
Acetylcholinesterase Inhibitory Activities of Essential Oils from Vietnamese Traditional Medicinal Plants

 ,
,  ,
,  , , , ,
, , , , 
Abstract
1. Introduction
2. Results and Discussion
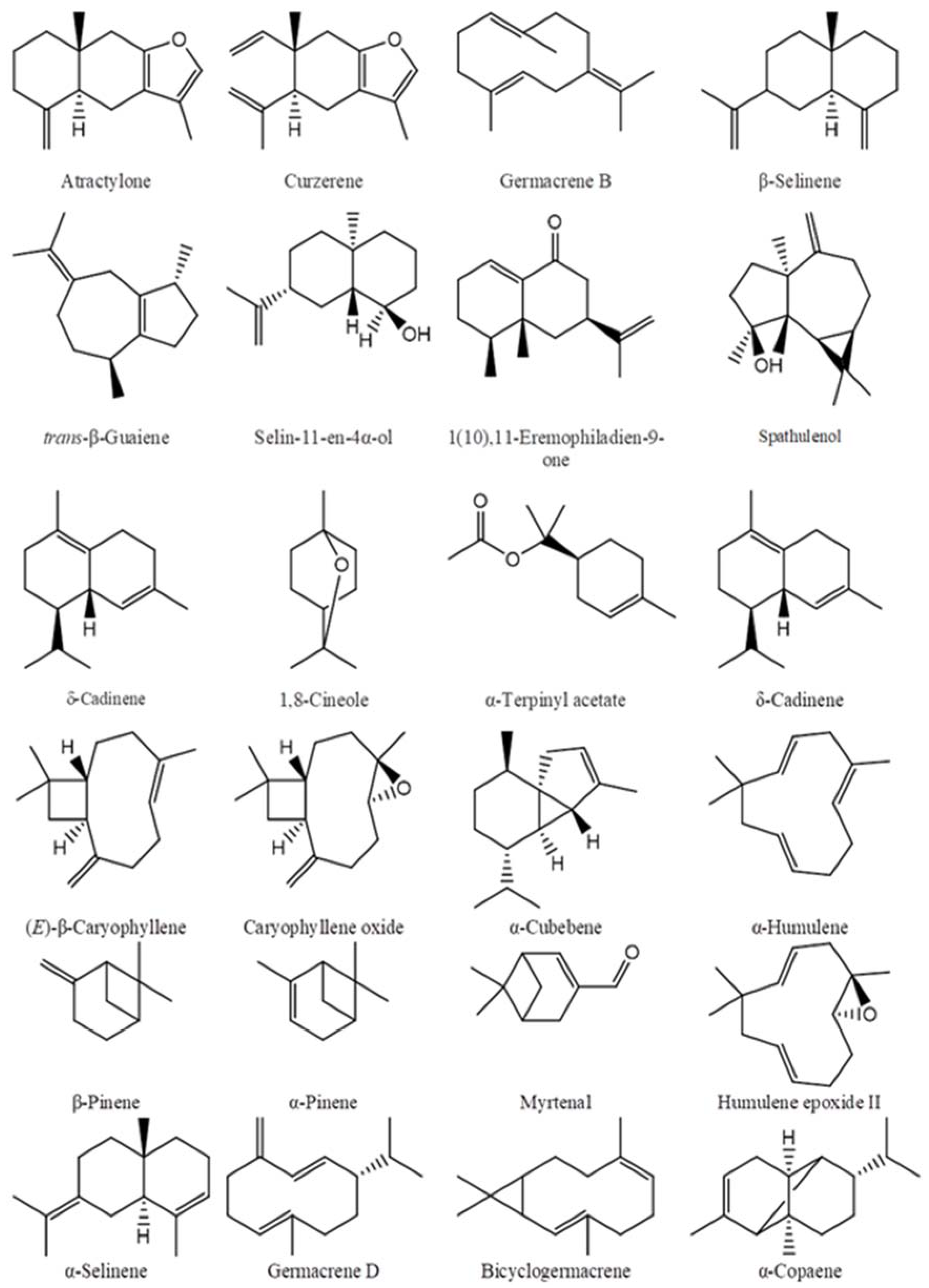
2.1. Chemical Compositions of the Essential Oils
2.2. Acetylcholinesterase Inhibitory Activity
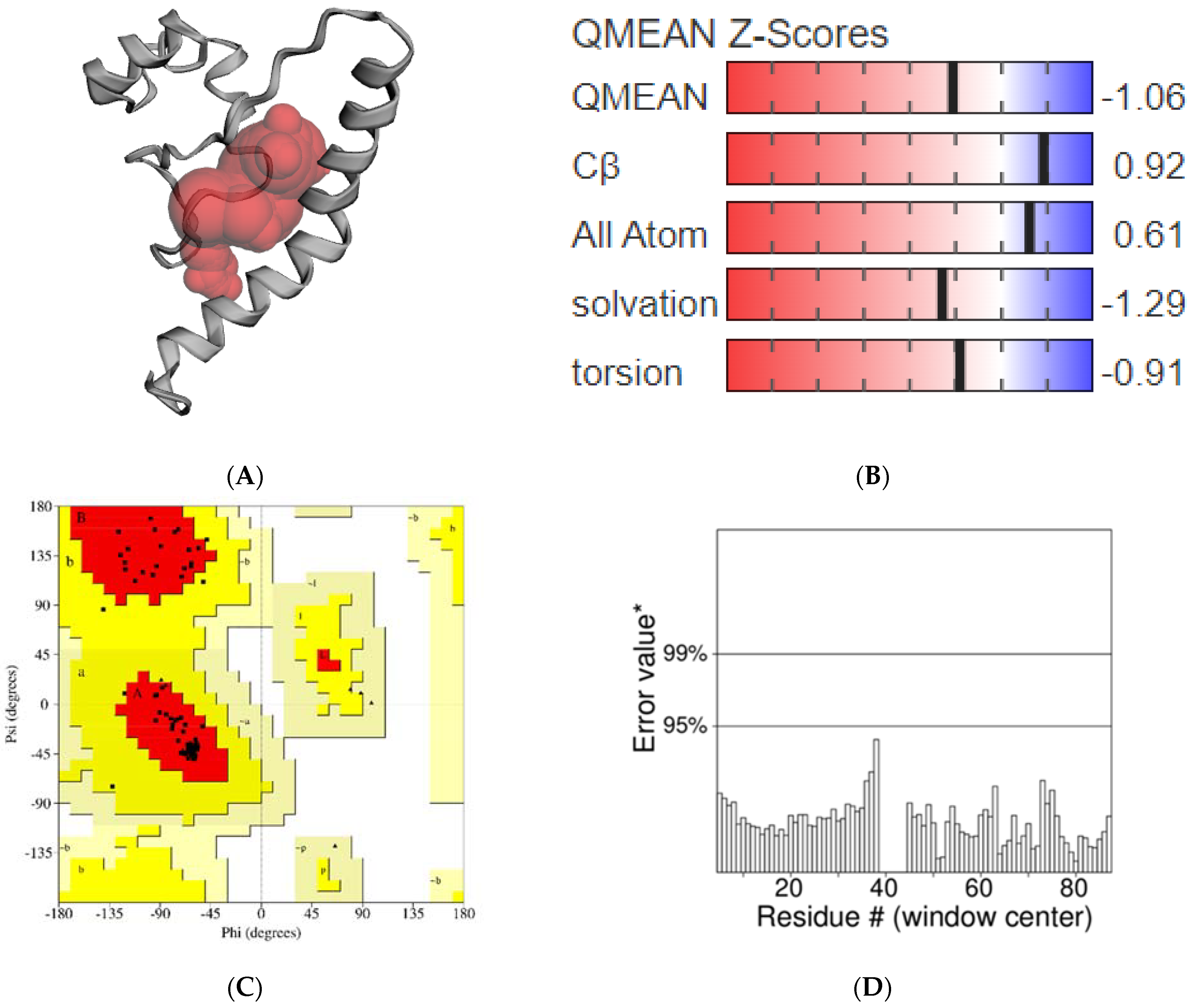
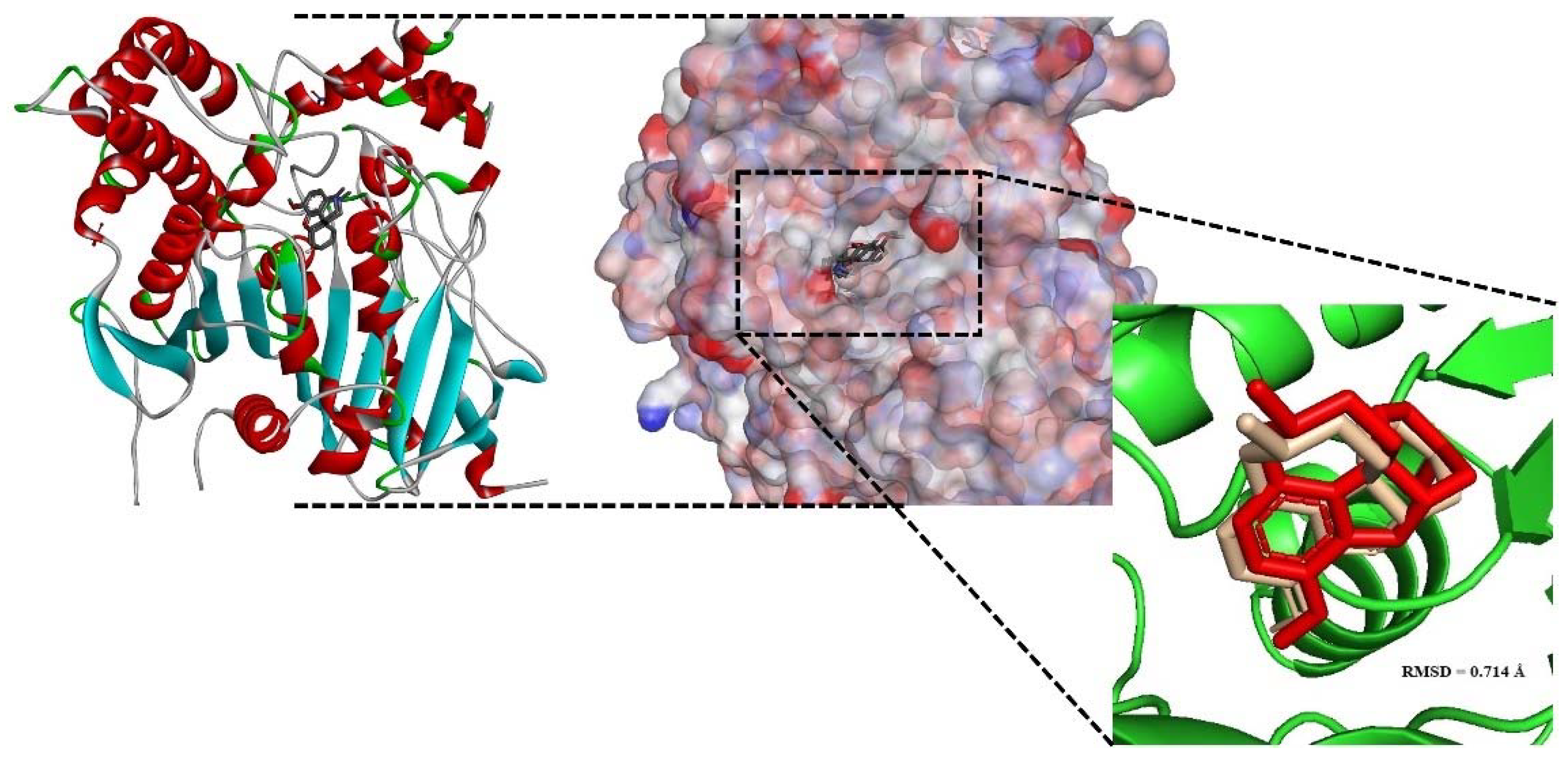
2.3. Homology Modeling Study of AChE1 Enzyme
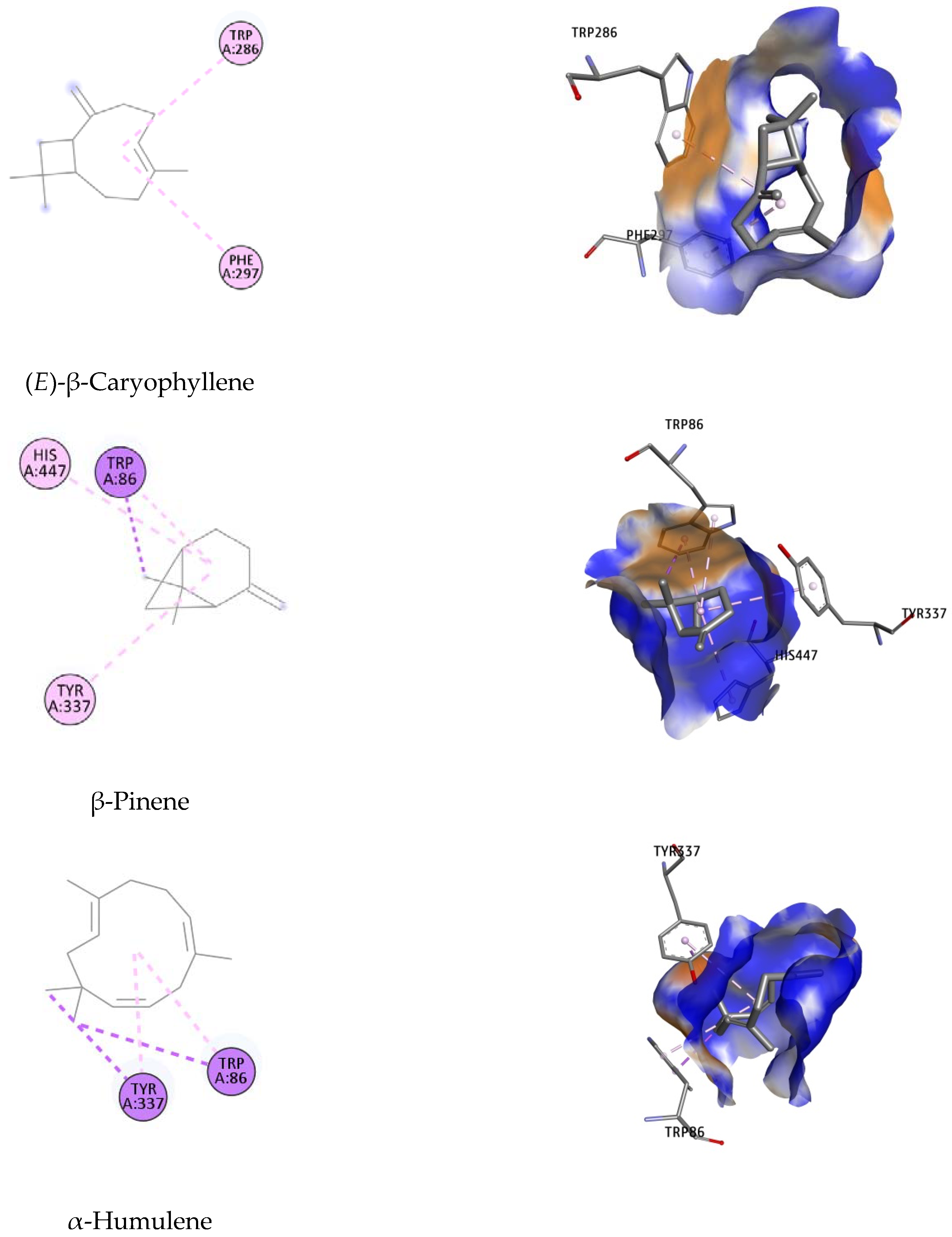
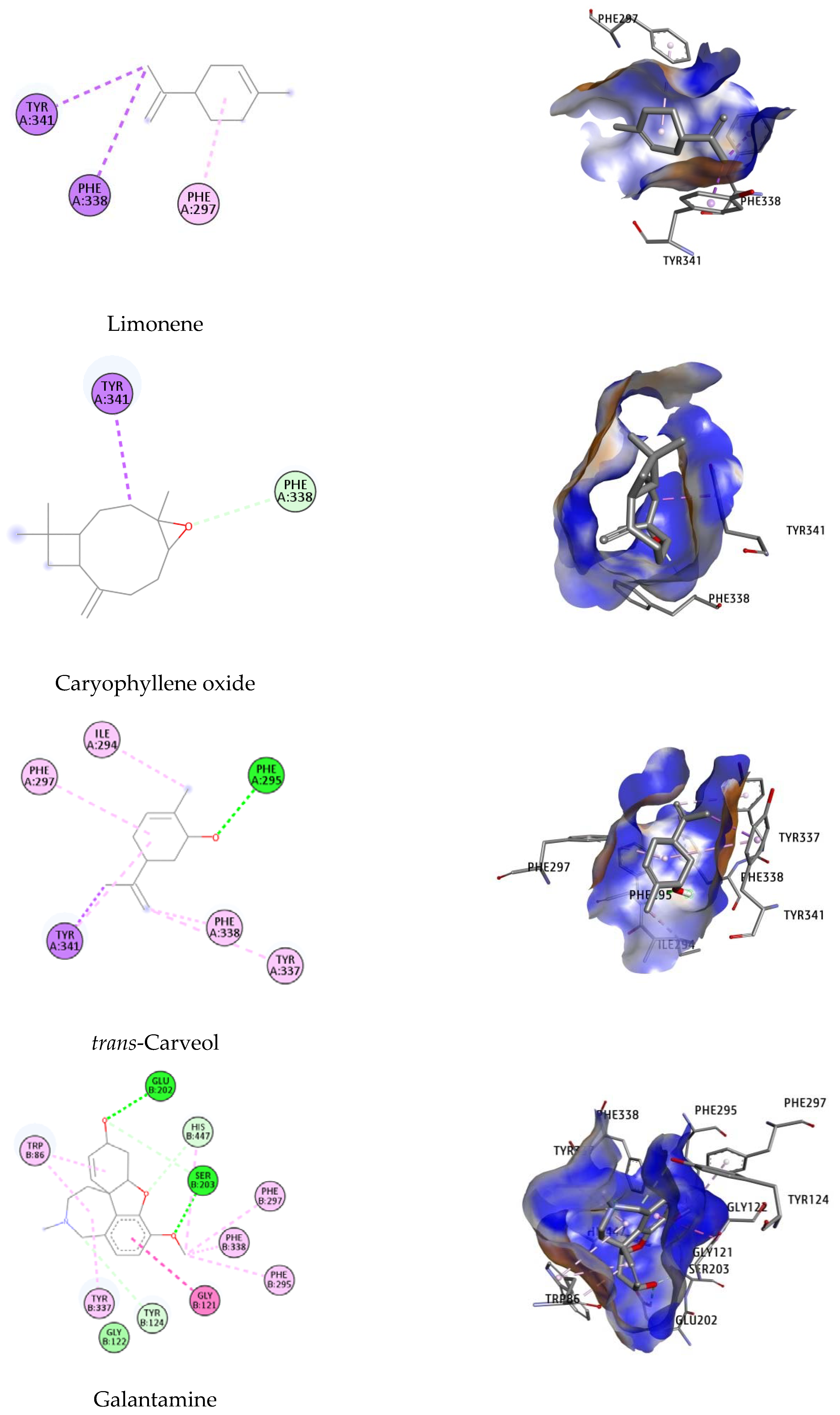
2.4. Molecular Docking Studies
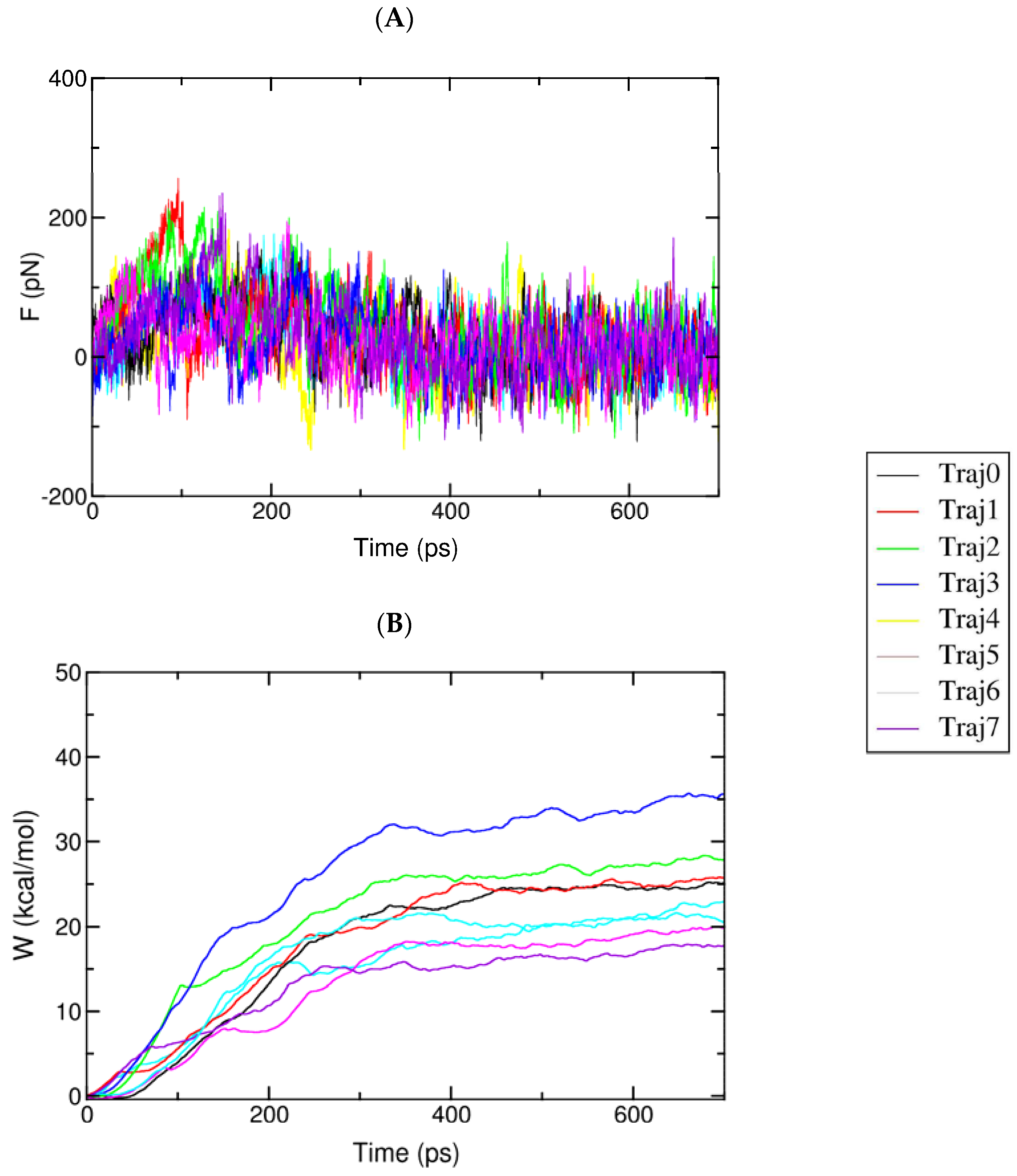
2.5. FPL Simulation
2.6. Drug-Likeness Studies
3. Conclusions
4. Materials and Methods
4.1. Source of Essential Oil
4.2. Acetylcholinesterase (AChE) Inhibition Assay
4.3. Molecular Docking Studies
4.4. Drug-Likeness Studies
4.5. Binding Affinity Calculation
4.6. Fast Pulling Ligand Simulation
4.7. Data Analysis
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Sample Availability
References
- Trang, A.; Khandhar, P.B. Physiology, Acetylcholinesterase. In StatPearls; StatPearls Publishing: Treasure Island, FL, USA, 2022. Available online: https://www.ncbi.nlm.nih.gov/books/NBK539735/ (accessed on 4 June 2022).
- Colovic, M.B.; Krstic, D.Z.; Lazarevic-Pasti, T.D.; Bondzic, A.M.; Vasic, V.M. Acetylcholinesterase inhibitors: Pharmacology and toxicology. Curr. Neuropharmacol. 2013, 11, 315–335. [Google Scholar] [CrossRef]
- Fukuto, T.R. Mechanism of action of organophosphorus and carbamate insecticides. Environ. Health Perspect. 1990, 87, 245–254. [Google Scholar] [CrossRef]
- Pope, C.; Karanth, S.; Liu, J. Pharmacology and toxicology of cholinesterase inhibitors: Uses and misuses of a common mechanism of action. Environ. Toxicol. Pharmacol. 2005, 19, 433–446. [Google Scholar] [CrossRef]
- Fournier, D. Mutations of acetylcholinesterase which confer insecticide resistance in insect populations. Chem. Biol. Interact. 2005, 157–158, 257–261. [Google Scholar] [CrossRef]
- Kihara, T.; Shimohama, S. Alzheimer’s disease and acetylcholine receptors. Acta Neurobiol. Exp. 2004, 64, 99–105. [Google Scholar]
- Mehta, M.; Adem, A.; Sabbagh, M. New acetylcholinesterase inhibitors for alzheimer’s disease. Int. J. Alzheimer’s Dis. 2012, 2012, 728983. [Google Scholar] [CrossRef]
- Singh, M.; Kaur, M.; Kukreja, H.; Chugh, R.; Silakari, O.; Singh, D. Acetylcholinesterase inhibitors as alzheimer therapy: From nerve toxins to neuroprotection. Eur. J. Med. Chem. 2013, 70, 165–188. [Google Scholar] [CrossRef]
- World Health Organization. Alzheimer’s Disease: Help for Caregivers; World Health Organization: Geneva, Switzerland, 1994. [Google Scholar]
- Knapp, M.J. A 30-week randomized controlled trial of high-dose tacrine in patients with alzheimer’s disease. JAMA J. Am. Med. Assoc. 1994, 271, 985. [Google Scholar] [CrossRef]
- Ibach, B.; Haen, E. Acetylcholinesterase inhibition in alzheimers disease. Curr. Pharm. Des. 2004, 10, 231–251. [Google Scholar] [CrossRef]
- Gust, C.; Pugliese, N.; Stern, G. Suspected donepezil toxicity: A case report. Clin. Case Rep. 2020, 8, 2817–2822. [Google Scholar] [CrossRef]
- Lee, D.H.; Choi, Y.H.; Cho, K.H.; Yun, S.Y.; Lee, H.M. A case of rivastigmine toxicity caused by transdermal patch. Am. J. Emerg. Med. 2011, 29, 695.e1–e2. [Google Scholar] [CrossRef]
- Dobetsberger, C.; Buchbauer, G. Actions of essential oils on the central nervous system: An updated review. Flavour Fragr. J. 2011, 26, 300–316. [Google Scholar] [CrossRef]
- Lahlou, M. Essential oils and fragrance compounds: Bioactivity and mechanisms of action. Flavour Fragr. J. 2004, 19, 159–165. [Google Scholar] [CrossRef]
- Lomarat, P.; Sripha, K.; Phanthong, P.; Kitphati, W.; Thirapanmethee, K.; Bunyapraphatsara, N. In vitro biological activities of black pepper essential oil and its major components relevant to the prevention of alzheimer’s disease. Thai. J. Pharm. Sci. 2015, 39, 94–101. [Google Scholar]
- Ayaz, M.; Junaid, M.; Ullah, F.; Sadiq, A.; Khan, M.A.; Ahmad, W.; Shah, M.R.; Imran, M.; Ahmad, S. Comparative chemical profiling, cholinesterase inhibitions and anti-radicals properties of essential oils from Polygonum hydropiper L.: A preliminary anti-alzheimer’s study. Lipids Health Dis. 2015, 14, 141. [Google Scholar] [CrossRef]
- Flanagan, N. The clinical use of aromatherapy in alzheimer’s patients. Altern. Complement. Ther. 1995, 1, 377–380. [Google Scholar] [CrossRef]
- Jimbo, D.; Kimura, Y.; Taniguchi, M.; Inoue, M.; Urakami, K. Effect of aromatherapy on patients with alzheimer’s disease. Psychogeriatrics 2009, 9, 173–179. [Google Scholar] [CrossRef]
- Fung, J.K.K.; Tsang, H.W.; Chung, R.C. A Systematic review of the use of aromatherapy in treatment of behavioral problems in dementia. Geriatr. Gerontol. Int. 2012, 12, 372–382. [Google Scholar] [CrossRef]
- Scuteri, D.; Morrone, L.A.; Rombolà, L.; Avato, P.R.; Bilia, A.R.; Corasaniti, M.T.; Sakurada, S.; Sakurada, T.; Bagetta, G. Aromatherapy and aromatic plants for the treatment of behavioural and psychological symptoms of dementia in patients with alzheimer’s disease: Clinical evidence and possible mechanisms. Evid.-Based Complement. Altern. Med. 2017, 2017, 9416305. [Google Scholar] [CrossRef]
- Lin, L.; Duan, R.; Yang, Q.; Li, T.; Zhou, H.; Hou, J.; Zhou, H. Effect of Aromatherapy in Patients with Alzheimer’s Disease: A Randomised Controlled Clinical Trial. Res. Sq. 2022. [Google Scholar] [CrossRef]
- Okuda, M.; Fujita, Y.; Takada-Takatori, Y.; Sugimoto, H.; Urakami, K. Aromatherapy improves cognitive dysfunction in senescence-accelerated mouse prone 8 by reducing the level of amyloid beta and tau phosphorylation. PLoS ONE 2020, 15, e0240378. [Google Scholar] [CrossRef] [PubMed]
- Chi, V.V. Dictionary of Vietnamese Medicinal Plants; Medical Publishing House: Hanoi, Vietnam, 2012. [Google Scholar]
- Loi, D.T. Vietnamese Medicinal Plants and Herbs; Medical Publishing House: Hanoi, Vietnam, 2005. [Google Scholar]
- Bich, D.H.; Chung, D.Q.; Chuong, B.X.; Dong, N.T.; Dam, D.T.; Hien, P.V.; Lo, V.N.; Mai, P.D.; Man, P.K.; Nhu, D.T.; et al. The Medicinal Plants and Animals in Vietnam; Science and Technology Publishing House: Hanoi, Vietnam, 2006; Volume 2. [Google Scholar]
- Hung, N.H.; Huong, L.T.; Chung, N.T.; Thuong, N.T.H.; Satyal, P.; Dung, N.A.; Tai, T.A.; Setzer, W.N. Callicarpa species from central Vietnam: Essential oil compositions and mosquito larvicidal activities. Plants 2020, 9, 113. [Google Scholar] [CrossRef] [PubMed]
- Hung, N.H.; Huong, L.T.; Chung, N.T.; Truong, N.C.; Dai, D.N.; Satyal, P.; Tai, T.A.; Hien, V.T.; Setzer, W.N. Premna species in Vietnam: Essential oil compositions and mosquito larvicidal activities. Plants 2020, 9, 1130. [Google Scholar] [CrossRef]
- Hung, N.H.; Dai, D.N.; Satyal, P.; Huong, L.T.; Chinh, B.T.; Tai, T.A.; Hien, V.T.; Setzer, W.N. Investigation of pesticidal activities of essential oils obtained from Vitex species. Rec. Nat. Prod. 2022, 16, 268–273. [Google Scholar] [CrossRef]
- Miyazawa, M.; Watanabe, H.; Kameoka, H. Inhibition of acetylcholinesterase activity by monoterpenoids with a p-menthane skeleton. J. Agric. Food Chem. 1997, 45, 677–679. [Google Scholar] [CrossRef]
- Bonesi, M.; Menichini, F.; Tundis, R.; Loizzo, M.R.; Conforti, F.; Passalacqua, N.G.; Statti, G.A.; Menichini, F. Acetylcholinesterase and butyrylcholinesterase inhibitory activity of Pinus species essential oils and their constituents. J. Enzym. Inhib. Med. Chem. 2010, 25, 622–628. [Google Scholar] [CrossRef]
- Savelev, S.U.; Okello, E.J.; Perry, E.K. Butyryl- and Acetyl-cholinesterase inhibitory activities in essential oils of Salvia species and their constituents. Phyther. Res. 2004, 18, 315–324. [Google Scholar] [CrossRef]
- Karakaya, S.; Yilmaz, S.V.; Özdemir, Ö.; Koca, M.; Pınar, N.M.; Demirci, B.; Yıldırım, K.; Sytar, O.; Turkez, H.; Baser, K.H.C. A caryophyllene oxide and other potential anticholinesterase and anticancer agent in Salvia verticillata subsp. amasiaca (Freyn & Bornm.) Bornm. (Lamiaceae). J. Essent. Oil Res. 2020, 32, 512–525. [Google Scholar] [CrossRef]
- Savelev, S.; Okello, E.; Perry, N.S.L.; Wilkins, R.M.; Perry, E.K. Synergistic and antagonistic interactions of anticholinesterase terpenoids in Salvia lavandulaefolia essential oil. Pharmacol. Biochem. Behav. 2003, 75, 661–668. [Google Scholar] [CrossRef]
- Miyazawa, M.; Yamafuji, C. Inhibition of acetylcholinesterase activity by bicyclic monoterpenoids. J. Agric. Food Chem. 2005, 53, 1765–1768. [Google Scholar] [CrossRef]
- Kaufmann, D.; Dogra, A.K.; Wink, M. Myrtenal inhibits acetylcholinesterase, a known Alzheimer target. J. Pharm. Pharmacol. 2011, 63, 1368–1371. [Google Scholar] [CrossRef] [PubMed]
- Salinas, M.; Bec, N.; Calva, J.; Ramirez, J.; Andrade, J.M.; Larroque, C.; Vidari, G.; Armijos, C. Chemical composition and anticholinesterase activity of the essential oil from the Ecuadorian plant Salvia pichinchensis Benth. Rec. Nat. Prod. 2020, 14, 276–285. [Google Scholar] [CrossRef]
- Ali, N.A.A.; Crouch, R.A.; Al-Fatimi, M.A.; Arnold, N.; Teichert, A.; Setzer, W.N.; Wessjohann, L. Chemical composition, antimicrobial, antiradical and anticholinesterase activity of the essential oil of Pulicaria stephanocarpa from Soqotra. Nat. Prod. Commun. 2012, 7, 113–116. [Google Scholar] [CrossRef]
- Siebert, D.A.; Tenfen, A.; Yamanaka, C.N.; de Cordova, C.M.M.; Scharf, D.R.; Simionatto, E.L.; Alberton, M.D. Evaluation of seasonal chemical composition, antibacterial, antioxidant and anticholinesterase activity of essential oil from Eugenia brasiliensis Lam. Nat. Prod. Res. 2015, 29, 289–292. [Google Scholar] [CrossRef]
- Valarezo, E.; Ludeña, J.; Echeverria-Coronel, E.; Cartuche, L.; Meneses, M.A.; Calva, J.; Morocho, V. Enantiomeric composition, antioxidant capacity and anticholinesterase activity of essential oil from leaves of Chirimoya (Annona cherimola Mill.). Plants 2022, 11, 367. [Google Scholar] [CrossRef]
- de Souza, A.; Lopes, E.M.C.; da Silva, M.C.; Cordeiro, I.; Young, M.C.M.; Sobral, M.E.G.; Moreno, P.R.H. Chemical composition and acetylcholinesterase inhibitory activity of essential oils of Myrceugenia myrcioides (Cambess.) O. Berg and Eugenia riedeliana O. Berg, Myrtaceae. Rev. Bras. Farmacogn. Braz. J. Pharmacogn. 2010, 20, 175–179. [Google Scholar] [CrossRef]
- Venditti, A.; Frezza, C.; Sciubba, F.; Serafini, M.; Bianco, A.; Cianfaglione, K.; Lupidi, G.; Quassinti, L.; Bramucci, M.; Maggi, F. Volatile components, polar constituents and biological activity of tansy daisy (Tanacetum macrophyllum (Waldst. et Kit.) Schultz Bip.). Ind. Crops Prod. 2018, 118, 225–235. [Google Scholar] [CrossRef]
- da Silva Barbosa, D.C.; Holanda, V.N.; de Assis, C.R.D.; de Oliveira Farias de Aguiar, J.C.R.; DoNascimento, P.H.; da Silva, W.V.; do Amaral Ferraz Navarro, D.M.; da Silva, M.V.; de Menezes Lima, V.L.; dos Santos Correia, M.T. Chemical composition and acetylcholinesterase inhibitory potential, in silico, of Myrciaria floribunda (H. West Ex Willd.) O. Berg fruit peel essential oil. Ind. Crops Prod. 2020, 151, 112372. [Google Scholar] [CrossRef]
- Miyazawa, M.; Nakahashi, H.; Usami, A.; Matsuda, N. Chemical composition, aroma evaluation, and inhibitory activity towards acetylcholinesterase of essential oils from Gynura bicolor DC. J. Nat. Med. 2016, 70, 282–289. [Google Scholar] [CrossRef]
- Liu, T.-T.; Chao, L.K.-P.; Hong, K.-S.; Huang, Y.-J.; Yang, T.-S. Composition and insecticidal activity of essential oil of Bacopa caroliniana and interactive effects of individual compounds on the activity. Insects 2019, 11, 23. [Google Scholar] [CrossRef]
- Lin, Z.; Xiao, Z.; Zhu, D.; Yan, Y.; Yu, B.; Wang, Q. Aqueous extracts of FBD, a chinese herb formula composed of Poria cocos, Atractylodes macrocephala, and Angelica sinensis reverse scopolamine induced memory deficit in ICR mice. Pharm. Biol. 2009, 47, 396–401. [Google Scholar] [CrossRef]
- Xu, J.; Guo, Y.; Zhao, P.; Xie, C.; Jin, D.-Q.; Hou, W.; Zhang, T. Neuroprotective cadinane sesquiterpenes from the resinous exudates of Commiphora myrrha. Fitoterapia 2011, 82, 1198–1201. [Google Scholar] [CrossRef] [PubMed]
- Amorim, A.C.L.; Lima, C.K.F.; Hovell, A.M.C.; Miranda, A.L.P.; Rezende, C.M. Antinociceptive and hypothermic evaluation of the leaf essential oil and isolated terpenoids from Eugenia uniflora L. (Brazilian Pitanga). Phytomedicine 2009, 16, 923–928. [Google Scholar] [CrossRef] [PubMed]
- Wilhelm, E.A.; Torres, M.L.C.P.; Pereira, C.F.; Vogt, A.G.; Cervo, R.; dos Santos, B.G.T.; Cargnelutti, R.; Luchese, C. Therapeutic potential of selanyl amide derivatives in the in vitro anticholinesterase activity and in in vivo antiamnesic action. Can. J. Physiol. Pharmacol. 2020, 98, 304–313. [Google Scholar] [CrossRef]
- Wright, B.S.; Bansal, A.; Moriarity, D.M.; Takaku, S.; Setzer, W.N. Cytotoxic leaf essential oils from Neotropical Lauraceae: Synergistic effects of essential oil components. Nat. Prod. Commun. 2007, 2, 1241–1244. [Google Scholar] [CrossRef]
- Menichini, F.; Tundis, R.; Loizzo, M.R.; Bonesi, M.; Marrelli, M.; Statti, G.A.; Menichini, F.; Conforti, F. Acetylcholinesterase and butyrylcholinesterase inhibition of ethanolic extract and monoterpenes from Pimpinella anisoides V Brig. (Apiaceae). Fitoterapia 2009, 80, 297–300. [Google Scholar] [CrossRef]
- Picollo, M.I.; Toloza, A.C.; Mougabure Cueto, G.; Zygadlo, J.; Zerba, E. Anticholinesterase and pediculicidal activities of monoterpenoids. Fitoterapia 2008, 79, 271–278. [Google Scholar] [CrossRef]
- Dohi, S.; Terasaki, M.; Makino, M. Acetylcholinesterase inhibitory activity and chemical composition of commercial essential oils. J. Agric. Food Chem. 2009, 57, 4313–4318. [Google Scholar] [CrossRef]
- Gupta, S.; Kathait, A.; Sharma, V. Computational sequence analysis and structure prediction of jack bean urease. Int. J. Adv. Res. 2015, 3, 185–191. [Google Scholar]
- Benkert, P.; Biasini, M.; Schwede, T. Toward the estimation of the absolute quality of individual protein structure models. Bioinformatics 2011, 27, 343–350. [Google Scholar] [CrossRef]
- Zhang, Y. TM-Align: A Protein structure alignment algorithm based on the TM-Score. Nucleic Acids Res. 2005, 33, 2302–2309. [Google Scholar] [CrossRef] [PubMed]
- Quan, P.M.; Huong, L.T.T.; Toan, T.Q.; Hung, N.P.; Nam, P.H.; Kiet, N.T.; Ha, N.X.; Le, D.T.T.; An, T.N.T.; Show, P.L.; et al. Cannabis sativa L. chemical compositions as potential plasmodium falciparum dihydrofolate reductase-thymidinesynthase enzyme inhibitors: An in silico study for drug development. Open Chem. 2021, 19, 1235–1241. [Google Scholar] [CrossRef]
- Bui, H.T.B.; Nguyen, P.H.; Pham, Q.M.; Tran, H.P.; Tran, D.Q.; Jung, H.; Hong, Q.V.; Nguyen, Q.C.; Nguyen, Q.P.; Le, H.T.; et al. Target design of novel histone deacetylase 6 selective inhibitors with 2-mercaptoquinazolinone as the cap moiety. Molecules 2022, 27, 2204. [Google Scholar] [CrossRef]
- Gohlke, H.; Hendlich, M.; Klebe, G. Knowledge-based scoring function to predict protein-ligand interactions. J. Mol. Biol. 2000, 295, 337–356. [Google Scholar] [CrossRef]
- Gowthaman, U.; Jayakanthan, M.; Sundar, D. Molecular docking studies of dithionitrobenzoic acid and its related compounds to protein disulfide isomerase: Computational screening of inhibitors to HIV-1 entry. BMC Bioinform. 2008, 9, S14. [Google Scholar] [CrossRef]
- Nguyen, N.T.; Nguyen, T.H.; Pham, T.N.H.; Huy, N.T.; Van Bay, M.; Pham, M.Q.; Nam, P.C.; Vu, V.V.; Ngo, S.T. Autodock vina adopts more accurate binding poses but autodock4 forms better binding affinity. J. Chem. Inf. Model. 2020, 60, 204–211. [Google Scholar] [CrossRef]
- Pham, M.Q.; Van Tran, T.H.; Pham, Q.L.; Gairin, J.E. In silico analysis of the binding properties of solasonine to mortalin and p53, and in vitro pharmacological studies of its apoptotic and cytotoxic effects on human HepG2 and Hep3b hepatocellular carcinoma cells. Fundam. Clin. Pharmacol. 2019, 33, 385–396. [Google Scholar] [CrossRef]
- Loh, Z.H.; Kwong, H.C.; Lam, K.W.; Teh, S.S.; Ee, G.C.L.; Quah, C.K.; Ho, A.S.H.; Mah, S.H. New 3-O-substituted xanthone derivatives as promising acetylcholinesterase inhibitors. J. Enzym. Inhib. Med. Chem. 2021, 36, 627–639. [Google Scholar] [CrossRef]
- Khodja, I.A.; Boulebd, H. Synthesis, biological evaluation, theoretical investigations, docking study and ADME parameters of some 1,4-bisphenylhydrazone derivatives as potent antioxidant agents and acetylcholinesterase inhibitors. Mol. Divers. 2021, 25, 279–290. [Google Scholar] [CrossRef] [PubMed]
- Ngo, S.T.; Hung, H.M.; Nguyen, M.T. Fast and accurate determination of the relative binding affinities of small compounds to HIV-1 protease using non-equilibrium work. J. Comput. Chem. 2016, 37, 2734–2742. [Google Scholar] [CrossRef]
- Hassan, S.S.U.; Abbas, S.Q.; Ali, F.; Ishaq, M.; Bano, I.; Hassan, M.; Jin, H.-Z.; Bungau, S.G. A Comprehensive In Silico exploration of pharmacological properties, bioactivities, molecular docking, and anticancer potential of vieloplain F from Xylopia vielana targeting B-Raf kinase. Molecules 2022, 27, 917. [Google Scholar] [CrossRef] [PubMed]
- Ellman, G.L.; Courtney, K.D.; Andres, V.; Featherstone, R.M. A New and rapid colorimetric determination of acetylcholinesterase activity. Biochem. Pharmacol. 1961, 7, 88–95. [Google Scholar] [CrossRef]
- Hung, N.H.; Dai, D.N.; Satyal, P.; Huong, L.T.; Chinh, B.T.; Hung, D.Q.; Tai, T.A.; Setzer, W.N. Lantana camara essential oils from Vietnam: Chemical composition, molluscicidal, and mosquito larvicidal activity. Chem. Biodivers. 2021, 18, e2100145. [Google Scholar] [CrossRef] [PubMed]
- Schrodinger, L. The PyMOL Molecular Graphics System, Version 1.3r1; Schrodinger: New York, NY, USA, 2010.
- Frisch, M.J.; Trucks, G.W.; Schlegel, H.B.; Scuseria, G.E.; Robb, M.A.; Cheeseman, J.R. Gaussian 09 Rev. d.01; Gaussian Inc.: Wallingford, CT, USA, 2009. [Google Scholar]
- Bourne, Y.; Grassi, J.; Bougis, P.E.; Marchot, P. Conformational flexibility of the acetylcholinesterase tetramer suggested by x-ray crystallography. J. Biol. Chem. 1999, 274, 30370–30376. [Google Scholar] [CrossRef]
- Morris, G.M.; Huey, R.; Lindstrom, W.; Sanner, M.F.; Belew, R.K.; Goodsell, D.S.; Olson, A.J. AutoDock4 and AutoDockTools4: Automated docking with selective receptor flexibility. J. Comput. Chem. 2009, 30, 2785–2791. [Google Scholar] [CrossRef]
- Dassault Systèmes BIOVIA. Discovery Studio Visualizer, V21.1.0.20298; Dassault Systèmes: San Diego, CA, USA, 2021. [Google Scholar]
- Cheung, J.; Rudolph, M.J.; Burshteyn, F.; Cassidy, M.S.; Gary, E.N.; Love, J.; Franklin, M.C.; Height, J.J. Structures of human acetylcholinesterase in complex with pharmacologically important ligands. J. Med. Chem. 2012, 55, 10282–10286. [Google Scholar] [CrossRef]
- Greenidge, P.A.; Kramer, C.; Mozziconacci, J.-C.; Wolf, R.M. MM/GBSA binding energy prediction on the PDBbind data set: Successes, failures, and directions for further improvement. J. Chem. Inf. Model. 2013, 53, 201–209. [Google Scholar] [CrossRef]
- Zhang, X.; Perez-Sanchez, H.; Lightstone, F.C. A comprehensive docking and MM/GBSA rescoring study of ligand recognition upon binding antithrombin. Curr. Top. Med. Chem. 2017, 17, 1631–1639. [Google Scholar] [CrossRef]
- Petrek, M.; Otyepka, M.; Banáš, P.; Košinová, P.; Koča, J.; Damborský, J. CAVER: A new tool to explore routes from protein clefts, pockets and cavities. BMC Bioinform. 2006, 7, 316. [Google Scholar] [CrossRef]
- Pham, M.Q.; Vu, K.B.; Pham, T.N.H.; Huong, L.T.T.; Tran, L.H.; Tung, N.T.; Vu, V.V.; Nguyen, T.H.; Ngo, S.T. Rapid prediction of possible inhibitors for SARS-CoV-2 main protease using docking and FPL simulations. RSC Adv. 2020, 10, 31991–31996. [Google Scholar] [CrossRef]
- Lindorff-Larsen, K.; Piana, S.; Palmo, K.; Maragakis, P.; Klepeis, J.L.; Dror, R.O.; Shaw, D.E. Improved side-chain torsion potentials for the amber Ff99SB protein force field. Proteins Struct. Funct. Bioinform. 2010, 78, 1950–1958. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Wang, W.; Kollman, P.A.; Case, D.A. Automatic atom type and bond type perception in molecular mechanical calculations. J. Mol. Graph. Model. 2006, 25, 247–260. [Google Scholar] [CrossRef] [PubMed]
- Jorgensen, W.L.; Chandrasekhar, J.; Madura, J.D.; Impey, R.W.; Klein, M.L. Comparison of simple potential functions for simulating liquid water. J. Chem. Phys. 1983, 79, 926–935. [Google Scholar] [CrossRef]
- Darden, T.; York, D.; Pedersen, L. Particle mesh Ewald: An N⋅log(N) method for Ewald sums in large systems. J. Chem. Phys. 1993, 98, 10089–10092. [Google Scholar] [CrossRef]
- Finney, D. Probit Analysis, Reissue, ed.; Cambridge University Press: Cambridge, UK, 2009; ISBN 978-0521135900. [Google Scholar]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Essential Oil | Major Components of the Essential Oils | Classification |
|---|---|---|
| Callicarpa candicans (Nghe An) [27] | Atractylone (37.7%), (E)-β-caryophyllene (19.0%), β-selinene (6.2%). | Ses: 95% (hydro: 49%; oxy: 46%) |
| Callicarpa candicans (Da Nang) [27] | Atractylone (42.4%), (E)-β-caryophyllene (15.3%), curzerene (5.3%), germacrene B (5.1%), β-selinene (4.5%). | Ses: 92.2% (hydro: 40.7%; oxy: 51.5%) |
| Callicarpa candicans (Quang Nam) [27] | Caryophyllene oxide (13.4%), (E)-β-caryophyllene (7.1%), β-selinene (5.7%). | Ses: 52.5% (hydro: 22.6%; oxy: 29.9%) compared with 59.7% of the identified compounds. |
| Callicarpa longifolia [27] | trans-β-Guaiene (22.2%), (E)-β-caryophyllene (11.8%), selin-11-en-4α-ol (8.0%), 1(10),11-eremophiladien-9-one (6.7%). | Ses: 92.4% (hydro: 63.0%; oxy: 29.4%). |
| Callicarpa sinuate [27] | α-Humulene (24.8%), α-copaene (12.6%), humulene epoxide II (6.7%), spathulenol (5.9%). | Ses: 91.6% (hydro: 67.5%; oxy: 24.1%). |
| Callicarpa petelotii [27] | α-Humulene (53.8%), α-selinene (12.8%), humulene epoxide II (8.1%). | Ses: 91.0% (hydro: 74.7%; oxy: 16.3%). |
| Callicarpa rubella (Tay Giang) [27] | (E)-Caryophyllene (18.0%), α-cubebene (17.4%). | Ses: 87.5% (hydro: 78.5%; oxy: 9.0%). |
| Callicarpa nudiflora [27] | β-Pinene (34.2%), caryophyllene oxide (20.1%), α-pinene (8.1%), myrtenal (6.8%). | Mono: 62.1% (hydro: 47.5%; oxy: 14.6%); ses: 32.8% (hydro: 5.7%, oxy: 27.1%). |
| Callicarpa erioclona * | Atractylone (34.6%), (E)-β-caryophyllene (11.1%), caryophyllene oxide (5.9%), β-selinene (5.1%). | Ses: 75.4% (hydro: 32.1%; oxy: 43.3%) compared with 79.6% of the identified compounds. |
| Callicarpa macrophylla (Pu Mat) * | (E)-Caryophyllene (25.2%), caryophyllene oxide (6.4%). | Ses: 73.9% (hydro: 62.0%; oxy: 11.9%). |
| Prmena cambodiana [28] | α-Copaene (23.3%), (E)-caryophyllene (12.8%), α-gurjunene (11.3%), δ-cadinene (5.5%). | Ses: 88.4% (hydro: 76.1%; oxy: 12.3%). |
| Prmena corymbosa [28] | allo-Aromadendrene (39.7%), (E)-caryophyllene (13.3%), α-copaene (8.1%). | Ses: 93.6% (hydro: 85.4%; oxy: 8.2%). |
| Vitex ajugifolia [29] | α-Copaene (17.0%), (E)-β-caryophyllene (11.7%), α-humulene (9.6%), spathulenol (8.7%). | Ses: 97.8% (hydro: 68.2%; oxy: 29.6%). |
| Vitex trifolia subsp. litoralis [29] | α-Pinene (18.7%), sabinene (15.2%), 1,8-cineole (14.5%), α-terpinyl acetate (12.7%). | Mono: 76.0% (hydro: 44.9%; oxy: 31.1%); ses: 1.6% (hydro: 1.1%, oxy: 0.5%). |
| Vitex pinnata [29] | (E)-β-Caryophyllene (32.7%), germacrene D (17.1%), bicyclogermacrene (11.1%). | Ses: 95.8% (hydro: 87.1%; oxy: 8.7%). |
| Vitex trifolia subsp. trifolia [29] | Sabinene (19.4%), 1,8-cineole (15.7%), (E)-β-caryophyllene (14.5%), α-pinene (11.7%), α-terpinyl acetate (8.3%). | Mono: 38.4% (hydro: 17.6%; oxy: 20.8%); ses: 39.2% (hydro: 31.5%, oxy: 7.7%). |
| Karomia fragrans * | (E)-Caryophyllene (26.5%), caryophyllene oxide (10.5%), α-humulene (10.1%), p-cymene (7.5%), δ-cadinene (5.2%). | Ses: 73.1% (hydro: 51.5%; oxy: 21.6%). |
| Concentration (µg/mL) | C. candicans (Nghe An) | C. candicans (Da Nang) | C. candicans (Quang Nam) | C. longifolia | C. sinuata | C. petelotii | C. rubella (Tay Giang) | C. nudiflora | ||
|---|---|---|---|---|---|---|---|---|---|---|
| 500 | 87.88 | 86.37 | 88.34 | 82.72 | 82.13 | 90.66 | 89.78 | 89.78 | ||
| 100 | 59.57 | 66.53 | 61.00 | 52.77 | 66.71 | 62.70 | 52.24 | 69.35 | ||
| 20 | 32.61 | 37.46 | 34.05 | 19.55 | 50.91 | 47.36 | 25.60 | 25.26 | ||
| 4 | 5.28 | 10.42 | 10.27 | 8.06 | 15.47 | 21.03 | 7.12 | 3.67 | ||
| IC50 | 58.38 ± 2.95 | 45.67 ± 1.84 | 52.88 ± 3.19 | 105.16 ± 5.61 | 34.15 ± 1.35 | 32.11 ± 3.85 | 89.38 ± 4.05 | 54.69 ± 3.05 | ||
| Concentration (µg/mL) | C. erioclona | C. macrophylla (Pu Mat) | P. cambodiana | P. corymbosa | V. ajugifolia | V. trifolia subsp. litoralis | V. pinnata | V. trifolia subsp. trifolia | K. fragrans | |
| 500 | 100.00 | 75.00 | 81.83 | 97.55 | 82.58 | 103.82 | 83.79 | 82.02 | 75.67 | |
| 100 | 74.06 | 33.60 | 42.30 | 59.14 | 65.02 | 49.53 | 44.37 | 57.21 | 40.66 | |
| 20 | 44.70 | 6.72 | 17.09 | 24.67 | 38.06 | 3.79 | 23.22 | 22.92 | 6.43 | |
| 4 | 26.77 | −1.56 | 8.61 | 11.54 | 7.55 | −4.77 | 17.59 | −1.19 | −0.41 | |
| IC50 | 28.71 ± 3.24 | 221.85 ± 15.32 | 157.06 ± 4.14 | 73.35 ± 4.61 | 50.93 ± 4.81 | 120.32 ± 16.64 | 144.33 ± 16.94 | 81.34 ± 3.57 | 187.91 ± 14.09 | |
| Concentration (µg/mL) | (E)-β-Caryophyllene | β-Pinene | α-Humulene | Limonene | Caryophyllene Oxide | trans-Carveol | Galantamine b |
|---|---|---|---|---|---|---|---|
| 500 | 79.05 | 86.54 | 72.83 | 90.82 | 74.00 | 76.90 | --- |
| 100 | 52.89 | 58.86 | 44.27 | 67.73 | 16.95 | 55.34 | --- |
| 20 | 26.28 | 28.88 | 25.10 | 27.04 | 0.49 | 16.79 | --- |
| 4 | 6.09 | 20.92 | 19.37 | 18.17 | −2.11 | 1.19 | --- |
| IC50 (μg/mL) | 89.10 ± 6.10 | 71.45 ± 5.77 | 160.48 ± 13.48 | 53.16 ± 4.08 | 320.16 ± 13.47 | 102.88 ± 7.84 | 1.78 ± 0.13b |
| IC50 (μM) | 436.0 ± 29.9 | 524.5 ± 42.4 | 785.3 ± 66.0 | 390.2 ± 30.0 | 1453 ± 61 | 675.8 ± 51.5 | 6.19 ± 0.45 |
| Parameters | |
|---|---|
| Number of amino acids | 91 |
| Molecular weight | 10,716.71 |
| Theoretical pI | 4.76 |
| Aliphatic index | 44.95 |
| Secondary Structure | Number of Amino Acids | Percentage (%) |
|---|---|---|
| Alpha helix | 33 | 36.26 |
| 310 helix | 0 | 0.00 |
| Pi helix | 0 | 0.00 |
| Beta bridge | 0 | 0.00 |
| Extended strand | 13 | 14.29 |
| Beta turn | 4 | 4.40 |
| Bend region | 0 | 0.00 |
| Random coil | 41 | 45.05 |
| Ambiguous states | 0 | 0.00 |
| Other states | 0 | 0.00 |
| Ligand | Dock Score (kcal/mol) | MM-GBSA (kcal/mol) | Interacting Residues |
|---|---|---|---|
| (E)-β-Caryophyllene | −8.79 | −105.41 | Trp286, Phe297 |
| β-Pinene | −9.24 | −116.32 | Trp86, Tyr337, His447 |
| α-Humulene | −6.95 | −43.19 | Trp86, Tyr337 |
| Limonene | −9.78 | −145.78 | Phe297, Phe338, Tyr341 |
| Caryophyllene oxide | −5.00 | −40.56 | Phe338, Tyr341 |
| trans-Carveol | −6.35 | −46.22 | Ile294, Phe295, Phe297, Tyr337, Phe338, Tyr34 |
| Galantamine | −12.76 | −154.63 | Trp86, Gly121, Gly122, Tyr124, Glu202, Ser203, Phe295, Phe297, Tyr337, Phe338, His447 |
| Compound | HIA (%) a | CNS (logPS) b | Mutagenic | Tumorigenic |
|---|---|---|---|---|
| (E)-β-Caryophyllene | 94.09 | −2.139 | NO | NO |
| β-Pinene | 95.43 | −1.847 | NO | NO |
| α-Humulene | 94.43 | −2.542 | NO | NO |
| Limonene | 95.40 | −2.356 | NO | NO |
| Caryophyllene oxide | 95.88 | −2.518 | NO | YES |
| trans-Carveol | 94.69 | −2.637 | NO | NO |
| Galantamine | 96.46 | −2.559 | NO | NO |
| Essential Oil | Yield (% v/w) | Part |
|---|---|---|
| Callicarpa candicans (Nghe An) | 0.15 | Leaves |
| Callicarpa candicans (Da Nang) | 0.17 | Leaves |
| Callicarpa candicans (Quang Nam) | 0.18 | Leaves |
| Callicarpa longifolia | 0.13 | Leaves |
| Callicarpa sinuata | 0.14 | Leaves |
| Callicarpa petelotii | 0.22 | Leaves |
| Callicarpa rubella (Tay Giang) | 0.12 | Leaves |
| Callicarpa nudiflora | 0.14 | Leaves |
| Callicarpa erioclona | 0.19 | Leaves |
| Callicarpa macrophylla (Pu Mat) | 0.24 | Leaves |
| Prmena cambodiana | 0.14 | Leaves |
| Prmena corymbosa | 0.25 | Leaves |
| Vitex ajugifolia | 0.09 | Leaves |
| Vitex trifolia subsp. litoralis | 0.12 | Leaves |
| Vitex pinnata | 0.14 | Leaves |
| Vitex trifolia subsp. trifolia | 0.12 | Leaves |
| Karomia fragrans | 0.12 | Leaves |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hung, N.H.; Quan, P.M.; Satyal, P.; Dai, D.N.; Hoa, V.V.; Huy, N.G.; Giang, L.D.; Ha, N.T.; Huong, L.T.; Hien, V.T.; et al. Acetylcholinesterase Inhibitory Activities of Essential Oils from Vietnamese Traditional Medicinal Plants. Molecules 2022, 27, 7092. https://doi.org/10.3390/molecules27207092
Hung NH, Quan PM, Satyal P, Dai DN, Hoa VV, Huy NG, Giang LD, Ha NT, Huong LT, Hien VT, et al. Acetylcholinesterase Inhibitory Activities of Essential Oils from Vietnamese Traditional Medicinal Plants. Molecules. 2022; 27(20):7092. https://doi.org/10.3390/molecules27207092
Chicago/Turabian StyleHung, Nguyen Huy, Pham Minh Quan, Prabodh Satyal, Do Ngoc Dai, Vo Van Hoa, Ngo Gia Huy, Le Duc Giang, Nguyen Thi Ha, Le Thi Huong, Vu Thi Hien, and et al. 2022. "Acetylcholinesterase Inhibitory Activities of Essential Oils from Vietnamese Traditional Medicinal Plants" Molecules 27, no. 20: 7092. https://doi.org/10.3390/molecules27207092
APA StyleHung, N. H., Quan, P. M., Satyal, P., Dai, D. N., Hoa, V. V., Huy, N. G., Giang, L. D., Ha, N. T., Huong, L. T., Hien, V. T., & Setzer, W. N. (2022). Acetylcholinesterase Inhibitory Activities of Essential Oils from Vietnamese Traditional Medicinal Plants. Molecules, 27(20), 7092. https://doi.org/10.3390/molecules27207092