
Chemical Composition, Antifungal and Anti-Biofilm Activities of Volatile Fractions of Convolvulus althaeoides L. Roots from Tunisia

 ,
,  ,
, 
Abstract
1. Introduction
2. Results and Discussion
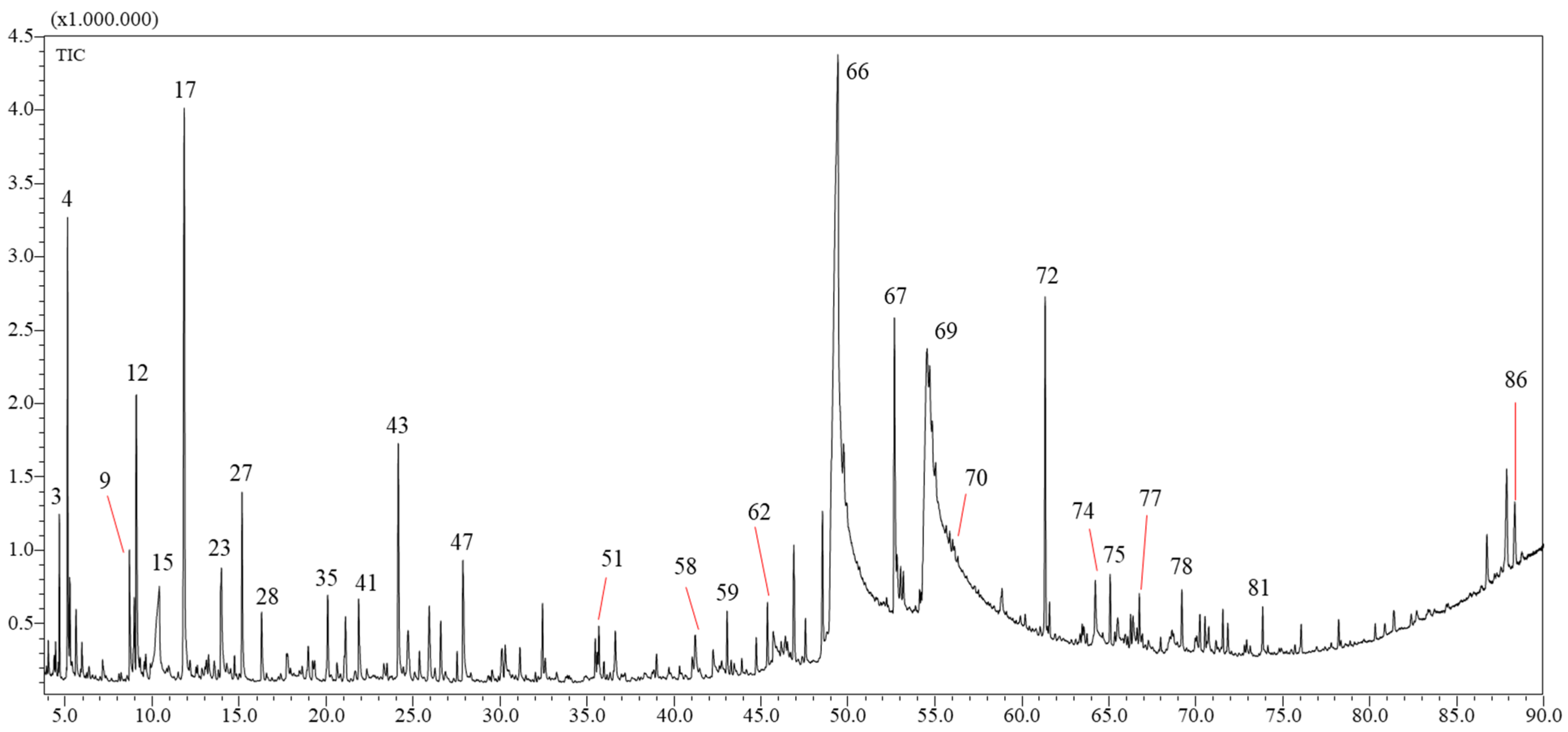
2.1. Yields and Chemical Composition of the VFs
2.2. Antifungal Activity
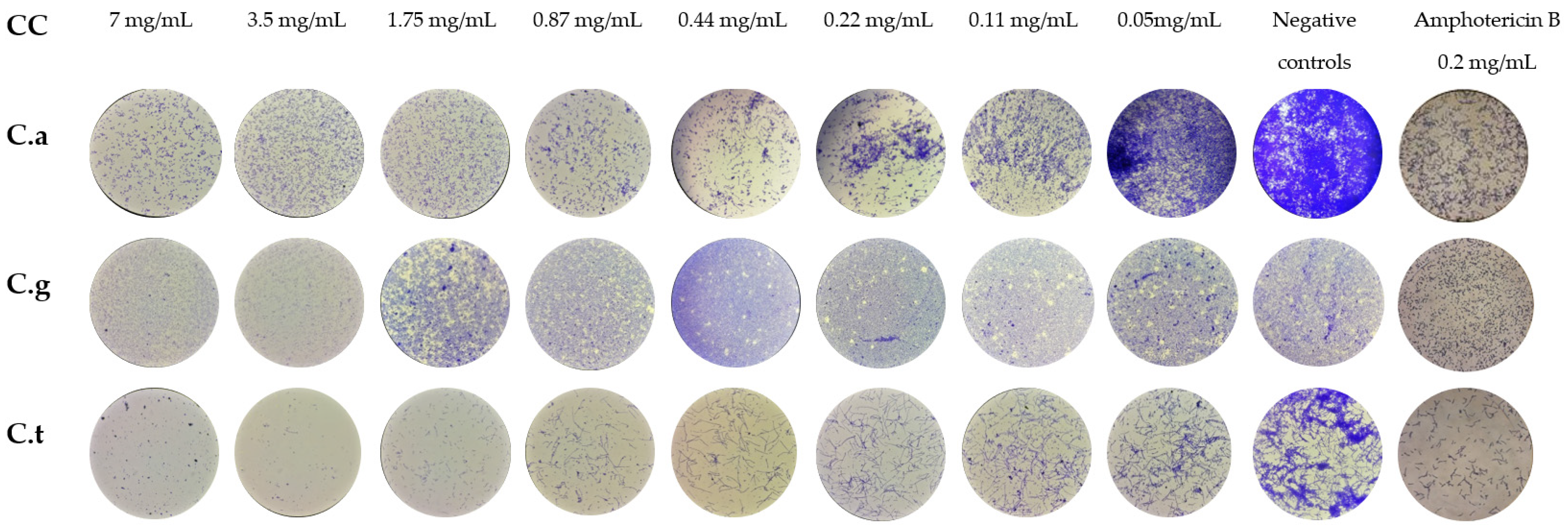
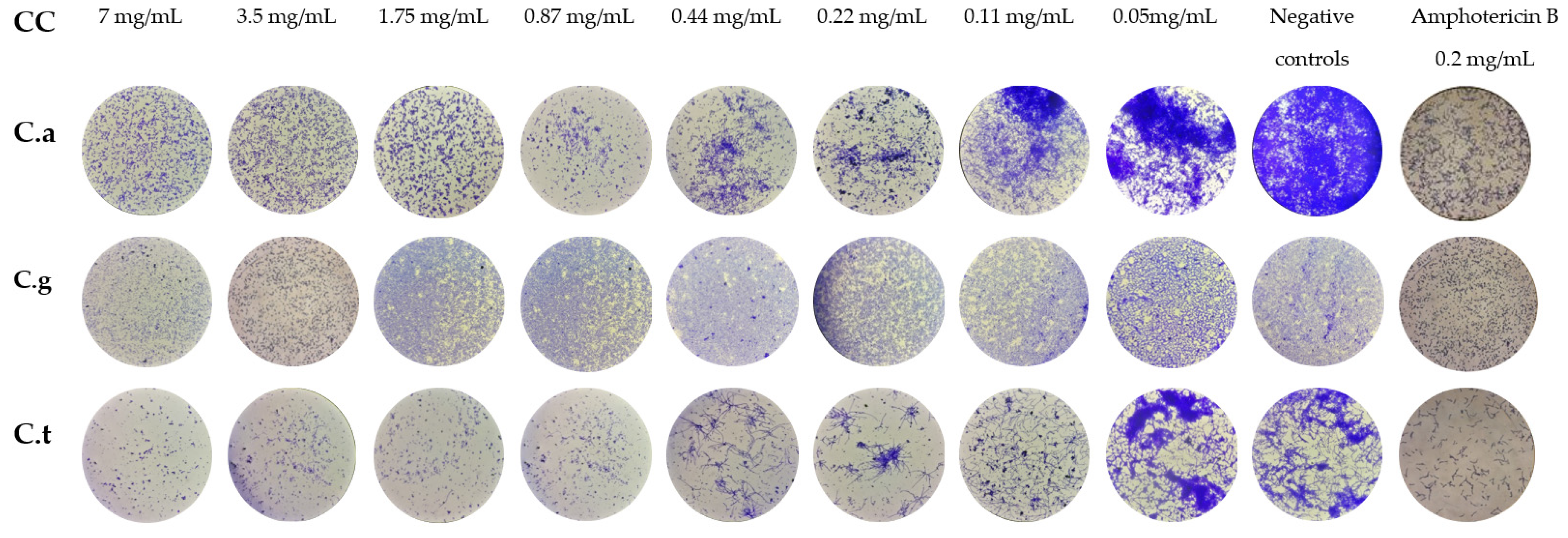
2.3. Anti-Biofilm Activity
2.3.1. Inhibition of Biofilm Formation (IBF)
2.3.2. Biofilm Sustained Inhibition (BSI)
3. Materials and Methods
3.1. Plant Material
3.2. Chemicals and Reagents
3.3. Preparation of VF from Roots of C. althaeoides L.
3.4. Gas Chromatography–Mass Spectrometry (GC–MS) Analysis
3.5. Antifungal Activity
3.5.1. Fungal Strains
3.5.2. Determination of the Minimum Inhibitory Concentrations (MICs)
3.5.3. Minimum fungicidal concentrations (MFCs)
3.6. Inhibition of Biofilm Formation (IBF)
3.7. Biofilm Sustained Inhibition (BSI)
3.8. Statistical Analysis
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Sample Availability
References
- Teodoro, G.R.; Ellepola, K.; Seneviratne, C.J.; Koga-Ito, C.Y. Potential Use of Phenolic Acids as Anti-Candida Agents: A Review. Front. Microbiol. 2015, 6, 1420. [Google Scholar] [CrossRef]
- El Hamdani, N.; Filali-Ansari, N.; Fdil, R.; El Abbouyi, A.; Khyari, S. Antifungal activity of the alkaloids extracts from aerial parts of Retama monosperma. Res. J. Pharm. Biol. Chem. Sci. 2016, 7, 965–971. [Google Scholar]
- Matin, M.; Bhattacharjee, S.; Hoque, M.; Ahamed, F. Antibacterial activity of some medicinal plants against carbapenem-resistant Acinetobacter baumannii isolated from patients. Eur. J. Pharm. Med. Res. 2019, 6, 111–116. [Google Scholar]
- Matin, P.; Rahman, M.; Huda, D.; Bakri, M.K.; Uddin, J.; Yurkin, Y.; Burkov, A.; Kuok, K.; Matin, M. Application of Synthetic Acyl Glucopyranosides for White-rot and Brown-rot Fungal Decay Resistance in Aspen and Pine Wood. Bioresources 2022, 17, 3025–3041. [Google Scholar] [CrossRef]
- Hamedi, S.; Shojaosadati, S.A.; Mohammadi, A. Evaluation of the catalytic, antibacterial and anti-biofilm activities of the Convolvulus arvensis extract functionalized silver nanoparticles. J. Photochem. Photobiol. B Biol. 2017, 167, 36–44. [Google Scholar] [CrossRef] [PubMed]
- Semwal, D.K.; Kumar, A.; Semwal, R.B.; Andola, H.C. Chapter 3.2.19—Convolvulus prostratus. In Naturally Occurring Chemicals Against Alzheimer’s Disease; Belwal, T., Nabavi, S.M., Nabavi, S.F., Dehpour, A.R., Shirooie, S., Eds.; Academic Press: Cambridge, MA, USA, 2021; pp. 409–424. [Google Scholar] [CrossRef]
- Al-Rifai, A.; Aqel, A.; Al-Warhi, T.; Wabaidur, S.M.; Al-Othman, Z.A.; Badjah-Hadj-Ahmed, A.Y. Antibacterial, Antioxidant Activity of Ethanolic Plant Extracts of Some Convolvulus Species and Their DART-ToF-MS Profiling. Evid. Based Complement. Altern. Med. 2017, 2017, 5694305. [Google Scholar] [CrossRef] [PubMed]
- Gupta, G.L.; Fernandes, J. Protective effect of Convolvulus pluricaulis against neuroinflammation associated depressive behavior induced by chronic unpredictable mild stress in rat. Biomed. Pharmacother. 2019, 109, 1698–1708. [Google Scholar] [CrossRef]
- Azman, N.; Gallego, M.; Juliá, L.; Fajari, L.; Almajano, M. The Effect of Convolvulus arvensis Dried Extract as a Potential Antioxidant in Food Models. Antioxidants 2015, 4, 170–184. [Google Scholar] [CrossRef] [PubMed]
- Hrichi, S.; Chaabane-Banaoues, R.; Giuffrida, D.; Mangraviti, D.; Oulad El Majdoub, Y.; Rigano, F.; Mondello, L.; Babba, H.; Mighri, Z.; Cacciola, F. Effect of seasonal variation on the chemical composition and antioxidant and antifungal activities of Convolvulus althaeoides L. leaf extracts. Arab. J. Chem. 2020, 13, 5651–5668. [Google Scholar] [CrossRef]
- Hassine, M.; Zardi-Berguaoui, A.; Znati, M.; Flamini, G.; Ben Jannet, H.; Hamza, M.A. Chemical composition, antibacterial and cytotoxic activities of the essential oil from the flowers of Tunisian Convolvulus althaeoides L. Nat. Prod. Res. 2014, 28, 769–775. [Google Scholar] [CrossRef]
- Cabrita, L. A Novel Acylated Anthocyanin with a Linear Trisaccharide from Flowers of Convolvulus althaeoides. Nat. Prod. Commun. 2015, 10, 1965–1968. [Google Scholar] [CrossRef] [PubMed]
- Granato, D.; Shahidi, F.; Wrolstad, R.; Kilmartin, P.; Melton, L.D.; Hidalgo, F.J.; Miyashita, K.; van Camp, J.; Alasalvar, C.; Ismail, A.B.; et al. Antioxidant activity, total phenolics and flavonoids contents: Should we ban in vitro screening methods? Food Chem. 2018, 264, 471–475. [Google Scholar] [CrossRef]
- Pfaller, M.A.; Diekema, D.J. Epidemiology of Invasive Candidiasis: A Persistent Public Health Problem. Clin. Microbiol. Rev. 2007, 20, 133–163. [Google Scholar] [CrossRef]
- Bhattacharya, S.; Sae-Tia, S.; Fries, B.C. Candidiasis and Mechanisms of Antifungal Resistance. Antibiotics 2020, 9, 312. [Google Scholar] [CrossRef]
- Deorukhkar, S.C.; Saini, S.; Mathew, S. Virulence Factors Contributing to Pathogenicity of Candida tropicalis and Its Antifungal Susceptibility Profile. Int. J. Microbiol. 2014, 2014, 456878. [Google Scholar] [CrossRef]
- Ramage, G.; Saville, S.P.; Thomas, D.P.; López-Ribot, J.L. Candida Biofilms: An Update. Eukaryot. Cell 2005, 4, 633–638. [Google Scholar] [CrossRef]
- Pinto, E.; Salgueiro, L.R.; Cavaleiro, C.; Palmeira, A.; Gonçalves, M.J. In vitro susceptibility of some species of yeasts and filamentous fungi to essential oils of Salvia officinalis. Ind. Crops Prod. 2007, 26, 135–141. [Google Scholar] [CrossRef]
- Achimón, F.; Brito, V.D.; Pizzolitto, R.P.; Ramirez Sanchez, A.; Gómez, E.A.; Zygadlo, J.A. Chemical composition and antifungal properties of commercial essential oils against the maize phytopathogenic fungus Fusarium verticillioides. Rev. Argent. Microbiol. 2021, 53, 292–303. [Google Scholar] [CrossRef]
- Karthikeyan, M.; Subramanian, P.; Ramalingam, S. Phytochemical analysis in economically important Ficus Benghalensis L. and Ficus Krishnae C.DC. using GC-MS. Int. J. Pharma. Bio. Sci. 2019, 10, 5–13. [Google Scholar] [CrossRef]
- Zyakun, A.; Nii-Annang, S.; Franke, G.; Fischer, T.; Buegger, F.; Dilly, O. Microbial Activity and 13C/12C Ratio as Evidence of N-Hexadecane and N-Hexadecanoic Acid Biodegradation in Agricultural and Forest Soils. Geomicrobiol. J. 2012, 29, 570–584. [Google Scholar] [CrossRef]
- Azadfar, M.; Gao, A.H.; Bule, M.V.; Chen, S. Structural characterization of lignin: A potential source of antioxidants guaiacol and 4-vinylguaiacol. Int. J. Biol. Macromol. 2015, 75, 58–66. [Google Scholar] [CrossRef] [PubMed]
- Behairy, A.; Abd El-Rahman, G.I.; Aly, S.S.H.; Fahmy, E.M.; Abd-Elhakim, Y.M. Di(2-ethylhexyl) adipate plasticizer triggers hepatic, brain, and cardiac injury in rats: Mitigating effect of Peganum harmala oil. Ecotoxicol. Environ. Saf. 2021, 208, 111620. [Google Scholar] [CrossRef] [PubMed]
- Al-Akeel, R.A.; El-Kersh, T.A.; Al-Sheikh, Y.A.; Al-Ahmadey, Z.Z. Heparin-benzyl alcohol enhancement of biofilms formation and antifungal susceptibility of vaginal Candida species isolated from pregnant and nonpregnant Saudi women. Bioinformation 2013, 9, 357–362. [Google Scholar] [CrossRef] [PubMed]
- Qiu, M.; Wang, Y.; Sun, L.; Deng, Q.; Zhao, J. Fatty Acids and Oxylipins as Antifungal and Anti-Mycotoxin Agents in Food: A Review. Toxins 2021, 13, 852. [Google Scholar] [CrossRef] [PubMed]
- Ma, K.; Kou, J.; Rahman, M.K.U.; Du, W.; Liang, X.; Wu, F.; Li, W.; Pan, K. Palmitic acid mediated change of rhizosphere and alleviation of Fusarium wilt disease in watermelon. Saudi J. Biol. Sci. 2021, 28, 3616–3623. [Google Scholar] [CrossRef]
- Ahsan, T.; Chen, J.; Zhao, X.; Irfan, M.; Wu, Y. Extraction and identification of bioactive compounds (eicosane and dibutyl phthalate) produced by Streptomyces strain KX852460 for the biological control of Rhizoctonia solani AG-3 strain KX852461 to control target spot disease in tobacco leaf. AMB Express 2017, 7, 54. [Google Scholar] [CrossRef]
- Morcia, C.; Malnati, M.; Terzi, V. In vitro antifungal activity of terpinen-4-ol, eugenol, carvone, 1,8-cineole (eucalyptol) and thymol against mycotoxigenic plant pathogens. Food Addit. Contam. Part A Chem. Anal. Control Expo. Risk Assess 2012, 29, 415–422. [Google Scholar] [CrossRef][Green Version]
- Li, Q.; Zhu, X. Vanillin and its derivatives, potential promising antifungal agents, inhibit Aspergillus flavus spores via destroying the integrity of cell membrane rather than cell wall. Grain Oil Sci. Technol. 2021, 4, 54–61. [Google Scholar] [CrossRef]
- Gao, T.; Zhang, Y.; Shi, J.; Mohamed, S.R.; Xu, J.; Liu, X. The Antioxidant Guaiacol Exerts Fungicidal Activity Against Fungal Growth and Deoxynivalenol Production in Fusarium graminearum. Front. Microbiol. 2021, 12, 762844. [Google Scholar] [CrossRef]
- Anum Mengal, S.; Baqi, A. 19. Antibacterial and antifungal activities of extracts of Convolvulus leiocalycinus and Haloxylon griffithii of Balochistan, Pakistan. Pure Appl. Biol. 2019, 8, 2286–2294. [Google Scholar]
- Amin, H.; Dhiman, K.; Sharma, R.; Vyas, M.; Prajapati, P. Shankhapushpi (Convolvulus pluricaulis Choisy): Validation of the Ayurvedic therapeutic claims through contemporary studies. Int. J. Green Pharm. 2014, 8, 193. [Google Scholar] [CrossRef] [PubMed]
- Essid, R.; Gharbi, D.; Abid, G.; Karkouch, I.; Hamouda, T.B.; Fares, N.; Trabelsi, D.; Mhadhbi, H.; Elkahoui, S.; Limam, F.; et al. Combined effect of Thymus capitatus and Cinnamomum verum essential oils with conventional drugs against Candida albicans biofilm formation and elucidation of the molecular mechanism of action. Ind. Crops Prod. 2019, 140, 111720. [Google Scholar] [CrossRef]
- Gulati, M.; Nobile, C.J. Candida albicans biofilms: Development, regulation, and molecular mechanisms. Microbes Infect. 2016, 18, 310–321. [Google Scholar] [CrossRef] [PubMed]
- Somma, A.D.; Moretta, A.; Canè, C.; Cirillo, A.; Duilio, A. Inhibition of Bacterial Biofilm Formation; IntechOpen: London, UK, 2020. [Google Scholar] [CrossRef]
- Tits, J.; Cammue, B.P.A.; Thevissen, K. Combination Therapy to Treat Fungal Biofilm-Based Infections. Int. J. Mol. Sci. 2020, 21, 8873. [Google Scholar] [CrossRef] [PubMed]
- Kim, Y.-G.; Lee, J.-H.; Lee, J. Antibiofilm activities of fatty acids including myristoleic acid against Cutibacterium acnes via reduced cell hydrophobicity. Phytomedicine 2021, 91, 153710. [Google Scholar] [CrossRef]
- Raut, J.S.; Shinde, R.B.; Chauhan, N.M.; Karuppayil, S.M. Terpenoids of plant origin inhibit morphogenesis, adhesion, and biofilm formation by Candida albicans. Biofouling 2013, 29, 87–96. [Google Scholar] [CrossRef]
- Caesar, L.K.; Cech, N.B. Synergy and antagonism in natural product extracts: When 1 + 1 does not equal 2. Nat. Prod. Rep. 2019, 36, 869–888. [Google Scholar] [CrossRef]
- Hrichi, S.; Chaabane-Banaoues, R.; Bayar, S.; Flamini, G.; Oulad El Majdoub, Y.; Mangraviti, D.; Mondello, L.; El Mzoughi, R.; Babba, H.; Mighri, Z.; et al. Botanical and Genetic Identification Followed by Investigation of Chemical Composition and Biological Activities on the Scabiosa atropurpurea L. Stem from Tunisian Flora. Molecules 2020, 25, 5032. [Google Scholar] [CrossRef]
- Li, H.; Zhang, M.; Addo, K.A.; Yu, Y.; Xiao, X. Action mode of cuminaldehyde against Staphylococcus aureus and its application in sauced beef. LWT 2022, 155, 112924. [Google Scholar] [CrossRef]
- Yano, T.; Miyahara, Y.; Morii, N.; Okano, T.; Kubota, H. Pentanol and Benzyl Alcohol Attack Bacterial Surface Structures Differently. Appl. Environ. Microbiol. 2016, 82, 402–408. [Google Scholar] [CrossRef]
- Guimarães, A.; Venâncio, A. The Potential of Fatty Acids and Their Derivatives as Antifungal Agents: A Review. Toxins 2022, 14, 188. [Google Scholar] [CrossRef] [PubMed]
- Prasath, K.G.; Tharani, H.; Kumar, M.S.; Pandian, S.K. Palmitic Acid Inhibits the Virulence Factors of Candida tropicalis: Biofilms, Cell Surface Hydrophobicity, Ergosterol Biosynthesis, and Enzymatic Activity. Front. Microbiol. 2020, 11, 864. [Google Scholar] [CrossRef] [PubMed]
- Aswathanarayan, J.B.; Vittal, R.R. Inhibition of biofilm formation and quorum sensing mediated phenotypes by berberine in Pseudomonas aeruginosa and Salmonella typhimurium. RSC Adv. 2018, 8, 36133–36141. [Google Scholar] [CrossRef] [PubMed]
- Nair, R.B.; Lennartsson, P.R.; Taherzadeh, M.J. Mycelial pellet formation by edible ascomycete filamentous fungi, Neurospora intermedia. AMB Express 2016, 6, 31. [Google Scholar] [CrossRef] [PubMed]
- Lohse, M.B.; Gulati, M.; Craik, C.S.; Johnson, A.D.; Nobile, C.J. Combination of Antifungal Drugs and Protease Inhibitors Prevent Candida albicans Biofilm Formation and Disrupt Mature Biofilms. Front. Microbiol. 2020, 11, 1027. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| N. Peak | Compound | Area% | LRI (Lib) | LRI (Exp) | Library | Compound Class | Formula |
|---|---|---|---|---|---|---|---|
| 15 | Myrtenol | 0.93 | 1202 | 1198 | FFNSC 4.0 | Oxygenated monoterpene | C10H16O |
| 17 | Cuminaldehyde | 0.79 | 1243 | 1245 | FFNSC 4.0 | Oxygenated monoterpene | C10H12O |
| 18 | Carvone | 1.77 | 1246 | 1246 | FFNSC 4.0 | Oxygenated monoterpene | C10H14O |
| 20 | 4-Vinylguaiacol | 12.2 | 1309 | 1315 | FFNSC 4.0 | Alcohol | C9H10O2 |
| 21 | Methyl 4-formylbenzoate | 1.03 | - | 1370 | W11N17 | Ester | C9H8O3 |
| 24 | Tetradec-1-ene | 0.9 | 1379 | 1391 | FFNSC 4.0 | Alkene | C14H28 |
| 26 | α-Methoxynaphthalene | 1.52 | 1450 | 1453 | FFNSC 4.0 | Alkene | C11H10O |
| 28 | 2,4-bis(1,1-dimethylethyl)-phenol | 1.47 | - | 1509 | W11N17 | Alcohol | C14H22O |
| 31 | n-Hexadecene | 1.1 | 1593 | 1592 | FFNSC 4.0 | Alkene | C16H32 |
| 39 | Octadec-1-ene | 1.03 | 1793 | 1792 | FFNSC 4.0 | Alkene | C18H36 |
| 42 | diisobutyl phthalate | 1.55 | 1858 | 1860 | FFNSC 4.0 | Ester | C16H22O4 |
| 48 | n-Hexadecanoic acid | 29.77 | 1977 | 1983 | FFNSC 4.0 | Fatty acid | C16H32O2 |
| 49 | n-Eicosene | 3.98 | 1994 | 1993 | FFNSC 4.0 | Alkene | C20H40 |
| 50 | n-Eicosane | 2.11 | 2000 | 1999 | FFNSC 4.0 | Alkane | C20H42 |
| 51 | n-Octadecanol | 2.89 | 2081 | 2087 | FFNSC 4.0 | Alcohol | C18H38O |
| 53 | Ethyl linoleate | 1.75 | 2164 | 2160 | FFNSC 4.0 | Fatty acid | C20H36O2 |
| 54 | Ethyl linolenate | 1.47 | 2165 | 2166 | FFNSC 4.0 | Fatty acid | C20H34O2 |
| 56 | n-Tricosane | 0.81 | 2300 | 2299 | FFNSC 4.0 | Alkane | C23H48 |
| 57 | Bis(2-ethylhexyl)-adipate | 9.69 | 2392 | 2391 | FFNSC 4.0 | Ester | C22H42O4 |
| 58 | n-Tetracosane | 1.23 | 2400 | 2399 | FFNSC 4.0 | Alkane | C24H50 |
| 59 | n-Pentacosane | 1.59 | 2500 | 2499 | FFNSC 4.0 | Alkane | C25H52 |
| 62 | n-Hexacosane | 1.77 | 2600 | 2598 | FFNSC 4.0 | Alkane | C26H54 |
| 64 | n-Heptacosane | 1.64 | 2700 | 2699 | FFNSC 4.0 | Alkane | C27H56 |
| 66 | n-Octacosane | 1.37 | 2800 | 2798 | FFNSC 4.0 | Alkane | C28H58 |
| 68 | n-Nonacosane | 1.12 | 900 | 2898 | FFNSC 4.0 | Alkane | C29H60 |
| 73 | 3,5-bis(1,1-dimethylthyl)-4-hydroxy-, octadecyl ester | 2.71 | - | 3596 | W11N17 | Carboxylic acid | C35H62O3 |
| N. Peak | Compound | Area% | LRI (Lib) | LRI (Exp) | Library | Compound Class | Formula |
|---|---|---|---|---|---|---|---|
| 3 | Hexan-2-ol | 0.91 | 802 | 812 | FFNSC 4.0 | Alcohol | C6H14O |
| 4 | Furfural | 3.24 | 845 | 832 | FFNSC 4.0 | Aldehyde | C5H4O2 |
| 9 | β-Lutidine | 1.29 | 955 | 955 | FFNSC 4.0 | Pyridine | C7H9N |
| 12 | 3-ethenyl- Pyridine | 2.43 | - | 966 | W11N17 | Pyridine | C7H7N |
| 15 | n-Hexanoic acid | 1.91 | 997 | 1004 | FFNSC 4.0 | Carboxylic acid | C6H12O2 |
| 17 | Benzyl alcohol | 7.86 | 1040 | 1037 | FFNSC 4.0 | Alcohol | C7H8O |
| 23 | 2-Methoxyphenol | 1.27 | - | 1088 | W11N17 | Alcohol | C7H8O2 |
| 27 | Phenethyl alcohol | 2.07 | 1113 | 1115 | FFNSC 4.0 | Alcohol | C8H10O |
| 35 | 4-Vinylphenol | 0.98 | 1217 | 1223 | FFNSC 4.0 | Alcohol | C8H8O |
| 41 | Benzopyridine | 1 | 1259 | 1263 | FFNSC 4.0 | Pyridine | C9H7N |
| 43 | 4-Vinylguaiacol | 2.87 | 1309 | 1314 | FFNSC 4.0 | Alcohol | C7H8O2 |
| 47 | Vanillin | 1.49 | 1394 | 1399 | FFNSC 4.0 | Aldehyde | C8H8O3 |
| 66 | n-Hexadecanoic acid | 34.01 | 1977 | 1983 | FFNSC 4.0 | Fatty acid | C16H32O2 |
| 67 | n-Octadecanol | 3.53 | 2081 | 2087 | FFNSC 4.0 | Alcohol | C18H38O |
| 69 | Linoleic acid | 7.3 | 2144 | 2154 | FFNSC 4.0 | Fatty acid | C18H32O2 |
| 70 | n-Docosane | 2.43 | 2200 | 2199 | FFNSC 4.0 | Alkane | C22H46 |
| 72 | Bis(2-ethylhexyl)-adipate | 3.57 | 2392 | 2390 | FFNSC 4.0 | Ester | C22H42O4 |
| 74 | n-Pentacosane | 1.47 | 2500 | 2499 | FFNSC 4.0 | Alkane | C25H52 |
| 86 | 3,5-bis(1,1-dimethylthyl)-4-hydroxy-, octadecyl ester | 0.95 | - | 3595 | W11N17 | Carboxylic acid | C35H62O3 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hrichi, S.; Chaâbane-Banaoues, R.; Alibrando, F.; Altemimi, A.B.; Babba, O.; Majdoub, Y.O.E.; Nasri, H.; Mondello, L.; Babba, H.; Mighri, Z.; et al. Chemical Composition, Antifungal and Anti-Biofilm Activities of Volatile Fractions of Convolvulus althaeoides L. Roots from Tunisia. Molecules 2022, 27, 6834. https://doi.org/10.3390/molecules27206834
Hrichi S, Chaâbane-Banaoues R, Alibrando F, Altemimi AB, Babba O, Majdoub YOE, Nasri H, Mondello L, Babba H, Mighri Z, et al. Chemical Composition, Antifungal and Anti-Biofilm Activities of Volatile Fractions of Convolvulus althaeoides L. Roots from Tunisia. Molecules. 2022; 27(20):6834. https://doi.org/10.3390/molecules27206834
Chicago/Turabian StyleHrichi, Soukaina, Raja Chaâbane-Banaoues, Filippo Alibrando, Ammar B. Altemimi, Oussama Babba, Yassine Oulad El Majdoub, Habib Nasri, Luigi Mondello, Hamouda Babba, Zine Mighri, and et al. 2022. "Chemical Composition, Antifungal and Anti-Biofilm Activities of Volatile Fractions of Convolvulus althaeoides L. Roots from Tunisia" Molecules 27, no. 20: 6834. https://doi.org/10.3390/molecules27206834
APA StyleHrichi, S., Chaâbane-Banaoues, R., Alibrando, F., Altemimi, A. B., Babba, O., Majdoub, Y. O. E., Nasri, H., Mondello, L., Babba, H., Mighri, Z., & Cacciola, F. (2022). Chemical Composition, Antifungal and Anti-Biofilm Activities of Volatile Fractions of Convolvulus althaeoides L. Roots from Tunisia. Molecules, 27(20), 6834. https://doi.org/10.3390/molecules27206834