

Autotoxicity of Endogenous Organic Acid Stress in Two Ganoderma lucidum Cultivars

Abstract

1. Introduction
2. Results and Discussion
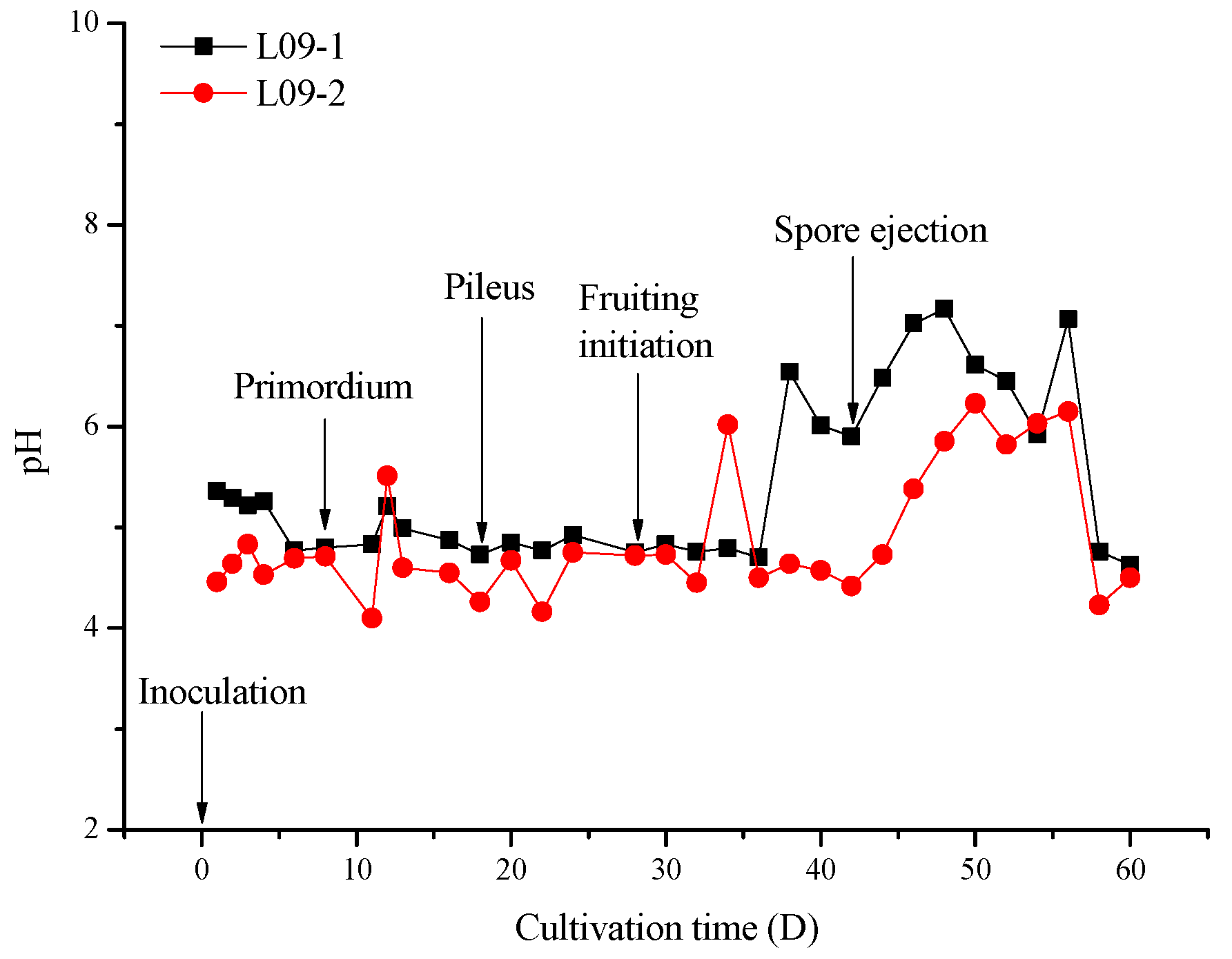
2.1. pH Variation of Germ Substrates during Cultivation
2.2. Acquisition of Mass Spectrum Data
2.3. Comparison of Organic Acids and Derivatives between L09-1 and L09-2
3. Materials and Methods
3.1. Strains and Inoculation
3.2. Sample Preparations
3.3. UPLC-MS/MS Data Acquisition
3.4. Processing and Statistical Analysis of MS Spectra Data
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Sample Availability
References
- Bhardwaj, A.; Gupta, P.; Kumar, N.; Mishra, J.; Kumar, A.; Rakhee; Misra, K. Lingzhi or Reishi medicinal mushroom, Ganoderma lucidum (Agaricomycetes), inhibits Candida biofilms: A metabolomic approach. Int. J. Med. Mushroom 2017, 19, 685–696. [Google Scholar] [CrossRef]
- Kurd-Anjaraki, S.; Ramezan, D.; Ramezani, S.; Samzadeh-Kermani, A.; Pirnia, M.; Shahi, B.Y. Potential of waste reduction of agro-biomasses through Reishi medicinal mushroom (Ganoderma lucidum) production using different substrates and techniques. Acta Ecol. Sin. 2022, 42, 90–101. [Google Scholar] [CrossRef]
- Kahveci, R.; Kahveci, F.O.; Gokce, E.C.; Gokce, A.; Kisa, U.; Sargon, M.F.; Fesli, R.; Gurer, B. Effects of Ganoderma lucidum Polysaccharides on Different Pathways Involved in the Development of Spinal Cord Ischemia Reperfusion Injury: Biochemical, Histopathologic, and Ultrastructural Analysis in a Rat Model. World Neurosurg. 2021, 150, e287–e297. [Google Scholar] [CrossRef] [PubMed]
- Soccol, C.R.; Bissoqui, L.Y.; Rodrigues, C.; Rubel, R.; Sella, S.R.B.R.; Leifa, F.; Vandenberghe, L.P.D.S.; Soccol, V.T. Pharmacological properties of biocompounds from spores of the Lingzhi or Reishi medicinal mushroom Ganoderma lucidum (Agaricomycetes): A review. Int. J. Med. Mushroom 2016, 18, 757–767. [Google Scholar] [CrossRef] [PubMed]
- Xie, C.; Yan, S.; Zhang, Z.; Gong, W.; Zhu, Z.; Zhou, Y.; Yan, L.; Hu, Z.; Ai, L.; Peng, Y. Mapping the metabolic signatures of fermentation broth, mycelium, fruiting body and spores powder from Ganoderma lucidum by untargeted metabolomics. LWT 2020, 129, 109494. [Google Scholar] [CrossRef]
- Diling, C.; Yinrui, G.; Longkai, Q.; Xiaocui, T.; Yadi, L.; Jiaxin, F.; Xiangxiang, Z.; Miao, Z.; Ou, S.; Dongdong, W.; et al. Metabolic regulation of Ganoderma lucidum extracts in high sugar and fat diet-induced obese mice by regulating the gut-brain axis. J. Funct. Foods 2020, 65, 103639. [Google Scholar] [CrossRef]
- Hapuarachchi, K.K. Current status of global Ganoderma cultivation, products, industry and market. Mycosphere 2018, 9, 1025–1052. [Google Scholar] [CrossRef]
- Bonanomi, G.; Lorito, M.; Vinale, F.; Woo, S.L. Organic amendments, beneficial microbes, and soil microbiota: Toward a unified framework for disease suppression. Annu. Rev. Phytopathol. 2018, 56, 1–20. [Google Scholar] [CrossRef]
- Li, S.; Wu, F. Diversity and co-occurrence patterns of soil bacterial and fungal communities in seven intercropping systems. Front. Microbiol. 2018, 9, 1521. [Google Scholar] [CrossRef] [PubMed]
- Wolińska, A.; Kuźniar, A.; Zielenkiewicz, U.; Banach, A.; Błaszczyk, M. Indicators of arable soils fatigue—Bacterial families and genera: A metagenomic approach. Ecol. Indic. 2018, 93, 490–500. [Google Scholar] [CrossRef]
- Shaoli, C.; Baoli, Z.; Shanshan, L.; Xia, L.; Xueling, Y. Accumulation of cinnamic acid and vanillin in eggplant root exudates and the relationship with continuous cropping obstacle. AFR J. Biotechnol. 2011, 10, 2659–2665. [Google Scholar] [CrossRef]
- Cai, M.; Liang, X.; Liu, Y.; Hu, H.; Xie, Y.; Chen, S.; Gao, X.; Li, X.; Xiao, C.; Chen, D.; et al. Transcriptional dynamics of genes purportedly involved in the control of meiosis, carbohydrate, and secondary metabolism during sporulation in Ganoderma lucidum. Genes 2021, 12, 504. [Google Scholar] [CrossRef] [PubMed]
- Marcos-Arias, C.; Eraso, E.; Madariaga, L.; Quindos, G. In vitro activities of natural products against oral Candida isolates from denture wearers. BMC Complement Altern. Med. 2011, 11, 119. [Google Scholar] [CrossRef]
- Morath, S.U.; Hung, R.; Bennett, J.W. Fungal volatile organic compounds: A review with emphasis on their biotechnological potential. Fungal Biol. Rev. 2012, 26, 73–83. [Google Scholar] [CrossRef]
- Yao, S.; Cao, Q.; Xie, J.; Deng, L.; Zeng, K. Alteration of sugar and organic acid metabolism in postharvest granulation of Ponkan fruit revealed by transcriptome profiling. Postharvest Biol. Technol. 2018, 139, 2–11. [Google Scholar] [CrossRef]
- Luo, J.; Hu, K.; Qu, F.; Ni, D.; Zhang, H.; Liu, S.; Chen, Y. Metabolomics analysis reveals major differential metabolites and metabolic alterations in tea plant leaves (Camellia sinensis L.) under different fluorine conditions. J. Plant Growth Regul. 2020, 40, 798–810. [Google Scholar] [CrossRef]
- Krzesłowska, M. The cell wall in plant cell response to trace metals: Polysaccharide remodeling and its role in defense strategy. Acta Physiol. Plant. 2010, 33, 35–51. [Google Scholar] [CrossRef]
- Wang, B.; Lv, Y.; Li, X.; Lin, Y.; Deng, H.; Pan, L. Profiling of secondary metabolite gene clusters regulated by LaeA in Aspergillus niger FGSC A1279 based on genome sequencing and transcriptome analysis. Res. Microbiol. 2018, 169, 67–77. [Google Scholar] [CrossRef]
- Ahmad, M.F. Ganoderma lucidum: Persuasive biologically active constituents and their health endorsement. Biomed. Pharmacother. 2018, 107, 507–519. [Google Scholar] [CrossRef]
- Sharma, C.; Bhardwaj, N.; Sharma, A.; Tuli, H.S.; Batra, P.; Beniwal, V.; Gupta, G.K.; Sharma, A.K. Bioactive metabolites of Ganoderma lucidum: Factors, mechanism and broad spectrum therapeutic potential. J. Herb. Med 2019, 17–18, 100268. [Google Scholar] [CrossRef]
- Wu, Z.; Tu, M.; Yang, X.; Xu, J.; Yu, Z. Effect of cutting and storage temperature on sucrose and organic acids metabolism in postharvest melon fruit. Postharvest Biol. Technol. 2020, 161, 111081. [Google Scholar] [CrossRef]
- Zhang, J.; Zeng, B.; Mao, Y.; Kong, X.; Wang, X.; Yang, Y.; Zhang, J.; Xu, J.; Rengel, Z.; Chen, Q. Melatonin alleviates aluminium toxicity through modulating antioxidative enzymes and enhancing organic acid anion exudation in soybean. Funct. Plant Biol. 2017, 44, 961–968. [Google Scholar] [CrossRef] [PubMed]
- Samanta, S.; Singh, A.; Banerjee, A.; Roychoudhury, A. Exogenous supplementation of melatonin alters representative organic acids and enzymes of respiratory cycle as well as sugar metabolism during arsenic stress in two contrasting indica rice cultivars. J. Biotechnol. 2020, 324, 220–232. [Google Scholar] [CrossRef]
- Zhang, N.; Zhang, H.J.; Sun, Q.Q.; Cao, Y.Y.; Li, X.; Zhao, B.; Wu, P.; Guo, Y.D. Proteomic analysis reveals a role of melatonin in promoting cucumber seed germination under high salinity by regulating energy production. Sci. Rep. 2017, 7, 503. [Google Scholar] [CrossRef]
- Wang, J.; Tian, T.; Wang, H.; Cui, J.; Shi, X.; Song, J.; Li, T.; Li, W.; Zhong, M.; Zhang, W. Chitosan-coated compound fertilizer application and crop rotation alleviate continuous cotton cropping obstacles by modulating root exudates. Rhizosphere 2022, 23, 100581. [Google Scholar] [CrossRef]
- Wang, H.-W.; Zhu, Y.-X.; Xu, M.; Cai, X.-Y.; Tian, F. Co-application of spent mushroom substrate and PGPR alleviates tomato continuous cropping obstacle by regulating soil microbial properties. Rhizosphere 2022, 23, 100563. [Google Scholar] [CrossRef]
- Atila, F. Comparative study on the mycelial growth and yield of Ganoderma lucidum (Curt.: Fr.) Karst. on different lignocellulosic wastes. Acta Ecol. Sin. 2020, 40, 153–157. [Google Scholar] [CrossRef]
- Royse, D.J.; Sanchez, J.E. Ground wheat straw as a substitute for portions of oak wood chips used in shiitake (Lentinula edodes) substrate formulae. Bioresour. Technol. 2007, 98, 2137–2141. [Google Scholar] [CrossRef]
- Harith, N.; Abdullah, N.; Sabaratnam, V. Cultivation of Flammulina velutipes mushroom using various agro-residues as a fruiting substrate. Pesqui. Agropecu. Bras. 2014, 49, 181–188. [Google Scholar] [CrossRef]
- Zhang, Y.X.; Yang, X.; Zou, P.; Du, P.F.; Wang, J.; Jin, F.; Jin, M.J.; She, Y.X. Nonylphenol Toxicity Evaluation and Discovery of Biomarkers in Rat Urine by a Metabolomics Strategy through HPLC-QTOF-MS. Int. J. Environ. Res. Public Health 2016, 13, 501. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| No. | Differential Compounds | Content Change |
|---|---|---|
| 1 | 5-hydroxy-2-oxo-4-ureido-2,5-dihydro-1H-imidazole-5-carboxylic acid | UP |
| 2 | malonic acid | UP |
| 3 | 2-(2-Carboxyethyl)-5-hydroxyphenyl hexopyranosiduronic acid | UP |
| 4 | 3-C-Carboxy-2-deoxy-4-O-[(2E)-3-(4-hydroxyphenyl)-2-propenoyl]pentaric acid | UP |
| 5 | 2-Deoxy-4-O-[(2E)-3-(4-hydroxyphenyl)-2-propenoyl]pentaric acid | UP |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zou, P.; Guo, Y.; Ding, S.; Song, Z.; Cui, H.; Zhang, Y.; Zhang, Z.; Chen, X. Autotoxicity of Endogenous Organic Acid Stress in Two Ganoderma lucidum Cultivars. Molecules 2022, 27, 6734. https://doi.org/10.3390/molecules27196734
Zou P, Guo Y, Ding S, Song Z, Cui H, Zhang Y, Zhang Z, Chen X. Autotoxicity of Endogenous Organic Acid Stress in Two Ganoderma lucidum Cultivars. Molecules. 2022; 27(19):6734. https://doi.org/10.3390/molecules27196734
Chicago/Turabian StyleZou, Pan, Yongze Guo, Shu Ding, Zhaowei Song, Hanyuan Cui, Yue Zhang, Zhijun Zhang, and Xiaoming Chen. 2022. "Autotoxicity of Endogenous Organic Acid Stress in Two Ganoderma lucidum Cultivars" Molecules 27, no. 19: 6734. https://doi.org/10.3390/molecules27196734
APA StyleZou, P., Guo, Y., Ding, S., Song, Z., Cui, H., Zhang, Y., Zhang, Z., & Chen, X. (2022). Autotoxicity of Endogenous Organic Acid Stress in Two Ganoderma lucidum Cultivars. Molecules, 27(19), 6734. https://doi.org/10.3390/molecules27196734