
Polydatin: Pharmacological Mechanisms, Therapeutic Targets, Biological Activities, and Health Benefits



Abstract
1. Introduction
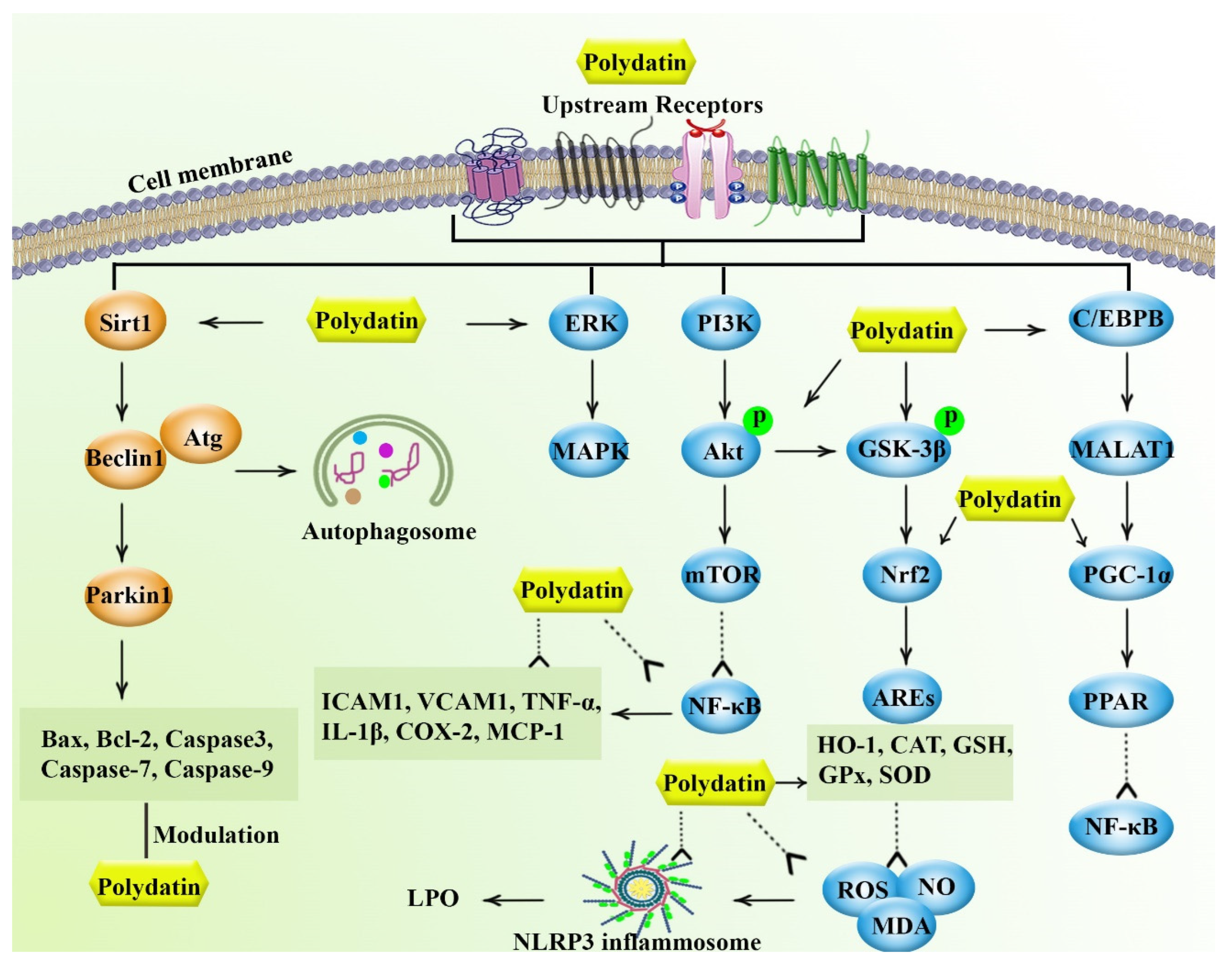
2. Pharmacological Mechanisms of Polydatin
2.1. Anti-Inflammatory Effects
2.2. Antioxidant Effects
2.3. Anti-Apoptotic Effects
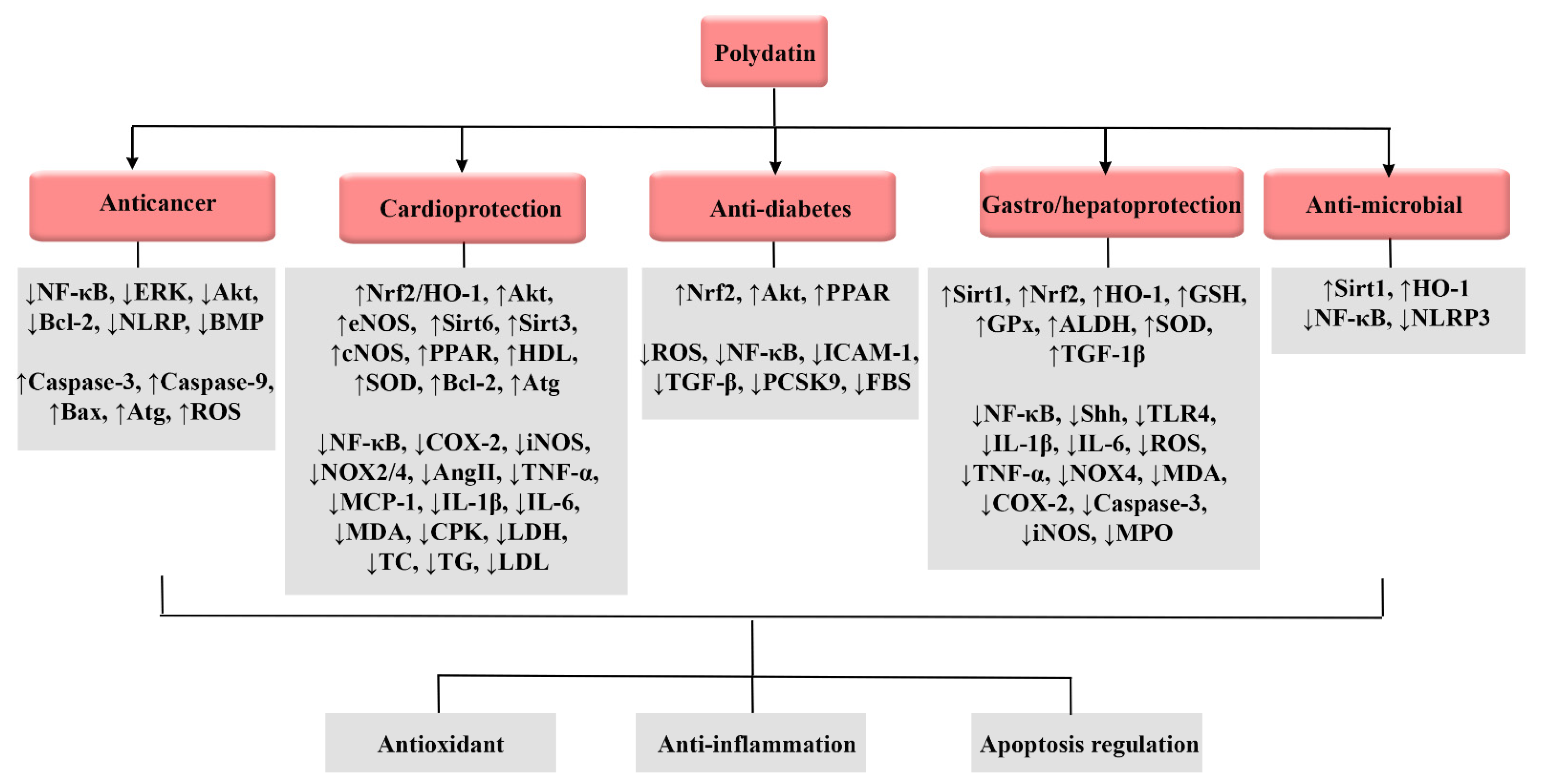
3. Biological Activities of Polydatin
3.1. Anticancer Effects
3.2. Cardioprotective Effects
3.3. Anti-Diabetic Effects
3.4. Effects on Gastrointestinal Tract
3.5. Anti-Microbial Effects
3.6. Hepatoprotective Effects
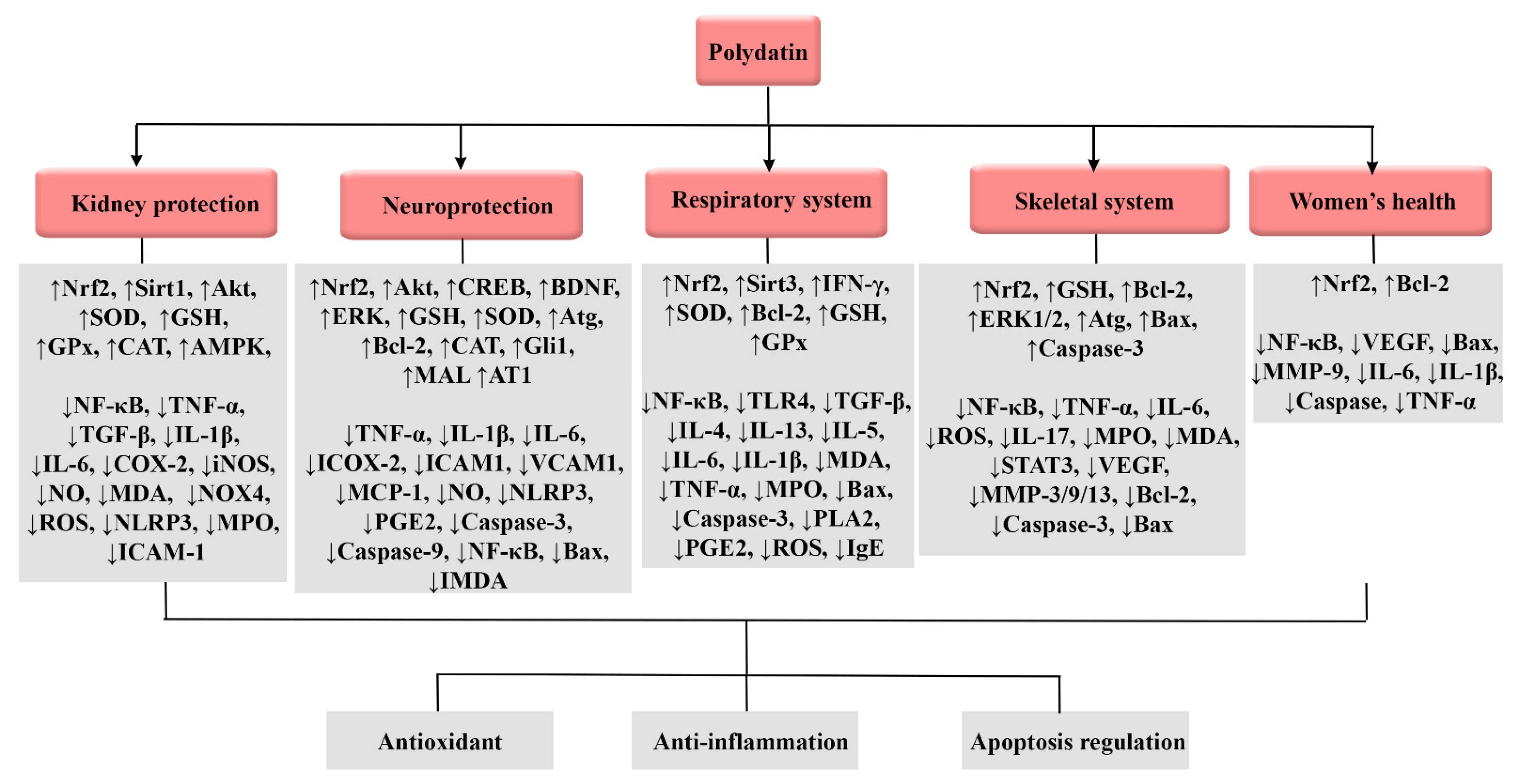
3.7. Neuroprotective Effects
3.8. Effects on Renal System
3.9. Effects on Respiratory System
3.10. Effects on Rheumatoid Diseases
3.11. Effects on the Skeletal System
3.12. Effects on Women’s Health
3.13. Miscellaneous Effects
4. Clinical Trials on Polydatin
5. Novel Delivery Systems of Polydatin
6. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Mele, L.; Paino, F.; Papaccio, F.; Regad, T.; Boocock, D.; Stiuso, P.; Lombardi, A.; Liccardo, D.; Aquino, G.; Barbieri, A. A new inhibitor of glucose-6-phosphate dehydrogenase blocks pentose phosphate pathway and suppresses malignant proliferation and metastasis in vivo. Cell Death Dis. 2018, 9, 572. [Google Scholar] [CrossRef] [PubMed]
- Zhang, M.; Zhao, Z.; Shen, M.; Zhang, Y.; Duan, J.; Guo, Y.; Zhang, D.; Hu, J.; Lin, J.; Man, W. Polydatin protects cardiomyocytes against myocardial infarction injury by activating Sirt3. Biochim. Biophys. Acta (BBA)-Mol. Basis Dis. 2017, 1863, 1962–1972. [Google Scholar] [CrossRef] [PubMed]
- Şöhretoğlu, D.; Baran, M.Y.; Arroo, R.; Kuruüzüm-Uz, A. Recent advances in chemistry, therapeutic properties and sources of polydatin. Phytochem. Rev. 2018, 17, 973–1005. [Google Scholar] [CrossRef]
- Mei, X.; Wang, Y.; Li, J.; Liu, Z.; Lang, S.; Ouyang, W.; Zhang, J. Comprehensive metabolism study of polydatin in rat plasma and urine using ultra-high performance liquid chromatography coupled with high-resolution mass spectrometry. J. Chromatogr. B 2019, 1117, 22–35. [Google Scholar] [CrossRef]
- San Tang, K.; Tan, J.S. The protective mechanisms of polydatin in cerebral ischemia. Eur. J. Pharmacol. 2019, 842, 133–138. [Google Scholar] [CrossRef]
- Peng, W.; Qin, R.; Li, X.; Zhou, H. Botany, phytochemistry, pharmacology, and potential application of Polygonum cuspidatum Sieb. et Zucc.: A review. J. Ethnopharmacol. 2013, 148, 729–745. [Google Scholar] [CrossRef]
- Negrea, B.-M.; Stoilov-Linu, V.; Pop, C.-E.; Deák, G.; Crăciun, N.; Făgăraș, M.M. Expansion of the Invasive Plant Species Reynoutria japonica Houtt in the Upper Bistrița Mountain River Basin with a Calculus on the Productive Potential of a Mountain Meadow. Sustainability 2022, 14, 5737. [Google Scholar] [CrossRef]
- Neag, M.A.; Mocan, A.; Echeverría, J.; Pop, R.M.; Bocsan, C.I.; Crişan, G.; Buzoianu, A.D. Berberine: Botanical occurrence, traditional uses, extraction methods, and relevance in cardiovascular, metabolic, hepatic, and renal disorders. Front. Pharmacol. 2018, 9, 557. [Google Scholar] [CrossRef]
- Potdar, S.; Parmar, M.S.; Ray, S.D.; Cavanaugh, J.E. Protective effects of the resveratrol analog piceid in dopaminergic SH-SY5Y cells. Arch. Toxicol. 2018, 92, 669–677. [Google Scholar] [CrossRef]
- Du, Q.-H.; Peng, C.; Zhang, H. Polydatin: A review of pharmacology and pharmacokinetics. Pharm. Biol. 2013, 51, 1347–1354. [Google Scholar] [CrossRef]
- Lanzilli, G.; Cottarelli, A.; Nicotera, G.; Guida, S.; Ravagnan, G.; Fuggetta, M.P. Anti-inflammatory effect of resveratrol and polydatin by in vitro IL-17 modulation. Inflammation 2012, 35, 240–248. [Google Scholar] [CrossRef] [PubMed]
- Muzio, L.L.; Bizzoca, M.E.; Ravagnan, G. New intriguing possibility for prevention of coronavirus pneumonitis: Natural purified polyphenols. Oral Dis. 2020, 28, 899–903. [Google Scholar] [CrossRef] [PubMed]
- Zhao, G.; Yang, L.; Zhong, W.; Hu, Y.; Tan, Y.; Ren, Z.; Ban, Q.; Yang, C.S.; Wang, Y.; Wang, Z. Polydatin, A Glycoside of Resveratrol, Is Better Than Resveratrol in Alleviating Non-alcoholic Fatty Liver Disease in Mice Fed a High-Fructose Diet. Front. Nutr. 2022, 9, 857879. [Google Scholar] [CrossRef] [PubMed]
- Bokkenheuser, V.D.; Shackleton, C.; Winter, J. Hydrolysis of dietary flavonoid glycosides by strains of intestinal Bacteroides from humans. Biochem. J. 1987, 248, 953–956. [Google Scholar] [CrossRef]
- Marín, L.; Miguélez, E.M.; Villar, C.J.; Lombó, F. Bioavailability of dietary polyphenols and gut microbiota metabolism: Antimicrobial properties. BioMed Res. Int. 2015, 2015, 905215. [Google Scholar] [CrossRef]
- Kim, M.; Lee, J.; Han, J. Deglycosylation of isoflavone C-glycosides by newly isolated human intestinal bacteria. J. Sci. Food Agric. 2015, 95, 1925–1931. [Google Scholar] [CrossRef]
- de Lima, M.T.R.; Waffo-Téguo, P.; Teissedre, P.L.; Pujolas, A.; Vercauteren, J.; Cabanis, J.C.; Mérillon, J.M. Determination of stilbenes (trans-astringin, cis-and trans-piceid, and cis-and trans-resveratrol) in Portuguese wines. J. Agric. Food Chem. 1999, 47, 2666–2670. [Google Scholar] [CrossRef]
- Wang, D.-G.; Liu, W.-Y.; Chen, G.-T. A simple method for the isolation and purification of resveratrol from Polygonum cuspidatum. J. Pharm. Anal. 2013, 3, 241–247. [Google Scholar] [CrossRef]
- Romero-Pérez, A.I.; Lamuela-Raventós, R.M.; Andrés-Lacueva, C.; de la Torre-Boronat, M.C. Method for the quantitative extraction of resveratrol and piceid isomers in grape berry skins. Effect of powdery mildew on the stilbene content. J. Agric. Food Chem. 2001, 49, 210–215. [Google Scholar] [CrossRef]
- Fakhri, S.; Gravandi, M.M.; Abdian, S.A.-O.; Akkol, E.A.-O.; Farzaei, M.A.-O.; Sobarzo-Sánchez, E.A.-O. The Neuroprotective Role of Polydatin: Neuropharmacological Mechanisms, Molecular Targets, Therapeutic Potentials, and Clinical Perspective. Molecules 2021, 26, 5985. [Google Scholar] [CrossRef]
- Mele, L.; la Noce, M.; Paino, F.; Regad, T.; Wagner, S.; Liccardo, D.; Papaccio, G.; Lombardi, A.; Caraglia, M.; Tirino, V. Glucose-6-phosphate dehydrogenase blockade potentiates tyrosine kinase inhibitor effect on breast cancer cells through autophagy perturbation. J. Exp. Clin. Cancer Res. 2019, 38, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Ming, D.; Songyan, L.; Yawen, C.; Na, Z.; Jing, M.; Zhaowen, X.; Ye, L.; Wa, D.; Jie, L. trans-Polydatin protects the mouse heart against ischemia/reperfusion injury via inhibition of the renin–angiotensin system (RAS) and Rho kinase (ROCK) activity. Food Funct. 2017, 8, 2309–2321. [Google Scholar] [CrossRef] [PubMed]
- Gong, W.; Li, J.; Chen, Z.; Huang, J.; Chen, Q.; Cai, W.; Liu, P.; Huang, H. Polydatin promotes Nrf2-ARE anti-oxidative pathway through activating CKIP-1 to resist HG-induced up-regulation of FN and ICAM-1 in GMCs and diabetic mice kidneys. Free Radic. Biol. Med. 2017, 106, 393–405. [Google Scholar] [CrossRef] [PubMed]
- Lv, R.; Du, L.; Zhang, L.; Zhang, Z. Polydatin attenuates spinal cord injury in rats by inhibiting oxidative stress and microglia apoptosis via Nrf2/HO-1 pathway. Life Sci. 2019, 217, 119–127. [Google Scholar] [CrossRef] [PubMed]
- Zhao, X.-J.; Yu, H.-W.; Yang, Y.-Z.; Wu, W.-Y.; Chen, T.-Y.; Jia, K.-K.; Kang, L.-L.; Jiao, R.-Q.; Kong, L.-D. Polydatin prevents fructose-induced liver inflammation and lipid deposition through increasing miR-200a to regulate Keap1/Nrf2 pathway. Redox Biol. 2018, 18, 124–137. [Google Scholar] [CrossRef]
- Li, T.; Liu, Y.; Xu, W.; Dai, X.; Liu, R.; Gao, Y.; Chen, Z.; Li, Y. Polydatin mediates Parkin-dependent mitophagy and protects against mitochondria-dependent apoptosis in acute respiratory distress syndrome. Lab. Investig. 2019, 99, 819–829. [Google Scholar] [CrossRef]
- Zeng, Z.; Chen, Z.; Xu, S.; Song, R.; Yang, H.; Zhao, K.-S. Polydatin alleviates small intestine injury during hemorrhagic shock as a SIRT1 activator. Oxidative Med. Cell. Longev. 2015, 2015, 965961. [Google Scholar] [CrossRef]
- Jiang, K.-F.; Zhao, G.; Deng, G.-Z.; Wu, H.-C.; Yin, N.-N.; Chen, X.-Y.; Qiu, C.-W.; Peng, X.-L. Polydatin ameliorates Staphylococcus aureus-induced mastitis in mice via inhibiting TLR2-mediated activation of the p38 MAPK/NF-κB pathway. Acta Pharmacol. Sin. 2017, 38, 211–222. [Google Scholar] [CrossRef]
- Masodsai, K.; Lin, Y.Y.; Chaunchaiyakul, R.; Su, C.T.; Lee, S.D.; Yang, A.L. Twelve-Week Protocatechuic Acid Administration Improves Insulin-Induced and Insulin-Like Growth Factor-1-Induced Vasorelaxation and Antioxidant Activities in Aging Spontaneously Hypertensive Rats. Nutrients 2019, 11, 699. [Google Scholar] [CrossRef]
- Kang, L.; Liu, S.; Li, J.; Tian, Y.; Xue, Y.; Liu, X. Parkin and Nrf2 prevent oxidative stress-induced apoptosis in intervertebral endplate chondrocytes via inducing mitophagy and anti-oxidant defenses. Life Sci. 2020, 243, 117244. [Google Scholar] [CrossRef]
- Li, R.; Maimai, T.; Yao, H.; Liu, X.; He, Z.; Xiao, C.; Wang, Y.; Xie, G. Protective effects of polydatin on LPS-induced endometritis in mice. Microb. Pathog. 2019, 137, 103720. [Google Scholar] [CrossRef] [PubMed]
- Tang, K.S. Protective Effects of Polydatin Against Dementia-Related Disorders. Curr. Neuropharmacol. 2020, 19, 127–135. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Li, Y.; Xun, Y.; Liu, H.; Wei, C.; Wang, H.; Yang, X.; Yuan, S.; Liu, N.; Xiang, S. Polydatin protects neuronal cells from hydrogen peroxide damage by activating CREB/Ngb signaling. Mol. Med. Rep. 2022, 25, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Wu, M.; Li, X.; Wang, S.; Yang, S.; Zhao, R.; Xing, Y.; Liu, L. Polydatin for treating atherosclerotic diseases: A functional and mechanistic overview. Biomed. Pharmacother. 2020, 128, 110308. [Google Scholar] [CrossRef]
- Ahmad, P.; Alvi, S.S.; Iqbal, D.; Khan, M.S. Insights into pharmacological mechanisms of polydatin in targeting risk factors-mediated atherosclerosis. Life Sci. 2020, 254, 117756. [Google Scholar] [CrossRef]
- Sun, Z.; Wang, X. Protective effects of polydatin on multiple organ ischemia-reperfusion injury. Bioorganic Chem. 2019, 94, 103485. [Google Scholar] [CrossRef]
- Liu, C.; Wang, W.; Zhang, K.; Liu, Q.; Ma, T.; Tan, L.; Ma, L. Protective Effects of Polydatin from Grapes and Reynoutria japonica Houtt. on Damaged Macrophages Treated with Acetaminophen. Nutrients 2022, 14, 2077. [Google Scholar] [CrossRef]
- Salem, M.A.; Serag, A.; El-Seedi, H.R.; Hamdan, D.I.; Ezzat, S.M.; Zayed, A. Identification and analysis of toxic phytochemicals. In Phytochemistry, the Military and Health; Elsevier: Amsterdam, The Netherlands, 2021; pp. 443–479. [Google Scholar]
- Rajendran, P.; Chen, Y.F.; Chen, Y.F.; Chung, L.C.; Tamilselvi, S.; Shen, C.Y.; Day, C.H.; Chen, R.J.; Viswanadha, V.P.; Kuo, W.W. The multifaceted link between inflammation and human diseases. J. Cell. Physiol. 2018, 233, 6458–6471. [Google Scholar] [CrossRef]
- Furman, D.; Campisi, J.; Verdin, E.; Carrera-Bastos, P.; Targ, S.; Franceschi, C.; Ferrucci, L.; Gilroy, D.W.; Fasano, A.; Miller, G.W. Chronic inflammation in the etiology of disease across the life span. Nat. Med. 2019, 25, 1822–1832. [Google Scholar] [CrossRef]
- Chen, L.; Deng, H.; Cui, H.; Fang, J.; Zuo, Z.; Deng, J.; Li, Y.; Wang, X.; Zhao, L. Inflammatory responses and inflammation-associated diseases in organs. Oncotarget 2017, 9, 7204–7218. [Google Scholar] [CrossRef]
- Mitchell, S.; Vargas, J.; Hoffmann, A. Signaling via the NFκB system. Wiley Interdiscip. Rev. Syst. Biol. Med. 2016, 8, 227–241. [Google Scholar] [CrossRef] [PubMed]
- Wang, R.X.; Zhou, M.; Ma, H.L.; Qiao, Y.B.; Li, Q.S. The Role of Chronic Inflammation in Various Diseases and Anti-inflammatory Therapies Containing Natural Products. ChemMedChem 2021, 16, 1576–1592. [Google Scholar] [CrossRef] [PubMed]
- Mikulski, D.; Molski, M. Quantitative structure–antioxidant activity relationship of trans-resveratrol oligomers, trans-4,4′-dihydroxystilbene dimer, trans-resveratrol-3-O-glucuronide, glucosides: Trans-piceid, cis-piceid, trans-astringin and trans-resveratrol-4′-O-β-D-glucopyranoside. Eur. J. Med. Chem. 2010, 45, 2366–2380. [Google Scholar]
- Tang, J.; Li, Y.; Wang, J.; Wu, Q.; Yan, H. Polydatin suppresses the development of lung inflammation and fibrosis by inhibiting activation of the NACHT domain-, leucine-rich repeat-, and pyd-containing protein 3 inflammasome and the nuclear factor-κB pathway after Mycoplasma pneumoniae infection. J. Cell. Biochem. 2019, 120, 10137–10144. [Google Scholar] [CrossRef]
- Chen, L.; Lan, Z. Polydatin attenuates potassium oxonate-induced hyperuricemia and kidney inflammation by inhibiting NF-κB/NLRP3 inflammasome activation via the AMPK/SIRT1 pathway. Food Funct. 2017, 8, 1785–1792. [Google Scholar] [CrossRef] [PubMed]
- Peritore, A.F.; D’Amico, R.; Cordaro, M.; Siracusa, R.; Fusco, R.; Gugliandolo, E.; Genovese, T.; Crupi, R.; Di Paola, R.; Cuzzocrea, S. PEA/Polydatin: Anti-Inflammatory and Antioxidant Approach to Counteract DNBS-Induced Colitis. Antioxidants 2021, 10, 464. [Google Scholar] [CrossRef]
- Huang, Q.-H.; Xu, L.-Q.; Liu, Y.-H.; Wu, J.-Z.; Wu, X.; Lai, X.-P.; Li, Y.-C.; Su, Z.-R.; Chen, J.-N.; Xie, Y.-L. Polydatin Protects Rat Liver against Ethanol-Induced Injury: Involvement of CYP2E1/ROS/Nrf2 and TLR4/NF-κB p65 Pathway. Evid. Based Complement. Altern. Med. 2017, 2017, 7953850. [Google Scholar] [CrossRef]
- Lv, R.; Du, L.; Liu, X.; Zhou, F.; Zhang, Z.; Zhang, L. Polydatin alleviates traumatic spinal cord injury by reducing microglial inflammation via regulation of iNOS and NLRP3 inflammasome pathway. Int. Immunopharmacol. 2019, 70, 28–36. [Google Scholar] [CrossRef]
- Liu, H.-B.; Meng, Q.-H.; Huang, C.; Wang, J.-B.; Liu, X.-W. Nephroprotective Effects of Polydatin against Ischemia/Reperfusion Injury: A Role for the PI3K/Akt Signal Pathway. Oxidative Med. Cell. Longev. 2015, 2015, 362158. [Google Scholar] [CrossRef]
- Chen, G.; Yang, Z.; Wen, D.; Guo, J.; Xiong, Q.; Li, P.; Zhao, L.; Wang, J.; Wu, C.; Dong, L. Polydatin has anti-inflammatory and antioxidant effects in LPS-induced macrophages and improves DSS-induced mice colitis. Immun. Inflamm. Dis. 2021, 9, 959–970. [Google Scholar] [CrossRef]
- Chen, L.; Lan, Z.; Lin, Q.; Mi, X.; He, Y.; Wei, L.; Lin, Y.; Zhang, Y.; Deng, X. Polydatin ameliorates renal injury by attenuating oxidative stress-related inflammatory responses in fructose-induced urate nephropathic mice. Food Chem. Toxicol. 2013, 52, 28–35. [Google Scholar] [CrossRef] [PubMed]
- Ruan, W.; Li, J.; Xu, Y.; Wang, Y.; Zhao, F.; Yang, X.; Jiang, H.; Zhang, L.; Saavedra, J.M.; Shi, L.; et al. MALAT1 Up-Regulator Polydatin Protects Brain Microvascular Integrity and Ameliorates Stroke Through C/EBPβ/MALAT1/CREB/PGC-1α/PPARγ Pathway. Cell. Mol. Neurobiol. 2019, 39, 265–286. [Google Scholar] [CrossRef] [PubMed]
- Shiyu, S.; Zhiyu, L.; Mao, Y.; Lin, B.; Lijia, W.; Tianbao, Z.; Jie, C.; Tingyu, L. Polydatin up-regulates Clara cell secretory protein to suppress phospholipase A2 of lung induced by LPS in vivo and in vitro. BMC Cell Biol. 2011, 12, 31. [Google Scholar] [CrossRef] [PubMed]
- Wu, M.J.; Gong, X.; Jiang, R.; Zhang, L.; Li, X.H.; Wan, J.Y. Polydatin Protects against Lipopolysaccharide-Induced Fulminant Hepatic Failure in D-Galactosamine-Sensitized Mice. Int. J. Immunopathol. Pharmacol. 2012, 25, 923–934. [Google Scholar] [CrossRef]
- Kamel, K.M.; Gad, A.M.; Mansour, S.M.; Safar, M.M.; Fawzy, H.M. Novel anti-arthritic mechanisms of Polydatin in complete Freund’s adjuvant-induced arthritis in rats: Involvement of IL-6, STAT-3, IL-17, and NF-κB. Inflammation 2018, 41, 1974–1986. [Google Scholar] [CrossRef]
- Pizzino, G.; Irrera, N.; Cucinotta, M.; Pallio, G.; Mannino, F.; Arcoraci, V.; Squadrito, F.; Altavilla, D.; Bitto, A. Oxidative Stress: Harms and Benefits for Human Health. Oxidative Med. Cell. Longev. 2017, 2017, 8416763. [Google Scholar] [CrossRef]
- Storz, G.; Imlayt, J.A. Oxidative stress. Curr. Opin. Microbiol. 1999, 2, 188–194. [Google Scholar] [CrossRef]
- Liu, Y.-H.; Huang, Q.-H.; Wu, X.; Wu, J.-Z.; Liang, J.-L.; Lin, G.-S.; Xu, L.-Q.; Lai, X.-P.; Su, Z.-R.; Chen, J.-N. Polydatin protects against acetaminophen-induced hepatotoxicity in mice via anti-oxidative and anti-apoptotic activities. Food Funct. 2018, 9, 5891–5902. [Google Scholar] [CrossRef]
- Liu, M.; Huang, Q.; Zhu, Y.; Chen, L.; Li, Y.; Gong, Z.; Ai, K. Harnessing reactive oxygen/nitrogen species and inflammation: Nanodrugs for liver injury. Mater. Today Bio 2022, 13, 100215. [Google Scholar] [CrossRef]
- Kobayashi, E.; Suzuki, T.; Yamamoto, M. Roles Nrf2 Plays in Myeloid Cells and Related Disorders. Oxidative Med. Cell. Longev. 2013, 2013, 529219. [Google Scholar] [CrossRef]
- Shah, F.A.; Kury, L.A.; Li, T.; Zeb, A.; Koh, P.O.; Liu, F.; Zhou, Q.; Hussain, I.; Khan, A.U.; Jiang, Y.; et al. Polydatin Attenuates Neuronal Loss via Reducing Neuroinflammation and Oxidative Stress in Rat MCAO Models. Front. Pharmacol. 2019, 10, 663. [Google Scholar] [CrossRef] [PubMed]
- Ji, H.; Zhang, X.; Du, Y.; Liu, H.; Li, S.; Li, L. Polydatin modulates inflammation by decreasing NF-κB activation and oxidative stress by increasing Gli1, Ptch1, SOD1 expression and ameliorates blood–brain barrier permeability for its neuroprotective effect in pMCAO rat brain. Brain Res. Bull. 2012, 87, 50–59. [Google Scholar] [CrossRef] [PubMed]
- Meng, Q.-h.; Liu, H.-b.; Wang, J.-b. Polydatin ameliorates renal ischemia/reperfusion injury by decreasing apoptosis and oxidative stress through activating sonic hedgehog signaling pathway. Food Chem. Toxicol. 2016, 96, 215–225. [Google Scholar] [CrossRef] [PubMed]
- Huang, K.; Chen, C.; Hao, J.; Huang, J.; Wang, S.; Liu, P.; Huang, H. Polydatin promotes Nrf2-ARE anti-oxidative pathway through activating Sirt1 to resist AGEs-induced upregulation of fibronetin and transforming growth factor-β1 in rat glomerular messangial cells. Mol. Cell. Endocrinol. 2015, 399, 178–189. [Google Scholar] [CrossRef]
- Yu, L.; Li, Z.; Dong, X.; Xue, X.; Liu, Y.; Xu, S.; Zhang, J.; Han, J.; Yang, Y.; Wang, H. Polydatin Protects Diabetic Heart against Ischemia-Reperfusion Injury via Notch1/Hes1-Mediated Activation of Pten/Akt Signaling. Oxidative Med. Cell. Longev. 2018, 2018, 2750695. [Google Scholar] [CrossRef]
- Tan, Y.-Y.; Chen, L.-X.; Fang, L.; Zhang, Q. Cardioprotective effects of polydatin against myocardial injury in diabetic rats via inhibition of NADPH oxidase and NF-κB activities. BMC Complement. Med. Ther. 2020, 20, 12. [Google Scholar] [CrossRef]
- Dan, S.; Bangle, Z.; Menglei, H.; Qiuju, H.; Chunlai, Z.; Siyuan, Z. Comparison of Resveratrol and Polydatin on Anti-oxidative Activities. China Pharm. 2010, 4, 471–473. [Google Scholar]
- Chai, Y.-y.; Wang, F.; Li, Y.-l.; Liu, K.; Xu, H. Antioxidant Activities of Stilbenoids from Rheum emodi Wall. Evid. Based Complement. Altern. Med. 2012, 2012, 603678. [Google Scholar] [CrossRef]
- Zhang, H.; Yu, C.-H.; Jiang, Y.-P.; Peng, C.; He, K.; Tang, J.-Y.; Xin, H.-L. Protective effects of polydatin from Polygonum cuspidatum against carbon tetrachloride-induced liver injury in mice. PLoS ONE 2012, 7, e46574. [Google Scholar] [CrossRef]
- Miao, Q.; Wang, S.; Miao, S.; Wang, J.; Xie, Y.; Yang, Q. Cardioprotective effect of polydatin against ischemia/reperfusion injury: Roles of protein kinase C and mito KATP activation. Phytomedicine 2011, 19, 8–12. [Google Scholar] [CrossRef]
- Pang, N.; Chen, T.; Deng, X.; Chen, N.; Li, R.; Ren, M.; Li, Y.; Luo, M.; Hao, H.; Wu, J.; et al. Polydatin Prevents Methylglyoxal-Induced Apoptosis through Reducing Oxidative Stress and Improving Mitochondrial Function in Human Umbilical Vein Endothelial Cells. Oxidative Med. Cell. Longev. 2017, 2017, 7180943. [Google Scholar] [CrossRef] [PubMed]
- Lan, T.; Zhuang, L.; Li, S.; Yang, G.; Xuan, Y.; Guo, J. Polydatin attenuates hepatic stellate cell proliferation and liver fibrosis by suppressing sphingosine kinase 1. Biomed. Pharmacother. 2020, 130, 110586. [Google Scholar] [CrossRef] [PubMed]
- Carson, D.A.; Ribeiro, J.M. Apoptosis and disease. Lancet 1993, 341, 1251–1254. [Google Scholar] [CrossRef]
- Lv, T.; Shen, L.; Yang, L.; Diao, W.; Yang, Z.; Zhang, Y.; Yu, S.; Li, Y. Polydatin ameliorates dextran sulfate sodium-induced colitis by decreasing oxidative stress and apoptosis partially via Sonic hedgehog signaling pathway. Int. Immunopharmacol. 2018, 64, 256–263. [Google Scholar] [CrossRef]
- Yu, H.I.; Shen, H.C.; Chen, S.H.; Lim, Y.P.; Chuang, H.H.; Tai, T.S.; Kung, F.P.; Lu, C.H.; Hou, C.Y.; Lee, Y.R. Autophagy Modulation in Human Thyroid Cancer Cells following Aloperine Treatment. Int. J. Mol. Sci. 2019, 20, 5315. [Google Scholar] [CrossRef]
- Wu, Z.; Luan, Z.; Zhang, X.; Zou, K.; Ma, S.; Yang, Z.; Feng, W.; He, M.; Jiang, L.; Li, J.; et al. Retraction Note: Chondro-protective effects of polydatin in osteoarthritis through its effect on restoring dysregulated autophagy via modulating MAPK, and PI3K/Akt signaling pathways. Sci. Rep. 2020, 10, 11511. [Google Scholar] [CrossRef]
- Fakhri, S.; Abbaszadeh, F.; Moradi, S.Z.; Cao, H.; Khan, H.; Xiao, J. Effects of Polyphenols on Oxidative Stress, Inflammation, and Interconnected Pathways during Spinal Cord Injury. Oxidative Med. Cell. Longev. 2022, 2022, 8100195. [Google Scholar] [CrossRef]
- Fakhri, S.; Piri, S.; Moradi, S.Z.; Khan, H. Phytochemicals Targeting Oxidative Stress, Interconnected Neuroinflammatory, and Neuroapoptotic Pathways Following Radiation. Curr. Neuropharmacol. 2022, 20, 836–856. [Google Scholar] [CrossRef]
- Mattiuzzi, C.; Lippi, G. Current cancer epidemiology. J. Epidemiol. Glob. Health 2019, 9, 217. [Google Scholar] [CrossRef]
- Tong, Y.; Wang, K.; Sheng, S.; Cui, J. Polydatin ameliorates chemotherapy-induced cognitive impairment (chemobrain) by inhibiting oxidative stress, inflammatory response, and apoptosis in rats. Biosci. Biotechnol. Biochem. 2020, 84, 1201–1210. [Google Scholar] [CrossRef]
- Schirrmacher, V. From chemotherapy to biological therapy: A review of novel concepts to reduce the side effects of systemic cancer treatment. Int. J. Oncol. 2019, 54, 407–419. [Google Scholar] [PubMed]
- Fakhri, S.; Abbaszadeh, F.; Jorjani, M.; Pourgholami, M.H. The effects of anticancer medicinal herbs on vascular endothelial growth factor based on pharmacological aspects: A review study. Nutr. Cancer 2021, 73, 1–15. [Google Scholar] [CrossRef]
- Fakhri, S.; Khodamorady, M.; Naseri, M.; Farzaei, M.H.; Khan, H. The ameliorating effects of anthocyanins on the cross-linked signaling pathways of cancer dysregulated metabolism. Pharmacol. Res. 2020, 159, 104895. [Google Scholar] [CrossRef]
- Nouri, Z.; Fakhri, S.; Nouri, K.; Wallace, C.E.; Farzaei, M.H.; Bishayee, A. Targeting multiple signaling pathways in cancer: The rutin therapeutic approach. Cancers 2020, 12, 2276. [Google Scholar] [CrossRef] [PubMed]
- Cao, W.J.; Wu, K.; Wang, C.; Wan, D.M. Polydatin-induced cell apoptosis and cell cycle arrest are potentiated by Janus kinase 2 inhibition in leukemia cells. Mol. Med. Rep. 2016, 13, 3297–3302. [Google Scholar] [CrossRef] [PubMed]
- Quagliariello, V.; Berretta, M.; Buccolo, S.; Iovine, M.; Paccone, A.; Cavalcanti, E.; Taibi, R.; Montopoli, M.; Botti, G.; Maurea, N. Polydatin reduces cardiotoxicity and enhances the anticancer effects of sunitinib by decreasing pro-oxidative stress, pro-inflammatory cytokines, and NLRP3 inflammasome expression. Front. Oncol. 2021, 11, 2188. [Google Scholar] [CrossRef] [PubMed]
- Jin, Y.; Zhan, X.; Zhang, B.; Chen, Y.; Liu, C.; Yu, L. Polydatin exerts an antitumor effect through regulating the miR-382/PD-L1 axis in colorectal cancer. Cancer Biother. Radiopharm. 2020, 35, 83–91. [Google Scholar] [CrossRef]
- Chen, Q.; Zeng, Y.-N.; Zhang, K.; Zhao, Y.; Wu, Y.-Y.; Li, G.; Cheng, H.-Y.; Zhang, M.; Lai, F.; Wang, J.-B. Polydatin increases radiosensitivity by inducing apoptosis of stem cells in colorectal cancer. Int. J. Biol. Sci. 2019, 15, 430. [Google Scholar] [CrossRef]
- Wang, C.; Luo, Y.; Lu, J.; Wang, Y.; Sheng, G. Polydatin induces apoptosis and inhibits growth of acute monocytic leukemia cells. J. Biochem. Mol. Toxicol. 2016, 30, 200–205. [Google Scholar] [CrossRef]
- Li, H.; Shi, B.; Li, Y.; Yin, F. Polydatin inhibits cell proliferation and induces apoptosis in laryngeal cancer and HeLa cells via suppression of the PDGF/AKT signaling pathway. J. Biochem. Mol. Toxicol. 2017, 31, e21900. [Google Scholar] [CrossRef]
- Liu, H.; Zhao, S.; Zhang, Y.; Wu, J.; Peng, H.; Fan, J.; Liao, J. Reactive oxygen species-mediated endoplasmic reticulum stress and mitochondrial dysfunction contribute to polydatin-induced apoptosis in human nasopharyngeal carcinoma CNE cells. J. Cell. Biochem. 2011, 112, 3695–3703. [Google Scholar] [CrossRef] [PubMed]
- Hogg, S.J.; Chitcholtan, K.; Hassan, W.; Sykes, P.H.; Garrill, A. Resveratrol, acetyl-resveratrol, and polydatin exhibit antigrowth activity against 3D cell aggregates of the SKOV-3 and OVCAR-8 ovarian cancer cell lines. Obstet. Gynecol. Int. 2015, 2015, 279591. [Google Scholar] [CrossRef] [PubMed]
- Zou, J.; Yang, Y.; Yang, Y.; Liu, X. Polydatin suppresses proliferation and metastasis of non-small cell lung cancer cells by inhibiting NLRP3 inflammasome activation via NF-κB pathway. Biomed. Pharmacother. 2018, 108, 130–136. [Google Scholar] [CrossRef] [PubMed]
- Verma, N.; Tiku, A.B. Polydatin-Induced Direct and Bystander Effects in A549 Lung Cancer Cell Line. Nutr. Cancer 2021, 74, 237–249. [Google Scholar] [CrossRef]
- Zhang, Y.; Zhuang, Z.; Meng, Q.; Jiao, Y.; Xu, J.; Fan, S. Polydatin inhibits growth of lung cancer cells by inducing apoptosis and causing cell cycle arrest. Oncol. Lett. 2014, 7, 295–301. [Google Scholar] [CrossRef]
- Chen, Y.; Niu, J.; Li, L.; Li, Z.; Jiang, J.; Zhu, M.; Dong, T.; Zhang, J.; Shi, C.; Xu, P. Polydatin executes anticancer effects against glioblastoma multiforme by inhibiting the EGFR-AKT/ERK1/2/STAT3-SOX2/Snail signaling pathway. Life Sci. 2020, 258, 118158. [Google Scholar] [CrossRef]
- Yang, B.; Zhao, S. Polydatin regulates proliferation, apoptosis and autophagy in multiple myeloma cells through mTOR/p70s6k pathway. OncoTargets Ther. 2017, 10, 935. [Google Scholar] [CrossRef]
- Pan, J.-H.; Wang, H.-B.; Du, X.-F.; Liu, J.-Y.; Zhang, D.-J. Polydatin induces human cervical cancer cell apoptosis via PI3K/AKT/mTOR signaling pathway. Zhongguo Zhong Yao Za Zhi China J. Chin. Mater. Med. 2017, 42, 2345–2349. [Google Scholar]
- Bai, L.; Ma, Y.; Wang, X.; Feng, Q.; Zhang, Z.; Wang, S.; Zhang, H.; Lu, X.; Xu, Y.; Zhao, E. Polydatin Inhibits Cell Viability, Migration, and Invasion Through Suppressing the c-Myc Expression in Human Cervical Cancer. Front. Cell Dev. Biol. 2021, 9, 587218. [Google Scholar] [CrossRef]
- Chen, S.; Tao, J.; Zhong, F.; Jiao, Y.; Xu, J.; Shen, Q.; Wang, H.; Fan, S.; Zhang, Y. Polydatin down-regulates the phosphorylation level of Creb and induces apoptosis in human breast cancer cell. PLoS ONE 2017, 12, e0176501. [Google Scholar] [CrossRef]
- Zhang, T.; Zhu, X.; Wu, H.; Jiang, K.; Zhao, G.; Shaukat, A.; Deng, G.; Qiu, C. Targeting the ROS/PI3K/AKT/HIF-1α/HK2 axis of breast cancer cells: Combined administration of Polydatin and 2-Deoxy-d-glucose. J. Cell. Mol. Med. 2019, 23, 3711–3723. [Google Scholar] [CrossRef] [PubMed]
- Zhao, W.; Chen, Z.; Guan, M. Polydatin enhances the chemosensitivity of osteosarcoma cells to paclitaxel. J. Cell. Biochem. 2019, 120, 17481–17490. [Google Scholar] [CrossRef] [PubMed]
- Jiang, C.-q.; Ma, L.-l.; Lv, Z.-d.; Feng, F.; Chen, Z.; Liu, Z.-D. Polydatin induces apoptosis and autophagy via STAT3 signaling in human osteosarcoma MG-63 cells. J. Nat. Med. 2020, 74, 533–544. [Google Scholar] [CrossRef] [PubMed]
- Xu, G.; Kuang, G.; Jiang, W.; Jiang, R.; Jiang, D. Polydatin promotes apoptosis through upregulation the ratio of Bax/Bcl-2 and inhibits proliferation by attenuating the β-catenin signaling in human osteosarcoma cells. Am. J. Transl. Res. 2016, 8, 922. [Google Scholar] [PubMed]
- Hu, T.; Fei, Z.; Su, H.; Xie, R.; Chen, L. Polydatin inhibits proliferation and promotes apoptosis of doxorubicin-resistant osteosarcoma through LncRNA TUG1 mediated suppression of Akt signaling. Toxicol. Appl. Pharmacol. 2019, 371, 55–62. [Google Scholar] [CrossRef]
- Jiao, Y.; Wu, Y.; Du, D. Polydatin inhibits cell proliferation, invasion and migration, and induces cell apoptosis in hepatocellular carcinoma. Braz. J. Med. Biol. Res. 2018, 51, e6867. [Google Scholar] [CrossRef]
- Jiang, J.; Chen, Y.; Dong, T.; Yue, M.; Zhang, Y.; An, T.; Zhang, J.; Liu, P.; Yang, X. Polydatin inhibits hepatocellular carcinoma via the AKT/STAT3-FOXO1 signaling pathway Corrigendum in /10.3892/ol.2019.10856. Oncol. Lett. 2019, 17, 4505–4513. [Google Scholar]
- Bang, T.-H.; Park, B.-S.; Kang, H.-M.; Kim, J.-H.; Kim, I.-R. Polydatin, a Glycoside of Resveratrol, Induces Apoptosis and Inhibits Metastasis Oral Squamous Cell Carcinoma Cells In Vitro. Pharmaceuticals 2021, 14, 902. [Google Scholar] [CrossRef]
- Ince, S.; Acaroz, D.A.; Neuwirth, O.; Demirel, H.H.; Denk, B.; Kucukkurt, I.; Turkmen, R. Protective effect of polydatin, a natural precursor of resveratrol, against cisplatin-induced toxicity in rats. Food Chem. Toxicol. 2014, 72, 147–153. [Google Scholar] [CrossRef]
- Xu, Z.; Feng, W.; Shen, Q.; Yu, N.; Yu, K.; Wang, S.; Chen, Z.; Shioda, S.; Guo, Y. Rhizoma Coptidis and Berberine as a Natural Drug to Combat Aging and Aging-Related Diseases via Anti-Oxidation and AMPK Activation. Aging Dis. 2017, 8, 760–777. [Google Scholar] [CrossRef]
- Fakhri, S.; Abbaszadeh, F.; Dargahi, L.; Jorjani, M. Astaxanthin: A mechanistic review on its biological activities and health benefits. Pharmacol. Res. 2018, 136, 1–20. [Google Scholar] [CrossRef] [PubMed]
- Goldstein, L.; Brown, S. The low-density lipoprotein pathway and its relation to atherosclerosis. Annu. Rev. Biochem. 1977, 46, 897–930. [Google Scholar] [CrossRef] [PubMed]
- Chen, G.; Liu, G.; Cao, D.; Jin, M.; Guo, D.; Yuan, X. Polydatin protects against acute myocardial infarction-induced cardiac damage by activation of Nrf2/HO-1 signaling. J. Nat. Med. 2019, 73, 85–92. [Google Scholar] [CrossRef]
- Ma, Y.; Gong, X.; Mo, Y.; Wu, S. Polydatin inhibits the oxidative stress-induced proliferation of vascular smooth muscle cells by activating the eNOS/SIRT1 pathway. Int. J. Mol. Med. 2016, 37, 1652–1660. [Google Scholar] [CrossRef]
- Miao, Q.; Shi, X.-P.; Ye, M.-X.; Zhang, J.; Miao, S.; Wang, S.-W.; Li, B.; Jiang, X.-X.; Zhang, S.; Hu, N. Polydatin attenuates hypoxic pulmonary hypertension and reverses remodeling through protein kinase C mechanisms. Int. J. Mol. Sci. 2012, 13, 7776–7787. [Google Scholar] [CrossRef] [PubMed]
- Gugliandolo, E.; Fusco, R.; Biundo, F.; D’Amico, R.; Benedetto, F.; Di Paola, R.; Cuzzocrea, S. Palmitoylethanolamide and Polydatin combination reduces inflammation and oxidative stress in vascular injury. Pharmacol. Res. 2017, 123, 83–92. [Google Scholar] [CrossRef]
- Yuan, X.; Chen, G.; Guo, D.; Xu, L.; Gu, Y. Polydatin alleviates septic myocardial injury by promoting SIRT6-mediated autophagy. Inflammation 2020, 43, 785–795. [Google Scholar] [CrossRef]
- Gao, J.P.; Chen, C.X.; Gu, W.L.; Wu, Q.; Wang, Y.; Lü, J. Effects of polydatin on attenuating ventricular remodeling in isoproterenol-induced mouse and pressure-overload rat models. Fitoterapia 2010, 81, 953–960. [Google Scholar] [CrossRef]
- Gao, Y.; Gao, J.; Chen, C.; Wang, H.; Guo, J.; Wu, R. Cardioprotective effect of polydatin on ventricular remodeling after myocardial infarction in coronary artery ligation rats. Planta Med. 2015, 81, 568–577. [Google Scholar] [CrossRef]
- Zhang, Q.; Tan, Y.; Zhang, N.; Yao, F. Polydatin prevents angiotensin II-induced cardiac hypertrophy and myocardial superoxide generation. Exp. Biol. Med. 2015, 240, 1352–1361. [Google Scholar] [CrossRef]
- Zhang, M.; Wang, S.; Cheng, Z.; Xiong, Z.; Lv, J.; Yang, Z.; Li, T.; Jiang, S.; Gu, J.; Sun, D. Polydatin ameliorates diabetic cardiomyopathy via Sirt3 activation. Biochem. Biophys. Res. Commun. 2017, 493, 1280–1287. [Google Scholar] [CrossRef] [PubMed]
- Pessina, A.; Di Vincenzo, M.; Maradonna, F.; Marchegiani, F.; Olivieri, F.; Randazzo, B.; Gioacchini, G.; Carnevali, O. Polydatin Beneficial Effects in Zebrafish Larvae Undergoing Multiple Stress Types. Int. J. Environ. Res. Public Health 2021, 18, 1116. [Google Scholar] [CrossRef]
- Huang, B.; Xue, L.; Wu, Y.; Jiang, Q. Effect and mechanism of polydatin on diabetic myocardial hypertrophy in mice. Zhongguo Zhong Yao Za Zhi China J. Chin. Mater. Med. 2015, 40, 4256–4261. [Google Scholar]
- Du, J.; Sun, L.-N.; Xing, W.-W.; Huang, B.-K.; Jia, M.; Wu, J.-Z.; Zhang, H.; Qin, L.-P. Lipid-lowering effects of polydatin from Polygonum cuspidatum in hyperlipidemic hamsters. Phytomedicine 2009, 16, 652–658. [Google Scholar] [CrossRef] [PubMed]
- Xing, W.-W.; Wu, J.-Z.; Jia, M.; Du, J.; Zhang, H.; Qin, L.-P. Effects of polydatin from Polygonum cuspidatum on lipid profile in hyperlipidemic rabbits. Biomed. Pharmacother. 2009, 63, 457–462. [Google Scholar] [CrossRef] [PubMed]
- Peng, Y.; Xu, J.; Zeng, Y.; Chen, L.; Le Xu, X. Polydatin attenuates atherosclerosis in apolipoprotein E-deficient mice: Role of reverse cholesterol transport. Phytomedicine 2019, 62, 152935. [Google Scholar] [CrossRef]
- Huang, Z.; Tian, G.; Cheng, S.; Zhao, D.; Zhang, Y.; Jia, Y.; Zhou, F. Polydatin attenuates atherosclerosis in ApoE−/− mice through PBEF mediated reduction of cholesterol deposition. Am. J. Chin. Med. 2018, 46, 1841–1859. [Google Scholar] [CrossRef]
- Xiong, Q.; Yan, Z.; Liang, J.; Yuan, J.; Chen, X.; Zhou, L.; Hu, Y.; Wu, J.; Jing, Y.; Zhang, Q. Polydatin alleviates high-fat diet induced atherosclerosis in apolipoprotein E-deficient mice by autophagic restoration. Phytomedicine 2021, 81, 153301. [Google Scholar] [CrossRef]
- Zheng, L.; Wu, J.; Mo, J.; Guo, L.; Wu, X.; Bao, Y. Polydatin inhibits adipose tissue inflammation and ameliorates lipid metabolism in high-fat-fed mice. BioMed Res. Int. 2019, 2019, 7196535. [Google Scholar] [CrossRef]
- Wu, M.; Liu, M.; Guo, G.; Zhang, W.; Liu, L. Polydatin inhibits formation of macrophage-derived foam cells. Evid. Based Complement. Altern. Med. 2015, 2015, 729017. [Google Scholar] [CrossRef]
- Zhang, L.-P.; Ma, H.-J.; Bu, H.-M.; Wang, M.-L.; Li, Q.; Qi, Z.; Zhang, Y. Polydatin attenuates ischemia/reperfusion-induced apoptosis in myocardium of the rat. Sheng Li Xue Bao 2009, 61, 367–372. [Google Scholar] [PubMed]
- Zhang, L.-P.; Yang, C.-Y.; Wang, Y.-P.; Cui, F.; Zhang, Y. Protective effect of polydatin against ischemia/reperfusion injury in rat heart. Sheng Li Xue Bao [Acta Physiol. Sin.] 2008, 60, 161–168. [Google Scholar]
- Ling, Y.; Chen, G.; Deng, Y.; Tang, H.; Ling, L.; Zhou, X.; Song, X.; Yang, P.; Liu, Y.; Li, Z. Polydatin post-treatment alleviates myocardial ischaemia/reperfusion injury by promoting autophagic flux. Clin. Sci. 2016, 130, 1641–1653. [Google Scholar] [CrossRef] [PubMed]
- Jiang, X.; Liu, W.; Deng, J.; Lan, L.; Xue, X.; Zhang, C.; Cai, G.; Luo, X.; Liu, J. Polydatin protects cardiac function against burn injury by inhibiting sarcoplasmic reticulum Ca2+ leak by reducing oxidative modification of ryanodine receptors. Free Radic. Biol. Med. 2013, 60, 292–299. [Google Scholar] [CrossRef] [PubMed]
- Yousef, A.I.; Shawki, H.H.; El-Shahawy, A.A.; Abd El-Twab, S.M.; Abdel-Moneim, A.; Oishi, H. Polydatin mitigates pancreatic β-cell damage through its antioxidant activity. Biomed. Pharmacother. 2021, 133, 111027. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Q.; Tan, Y.; Zhang, N.; Yao, F. Polydatin supplementation ameliorates diet-induced development of insulin resistance and hepatic steatosis in rats. Mol. Med. Rep. 2015, 11, 603–610. [Google Scholar] [CrossRef]
- Xie, X.; Peng, J.; Huang, K.; Huang, J.; Shen, X.; Liu, P.; Huang, H. Polydatin ameliorates experimental diabetes-induced fibronectin through inhibiting the activation of NF-κB signaling pathway in rat glomerular mesangial cells. Mol. Cell. Endocrinol. 2012, 362, 183–193. [Google Scholar] [CrossRef]
- Chen, C.; Huang, K.; Hao, J.; Huang, J.; Yang, Z.; Xiong, F.; Liu, P.; Huang, H. Polydatin attenuates AGEs-induced upregulation of fibronectin and ICAM-1 in rat glomerular mesangial cells and db/db diabetic mice kidneys by inhibiting the activation of the SphK1-S1P signaling pathway. Mol. Cell. Endocrinol. 2016, 427, 45–56. [Google Scholar] [CrossRef]
- Wang, Y.; Ye, J.; Li, J.; Chen, C.; Huang, J.; Liu, P.; Huang, H. Polydatin ameliorates lipid and glucose metabolism in type 2 diabetes mellitus by downregulating proprotein convertase subtilisin/kexin type 9 (PCSK9). Cardiovasc. Diabetol. 2016, 15, 1–13. [Google Scholar] [CrossRef]
- Hao, J.; Chen, C.; Huang, K.; Huang, J.; Li, J.; Liu, P.; Huang, H. Polydatin improves glucose and lipid metabolism in experimental diabetes through activating the Akt signaling pathway. Eur. J. Pharmacol. 2014, 745, 152–165. [Google Scholar] [CrossRef]
- Wu, Y.; Xue, L.; Du, W.; Huang, B.; Tang, C.; Liu, C.; Qiu, H.; Jiang, Q. Polydatin restores endothelium-dependent relaxation in rat aorta rings impaired by high glucose: A Novel insight into the PPARβ-NO signaling pathway. PLoS ONE 2015, 10, e0126249. [Google Scholar] [CrossRef] [PubMed]
- Wang, D.; Zhou, Z.; Yuan, L. Polydatin reverses oxidation low lipoprotein (oxLDL)-induced apoptosis of human umbilical vein endothelial cells via regulating the miR-26a-5p/BID axis. Eur. J. Histochem. 2022, 66. [Google Scholar] [CrossRef] [PubMed]
- Bheereddy, P.; Yerra, V.G.; Kalvala, A.K.; Sherkhane, B.; Kumar, A. SIRT1 activation by polydatin alleviates oxidative damage and elevates mitochondrial biogenesis in experimental diabetic neuropathy. Cell. Mol. Neurobiol. 2020, 41, 1563–1577. [Google Scholar] [CrossRef] [PubMed]
- Liu, B.; Li, S.; Sui, X.; Guo, L.; Liu, X.; Li, H.; Gao, L.; Cai, S.; Li, Y.; Wang, T. Root extract of Polygonum cuspidatum Siebold & Zucc. ameliorates DSS-induced ulcerative colitis by affecting NF-kappaB signaling pathway in a mouse model via synergistic effects of polydatin, resveratrol, and emodin. Front. Pharmacol. 2018, 9, 347. [Google Scholar]
- Yao, J.; Wang, J.-Y.; Liu, L.; Zeng, W.-S.; Li, Y.-X.; Xun, A.-Y.; Zhao, L.; Jia, C.-H.; Feng, J.-L.; Wei, X.-X. Polydatin ameliorates DSS-induced colitis in mice through inhibition of nuclear factor-kappaB activation. Planta Med. 2011, 77, 421–427. [Google Scholar] [CrossRef]
- Zeng, Z.; Yang, Y.; Dai, X.; Xu, S.; Li, T.; Zhang, Q.; Zhao, K.-s.; Chen, Z. Polydatin ameliorates injury to the small intestine induced by hemorrhagic shock via SIRT3 activation-mediated mitochondrial protection. Expert Opin. Ther. Targets 2016, 20, 645–652. [Google Scholar] [CrossRef]
- Li, P.; Wang, X.; Zhao, M.; Song, R.; Zhao, K.-s. Polydatin protects hepatocytes against mitochondrial injury in acute severe hemorrhagic shock via SIRT1-SOD2 pathway. Expert Opin. Ther. Targets 2015, 19, 997–1010. [Google Scholar] [CrossRef]
- Zhao, G.; Jiang, K.; Wu, H.; Qiu, C.; Deng, G.; Peng, X. Polydatin reduces Staphylococcus aureus lipoteichoic acid-induced injury by attenuating reactive oxygen species generation and TLR 2-NF κB signalling. J. Cell. Mol. Med. 2017, 21, 2796–2808. [Google Scholar] [CrossRef]
- Qiu, Y.; Pan, X.; Hu, Y. Polydatin ameliorates pulmonary fibrosis by suppressing inflammation and the epithelial mesenchymal transition via inhibiting the TGF-β/Smad signaling pathway. RSC Adv. 2019, 9, 8104–8112. [Google Scholar] [CrossRef]
- Li, X.-h.; Gong, X.; Zhang, L.; Jiang, R.; Li, H.-z.; Wu, M.-j.; Wan, J.-y. Protective effects of polydatin on septic lung injury in mice via upregulation of HO-1. Mediat. Inflamm. 2013, 2013, 354087. [Google Scholar] [CrossRef]
- Ricordi, C.; Pacifici, F.; Lanzoni, G.; Palamara, A.T.; Garaci, E.; Della-Morte, D. Dietary and protective factors to halt or mitigate progression of autoimmunity, COVID-19 and its associated metabolic diseases. Int. J. Mol. Sci. 2021, 22, 3134. [Google Scholar] [CrossRef] [PubMed]
- Lai, Y.; Zhou, C.; Huang, P.; Dong, Z.; Mo, C.; Xie, L.; Lin, H.; Zhou, Z.; Deng, G.; Liu, Y. Polydatin alleviated alcoholic liver injury in zebrafish larvae through ameliorating lipid metabolism and oxidative stress. J. Pharmacol. Sci. 2018, 138, 46–53. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.; Chan, H.; Zhang, L.; Liu, X.; Ho, I.H.; Zhang, X.; Ho, J.; Hu, W.; Tian, Y.; Kou, S. The phytochemical polydatin ameliorates non-alcoholic steatohepatitis by restoring lysosomal function and autophagic flux. J. Cell. Mol. Med. 2019, 23, 4290–4300. [Google Scholar] [CrossRef]
- Luo, J.-L.; Chen, S.; Wang, L.; Zhao, X.-H.; Piao, C.-L. Pharmacological effects of polydatin in the treatment of metabolic diseases: A review. Phytomedicine 2022, 154161. [Google Scholar] [CrossRef]
- Zhang, J.; Tan, Y.; Yao, F.; Zhang, Q. Polydatin alleviates non-alcoholic fatty liver disease in rats by inhibiting the expression of TNF-α and SREBP-1c. Mol. Med. Rep. 2012, 6, 815–820. [Google Scholar] [CrossRef]
- Li, R.; Li, J.; Huang, Y.; Li, H.; Yan, S.; Lin, J.; Chen, Y.; Wu, L.; Liu, B.; Wang, G. Polydatin attenuates diet-induced nonalcoholic steatohepatitis and fibrosis in mice. Int. J. Biol. Sci. 2018, 14, 1411. [Google Scholar] [CrossRef]
- Zhang, H.; Chen, Y.; Pei, Z.; Gao, H.; Shi, W.; Sun, M.; Xu, Q.; Zhao, J.; Meng, W.; Xiao, K. Protective effects of polydatin against sulfur mustard-induced hepatic injury. Toxicol. Appl. Pharmacol. 2019, 367, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Zhao, X.; Li, R.; Liu, Y.; Zhang, X.; Zhang, M.; Zeng, Z.; Wu, L.; Gao, X.; Lan, T.; Wang, Y. Polydatin protects against carbon tetrachloride-induced liver fibrosis in mice. Arch. Biochem. Biophys. 2017, 629, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Huang, Z.-S.; Wang, Z.-W.; Liu, M.-P.; Zhong, S.-Q.; Li, Q.-M.; Rong, X.-L. Protective effects of polydatin against CCl4-induced injury to primarily cultured rat hepatocytes. World J. Gastroenterol. 1999, 5, 41. [Google Scholar] [CrossRef]
- Zeng, Z.; Chen, Z.; Li, T.; Zhang, J.; Gao, Y.; Xu, S.; Cai, S.; Zhao, K.S. Polydatin: A new therapeutic agent against multiorgan dysfunction. J. Surg. Res. 2015, 198, 192–199. [Google Scholar] [CrossRef]
- Dumurgier, J.; Tzourio, C. Epidemiology of neurological diseases in older adults. Rev. Neurol. 2020, 176, 642–648. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Zhang, D.-q.; Liao, Z.; Wang, B.; Gong, S.; Wang, C.; Zhang, M.-z.; Wang, G.-h.; Cai, H.; Liao, F.-F. Anti-oxidant polydatin (piceid) protects against substantia nigral motor degeneration in multiple rodent models of Parkinson’s disease. Mol. Neurodegener. 2015, 10, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Zhao, X.; Qin, J.; Li, H.; Feng, X.; Lv, Y.; Yang, J. Effect of Polydatin on Neurological Function and the Nrf2 Pathway during Intracerebral Hemorrhage. J. Mol. Neurosci. 2020, 70, 1332–1337. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Song, R.; Chen, Y.; Zhao, M.; Zhao, K.-s. Polydatin–a new mitochondria protector for acute severe hemorrhagic shock treatment. Expert Opin. Investig. Drugs 2013, 22, 169–179. [Google Scholar] [CrossRef]
- Wang, X.; Song, R.; Bian, H.N.; Brunk, U.T.; Zhao, M.; Zhao, K.-s. Polydatin, a natural polyphenol, protects arterial smooth muscle cells against mitochondrial dysfunction and lysosomal destabilization following hemorrhagic shock. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2012, 302, R805–R814. [Google Scholar] [CrossRef]
- Li, R.-P.; Wang, Z.-Z.; Sun, M.-X.; Hou, X.-L.; Sun, Y.; Deng, Z.-F.; Xiao, K. Polydatin protects learning and memory impairments in a rat model of vascular dementia. Phytomedicine 2012, 19, 677–681. [Google Scholar] [CrossRef]
- Huang, L.; He, S.; Cai, Q.; Li, F.; Wang, S.; Tao, K.; Xi, Y.; Qin, H.; Gao, G.; Feng, D. Polydatin alleviates traumatic brain injury: Role of inhibiting ferroptosis. Biochem. Biophys. Res. Commun. 2021, 556, 149–155. [Google Scholar] [CrossRef]
- Huang, B.; Liu, J.; Meng, T.; Li, Y.; He, D.; Ran, X.; Chen, G.; Guo, W.; Kan, X.; Fu, S. Polydatin prevents lipopolysaccharide (LPS)-induced Parkinson’s disease via regulation of the AKT/GSK3β-Nrf2/NF-κB signaling axis. Front. Immunol. 2018, 9, 2527. [Google Scholar] [CrossRef]
- Bai, H.; Ding, Y.; Li, X.; Kong, D.; Xin, C.; Yang, X.; Zhang, C.; Rong, Z.; Yao, C.; Lu, S. Polydatin protects SH-SY5Y in models of Parkinson’s disease by promoting Atg5-mediated but parkin-independent autophagy. Neurochem. Int. 2020, 134, 104671. [Google Scholar] [CrossRef]
- Zhang, S.; Wang, S.; Shi, X.; Feng, X. Polydatin alleviates parkinsonism in MPTP-model mice by enhancing glycolysis in dopaminergic neurons. Neurochem. Int. 2020, 139, 104815. [Google Scholar] [CrossRef]
- Schimith, L.E.; Dos Santos, M.G.; Arbo, B.D.; André-Miral, C.; Muccillo-Baisch, A.L.; Hort, M.A. Polydatin as a therapeutic alternative for central nervous system disorders: A systematic review of animal studies. Phytother. Res. PTR 2022, 36, 2852–2877. [Google Scholar] [CrossRef] [PubMed]
- Gao, Y.; Chen, T.; Lei, X.; Li, Y.; Dai, X.; Cao, Y.; Ding, Q.; Lei, X.; Li, T.; Lin, X. Neuroprotective effects of polydatin against mitochondrial-dependent apoptosis in the rat cerebral cortex following ischemia/reperfusion injury. Mol. Med. Rep. 2016, 14, 5481–5488. [Google Scholar] [CrossRef]
- Bonucci, M.; Raggi, R.; Vacca, R.A. Polydatin and its potential protective effect on COVID-19. Clin. Nutr. 2020, 39, 3850–3851. [Google Scholar] [CrossRef] [PubMed]
- Cheng, Y.; Zhang, H.-T.; Sun, L.; Guo, S.; Ouyang, S.; Zhang, Y.; Xu, J. Involvement of cell adhesion molecules in polydatin protection of brain tissues from ischemia–reperfusion injury. Brain Res. 2006, 1110, 193–200. [Google Scholar] [CrossRef] [PubMed]
- Chen, F.; Fang, X.; Zhang, H. Effect of polydatin on expression of p53 and Notch1 in brain tissue of ischemic cerebrovascular disease. J. Biol. Regul. Homeost. Agents 2018, 32, 133–138. [Google Scholar]
- Xu, C.-Y.; Li, S.; Hao, W.; Zhang, R.-L. Effect of polydatin on expression of CDK5 in the prefrontal cortex of rats with chronic alcoholism. Zhongguo Ying Yong Sheng Li Xue Za Zhi Chin. J. Appl. Physiol. 2012, 28, 158–188. [Google Scholar]
- Sun, J.; Qu, Y.; He, H.; Fan, X.; Qin, Y.; Mao, W.; Xu, L. Protective effect of polydatin on learning and memory impairments in neonatal rats with hypoxic-ischemic brain injury by up-regulating brain-derived neurotrophic factor. Mol. Med. Rep. 2014, 10, 3047–3051. [Google Scholar] [CrossRef][Green Version]
- Chen, M.; Hou, Y.; Lin, D. Polydatin protects bone marrow stem cells against oxidative injury: Involvement of Nrf 2/ARE pathways. Stem Cells Int. 2016, 2016, 9394150. [Google Scholar] [CrossRef]
- Zhan, J.; Li, X.; Luo, D.; Yan, W.; Hou, Y.; Hou, Y.; Chen, S.; Luan, J.; Zhang, Q.; Lin, D. Polydatin Attenuates OGD/R-Induced Neuronal Injury and Spinal Cord Ischemia/Reperfusion Injury by Protecting Mitochondrial Function via Nrf2/ARE Signaling Pathway. Oxidative Med. Cell. Longev. 2021, 2021, 6687212. [Google Scholar] [CrossRef]
- Zhan, J.; Li, X.; Luo, D.; Hou, Y.; Hou, Y.; Chen, S.; Xiao, Z.; Luan, J.; Lin, D. Polydatin promotes the neuronal differentiation of bone marrow mesenchymal stem cells in vitro and in vivo: Involvement of Nrf2 signalling pathway. J. Cell. Mol. Med. 2020, 24, 5317–5329. [Google Scholar] [CrossRef]
- Liu, H.; Zhang, G.; Bie, X.; Liu, M.; Yang, J.; Wan, H.; Zhang, Y. Effect of polydatin on dynamic changes of excitatory amino acids in cerebrospinal fluid of cerebral hemorrhage rats. Zhongguo Zhong Yao Za Zhi China J. Chin. Mater. Med. 2010, 35, 3038–3042. [Google Scholar]
- Maoka, T.; Yasui, H.; Ohmori, A.; Tokuda, H.; Suzuki, N.; Osawa, A.; Shindo, K.; Ishibashi, T. Anti-oxidative, anti-tumor-promoting, and anti-carcinogenic activities of adonirubin and adonixanthin. J. Oleo Sci. 2012, 62, 181–186. [Google Scholar] [CrossRef] [PubMed]
- Wiwanitkit, V. Polydatin and COVID-19. Clin. Nutr. 2021. [Google Scholar] [CrossRef] [PubMed]
- Chen, Z.-q.; Sun, X.-h.; Li, X.-j.; Xu, Z.-c.; Yang, Y.; Lin, Z.-y.; Xiao, H.-m.; Zhang, M.; Quan, S.-j.; Huang, H.-q. Polydatin attenuates renal fibrosis in diabetic mice through regulating the Cx32-Nox4 signaling pathway. Acta Pharmacol. Sin. 2020, 41, 1587–1596. [Google Scholar] [CrossRef] [PubMed]
- Gu, L.; Liu, J.; Xu, D.; Lu, Y. Polydatin prevents LPS-induced acute kidney injury through inhibiting inflammatory and oxidative responses. Microb. Pathog. 2019, 137, 103688. [Google Scholar] [CrossRef]
- Gao, Y.; Dai, X.; Li, Y.; Li, G.; Lin, X.; Ai, C.; Cao, Y.; Li, T.; Lin, B. Role of Parkin-mediated mitophagy in the protective effect of polydatin in sepsis-induced acute kidney injury. J. Transl. Med. 2020, 18, 114. [Google Scholar] [CrossRef]
- Han, B.; Gong, M.; Li, Z.; Qiu, Y.; Zou, Z. NMR-based metabonomic study reveals intervention effects of polydatin on potassium oxonate-induced hyperuricemia in rats. Oxidative Med. Cell. Longev. 2020, 2020, 6943860. [Google Scholar] [CrossRef]
- Sun, Z.; Wang, X.; Xu, Z. SIRT1 provides new pharmacological targets for polydatin through its role as a metabolic sensor. Biomed. Pharmacother. 2021, 139, 111549. [Google Scholar] [CrossRef]
- Gao, Y.; Zeng, Z.; Li, T.; Xu, S.; Wang, X.; Chen, Z.; Lin, C. Polydatin inhibits mitochondrial dysfunction in the renal tubular epithelial cells of a rat model of sepsis-induced acute kidney injury. Anesth. Analg. 2015, 121, 1251–1260. [Google Scholar] [CrossRef]
- Zeng, Z.; Chen, Z.; Xu, S.; Zhang, Q.; Wang, X.; Gao, Y.; Zhao, K.-s. Polydatin protecting kidneys against hemorrhagic shock-induced mitochondrial dysfunction via SIRT1 activation and p53 deacetylation. Oxidative Med. Cell. Longev. 2016, 2016, 1737185. [Google Scholar] [CrossRef]
- Gu, T.-T.; Zhang, D.-M.; Wan, Z.-Y.; Li, T.-S.; Jiao, R.-Q.; Chen, T.-Y.; Zhao, X.-J.; Kong, L.-D. Polydatin enhances glomerular podocyte autophagy homeostasis by improving Nrf2-dependent antioxidant capacity in fructose-fed rats. Mol. Cell. Endocrinol. 2021, 520, 111079. [Google Scholar] [CrossRef] [PubMed]
- Cao, K.; Lei, X.; Liu, H.; Zhao, H.; Guo, J.; Chen, Y.; Xu, Y.; Cheng, Y.; Liu, C.; Cui, J. Polydatin alleviated radiation-induced lung injury through activation of Sirt3 and inhibition of epithelial–mesenchymal transition. J. Cell. Mol. Med. 2017, 21, 3264–3276. [Google Scholar] [CrossRef] [PubMed]
- Hanna, D.A.; Khalaf, M.M.; Abo-Saif, A.A. Polydatin protects against ovalbumin-induced bronchial asthma in rats; involvement of urocortin and surfactant-D expression. Immunopharmacol. Immunotoxicol. 2019, 41, 403–412. [Google Scholar] [CrossRef] [PubMed]
- Zeng, H.; Wang, Y.; Gu, Y.; Wang, J.; Zhang, H.; Gao, H.; Jin, Q.; Zhao, L. Polydatin attenuates reactive oxygen species-induced airway remodeling by promoting Nrf2-mediated antioxidant signaling in asthma mouse model. Life Sci. 2019, 218, 25–30. [Google Scholar] [CrossRef]
- Liu, Y.-L.; Chen, B.-Y.; Nie, J.; Zhao, G.-H.; Zhuo, J.-Y.; Yuan, J.; Li, Y.-C.; Wang, L.-L.; Chen, Z.-W. Polydatin prevents bleomycin-induced pulmonary fibrosis by inhibiting the TGF-β/Smad/ERK signaling pathway. Exp. Ther. Med. 2020, 20, 1. [Google Scholar] [CrossRef]
- Jin, X.-F.; Xu, Z.-J.; Wang, W.-T.; Xu, Y.-X.; Zhang, X.-L. The regulative effects of polydatin on toll-like receptor 4 signal transduction pathway in lung ischemia/reperfusion injury in rabbits. Zhongguo Ying Yong Sheng Li Xue Za Zhi Chin. J. Appl. Physiol. 2009, 25, 41–44. [Google Scholar]
- Wang, F.-Y.; Xu, Z.-J.; Zhang, X.-L.; Wang, W.-T.; Ha, M.-L.; Wang, Y. Protective effects of polydatin against lung ischemia/reperfusion injury and the initial exploration for its mechanism. Zhongguo Ying Yong Sheng Li Xue Za Zhi Chin. J. Appl. Physiol. 2008, 24, 62–65. [Google Scholar]
- Li, T.; Liu, Y.; Li, G.; Wang, X.; Zeng, Z.; Cai, S.; Li, F.; Chen, Z. Polydatin attenuates ipopolysaccharide-induced acute lung injury in rats. Int. J. Clin. Exp. Pathol. 2014, 7, 8401. [Google Scholar]
- Li, T.; Cai, S.; Zeng, Z.; Zhang, J.; Gao, Y.; Wang, X.; Chen, Z. Protective effect of polydatin against burn-induced lung injury in rats. Respir. Care 2014, 59, 1412–1421. [Google Scholar] [CrossRef]
- Matacchione, G.; Gurău, F.; Silvestrini, A.; Tiboni, M.; Mancini, L.; Valli, D.; Rippo, M.R.; Recchioni, R.; Marcheselli, F.; Carnevali, O.; et al. Anti-SASP and anti-inflammatory activity of resveratrol, curcumin and β-caryophyllene association on human endothelial and monocytic cells. Biogerontology 2021, 22, 297–313. [Google Scholar] [CrossRef]
- Jiang, Q.; Yi, M.; Guo, Q.; Wang, C.; Wang, H.; Meng, S.; Liu, C.; Fu, Y.; Ji, H.; Chen, T. Protective effects of polydatin on lipopolysaccharide-induced acute lung injury through TLR4-MyD88-NF-κB pathway. Int. Immunopharmacol. 2015, 29, 370–376. [Google Scholar] [CrossRef] [PubMed]
- Shu, S.; Wang, X.; Ling, Z.; Lu, Z. Effect of polydatin on phospholipase A2 in lung tissues in rats with endotoxic shock. Chin. J. Traumatol. 2004, 7, 239–243. [Google Scholar] [PubMed]
- Zhang, Y.; Chen, L.; Liang, B. Effects of polydatin on bleomycin-induced pulmonary fibrosis in rats. Zhongguo Zhong Yao Za Zhi China J. Chin. Mater. Med. 2011, 36, 3528–3534. [Google Scholar]
- Fu, Y.; Yan, M.; Xie, C.; Hu, J.; Zeng, X.; Hu, Q. Polydatin relieves paraquat-induced human MRC-5 fibroblast injury through inhibiting the activation of the NLRP3 inflammasome. Ann. Transl. Med. 2020, 8, 765. [Google Scholar] [CrossRef]
- Li, B.; Wang, X.L. Effective treatment of polydatin weakens the symptoms of collagen-induced arthritis in mice through its anti-oxidative and anti-inflammatory effects and the activation of MMP-9. Mol. Med. Rep. 2016, 14, 5357–5362. [Google Scholar] [CrossRef][Green Version]
- Liao, P.; He, Y.; Yang, F.; Luo, G.; Zhuang, J.; Zhai, Z.; Zhuang, L.; Lin, Z.; Zheng, J.; Sun, E. Polydatin effectively attenuates disease activity in lupus-prone mouse models by blocking ROS-mediated NET formation. Arthritis Res. Ther. 2018, 20, 1–11. [Google Scholar] [CrossRef]
- Yang, F.; Luo, X.; Luo, G.; Zhai, Z.; Zhuang, J.; He, J.; Han, J.; Zhang, Y.; Zhuang, L.; Sun, E. Inhibition of NET formation by polydatin protects against collagen-induced arthritis. Int. Immunopharmacol. 2019, 77, 105919. [Google Scholar] [CrossRef]
- Zhou, Q.L.; Qin, R.Z.; Yang, Y.X.; Huang, K.B.; Yang, X.W. Polydatin possesses notable anti-osteoporotic activity via regulation of OPG, RANKL and β-catenin. Mol. Med. Rep. 2016, 14, 1865–1869. [Google Scholar] [CrossRef]
- Shen, Y.-S.; Chen, X.-J.; Wuri, S.-N.; Yang, F.; Pang, F.-X.; Xu, L.-L.; He, W.; Wei, Q.-S. Polydatin improves osteogenic differentiation of human bone mesenchymal stem cells by stimulating TAZ expression via BMP2-Wnt/β-catenin signaling pathway. Stem Cell Res. Ther. 2020, 11, 204. [Google Scholar] [CrossRef]
- Di Benedetto, A.; Posa, F.; De Maria, S.; Ravagnan, G.; Ballini, A.; Porro, C.; Trotta, T.; Grano, M.; Muzio, L.L.; Mori, G. Polydatin, natural precursor of resveratrol, promotes osteogenic differentiation of mesenchymal stem cells. Int. J. Med. Sci. 2018, 15, 944. [Google Scholar] [CrossRef]
- Chen, Z.; Wei, Q.; Hong, G.; Chen, D.; Liang, J.; He, W.; Chen, M.H. Polydatin induces bone marrow stromal cells migration by activation of ERK1/2. Biomed. Pharmacother. 2016, 82, 49–53. [Google Scholar] [CrossRef] [PubMed]
- Ma, C.; Wen, B.; Zhang, Q.; Shao, P.; Gu, W.; Qu, K.; Shi, Y.; Wang, B. Polydatin regulates the apoptosis and autophagy of fibroblasts obtained from patients with ankylosing spondylitis. Biol. Pharm. Bull. 2018, 42, 50–56. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Huang, C.; Lin, Z.; Pan, X.; Chen, J.; Zheng, G.; Tian, N.; Yan, Y.; Zhang, Z.; Hu, J. Polydatin suppresses nucleus pulposus cell senescence, promotes matrix homeostasis and attenuates intervertebral disc degeneration in rats. J. Cell. Mol. Med. 2018, 22, 5720–5731. [Google Scholar] [CrossRef] [PubMed]
- Indraccolo, U.; Indraccolo, S.R.; Mignini, F. Micronized palmitoylethanolamide/trans-polydatin treatment of endometriosis-related pain: A meta-analysis. Ann. Dell’istituto Super. Sanita 2017, 53, 125–134. [Google Scholar]
- Indraccolo, U.; Barbieri, F. Effect of palmitoylethanolamide–polydatin combination on chronic pelvic pain associated with endometriosis: Preliminary observations. Eur. J. Obstet. Gynecol. Reprod. Biol. 2010, 150, 76–79. [Google Scholar] [CrossRef]
- Di Paola, R.; Fusco, R.; Gugliandolo, E.; Crupi, R.; Evangelista, M.; Granese, R.; Cuzzocrea, S. Co-micronized palmitoylethanolamide/polydatin treatment causes endometriotic lesion regression in a rodent model of surgically induced endometriosis. Front. Pharmacol. 2016, 7, 382. [Google Scholar] [CrossRef]
- Loi, E.S.; Pontis, A.; Cofelice, V.; Pirarba, S.; Fais, M.F.; Daniilidis, A.; Melis, I.; Paoletti, A.M.; Angioni, S. Effect of ultramicronized-palmitoylethanolamide and co-micronized palmitoylethanolamide/polydatin on chronic pelvic pain and quality of life in endometriosis patients: An open-label pilot study. Int. J. Women’s Health 2019, 11, 443. [Google Scholar]
- Sheng, C.; Yu, Y.-h.; Zhao, K.-s.; Qin, W.; Wang, C.-h. Hypotensive resuscitation combined with polydatin improve microcirculation and survival in a rabbit model of uncontrolled hemorrhagic shock in pregnancy. J. Surg. Res. 2011, 168, 103–110. [Google Scholar] [CrossRef]
- Ou, C.; Song, H.; Zhou, Y.; Peng, J.; Peng, Q. Exploring the Molecular Mechanism of Qing Guang An Granule in Treating Glaucoma Using Network Pharmacology and Molecular Docking. Evid. Based Complement. Altern. Med. Ecam 2020, 2020, 8824150. [Google Scholar] [CrossRef]
- Ye, J.; Piao, H.; Jiang, J.; Jin, G.; Zheng, M.; Yang, J.; Jin, X.; Sun, T.; Choi, Y.H.; Li, L. Polydatin inhibits mast cell-mediated allergic inflammation by targeting PI3K/Akt, MAPK, NF-κB and Nrf2/HO-1 pathways. Sci. Rep. 2017, 7, 11895. [Google Scholar] [CrossRef]
- Yuan, M.; Li, J.; Lv, J.; Mo, X.; Yang, C.; Chen, X.; Liu, Z.; Liu, J. Polydatin (PD) inhibits IgE-mediated passive cutaneous anaphylaxis in mice by stabilizing mast cells through modulating Ca2+ mobilization. Toxicol. Appl. Pharmacol. 2012, 264, 462–469. [Google Scholar] [CrossRef] [PubMed]
- Yang, B.; Li, J.-J.; Cao, J.-J.; Yang, C.-B.; Liu, J.; Ji, Q.-M.; Liu, Z.-G. Polydatin attenuated food allergy via store-operated calcium channels in mast cell. World J. Gastroenterol. WJG 2013, 19, 3980. [Google Scholar] [CrossRef] [PubMed]
- Cordaro, M.; Impellizzeri, D.; Siracusa, R.; Gugliandolo, E.; Fusco, R.; Inferrera, A.; Esposito, E.; Di Paola, R.; Cuzzocrea, S. Effects of a co-micronized composite containing palmitoylethanolamide and polydatin in an experimental model of benign prostatic hyperplasia. Toxicol. Appl. Pharmacol. 2017, 329, 231–240. [Google Scholar] [CrossRef] [PubMed]
- Guan, S.-Y.; Zhang, K.; Wang, X.-S.; Yang, L.; Feng, B.; Tian, D.-D.; Gao, M.-R.; Liu, S.-B.; Liu, A.; Zhao, M.-G. Anxiolytic effects of polydatin through the blockade of neuroinflammation in a chronic pain mouse model. Mol. Pain 2020, 16, 1744806919900717. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Min, J.; Chen, Y.; Li, H.; Zhang, Y. Polydatin attenuates orbital oxidative stress in Graves’ orbitopathy through the NRF2 pathway. Chem. Biol. Interact. 2020, 315, 108894. [Google Scholar] [CrossRef] [PubMed]
- Tang, S.; Tang, Q.; Jin, J.; Zheng, G.; Xu, J.; Huang, W.; Li, X.; Shang, P.; Liu, H. Polydatin inhibits the IL-1β-induced inflammatory response in human osteoarthritic chondrocytes by activating the Nrf2 signaling pathway and ameliorates murine osteoarthritis. Food Funct. 2018, 9, 1701–1712. [Google Scholar] [CrossRef]
- Xu, L.-Q.; Xie, Y.-L.; Gui, S.-H.; Zhang, X.; Mo, Z.-Z.; Sun, C.-Y.; Li, C.-L.; Luo, D.-D.; Zhang, Z.-B.; Su, Z.-R. Polydatin attenuates d-galactose-induced liver and brain damage through its anti-oxidative, anti-inflammatory and anti-apoptotic effects in mice. Food Funct. 2016, 7, 4545–4555. [Google Scholar] [CrossRef]
- Ravagnan, G.; De Filippis, A.; Cartenì, M.; De Maria, S.; Cozza, V.; Petrazzuolo, M.; Tufano, M.A.; Donnarumma, G. Polydatin, a natural precursor of resveratrol, induces β-defensin production and reduces inflammatory response. Inflammation 2013, 36, 26–34. [Google Scholar] [CrossRef]
- Fuggetta, M.P.; Migliorino, M.R.; Ricciardi, S.; Osman, G.; Iacono, D.; Leone, A.; Lombardi, A.; Ravagnan, G.; Greco, S.; Remotti, D. Prophylactic dermatologic treatment of afatinib-induced skin toxicities in patients with metastatic lung cancer: A pilot study. Scientifica 2019, 2019, 9136249. [Google Scholar] [CrossRef]
- Cervigni, M.; Nasta, L.; Schievano, C.; Lampropoulou, N.; Ostardo, E. Micronized palmitoylethanolamide-polydatin reduces the painful symptomatology in patients with interstitial cystitis/bladder pain syndrome. BioMed Res. Int. 2019, 2019, 9828397. [Google Scholar] [CrossRef]
- Chen, L.; Chen, Z.; Xu, Z.; Feng, W.; Yang, X.; Qi, Z. Polydatin protects Schwann cells from methylglyoxal induced cytotoxicity and promotes crushed sciatic nerves regeneration of diabetic rats. Phytother. Res. 2021, 35, 4592–4604. [Google Scholar] [CrossRef] [PubMed]
- Soave, I.; Marci, R. Administration of micronized palmitoylethanolamide (PEA)-transpolydatin in the treatment of chronic pelvic pain in women affected by endometriosis: Preliminary results. Minerva Ginecol. 2013, 65, 453–463. [Google Scholar]
- Cremon, C.; Stanghellini, V.; Barbaro, M.; Cogliandro, R.; Bellacosa, L.; Santos, J.; Vicario, M.; Pigrau, M.; Alonso Cotoner, C.; Lobo, B. Randomised clinical trial: The analgesic properties of dietary supplementation with palmitoylethanolamide and polydatin in irritable bowel syndrome. Aliment. Pharmacol. Ther. 2017, 45, 909–922. [Google Scholar] [CrossRef] [PubMed]
- Pace, M.C.; Passavanti, M.B.; Aurilio, C.; Sansone, P.; Aurilio, R.; De Maria, S.; Lama, S.; Federico, A.; Ravagnan, G.; Caraglia, M. Polydatin administration improves serum biochemical parameters and oxidative stress markers during chronic alcoholism: A pilot study. Vivo 2015, 29, 405–408. [Google Scholar]
- Biswas, P.; Dellanoce, C.; Vezzoli, A.; Mrakic-Sposta, S.; Malnati, M.; Beretta, A.; Accinni, R. Antioxidant Activity with Increased Endogenous Levels of Vitamin C, E and A Following Dietary Supplementation with a Combination of Glutathione and Resveratrol Precursors. Nutrients 2020, 12, 3224. [Google Scholar] [CrossRef] [PubMed]
- Bavetta, M.; Silvaggio, D.; Campione, E.; Sollena, P.; Formica, V.; Coletta, D.; Graziani, G.; Romano MC, P.; Roselli, M.; Peris, K. The Effects of Association of Topical Polydatin Improves the Preemptive Systemic Treatment on EGFR Inhibitors Cutaneous Adverse Reactions. J. Clin. Med. 2021, 10, 466. [Google Scholar] [CrossRef]
- Mostafa, F.; Abdel-Moneim, A.; Abdul-Hamid, M.; Galaly, S.R.; Mohamed, H.M. Polydatin and polydatin-loaded chitosan nanoparticles attenuate diabetic cardiomyopathy in rats. J. Mol. Histol. 2021, 52, 135–152. [Google Scholar] [CrossRef]
- Wang, X.; Guan, Q.; Chen, W.; Hu, X.; Li, L. Novel nanoliposomal delivery system for polydatin: Preparation, characterization, and in vivo evaluation. Drug Des. Dev. Ther. 2015, 9, 1805. [Google Scholar]
- Lin, L.; Gong, H.; Li, R.; Huang, J.; Cai, M.; Lan, T.; Huang, W.; Guo, Y.; Zhou, Z.; An, Y. Nanodrug with ROS and pH Dual-Sensitivity Ameliorates Liver Fibrosis via Multicellular Regulation. Adv. Sci. 2020, 7, 1903138. [Google Scholar] [CrossRef]
- Xiao, J.; Wang, F.; Liu, J.; Wang, L.; Kai, G.; Yu, X. Effect of ZnO# ZnS QDs heterojunctures on the stilbenes–plasma proteins interactions. Mol. BioSystems 2011, 7, 2452–2458. [Google Scholar]
- Basta-Kaim, A.; Ślusarczyk, J.; Szczepanowicz, K.; Warszyński, P.; Leśkiewicz, M.; Regulska, M.; Trojan, E.; Lasoń, W. Protective effects of polydatin in free and nanocapsulated form on changes caused by lipopolysaccharide in hippocampal organotypic cultures. Pharmacol. Rep. 2019, 71, 603–613. [Google Scholar] [CrossRef] [PubMed]
- Abd El-Hameed, A.M.; Yousef, A.I.; Abd El-Twab, S.M.; El-Shahawy, A.A.; Abdel-Moneim, A. Hepatoprotective Effects of Polydatin-Loaded Chitosan Nanoparticles in Diabetic Rats: Modulation of Glucose Metabolism, Oxidative Stress, and Inflammation Biomarkers. Biochemistry 2021, 86, 179–189. [Google Scholar] [CrossRef] [PubMed]
- Abdel-Moneim, A.; El-Shahawy, A.; Yousef, A.I.; Abd El-Twab, S.M.; Elden, Z.E.; Taha, M. Novel polydatin-loaded chitosan nanoparticles for safe and efficient type 2 diabetes therapy: In silico, in vitro and in vivo approaches. Int. J. Biol. Macromol. 2020, 154, 1496–1504. [Google Scholar] [CrossRef]
- Abd El-Hameed, A.M. Polydatin-loaded chitosan nanoparticles ameliorates early diabetic nephropathy by attenuating oxidative stress and inflammatory responses in streptozotocin-induced diabetic rat. J. Diabetes Metab. Disord. 2020, 19, 1599. [Google Scholar] [CrossRef] [PubMed]
- Vijayalakshmi, S.; Mariadoss AV, A.; Ramachandran, V.; Shalini, V.; Agilan, B.; Sangeetha, C.C.; Balu, P.; Kotakadi, V.S.; Karthikkumar, V.; Ernest, D. Polydatin encapsulated poly [lactic-co-glycolic acid] nanoformulation counteract the 7, 12-dimethylbenz [a] anthracene mediated experimental carcinogenesis through the inhibition of cell proliferation. Antioxidants 2019, 8, 375. [Google Scholar] [CrossRef]
- Samra, Y.A.; Abdelghany, A.M.; Zaghloul, R.A. Polydatin gold nanoparticles potentiate antitumor effect of doxorubicin in Ehrlich ascites carcinoma-bearing mice. J. Biochem. Mol. Toxicol. 2021, 35, e22869. [Google Scholar] [CrossRef]
- Orlowski, P.; Zmigrodzka, M.; Tomaszewska, E.; Ranoszek-Soliwoda, K.; Pajak, B.; Slonska, A.; Cymerys, J.; Celichowski, G.; Grobelny, J.; Krzyzowska, M. Polyphenol-conjugated bimetallic Au@ AgNPs for improved wound healing. Int. J. Nanomed. 2020, 15, 4969. [Google Scholar] [CrossRef]
- Magalhães, P.R.; Reis, P.B.; Vila-Viçosa, D.; Machuqueiro, M.; Victor, B.L. Identification of Pan-Assay INterference compoundS (PAINS) Using an MD-Based Protocol. In Computational Design of Membrane Proteins; Springer: Berlin/Heidelberg, Germany, 2021; pp. 263–271. [Google Scholar]
- de Matos, A.M.; Blázquez-Sánchez, M.T.; Sousa, C.; Oliveira, M.C.; de Almeida, R.F.; Rauter, A.P. C-Glucosylation as a tool for the prevention of PAINS-induced membrane dipole potential alterations. Sci. Rep. 2021, 11, 4443. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Study Type | Biological Effects | Cell Lines, Animal Types | Doses | References |
|---|---|---|---|---|
| In vivo, in vitro | ⇅miR-382/PD-L1 axis activity, ┬cell proliferation, ↑cleaved caspase-3 and cleaved caspase-9 expression | Mice, colorectal cancer cell line (Caco-2, HCT 116, HEK293) | 150 mg/kg/day for 16 days, injection into tumor, 50, 100, 150 μM | [88] |
| In vivo, in vitro | ↑BMP signaling pathway activity, radiosensitivity, ┬proliferation | Mice, colon tumor cell line (CT26, HCT116) | 25 mg/kg i.p. once a week for 4 weeks, 0, 40, 60, 80, 100, and 120 μM | [89] |
| In vitro | ↑Bax and cyclin A expression, ↓cyclin D1 and Bc1-2 expression, ┬growth, ↑arresting cells in S phase | Human acute monocytic leukemia cell line (THP-1) | 10, 20, 40, 60, 80, 100, 120, 140, 160 μM for 24 and 48 h | [90] |
| In vitro | ↑Bcl-2-associated x expression, ↓cyclin D1 and cyclin B1 expression | Leukemia cell line (MOLT-4) | 0.5, 1, 2, 4, 10, or 20 μM | [86] |
| In vivo, in vitro | ┬PDGF/Akt signaling pathway activity, cell proliferation, ↓Bcl-2 expression, ↑Bax expression | Mice, human laryngeal cancer cell line (AMC-HN-8, HeLa Hep-2) | 50 mg/kg 3 times a week for 3 weeks, 2, 4, 6 μM for 24, 48, 72 h | [91] |
| In vitro | ↑ROS level, ER stress, cytosolic cytochrome c level, ↓mitochondrial cytochrome C level, Akt phosphorylation | Human nasopharyngeal carcinoma cell line (CNE), cervical carcinoma cell line (HeLa), hepatoma cell line (SMMC-7721), epidermal carcinoma cell line (A-431) | - | [92] |
| In vitro | ↓phosphorylated Her-2 and EGFR levels, ERK expression, secretion of VEGF | Human ovarian adenocarcinoma cell line (SKOV-3, OVCAR-8) | 5, 10, 50, 100 μM for 6 days | [93] |
| In vitro | ↓phosphor-NF-κB p6 expression, activation of NF-κB pathway, ┬NLRP3 inflammasome activation, proliferation and migration of NSCLC cells | NSCLC cell line (A549, H1299) | 25, 50, 100 μM for 24 h | [94] |
| In vitro | ↑Bax expression, ↓Bcl-2 and cyclin D1 expression, ⇅mTOR pathway activity | Lung cancer cell line (A549) | 2, 4, 6, 8 μM for 24 and 48 h | [95] |
| In vivo, in vitro | ┬EGFR-Akt/ERK1/2/STAT3-SOX2/Snail signaling pathway activity, ┬proliferation, migration, invasion, stemness, ↑Bax expression, ↓Bcl-2 and cyclin D1 expression | Mice, lung cancer cell lines (NCI-H1975, A549), breast cancer cell lines (MDA-MB-231, MCF-7), cervical cancer cell line (HeLa), ovarian cancer cell line (SKOV-3), liver cancer cell line (SMMC-7721), nasopharyngeal cancer cell line (CNE-1), leukemia cell line (HL-60, K562) | 50 mg/kg/day for 14 days i.p. injection, 0–8 μM for 24 and 48 h | [96] |
| In vitro | ↓phosphorylation of mTOR and p70s6k | Human multiple myeloma cell line (RPMI 8226) | 0, 5, 10, 25, 50, 100, 200, and 400 μM for 24 and 48 h | [98] |
| In vitro | ┬PI3K/Akt/mTOR signaling pathway activity and downstream gene expression | Human cervical cancer cell line (HeLa cells) | 50, 100, 150 μM | [99] |
| In vivo, in vitro | ┬c-Myc expression, ⇅cell cycle-related protein expression (p21, p27, CDK2, CDK4, Cyclin D1, Cyclin E1, EMT, markers (E-cadherin, N-cadherin, Snail and Slug)), ┬proliferation and metastasis | Mice, human cervical cancer cell lines (CaSki, C33A), human embryonic renal cell line (293FT) | 100 mg/kg/day injection for 3 weeks, 0.1, 10, 100, 500 μM for 24 and 48 h | [100] |
| In vivo, in vitro | ┬G6PD activity, ↑ROS level, ER stress | Mice, head, and neck squamous cell carcinoma (HNSCC), breast cancer cell line (MCF-7) | 100 mg/kg 3 times a week for 30 days, 2–100 μM for 24 or 48 h | [1] |
| In vitro | ┬G6PD activity, ↑autophagosomes formation | Breast cell lines (MCF-7) | 10, 20, 30 μM for 24 h | [21] |
| In vitro | ↓phosphorylated CREB level, cyclin D1 expression, ↑S-phase cell cycle arrest | Breast cancer cell lines (MDA-MB-231, MCF-7) | 0–8 μM for 24 and 48 h | [101] |
| In vivo, in vitro | ┬ROS/PI3K/Akt/HIF-1α/HK2 signaling axis activity | Mice, breast cell lines (4T1, MCF-7) | 100 mg/kg i.p. injection every other day for 3 weeks, 2 μM | [102] |
| In vitro | ┬Akt activation, ⇅Ki67, p21, cyclin A, cyclin E, CDK2, MMP-2, MMP-9, tissue metalloproteinase-1, PPAR 1, caspase-3, p-glycoprotein 1, lung resistance-related protein 1, growth arrest, and DNA damage-45α, glutathione S-transferase π and heat shock protein 27 expression, ↑chemosensitivity to paclitaxel | Human osteosarcoma cell lines (U-2OS, MG-63) | 0–400 μM for 24, 48, 72 h | [103] |
| In vitro | ┬STAT3 signaling pathway activity, ↑Atg12, Atg14, BECN, and PIC3K3 expression, autophagic cell death | Human normal osteoblast cell (hFOB1.19), human osteosarcoma cell line (MG-63) | 0–160 μM for 12–72 h | [104] |
| In vitro | ┬β-catenin signaling pathway activity, ↓cell proliferation, ↑Bax expression, ↓Bcl-2 expression, caspase-3 activity | Osteosarcoma cell lines (143B, MG63) | 0, 1, 10, 30, 100 μM for 24, 48, 72 h | [105] |
| In vivo, in vitro | ┬TUG1/Akt signaling pathway activity | Mice, osteosarcoma cell lines (Saos-2, MG-63), normal osteoblast cell line (hFOB), doxorubicin-resistant cell lines (Saos-2/Dox, MG- 63/Dox) | 150 mg/Kg/day i.p. injection for 20 days, 50, 100, 150, 200, 250 μM 24, 48, 72 h | [106] |
| In vivo, in vitro | ↑caspase-3 activity, caspase-3, caspase-9 and Bax expression, TUNEL activity, ↓expression of Bcl-2, Ki-67, ┬Wnt/beta-catenin signaling activity | Mice, normal liver cell lines (HL-7702, HCC), liver cancer cell lines (HepG2, SMMC-7721) | 25, 50, 100 mg/kg/day i.p. injection for 20 days, 1, 3, 10, 30, 100 μM | [107] |
| In vitro | ┬Akt/STAT3–FOXO1 signaling pathway activity | Human hepatocellular carcinoma cell line (HCCLM3), normal hepatic cell line LO2 | 0–800 μM for 24–72 h | [108] |
| Study Type | Biological Effects | Cell Lines, Animal Types | Doses | References |
|---|---|---|---|---|
| In vivo, in vitro | ↑Nrf2/HO-1 signaling activity, ↓ROS levels, hypoxia, AMI-induced myocardial damage | Rat, rat embryonic cardiomyocyte cells | 100 mg/kg/day i.p. injection for 30 days, 0, 10, 100, 250, 500 μM for 12 h | [114] |
| In vitro | ↑eNOS/Sirt1 pathway activity, ↓proliferation of VSMCs, ROS | vascular smooth muscle cells (VSMCs) | 10, 50,100 μM for 24 h | [115] |
| In vivo | ⇅NO, Ang II and ET expression, ┬protein kinase C activity | Rat | 5, 10, 20 mg/kg every other day i.p. injection for 3 weeks | [116] |
| In vivo | ↓ICAM-1, V-CAM, TNF-α, IL-1β, iNOS, PARP, and NF-κB expression, Bax and FAS ligand activity, vascular damage | Mice | 30 mg/kg orally for 14 days | [117] |
| In vivo, in vitro | ↑Sirt6-mediated autophagy and Bcl-2 expression, ↓serum TNF-α, IL-1β, IL-6, caspase-3, and Bax expression | Rat, cardiomyoblast cell line (H9c2) | 30 mg/kg i.p. injection once, 100 μM for 30 min | [118] |
| In vivo | ↑reverse cholesterol transport | Mice | 50, 100 mg/kg/day orally for 12 weeks | [127] |
| In vivo | ↓ROS levels, spark-mediated sarcoplasmic reticulum leak | Rat | 10 mg/kg i.v. injection once | [135] |
| In vivo | ↓size of cardiomyocyte, aldosterone, TNF-α, Ang II, and endothelin-1 expression, blood pressure | Rat, mice | 60, 120 mg/kg/day orally for 30 days | [119] |
| In vivo | ↓renin–angiotensin–aldosterone system activity, ┬peroxidation | Rat | 25, 50, 100 mg/kg/day orally for 7 weeks | [120] |
| In vivo, in vitro | ┬NADPH oxidase activity, superoxide production, ↓Ang II-mediated cardiac hypertrophy | Rat, ventricle cells of neonatal rats | 50 mg/kg/day orally for 7 days, 5–50 μM for 60 min | [121] |
| In vitro, in vivo | ↑Sirt3 expression, autophagy flux | Mice, cardiomyocytes of neonates, and adult mice | 7.5 mg/kg/day injection for 28 days | [122] |
| In vivo | ↓NOX4, NOX2, NF-κB expression, NADPH oxidase activity, ROS level, inflammatory cytokines | Rat, embryonic rat cardiac cell line H9c2 | 100 mg/kg/day orally for 8 weeks | [35] |
| In vivo | ↓myocardial superoxide generation, MDA level, and iNOS expression, NO metabolite levels, nitrotyrosine, ↑Notch1/Hes1-Pten/Akt signaling pathway activity, eNOS phosphorylation | Rat | 20 mg/kg/day orally for 4 days | [36,66,123] |
| In vivo | ↑PPARβ activation, ↓NF-κB p65, COX-2, and iNOS expression | Mice | 50, 100 mg/kg/day | [124] |
| In vivo | ↑LDL-C/HDL-C, TC/HDL-C ratios, ↓serum TC, TG, LDL-C, hepatic TG levels | Hamsters | 25, 50 and 100 mg/kg/day orally for 15 days | [125] |
| In vivo | ↓TC, TG, LDL-C, TC/HDL-C | Rabbit | 25, 50, 100 mg/kg/day orally for 3 weeks | [126] |
| In vitro | ↑eNOS/Sirt1 pathway activity, Kip1/p27 expression, ↓VSMCs proliferation, ROS production | VSMCs of the mouse thoracic aorta | 10, 50,100 μM for 24 h | [115] |
| In vivo, in vitro | ↓PBEF-inducing cholesterol deposition in macrophages | Mice, peritoneal macrophages of mouse | 100 mg/kg/day i.p. injection for 12 weeks, 50, 100, 200 μg/mL for 2 h | [128] |
| In vivo, in vitro | ↓TC, TG, LDL-C, PPARγ expression, MCP-1 and TNF-α expression, ↑HDL, leptin expression | Mice, mouse 3T3-L1 preadipocytes | 100 mg/kg/d orally for 4 weeks, 20 μM for 2 days | [130] |
| In vitro | ⇅PPARγ signaling pathways | Mouse peritoneal macrophages | 8.9 μg/mL | [131] |
| In vivo | ↑PKC--dependent signaling activity, ↓CPK and LDH leakage, ⇅SOD and MDA content | Rat | 20 μg/kg, i.v. once | [71] |
| In vitro | ↑Bcl-2 expression, ↓Bax expression, ↓I/R-induced apoptosis | Rat heart | 50 μM for 10 min | [132] |
| In vivo, in vitro | ┬RAS and the downstream ROCK pathway activity | Rat, ventricular myocytes of rats | 50 mg/kg/day orally for 8 days, 30, 50 μM for 1 h | [22] |
| In vivo, in vitro | ↑SOD activity, NO, NOS activity, cNOS, ↓MDA content | Rat, hearts of rats | 0.2 mL/100 g body weight (0.1% polydatin solution) i.v. injection before model establishment, 0.05 μM 15 min before model establishment | [133] |
| In vivo, in vitro | ↑Sirt3 activity, ↑mitochondrial biogenesis, upregulation of autophagy | Mice, ventricular cardiomyocytes | 7.5 mg/kg/day i.p. injection for 10 days, 10 μM | [2] |
| Study Type | Biological Effects | Animal Types, Cell Lines | Doses | References |
|---|---|---|---|---|
| In vitro, in vivo | ↓CYP2Y3, CYP3A65, HMGCRa, HMGCRb, FASN, lipid and ethanol metabolism, hepatic fat accumulation, oxidative stress, DNA damage↑CHOP and GADD45aa expression | Zebrafish larvae | 6.25, 12.5, 25 μg/mL for 48 h | [153] |
| In vivo | ⇅TLR4 and NF-κB activity, ↓TNF-α, IL-1β, IL-6, CYP2E1, and ROS levels, ↑Nrf2/HO-1, SOD activity, GSH-Px, CAT, ADH, and ALDH activity | Rat | 25, 50, 100 mg/kg/day orally for 7 days | [48,155] |
| In vivo | ↓lipid peroxidation, TNF-α expression, lipogenesis | Rat | 30, 90 mg/kg/day orally for 8 weeks | [156] |
| In vivo, in vitro | ┬mTOR signaling activity, ↑TFEB expression, autophagic flux | Mice, human hepatocyte cell line (LO2) | 100 mg/kg every other day orally for 4 weeks, 24 μM for 24 h | [154] |
| In vivo, in vitro | ↓NOX4 enzymes, oxidative stress, CD68, FAS, SREBP-1c, LDH, cleaved caspase-3, annexin V, ALT, AST levels, ┬TLR-4/NF-κB p65 signaling pathway activity, | Mice, human hepatocellular carcinoma cell line (HepG2) | 5 mg/kg, every other day, i.p. injection for 4 weeks, 5, 10, 20 μM for 24 h | [157] |
| In vivo, in vitro | ↑Sirt1/Nrf2 pathway activity, ↓serum aminotransferase levels, hepatic pathological damage | Mice, human hepatocyte cell line (L02) | 100, 200, 400 mg/kg/day orally for 7 days, 50 μM for 24 h | [158] |
| In vivo | ↓4-HNE production, NOX4 expression | Mice | 5 mg/kg i.p. injection | [159] |
| In vivo | ↓AST, ALT, MDA, TNF-α, IL-1β, COX-2, iNOs, NF-κB, ↑GSH content, GST, SOD, CAT, and GSH-Px activity, hepatic TGF-b1 expression | Mice | 25, 50, 100 mg/kg/day orally for 5 days | [70] |
| In vivo | ┬NF-κB activation, ↓TNF-α expression, MPO activity, ICAM-l and ECAM-l expression, caspase-3 activity | Mice | 10, 30,100 mg/kg i.p. injection | [55,161] |
| Study Type | Biological Effects | Cell lines, Animal Types | Doses | References |
|---|---|---|---|---|
| In vivo | ↓MDA and activated caspase-3 levels, ↑GSH levels, SOD activity, p-Akt level, motor functions, ⇅TRX and ATP levels | Rat | 20, 40, 80 mg/kg/day orally for 5 weeks | [163] |
| In vivo, in vitro | ↑autophagy-related gene 5 (Atg5) activity, mTOR/Ulk-involved autophagy, ↓PGC1β/mfn2-involved mitochondrial fusion, ⇅autophagic processes, mitochondrial fusion | Flies, human neuroblastoma cell (SH-SY5Y) | 2 mM orally for 25 days, 1–500 μM for 6 h | [170] |
| In vivo, in vitro | ⇅Akt/GSK3β-Nrf2/NF-κB signaling axis activity | Rat, murine microglia cell line (BV-2 cells) | 25, 50, 100 mg/kg/day orally for 32 days, 100, 200, 400 μM for 1 h | [169] |
| In vitro | ↓caspase-3/7 activity, ↑Bcl-2 expression, ERK1/2, and ERK5 activity | Human dopaminergic neuroblastoma cell line (SH-SY5Y) | 10, 20, 30 μM for 1 h | [9] |
| In vivo | glucose metabolism enhancement | Mice | 20, 80 mg/kg/day orally for 10 days | [171] |
| In vivo | ⇅Nrf2/ARE pathway and downstream gene expression, ↑SOD, GSSG, and GSH levels, ↓NO and MDA levels | Rat | 50 mg/kg orally | [164] |
| In vivo, in vitro | ↓MDA production,↑SOD and CAT activity | Rat, rat cortical neuron | 12.5, 25, 50 mg/kg/day orally for 30 days, 12.5, 50 μg/mL for 1 h | [167] |
| In vivo | ↓Bax expression, caspase-9 activity, caspase-3 activity, ROS levels, infarcted volume, mitochondrial dysfunction, ↑Bcl-2 expression | Rat | 30 mg/kg i.v. injection | [173] |
| In vivo | ↓infarction volume, neurobehavioral deficits, neuronal apoptosis, ROS level, p38 mitogen-activated protein kinase activity, c-Jun N-terminal kinase activity, ↑Nrf2/HO-1/TRX activity | Rat | 30 mg/kg i.p. injection twice | [62,167,174] |
| In vivo | ↑Gli1, Ptch1, and SOD1 expression, ↓NF-κB expression, BBB permeability, infarcted volume, brain water content | Rat | 25, 50 mg/kg i.p. injection once or for 2 days | [63] |
| In vivo | ↓brain edema | Rat | 12.5, 50 mg/kg i.p. injection once | [176] |
| In vivo | ↑Nrf2 activity, MDA and GSH levels, ↓hippocampal apoptosis, TUNEL-positive cells, cleaved caspase-3, cleaved caspase-9, NF-κB, TNF-α, PGE-2, and COX-2 levels | Rat | 50 mg/kg/day orally for 3 or 4 weeks | [81] |
| In vivo, in vitro | ↓NO, IL-1β, IL-6, and TNF-α levels, NLRP3 inflammasome activation, ↑motor function | Rat, BV2 mouse microglia | 20, 40 mg/kg i.p. injection once, 1, 2, 4 μM for 24 h | [49] |
| In vivo, in vitro | ↑Nrf2/ARE pathway activity, ↓mitochondrial dysfunction | Mice, spinal cord motor neurons | 30 mg/kg/day orally for 9 days, 3.75, 7.5, 15, 30, or 60 μM | [180] |
| In vitro | ⇅Nrf 2/ARE pathway activity | BMSCs | 3, 10, 30 μM for 2 h | [179] |
| In vivo, in vitro | ↑Nrf2 pathway activity | Mice, BMSCs | 20 mg/kg/day gastrically perfused for 5 days, 3, 10, 30 μM | [181] |
| In vivo | ↑BDNF expression | Rat | 10 mg/kg/day orally for 10 days | [178] |
| In vivo | ↓ICAM-1, VCAM-1, E-selectin, L-selectin, and Integrins levels, ↓neurological deficits, volume of brain infarction | Rat | 7.5, 15, 30 mg/kg i.v. injection once | [175] |
| In vivo, in vitro | ↑MALAT1/CREB/PGC-1α/PPARγ signaling pathway activity, BBB integrity, ↓cerebral infarcted volume, TNF-α, IL-6, IL-1β, COX-2, ICAM-1, VCAM-1, and MCP-1 expression | Rat, human embryonic kidney cell line (HEK-293T), human umbilical vein endothelial cell line (HUVEC) | 30 mg/kg i.v. injection once, 20 μM for 24 h | [53,172] |
| In vivo | ↑intracellular ATP, lysosomal stability, ↓mitochondrial swelling, vasoresponsiveness, arteriolar smooth muscle cell hyperpolarization, KATP channel activity | Rat | 15, 30, and 45 mg/100 g body weight | [166] |
| Study Type | Biological Effects | Cell Lines, Animal Types | Doses | References |
|---|---|---|---|---|
| In vivo | ⇅renal expression of mURAT1, mGLUT9, mAGCG2, mOAT1, mOCT1, mOCTN1, mOCTN2, mUMOD, ↑antihyperuricemic, nephroprotective effect | Mice | 20, 40 mg/kg/day orally for 7 days | [183] |
| In vivo | ↑PI3K/Akt pathway activity, SOD, GST, GSH-Px, and CAT activity, GSH level, ↓TNF-α, IL-1, COX-2, iNOS, PGE-2, NO, and MDA levels | Mice | 10,20,40 mg/kg/day i.p. injection for 6 days | [10,50,184] |
| In vivo | ↓NF-κB p65, COX-2, and iNOS expression, ┬TNF-α, PGE-2, and IL-1β production | Mice | 12.5, 25.0, 50.0 mg/kg/day orally for 4 weeks | [52] |
| In vivo | ↑AMPK activity, Sirt1 expression, ┬NF-κB/NLRP3 inflammasome activity | Rats | 25, 50 mg/kg/day orally for 7 days | [46,189] |
| In vivo, in vitro | ↑Cx32 expression, K48-linked polyubiquitination activity, ↓NOX4, ROS, FN, and ICAM-1 levels | Mice, primary GMCs of rat | 100 mg/k/day orally 6 days a week for 12 weeks, 5, 10, 20, 40, 80, 160 μM for 24 h | [185] |
| In vivo | ┬NF-κB signaling, ↑Nrf2 signaling pathway activity, ↓TNF-α, IL-1β, and IL-6 expression, MPO activity, MDA content | Mice | 20, 40, 80 mg/kg i.p. injection once | [186] |
| In vivo, in vitro | ↑Sirt1/Nrf2/ARE pathway activity, ↓ROS, FN, and TGF-β1 levels | Rats, GMCs of rat | 150 mg/kg/day orally for 12 weeks, 5, 10, 20 μM | [65] |
| In vivo, in vitro | ↓AMPK/p38 MAPK signaling pathway activity, autophagy imbalance, ↑Nrf2 signaling pathway activity | Rat, conditionally immortalized human podocytes (HPCs) | 7.5, 15, 30 mg/kg/day orally for 6 weeks, 5, 10, 20 μM for 24 h | [192] |
| In vivo, in vitro | ↑Sirt1–p53 expression, ↓mitochondrial dysfunction | Rat, human renal proximal tubular epithelial cell line (HK-2) | 30 mg/kg infusion for 2 h, various concentrations for 2 h and 50 μM for 48 h | [191] |
| In vivo | ↓IL-6 level, lipid peroxide, lysosomal instability, mitochondrial swelling, ↑ATP level, mitochondrial membrane potential | Rats | 30 mg/kg i.v. injection three times (6, 12, and 18 h after model establishment) | [190] |
| In vivo | ↑Sirt1 activity, Parkin-dependent mitophagy, ↓NLRP3 inflammasome activity, mitochondrial dysfunction | mice | 30 mg/kg i.v. injection once | [187] |
| Study Type | Biological Effects | Cell Lines, Animal Types | Doses | References |
|---|---|---|---|---|
| In vivo, in vitro | ┬TGF-β1/Smad3 signaling pathway activity, epithelial–mesenchymal transition, ↓IL-4 and IL-3 levels, ↑Sirt3 expression, Nrf2 and PGC1a levels, INF-γ activity | Mice, human bronchial epithelial cell line (BEAS-2B) | 100 mg/kg/day i.p. injection for 31 days | [193] |
| In vivo | ↓MDA level, W/D, ultrastructure injuries, ↑SOD content | Rabbit | - | [198] |
| In vivo | ↓TLR4 and NF-κB expression, ┬inflammatory mediators | Rabbit | - | [197] |
| In vivo | ↓TNF-α, IL-1β, and IL-6 levels, MPO activity, total cells, PMNs in BALF, Bcl-xl expression, ↑Bax and caspase-3 activity | Rat | 15, 30, 45 mg/kg via caudal vein once | [199] |
| In vivo, in vitro | ↑mitophagy via Parkin, Bcl-2 expression, ↓mitochondrial-dependent apoptosis, Bax expression, cytochrome c release, caspase-3 activity | Mice, human bronchial epithelial cell line (BEAS 2B cells) | 45 mg/kg once, 50 μM for 6 h | [26] |
| In vivo | ┬PLA2 | Rat | 1 mg/kg i.v. injection once | [203] |
| In vitro, in vivo | ┬TGF-β/Smad/ERK pathway activity, ↓α-smooth muscle actin, collagen I, TNF-α, IL-6, IL-13, MPO, and MDA expression, ↑epithelial cell cadherin expression, SOD activity | Rat | 10, 40, and 160 mg/kg/day for 28 days | [196] |
| In vivo | ↓PLA2 activity, hydroxyproline, PGE-2, LTC4, and TGF-β1 levels in BALF | Rat | 10, 20, 40 mg/kg intraperitoneal injection once before model establishment | [204] |
| In vivo, in vitro | ┬TGF-β1 activity, ↓ROS activity, ↑Nrf2 signaling pathway activity | Mice, human lung epithelial cell line (BEAS-2B) | 100 mg/kg at days 0, 7, 14, 100 μM for up to 48 h | [195] |
| In vivo | ⇅SP-D and UCN expression, ↓serum IgE, IL-4, IL-13, IL-5, MDA, TNF-α, IFN-γ, iNOS, and eosinophil levels, ↑SOD activity, GSH level | Rats | 200 mg/kg/day orally for 14 days | [194] |
| In vitro | ↑GSH-Px and SOD content, ↓apoptosis, MDA content, activation-related proteins of the NLRP3 inflammasome, TNF-α, TGF-β, IL-1β, IL-6 | Embryonic lung fibroblast MRC-5 | 12.5, 25, 50, 100, 200, 400 μmol/L for 24 h | [205] |
| In vitro, in vivo | ↑CCSP expression, ┬PLA2 activation | Rats, human bronchial epithelia cells (BEAS-2B cells) | 1, 5, 10, 30 mg/kg i.v. injection once, 0.5 m mol/L for 4 h | [54,169,201] |
| In vivo | ↓TUNEL-positive lung cells, TNF-α, IL-1, and IL-6 levels, lung MPO activity, ┬Bax upregulation, caspase-3 activity, Bcl-xl downregulation | Rat | 45 mg/kg i.v. injection once | [200] |
| In vitro, in vivo | ┬TLR4/MyD88/NF-κB signaling pathway activity, W/D, MPO, neutrophil number, ↓IL-6, IL-1β, TNF-α, IL-8 levels | Mice, human bronchial epithelial cell line (BEAS-2B cells) | 20, 80 mg/kg i.p. injection once, 2 μM, 4 μM, 8 μM for 2 h | [202] |
| In vivo | ┬NF-κB activity, ↓TNF-α and IL-6 levels, lung COX-2 and iNOS expression | Mice | 15, 45, 100 mg/kg i.p. injection once | [151] |
| Study Type | Biological Effects | Cell Lines, Animal Types | Doses | References |
|---|---|---|---|---|
| In vivo, in vitro | ┬NET formation | Mice, neutrophils isolated from RA patients and mice | 45 mg/kg/day i.p. injection for 24 days, 0, 50, 75, 100, and 125 μg/mL | [208] |
| In vivo | ↓TNF-α, IL-6, IL-17, and MMP levels, MPO activity, MDA level, ↓RANKL and STAT3 expression, ┬VEGF, NF-κB, ↑GSH content | Rats | 200 mg/kg/day orally for 21 days | [56] |
| In vivo | ↓IL-1β, TNF-α, MDA, and SOD expression, ↑Bcl-2/Bax pathway expression, MMP-9 activity, ┬caspase-3/9 activity | Mice | 15, 30, 45 mg/kg i.d. injection for 24 h | [206] |
| In vivo, in vitro | ┬ROS-mediated NET formation | Mice, neutrophils from systematic lupus erythematous patients and healthy people | 45 mg/kg/day i.p. injection for 8 or 16 weeks, 50, 75, 100, 125, and 150 μg/mL | [207] |
| Study Type | Biological Effects | Cell Lines, Animal Types | Doses | References |
|---|---|---|---|---|
| In vivo, in vitro | ↑Nrf2 signaling pathway activity, Parkin expression | Rats, human endplate chondrocytes | 50 mg/kg/day orally for 4 weeks, 200 μM for 2 h | [30] |
| In vivo, in vitro | ⇅BMP2–Wnt/β-catenin, ↑TAZ, mRNAs RUNX2, osteopontin, DLX5, β-catenin, and osteocalcin expression | Mice, hBMSCs | 3 mg/kg every 2 days i.p. injection for 12 weeks, 0, 30, 10, 100 μM for 1, 2, 3, 7, 14 days | [210] |
| In vitro | ↑ERK ½ activity | Mouse BMSCs | 5, 10, 15, 30, 100 μM for 24 h | [212] |
| In vitro | ↑active caspase-3, Bax, LC3II, Beclin 1, and Atg5 expression, ↓Bcl2 expression | Fibroblasts of patients with AS | 0, 0.33, 1, 3, 10 μM for 24, 48, or 72 h | [213] |
| In vivo, In vitro | ↑aggrecan and collagen II levels, Nrf2 signaling pathway activity, ↓TNF-α, p53, p16, MMP-3, MMP-13, and ADAMTS-4 expression | Rats, nucleus pulposus cells of rats | 50 mg/kg/day orally for 4 weeks, 0, 200, 400 μmol/L for 24 h | [214] |
| In vivo, in vitro | ⇅osteoprotegerin, RANKL, β-catenin expression | Mice, mouse bone marrow stromal cell line ST2 | 10, 20, 40, mg/kg/day i.p. injection for 12 weeks, 20, 40 μg/mL for 48 h | [209] |
| Study Type | Biological Effects | Cell Lines, Animal Types | Doses | References |
|---|---|---|---|---|
| In vivo | ↓VEGF, nerve growth factor, ICAM and MMP-9 expression, lymphocyte accumulation, peroxynitrite formation, PARP activation, IkBa phosphorylation, NF-κB translocation | Rats | 10 mg/kg/day orally for 14 days | [217] |
| In vivo | ↑capillary perfusion, functional capillary density, survival time | Rabbits | single bolus infusion 4 mL/kg | [219] |
| In vivo | ↓NF-κB activity, IL-6, IL-1β, 8-OHdG, 4-HNE, IL-6, γ-H2AX, Bax, cleaved-caspase-3, IL-6, caspase-1, phosphorylated p6 levels | Mice | 50, 100 mg/kg/day orally for 28 days | [220] |
| In vivo | ┬NF-κB pathway activity, ↑Nrf2 signaling pathway activity, ↓TNF-α, IL-1β, and IL-6 expression | Mice | 20, 40, 80 mg/kg i.p. injection once | [31] |
| Clinical Study Type | Health Benefits | Gender | Dose | Study Type | References |
|---|---|---|---|---|---|
| Women’s health | ↓chronic pelvic pain, deep dyspareunia, dysmenorrhea, dyschezia, ↑life quality | Symptomatic women with endometriosis desiring pregnancy | 600 mg um-PEA orally twice a day for 10 days followed by 400/40 mg m(PEA/polydatin) orally twice a day for another 80 days | single-arm, non-randomized, open-label clinical study | [218] |
| Gastrointestinal | ↓abdominal pain severity | IBS patients and healthy people | 200/20 mg PEA/polydatin twice a day for 12 weeks | randomized, double-blind, placebo-controlled, multi-center clinical study | [234] |
| Liver disease | ↓AST and ALT levels, lipid peroxidation, ↓cognitive impairment | Alcoholic patients | 40 mg orally twice a day for 2 weeks | A pilot study | [235] |
| Antioxidant and anti-inflammatory activity | ↓oxidized species levels, neopterin levels,↑GSH level, other thiol species, vitamins C, E, and A | Healthy people | 35 mg orally twice a day for 8 weeks | randomized clinical study | [236] |
| Skin | ↓pruritus, skin manifestation such as papulopustular rash,↑quality of life | Cancer patients taking anti-EGFR treatment regimen with or without cutaneous adverse events | Cream (1.5% or 0.8%) twice a day topically for 6 months | prospective pilot study | [237] |
| Skin | ↓incidence of rash | Patients with mutated NSCLC taking afatinib | 1.5% cream twice a day topically for 6.4 months | pilot study | [230] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Karami, A.; Fakhri, S.; Kooshki, L.; Khan, H. Polydatin: Pharmacological Mechanisms, Therapeutic Targets, Biological Activities, and Health Benefits. Molecules 2022, 27, 6474. https://doi.org/10.3390/molecules27196474
Karami A, Fakhri S, Kooshki L, Khan H. Polydatin: Pharmacological Mechanisms, Therapeutic Targets, Biological Activities, and Health Benefits. Molecules. 2022; 27(19):6474. https://doi.org/10.3390/molecules27196474
Chicago/Turabian StyleKarami, Ahmad, Sajad Fakhri, Leila Kooshki, and Haroon Khan. 2022. "Polydatin: Pharmacological Mechanisms, Therapeutic Targets, Biological Activities, and Health Benefits" Molecules 27, no. 19: 6474. https://doi.org/10.3390/molecules27196474
APA StyleKarami, A., Fakhri, S., Kooshki, L., & Khan, H. (2022). Polydatin: Pharmacological Mechanisms, Therapeutic Targets, Biological Activities, and Health Benefits. Molecules, 27(19), 6474. https://doi.org/10.3390/molecules27196474