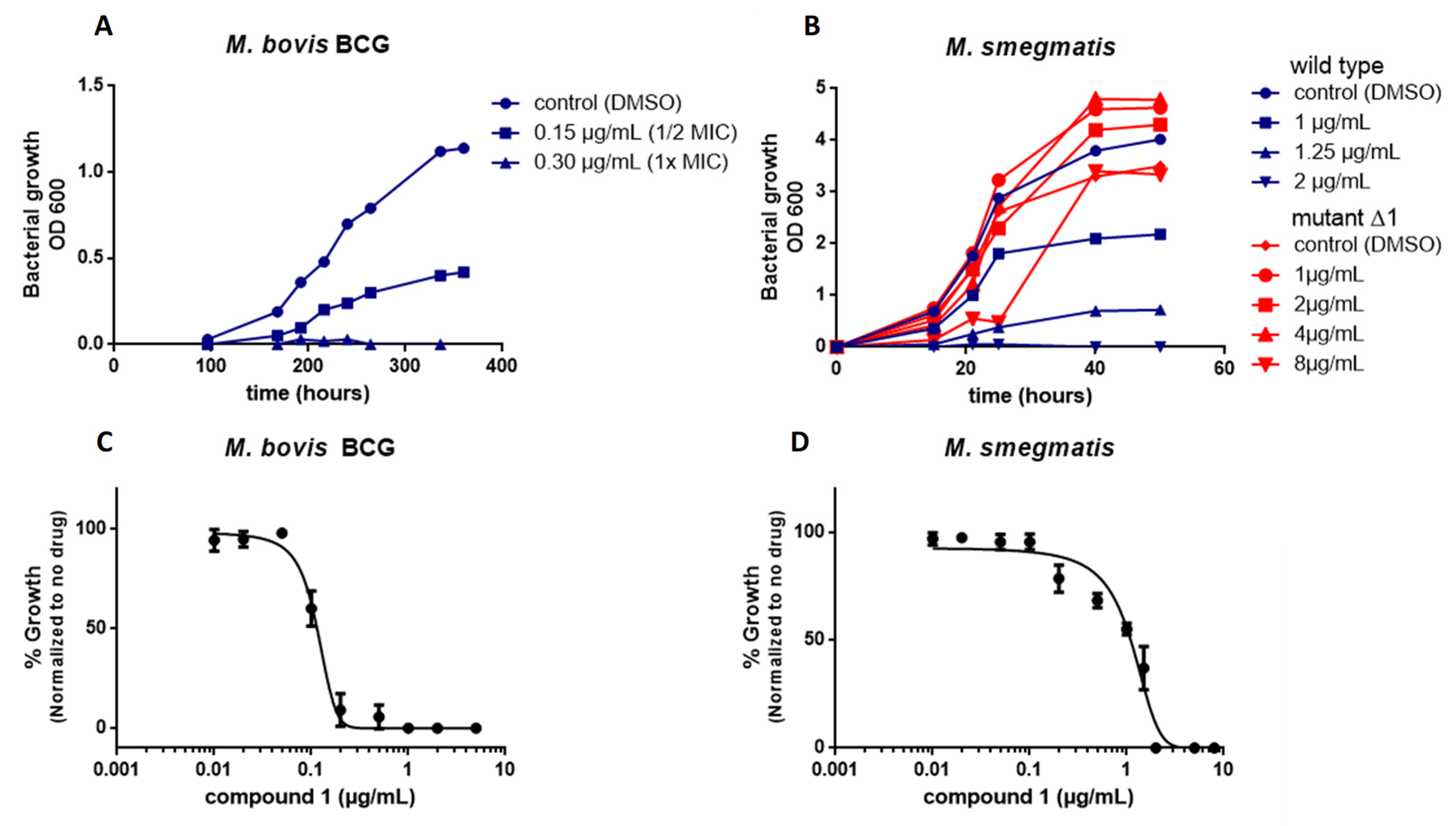
2.4. Mutations in MmpL3 Confer Resistance to Compound 1
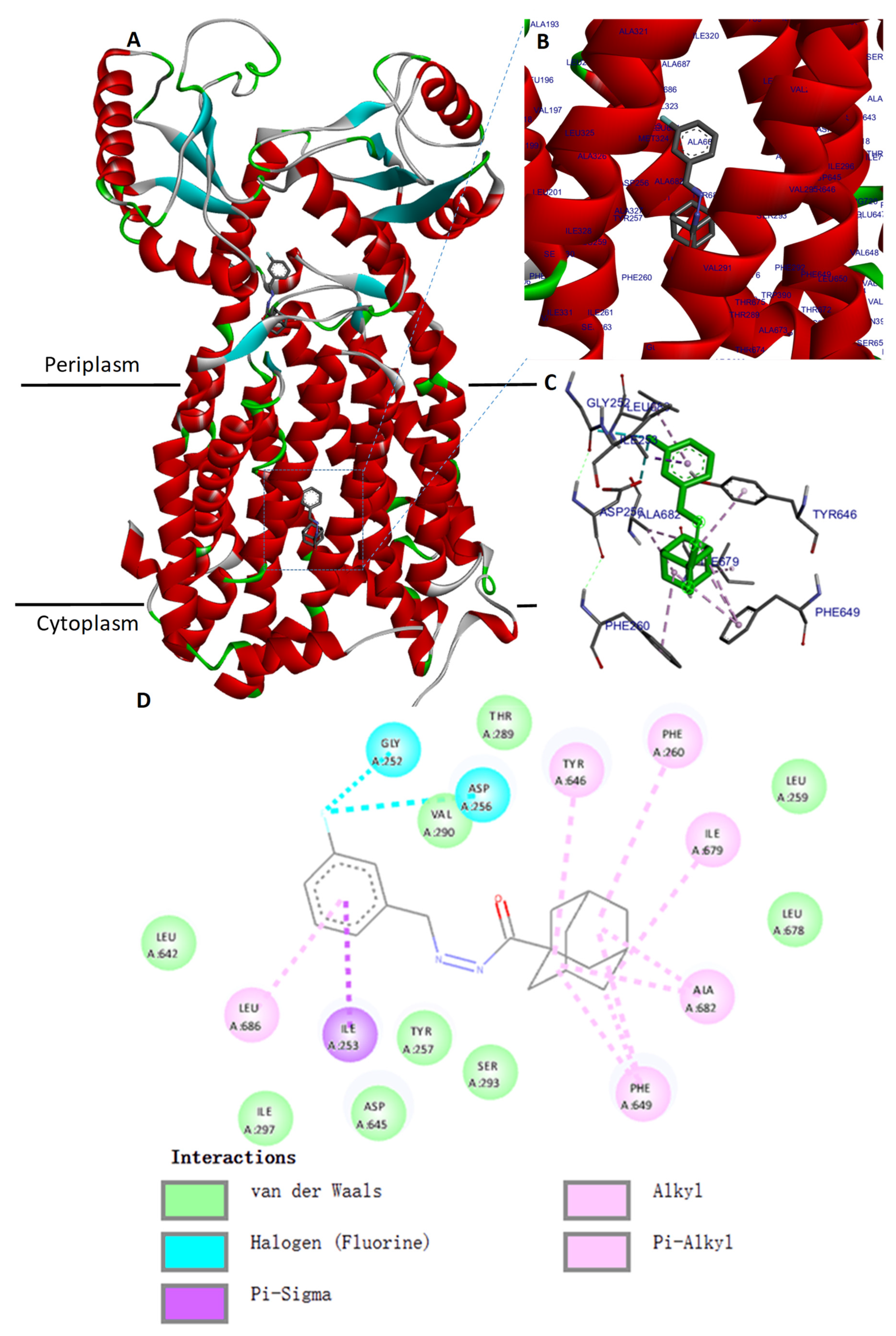
2.5. Molecular Docking of the MmpL3 Complex with Compound 1
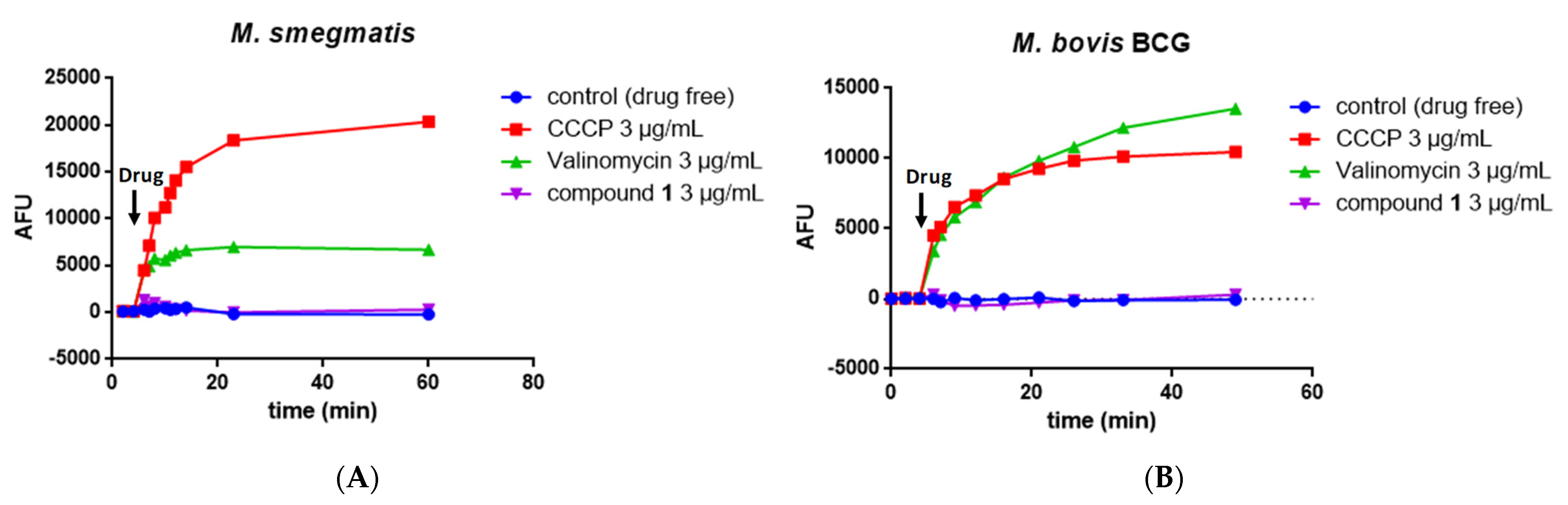
2.6. Measurement of the Mycobacterial Proton Motive Force
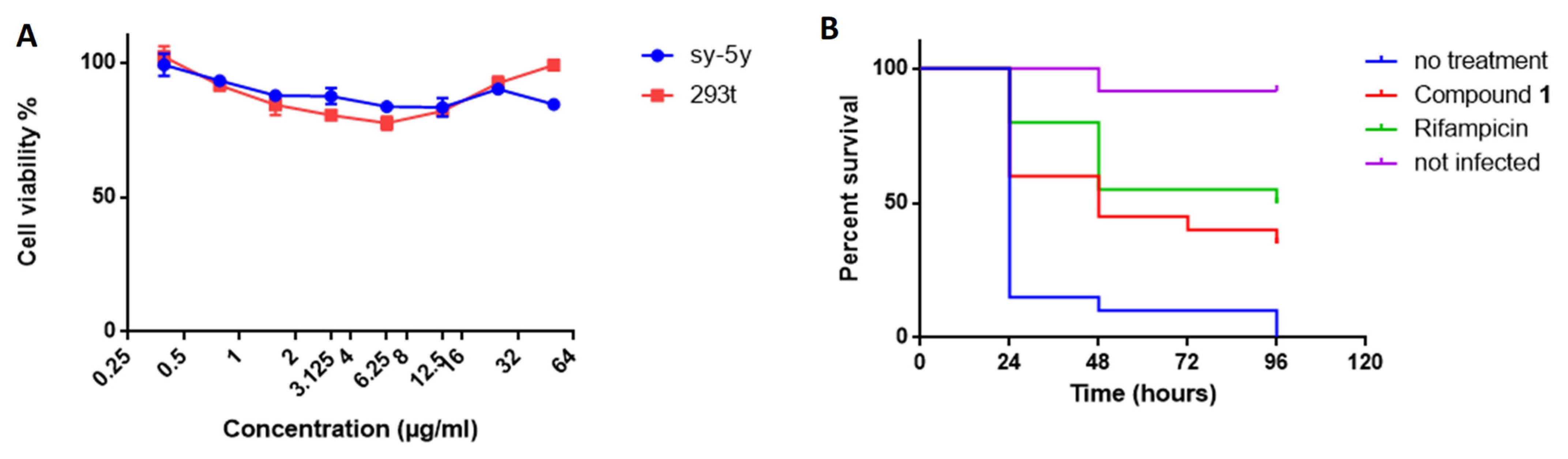
2.7. Compound 1 Shows No Cytotoxicity
2.8. Compound 1 Protects Galleria Mellonella Larvae against M. bovis BCG Infection
3. Materials and Methods
3.1. Drugs and Reagent Preparation
3.2. Strains and Growth Condition
3.3. Screening and MIC and MBC Determination
3.4. Dose–Response Curves
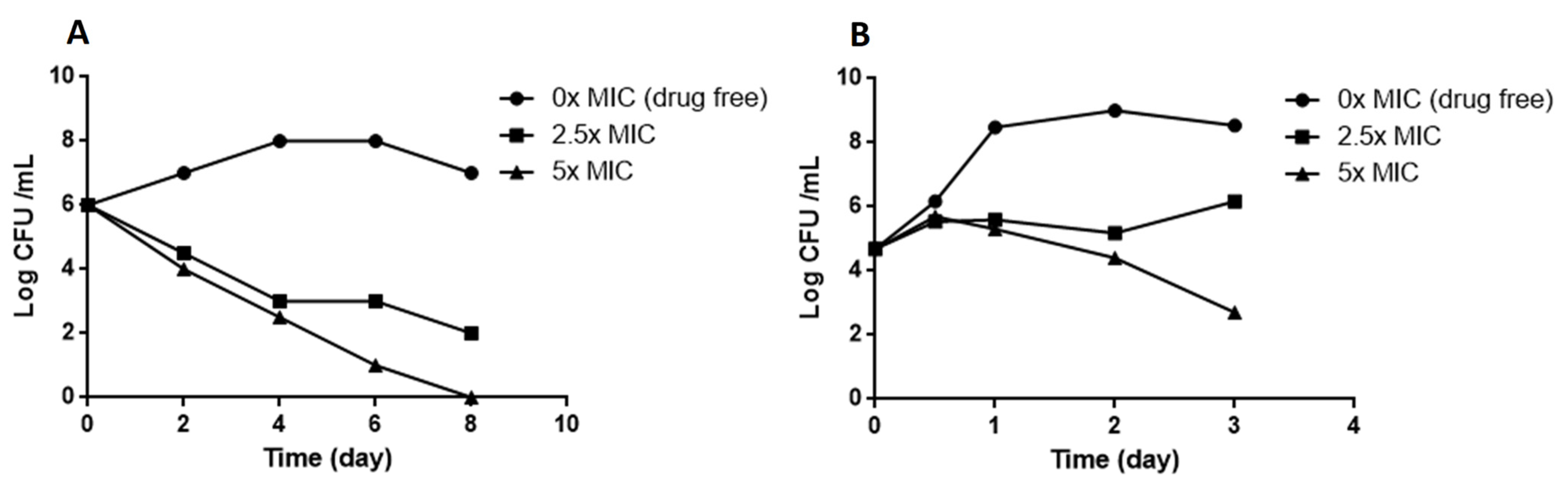
3.5. Kill Kinetics
3.6. Generation and Analysis of Resistant Mutants
3.7. Genome Sequencing and the Identification of Polymorphisms
3.8. Checkerboard Synergy Assay
3.9. Docking Studies
3.10. Measurement of the Mycobacterial Transmembrane Potential ΔΨ
3.11. Measurement of the Mycobacterial Chemical Proton Potential ΔpH
3.12. Adme Prediction
3.13. Cytotoxicity
3.14. Infection of Galleria Mellonella and Drug Treatment
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| CFU | colony-forming unit |
| DMSO | dimethyl sulfoxide |
| EMB | Ethambuthol |
| FBS | fetal bovine serum |
| FICI | fractional inhibitory concentration index |
| MBC | minimum bactericidal concentration |
| MDR | multidrug-resistant |
| MIC | minimum inhibitory concentration |
| MmpL3 | mycobacterial membrane protein large 3 |
| PZA | Pyrazinamide |
| Rif-R | rifampicin resistant |
| TMMs | trehalose monomycolates |
References
- World Health Organization. Global Tuberculosis Report 2021; Global Tuberculosis Programme; World Health Organization: Geneva, Switzerland, 2021; ISBN 9789240037021. [Google Scholar]
- Tandon, R.; Nath, M. Tackling Drug-Resistant Tuberculosis: Current Trends and Approaches. Mini Rev. Med. Chem. 2017, 17, 549–570. [Google Scholar] [CrossRef] [PubMed]
- Lee, A.; Xie, Y.L.; Barry, C.E.; Chen, R.Y. Current and Future Treatments for Tuberculosis. BMJ 2020, 368, m216. [Google Scholar] [CrossRef] [PubMed]
- Mase, S.R.; Chorba, T. Treatment of Drug-Resistant Tuberculosis. Clin. Chest Med. 2019, 40, 775–795. [Google Scholar] [CrossRef] [PubMed]
- World Health Organization. WHO Operational Handbook on Tuberculosis. Module 4: Treatment—Drug-Resistant Tuberculosis Treatment; World Health Organization: Geneva, Switzerland, 2020; ISBN 9789240006997. [Google Scholar]
- Andries, K.; Verhasselt, P.; Guillemont, J.; Gohlmann, H.W.; Neefs, J.M.; Winkler, H.; Van Gestel, J.; Timmerman, P.; Zhu, M.; Lee, E.; et al. A Diarylquinoline Drug Active on the ATP Synthase of Mycobacterium Tuberculosis. Science 2005, 307, 223–227. [Google Scholar] [CrossRef] [PubMed]
- Wilson, D.N.; Schluenzen, F.; Harms, J.M.; Starosta, A.L.; Connell, S.R.; Fucini, P. The Oxazolidinone Antibiotics Perturb the Ribosomal Peptidyl-Transferase Center and Effect TRNA Positioning. Proc. Natl. Acad. Sci. USA 2008, 105, 13339–13344. [Google Scholar] [CrossRef]
- Lee, M.; Lee, J.; Carroll, M.W.; Choi, H.; Min, S.; Song, T.; Via, L.E.; Goldfeder, L.C.; Kang, E.; Jin, B.; et al. Linezolid for Treatment of Chronic Extensively Drug-Resistant Tuberculosis. N. Engl. J. Med. 2012, 367, 1508–1518. [Google Scholar] [CrossRef]
- Tiberi, S.; Vjecha, M.J.; Zumla, A.; Galvin, J.; Migliori, G.B.; Zumla, A. Accelerating Development of New Shorter TB Treatment Regimens in Anticipation of a Resurgence of Multi-Drug Resistant TB Due to the COVID-19 Pandemic. Int. J. Infect. Dis. IJID Off. Publ. Int. Soc. Infect. Dis. 2021, 113, S96–S99. [Google Scholar] [CrossRef]
- Tyers, M.; Wright, G.D. Drug Combinations: A Strategy to Extend the Life of Antibiotics in the 21st Century. Nat. Rev. Microbiol. 2019, 17, 141–155. [Google Scholar] [CrossRef]
- Huang, W.; Briffotaux, J.; Wang, X.; Liu, L.; Hao, P.; Cimino, M.; Buchieri, M.V.; Namouchi, A.; Ainsa, J.-A.; Gicquel, B. Ionophore A23187 Shows Anti-Tuberculosis Activity and Synergy with Tebipenem. Tuberculosis 2017, 107, 111–118. [Google Scholar] [CrossRef]
- Briffotaux, J.; Liu, S.; Gicquel, B. Genome-Wide Transcriptional Responses of Mycobacterium to Antibiotics. Front. Microbiol. 2019, 10, 249. [Google Scholar] [CrossRef]
- Ioerger, T.R.; O’Malley, T.; Liao, R.; Guinn, K.M.; Hickey, M.J.; Mohaideen, N.; Murphy, K.C.; Boshoff, H.I.M.; Mizrahi, V.; Rubin, E.J.; et al. Identification of New Drug Targets and Resistance Mechanisms in Mycobacterium Tuberculosis. PLoS ONE 2013, 8, e75245. [Google Scholar] [CrossRef] [PubMed]
- Varela, C.; Rittmann, D.; Singh, A.; Krumbach, K.; Bhatt, K.; Eggeling, L.; Besra, G.S.; Bhatt, A. MmpL Genes Are Associated with Mycolic Acid Metabolism in Mycobacteria and Corynebacteria. Chem. Biol. 2012, 19, 498–506. [Google Scholar] [CrossRef] [PubMed]
- Xu, Z.; Meshcheryakov, V.A.; Poce, G.; Chng, S.-S. MmpL3 Is the Flippase for Mycolic Acids in Mycobacteria. Proc. Natl. Acad. Sci. USA 2017, 114, 7993–7998. [Google Scholar] [CrossRef]
- Zhou, Y.; Gammeltoft, K.A.; Galli, A.; Offersgaard, A.; Fahnøe, U.; Ramirez, S.; Bukh, J.; Gottwein, J.M. Efficacy of Ion-Channel Inhibitors Amantadine, Memantine and Rimantadine for the Treatment of SARS-CoV-2 In Vitro. Viruses 2021, 13, 2082. [Google Scholar] [CrossRef]
- Grzegorzewicz, A.E.; Pham, H.; Gundi, V.A.K.B.; Scherman, M.S.; North, E.J.; Hess, T.; Jones, V.; Gruppo, V.; Born, S.E.M.; Korduláková, J.; et al. Inhibition of Mycolic Acid Transport across the Mycobacterium Tuberculosis Plasma Membrane. Nat. Chem. Biol. 2012, 8, 334–341. [Google Scholar] [CrossRef] [PubMed]
- Protopopova, M.; Hanrahan, C.; Nikonenko, B.; Samala, R.; Chen, P.; Gearhart, J.; Einck, L.; Nacy, C.A. Identification of a New Antitubercular Drug Candidate, SQ109, from a Combinatorial Library of 1,2-Ethylenediamines. J. Antimicrob. Chemother. 2005, 56, 968–974. [Google Scholar] [CrossRef] [PubMed]
- Heinrich, N.; Dawson, R.; du Bois, J.; Narunsky, K.; Horwith, G.; Phipps, A.J.; Nacy, C.A.; Aarnoutse, R.E.; Boeree, M.J.; Gillespie, S.H.; et al. Early Phase Evaluation of SQ109 Alone and in Combination with Rifampicin in Pulmonary TB Patients. J. Antimicrob. Chemother. 2015, 70, 1558–1566. [Google Scholar] [CrossRef] [PubMed]
- Boeree, M.J.; Heinrich, N.; Aarnoutse, R.; Diacon, A.H.; Dawson, R.; Rehal, S.; Kibiki, G.S.; Churchyard, G.; Sanne, I.; Ntinginya, N.E.; et al. High-Dose Rifampicin, Moxifloxacin, and SQ109 for Treating Tuberculosis: A Multi-Arm, Multi-Stage Randomised Controlled Trial. Lancet Infect. Dis. 2017, 17, 39–49. [Google Scholar] [CrossRef]
- Popiołek, Ł. Hydrazide-Hydrazones as Potential Antimicrobial Agents: Overview of the Literature since 2010. Med. Chem. Res. 2017, 26, 287–301. [Google Scholar] [CrossRef]
- Popiołek, Ł. Updated Information on Antimicrobial Activity of Hydrazide-Hydrazones. Int. J. Mol. Sci. 2021, 22, 9389. [Google Scholar] [CrossRef]
- Rollas, S.; Gulerman, N.; Erdeniz, H. Synthesis and Antimicrobial Activity of Some New Hydrazones of 4-Fluorobenzoic Acid Hydrazide and 3-Acetyl-2,5-Disubstituted-1,3,4-Oxadiazolines. Farmaco 2002, 57, 171–174. [Google Scholar] [CrossRef]
- Bedia, K.-K.; Elçin, O.; Seda, U.; Fatma, K.; Nathaly, S.; Sevim, R.; Dimoglo, A. Synthesis and Characterization of Novel Hydrazide-Hydrazones and the Study of Their Structure-Antituberculosis Activity. Eur. J. Med. Chem. 2006, 41, 1253–1261. [Google Scholar] [CrossRef] [PubMed]
- Abdelrahman, M.A.; Salama, I.; Gomaa, M.S.; Elaasser, M.M.; Abdel-Aziz, M.M.; Soliman, D.H. Design, Synthesis and 2D QSAR Study of Novel Pyridine and Quinolone Hydrazone Derivatives as Potential Antimicrobial and Antitubercular Agents. Eur. J. Med. Chem. 2017, 138, 698–714. [Google Scholar] [CrossRef] [PubMed]
- Krátký, M.; Bősze, S.; Baranyai, Z.; Stolaříková, J.; Vinšová, J. Synthesis and Biological Evolution of Hydrazones Derived from 4-(Trifluoromethyl)Benzohydrazide. Bioorg. Med. Chem. Lett. 2017, 27, 5185–5189. [Google Scholar] [CrossRef]
- Kumar, P.; Narasimhan, B. Hydrazides/Hydrazones as Antimicrobial and Anticancer Agents in the New Millennium. Mini Rev. Med. Chem. 2013, 13, 971–987. [Google Scholar] [CrossRef]
- Zheng, H.; Williams, J.T.; Coulson, G.B.; Haiderer, E.R.; Abramovitch, R.B. HC2091 Kills Mycobacterium Tuberculosis by Targeting the MmpL3 Mycolic Acid Transporter. Antimicrob. Agents Chemother. 2018, 62, e02459-17. [Google Scholar] [CrossRef]
- Williams, J.T.; Haiderer, E.R.; Coulson, G.B.; Conner, K.N.; Ellsworth, E.; Chen, C.; Alvarez-Cabrera, N.; Li, W.; Jackson, M.; Dick, T.; et al. Identification of New MmpL3 Inhibitors by Untargeted and Targeted Mutant Screens Defines MmpL3 Domains with Differential Resistance. Antimicrob. Agents Chemother. 2019, 63, e00547-19. [Google Scholar] [CrossRef]
- Zampieri, M.; Enke, T.; Chubukov, V.; Ricci, V.; Piddock, L.; Sauer, U. Metabolic Constraints on the Evolution of Antibiotic Resistance. Mol. Syst. Biol. 2017, 13, 917. [Google Scholar] [CrossRef]
- Foss, M.H.; Pou, S.; Davidson, P.M.; Dunaj, J.L.; Winter, R.W.; Pou, S.; Licon, M.H.; Doh, J.K.; Li, Y.; Kelly, J.X.; et al. Diphenylether-Modified 1,2-Diamines with Improved Drug Properties for Development against Mycobacterium Tuberculosis. ACS Infect. Dis. 2016, 2, 500–508. [Google Scholar] [CrossRef]
- Li, W.; Obregón-Henao, A.; Wallach, J.B.; North, E.J.; Lee, R.E.; Gonzalez-Juarrero, M.; Schnappinger, D.; Jackson, M. Therapeutic Potential of the Mycobacterium Tuberculosis Mycolic Acid Transporter, MmpL3. Antimicrob. Agents Chemother. 2016, 60, 5198–5207. [Google Scholar] [CrossRef]
- Viljoen, A.; Dubois, V.; Girard-Misguich, F.; Blaise, M.; Herrmann, J.-L.; Kremer, L. The Diverse Family of MmpL Transporters in Mycobacteria: From Regulation to Antimicrobial Developments. Mol. Microbiol. 2017, 104, 889–904. [Google Scholar] [CrossRef] [PubMed]
- Bolla, J.R. Targeting MmpL3 for Anti-Tuberculosis Drug Development. Biochem. Soc. Trans. 2020, 48, 1463–1472. [Google Scholar] [CrossRef] [PubMed]
- Shao, M.; McNeil, M.; Cook, G.M.; Lu, X. MmpL3 Inhibitors as Antituberculosis Drugs. Eur. J. Med. Chem. 2020, 200, 112390. [Google Scholar] [CrossRef] [PubMed]
- Tahlan, K.; Wilson, R.; Kastrinsky, D.B.; Arora, K.; Nair, V.; Fischer, E.; Barnes, S.W.; Walker, J.R.; Alland, D.; Barry, C.E., III; et al. SQ109 Targets MmpL3, a Membrane Transporter of Trehalose Monomycolate Involved in Mycolic Acid Donation to the Cell Wall Core of Mycobacterium Tuberculosis. Antimicrob. Agents Chemother. 2012, 56, 1797–1809. [Google Scholar] [CrossRef]
- La Rosa, V.; Poce, G.; Canseco, J.O.; Buroni, S.; Pasca, M.R.; Biava, M.; Raju, R.M.; Porretta, G.C.; Alfonso, S.; Battilocchio, C.; et al. MmpL3 Is the Cellular Target of the Antitubercular Pyrrole Derivative BM212. Antimicrob. Agents Chemother. 2012, 56, 324–331. [Google Scholar] [CrossRef]
- Lun, S.; Guo, H.; Onajole, O.K.; Pieroni, M.; Gunosewoyo, H.; Chen, G.; Tipparaju, S.K.; Ammerman, N.C.; Kozikowski, A.P.; Bishai, W.R. Indoleamides Are Active against Drug-Resistant Mycobacterium Tuberculosis. Nat. Commun. 2013, 4, 2907. [Google Scholar] [CrossRef]
- Chawla, M.; Parikh, P.; Saxena, A.; Munshi, M.; Mehta, M.; Mai, D.; Srivastava, A.K.; Narasimhulu, K.V.; Redding, K.E.; Vashi, N.; et al. Mycobacterium Tuberculosis WhiB4 Regulates Oxidative Stress Response to Modulate Survival and Dissemination in Vivo. Mol. Microbiol. 2012, 85, 1148–1165. [Google Scholar] [CrossRef]
- Remuiñán, M.J.; Pérez-Herrán, E.; Rullás, J.; Alemparte, C.; Martínez-Hoyos, M.; Dow, D.J.; Afari, J.; Mehta, N.; Esquivias, J.; Jiménez, E.; et al. Tetrahydropyrazolo[1,5-a]Pyrimidine-3-Carboxamide and N-Benzyl-6’,7’-Dihydrospiro[Piperidine-4,4’-Thieno[3,2-c]Pyran] Analogues with Bactericidal Efficacy against Mycobacterium Tuberculosis Targeting MmpL3. PLoS ONE 2013, 8, e60933. [Google Scholar] [CrossRef]
- Dupont, C.; Chen, Y.; Xu, Z.; Roquet-Banères, F.; Blaise, M.; Witt, A.-K.; Dubar, F.; Biot, C.; Guérardel, Y.; Maurer, F.P.; et al. A Piperidinol-Containing Molecule Is Active against Mycobacterium Tuberculosis by Inhibiting the Mycolic Acid Flippase Activity of MmpL3. J. Biol. Chem. 2019, 294, 17512–17523. [Google Scholar] [CrossRef]
- Grover, S.; Engelhart, C.A.; Pérez-Herrán, E.; Li, W.; Abrahams, K.A.; Papavinasasundaram, K.; Bean, J.M.; Sassetti, C.M.; Mendoza-Losana, A.; Besra, G.S.; et al. Two-Way Regulation of MmpL3 Expression Identifies and Validates Inhibitors of MmpL3 Function in Mycobacterium Tuberculosis. ACS Infect. Dis. 2021, 7, 141–152. [Google Scholar] [CrossRef]
- Li, W.; Stevens, C.M.; Pandya, A.N.; Darzynkiewicz, Z.; Bhattarai, P.; Tong, W.; Gonzalez-Juarrero, M.; North, E.J.; Zgurskaya, H.I.; Jackson, M. Direct Inhibition of MmpL3 by Novel Antitubercular Compounds. ACS Infect. Dis. 2019, 5, 1001–1012. [Google Scholar] [CrossRef] [PubMed]
- Zhang, B.; Li, J.; Yang, X.; Wu, L.; Zhang, J.; Yang, Y.; Zhao, Y.; Zhang, L.; Yang, X.; Yang, X.; et al. Crystal Structures of Membrane Transporter MmpL3, an Anti-TB Drug Target. Cell 2019, 176, 636–648.e13. [Google Scholar] [CrossRef] [PubMed]
- Bernut, A.; Viljoen, A.; Dupont, C.; Sapriel, G.; Blaise, M.; Bouchier, C.; Brosch, R.; de Chastellier, C.; Herrmann, J.-L.; Kremer, L. Insights into the Smooth-to-Rough Transitioning in Mycobacterium Bolletii Unravels a Functional Tyr Residue Conserved in All Mycobacterial MmpL Family Members. Mol. Microbiol. 2016, 99, 866–883. [Google Scholar] [CrossRef] [PubMed]
- Su, C.-C.; Klenotic, P.A.; Bolla, J.R.; Purdy, G.E.; Robinson, C.V.; Yu, E.W. MmpL3 Is a Lipid Transporter That Binds Trehalose Monomycolate and Phosphatidylethanolamine. Proc. Natl. Acad. Sci. USA 2019, 116, 11241–11246. [Google Scholar] [CrossRef] [PubMed]
- Adams, O.; Deme, J.C.; Parker, J.L.; Fowler, P.W.; Lea, S.M.; Newstead, S. Cryo-EM Structure and Resistance Landscape of M. Tuberculosis MmpL3: An Emergent Therapeutic Target. Structure 2021, 29, 1182–1191.e4. [Google Scholar] [CrossRef]
- Umare, M.D.; Khedekar, P.B.; Chikhale, R.V. Mycobacterial Membrane Protein Large 3 (MmpL3) Inhibitors: A Promising Approach to Combat Tuberculosis. ChemMedChem 2021, 16, 3136–3148. [Google Scholar] [CrossRef]
- Moorey, A.R.; Cabanillas, A.; Batt, S.M.; Ghidelli-Disse, S.; Urones, B.; Sanz, O.; Lelievre, J.; Bantscheff, M.; Cox, L.R.; Besra, G.S. The Multi-Target Aspect of an MmpL3 Inhibitor: The BM212 Series of Compounds Bind EthR2, a Transcriptional Regulator of Ethionamide Activation. Cell Surf. 2021, 7, 100068. [Google Scholar] [CrossRef]
- Su, C.-C.; Klenotic, P.A.; Cui, M.; Lyu, M.; Morgan, C.E.; Yu, E.W. Structures of the Mycobacterial Membrane Protein MmpL3 Reveal Its Mechanism of Lipid Transport. PLoS Biol. 2021, 19, e3001370. [Google Scholar] [CrossRef]
- Li, W.; Upadhyay, A.; Fontes, F.L.; North, E.J.; Wang, Y.; Crans, D.C.; Grzegorzewicz, A.E.; Jones, V.; Franzblau, S.G.; Lee, R.E.; et al. Novel Insights into the Mechanism of Inhibition of MmpL3, a Target of Multiple Pharmacophores in Mycobacterium Tuberculosis. Antimicrob. Agents Chemother. 2014, 58, 6413–6423. [Google Scholar] [CrossRef]
- Daina, A.; Michielin, O.; Zoete, V. SwissADME: A Free Web Tool to Evaluate Pharmacokinetics, Drug-Likeness and Medicinal Chemistry Friendliness of Small Molecules. Sci. Rep. 2017, 7, 42717. [Google Scholar] [CrossRef]
- Asai, M.; Li, Y.; Khara, J.S.; Robertson, B.D.; Langford, P.R.; Newton, S.M. Galleria Mellonella: An Infection Model for Screening Compounds Against the Mycobacterium Tuberculosis Complex. Front. Microbiol. 2019, 10, 2630. [Google Scholar] [CrossRef] [PubMed]
- Entwistle, F.M.; Coote, P.J. Evaluation of Greater Wax Moth Larvae, Galleria Mellonella, as a Novel in Vivo Model for Non-Tuberculosis Mycobacteria Infections and Antibiotic Treatments. J. Med. Microbiol. 2018, 67, 585–597. [Google Scholar] [CrossRef] [PubMed]
- Mourik, B.C.; de Knegt, G.J.; Verbon, A.; Mouton, J.W.; Bax, H.I.; de Steenwinkel, J.E.M. Assessment of Bactericidal Drug Activity and Treatment Outcome in a Mouse Tuberculosis Model Using a Clinical Beijing Strain. Antimicrob. Agents Chemother. 2017, 61, e00696-17. [Google Scholar] [CrossRef] [PubMed]
- European Committee for Antimicrobial Susceptibility Testing (EUCAST) of the European Society of Clinical Microbiology and Infectious Dieases (ESCMID). EUCAST Definitive Document E. Def 1.2, May 2000: Terminology Relating to Methods for the Determination of Susceptibility of Bacteria to Antimicrobial Agents. Clin. Microbiol. Infect. Off. Publ. Eur. Soc. Clin. Microbiol. Infect. Dis. 2000, 6, 503–508. [Google Scholar] [CrossRef]
- Lechartier, B.; Hartkoorn, R.C.; Cole, S.T. In Vitro Combination Studies of Benzothiazinone Lead Compound BTZ043 against Mycobacterium Tuberculosis. Antimicrob. Agents Chemother. 2012, 56, 5790–5793. [Google Scholar] [CrossRef]
- Shetty, A.; Xu, Z.; Lakshmanan, U.; Hill, J.; Choong, M.L.; Chng, S.-S.; Yamada, Y.; Poulsen, A.; Dick, T.; Gengenbacher, M. Novel Acetamide Indirectly Targets Mycobacterial Transporter MmpL3 by Proton Motive Force Disruption. Front. Microbiol. 2018, 9, 2960. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Bacterial Strains | MIC | MBC |
|---|---|---|
| M. tuberculosis H37Rv | 0.2 | n.d. |
| M. bovis BCG | 0.3 | 0.6 |
| M. smegmatis | 1.5 | 3 |
| M. marinum | 12.5 | n.d. |
| M. abscessus | 12.5 | n.d. |
| M. avium | >25 | n.d. |
| S. aureus | >25 | n.d. |
| E. coli | >25 | n.d. |
| M. tuberculosis Clinical Strain | Strain Type | MIC |
|---|---|---|
| S17 | Sensitive | 0.2–0.39 |
| S23 | Sensitive | 0.2–0.39 |
| S31 | Sensitive | 0.2–0.39 |
| M9 | Rif-R | 0.78 |
| M20 | Rif-R | 1.56 |
| M34 | Rif-R | 0.39–0.78 |
| M7 | MDR | 0.2–0.39 |
| M. bovis BCG | M. smegmatis | M. tuberculosis | |
|---|---|---|---|
| Linezolid | 0.50 | 0.28 | 0.68 |
| REF | ALT | Amino Acid Change | Impact | Gene Name | Description | |
|---|---|---|---|---|---|---|
| MutantΔ1 | ||||||
| snp | TTC | CTC | Phe649Leu | missense | MSMEG_0250 | MmpL3 |
| snp | CAC | CAT | His80His | synonymous | IS1 family transposase IS1S | |
| MutantΔ2 | ||||||
| snp | TTC | CTC | Phe649Leu | missense | MSMEG_0250 | MmpL3 |
| snp | AGA | AGG | Arg54Arg | synonymous | IS1 family transposase IS1S | |
| MutantΔ3 | ||||||
| snp | TTC | CTC | Phe649Leu | missense | MSMEG_0250 | MmpL3 |
| Drug | pH Outside | pH Inside | ΔpH |
|---|---|---|---|
| DMSO | 6.80 | 7.20 (±0.01) | 0.40 (±0.01) |
| Nigericin 5 µg/mL | 6.80 | 6.76 (±0.01) | −0.04 (±0.01) |
| CCCP 1 µg/mL | 6.80 | 6.91 (±0.01) | 0.11 (±0.01) |
| Compound 1 10 µg/mL | 6.80 | 7.13 (±0.02) | 0.33 (±0.02) |
| Compound 1 1 µg/mL | 6.80 | 7.19 (±0.01) | 0.39 (±0.01) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Briffotaux, J.; Xu, Y.; Huang, W.; Hui, Z.; Wang, X.; Gicquel, B.; Liu, S. A Hydrazine–Hydrazone Adamantine Compound Shows Antimycobacterial Activity and Is a Probable Inhibitor of MmpL3. Molecules 2022, 27, 7130. https://doi.org/10.3390/molecules27207130
Briffotaux J, Xu Y, Huang W, Hui Z, Wang X, Gicquel B, Liu S. A Hydrazine–Hydrazone Adamantine Compound Shows Antimycobacterial Activity and Is a Probable Inhibitor of MmpL3. Molecules. 2022; 27(20):7130. https://doi.org/10.3390/molecules27207130
Chicago/Turabian StyleBriffotaux, Julien, Yanji Xu, Wei Huang, Zhen Hui, Xiao Wang, Brigitte Gicquel, and Shengyuan Liu. 2022. "A Hydrazine–Hydrazone Adamantine Compound Shows Antimycobacterial Activity and Is a Probable Inhibitor of MmpL3" Molecules 27, no. 20: 7130. https://doi.org/10.3390/molecules27207130
APA StyleBriffotaux, J., Xu, Y., Huang, W., Hui, Z., Wang, X., Gicquel, B., & Liu, S. (2022). A Hydrazine–Hydrazone Adamantine Compound Shows Antimycobacterial Activity and Is a Probable Inhibitor of MmpL3. Molecules, 27(20), 7130. https://doi.org/10.3390/molecules27207130