
Therapeutic and Nutraceutical Effects of Polyphenolics from Natural Sources

 ,
,  , ,
, ,
Abstract
1. Introduction
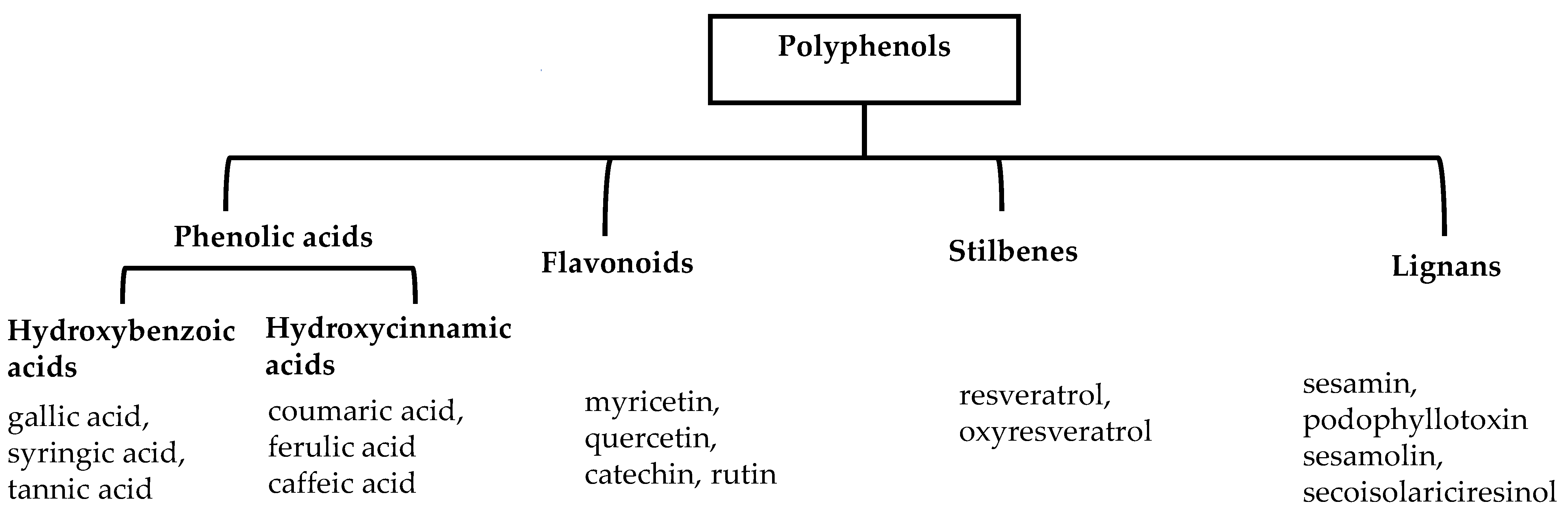
2. Classification of Polyphenols
2.1. Phenolic Acids
2.2. Flavonoids
2.3. Stilbenes
2.4. Lignans
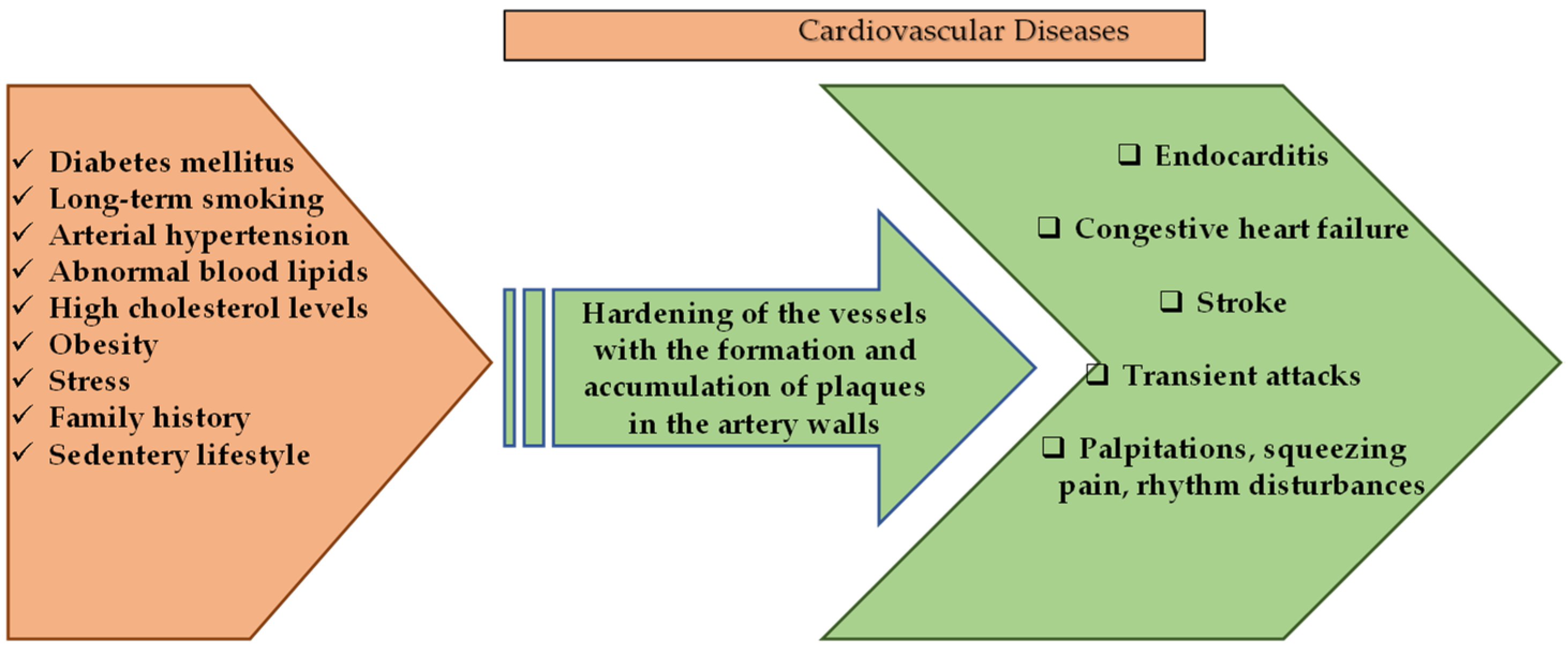
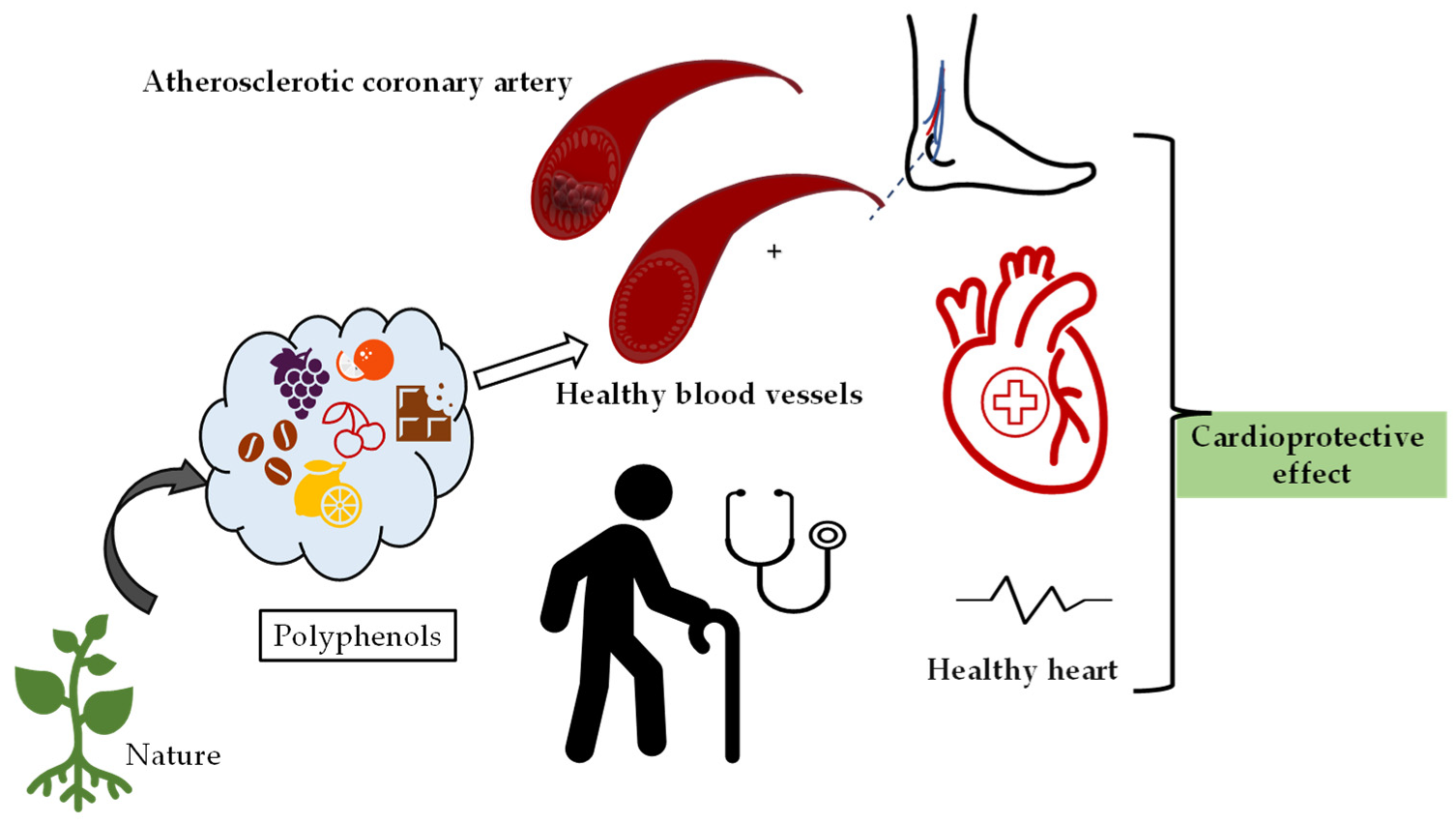
3. Cardioprotective Effects of Polyphenolic Compounds
3.1. Phenolic Acids
3.2. Certain Cardioprotective Effects of Flavonoids
3.3. Stilbenes
3.4. Lignans

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Study Design | Outcomes | Ref. |
|---|---|---|
| Caffeic acid (100 nM and 1 μM) human umbilical vein-derived endothelial cells (HUVEC) | Caffeic acid increased proliferation and angiogenesis, inhibited leukocyte adhesion and endothelial cell apoptosis under hypoxia or by the uremic toxins’ conditions | [111] |
| Caffeic acid phenethyl ester (CAPE) 30 mg/kg/day administered by oral gavage for 6 weeks. Streptozotocin induced diabetes-induced atherosclerosis in rat model | CAPE abolished the diabetes-associated atherosclerotic changes by improving important functional and structural disorders in vessels. CAPE alleviated the elevation in systolic and diastolic BP | [112] |
| High fructose (HFCS) induced diabetic rats’ subacute CAPE administrations (50 μmol/kg/day intraperitoneally for 2 weeks | CAPE ameliorated the elevation in blood pressure, vascular damage, and it increased eNOS levels. CAPE lowered homocysteine and cholesterol levels | [113] |
| Atherogenic diet (Ath)-induced rat model administrations with caffeic acid 50 mg/kg, p.o. | CA ameliorated lipid profile and reduced the oxidative stress level. In aorta revealed reduction of the atherosclerotic lesions | [114] |
| Dietary 10% flaxseed content 1.37 mg/g SDG, (the lignan secoisolariciresinol diglucoside) LDL receptor-deficient mouse (LDLrKO) fed a cholesterol-supplemented diet and an increase in atherosclerotic plaque formation | Flaxseed lowered plasma cholesterol levels and saturated fatty acids, increased plasma ALA levels, and inhibited plaque formation in the aorta and reduced the inflammatory markers (IL-6, mac-3, and VCAM-1) | [115] |
| A 40 g/day of ground flaxseed administered Sixty-two men and post-menopausal women (LDL-C between 130 and 200) mg/dl 10 weeks | Flaxseed lowered LDL-C short lived and did not affect inflammation or oxidative stress | [116] |
| Low-density lipoprotein receptor knockout (LDLR−/−) mice with 170 g/kg sesame oil diet 3 months of feeding | Reduced atherosclerotic lesion formation, plasma cholesterol, triglyceride, and LDL cholesterol levels. Anti-inflammatory property (reduced inflammatory cytokines, such as MCP-1, RANTES, IL-1a, IL-6, and CXCL-16) | [117,118] |
| Sesamol 50 mg/kg orally for 6 weeks (DOCA)-salt-induced hypertensive rats | Decreased systolic and diastolic blood pressure and lipid peroxidation and enhanced the antioxidant activity. Hypertensive rats showed cardiac muscle fiber rupture and mononuclear infiltration, but Sesamol 50 mg/kg group heart showed to near-normal architecture | [119] |
| Isoproterenol treated myocardial infarcted rats, pretreated with gallic acid (15 mg/kg) daily for a period of 10 days | Prevented the changes in the activities of cardiac marker enzymes (CK-MB and LDH), reduced the levels of lipid peroxidation products (LPO), glutathione and lysosomal membrane damage | [120] |
| Rats were infused with AGEs (advanced glycation end products play a role development of cardiovascular disorders) and then treated with gallic acid (GA) by oral gavage daily at a dose of 25 mg/kg BW/day for 30 days | AGEs induced cardiac fibrosis and augmented oxidative stress in the heart tissues. GA prevented the upregulation of pro-fibrotic genes and ECM proteins (↓TNF-α, TGF-β, MMP-2 and -9 expression). GA treatment effectively prevented cardiac remodeling | [121] |
| NG-nitro-L-arginine methyl ester (L-NAME)-induced hypertensive mice treatment with gallic acid 100 mg/kg per day by daily intraperitoneal injections 3 or 8 weeks | GA attenuated cardiac fibrosis and remodeling, reduced the expression of histone deacetylase 1 (HDAC1) and 2 (HDAC2). GA lowered the elevated SBP | [122] |
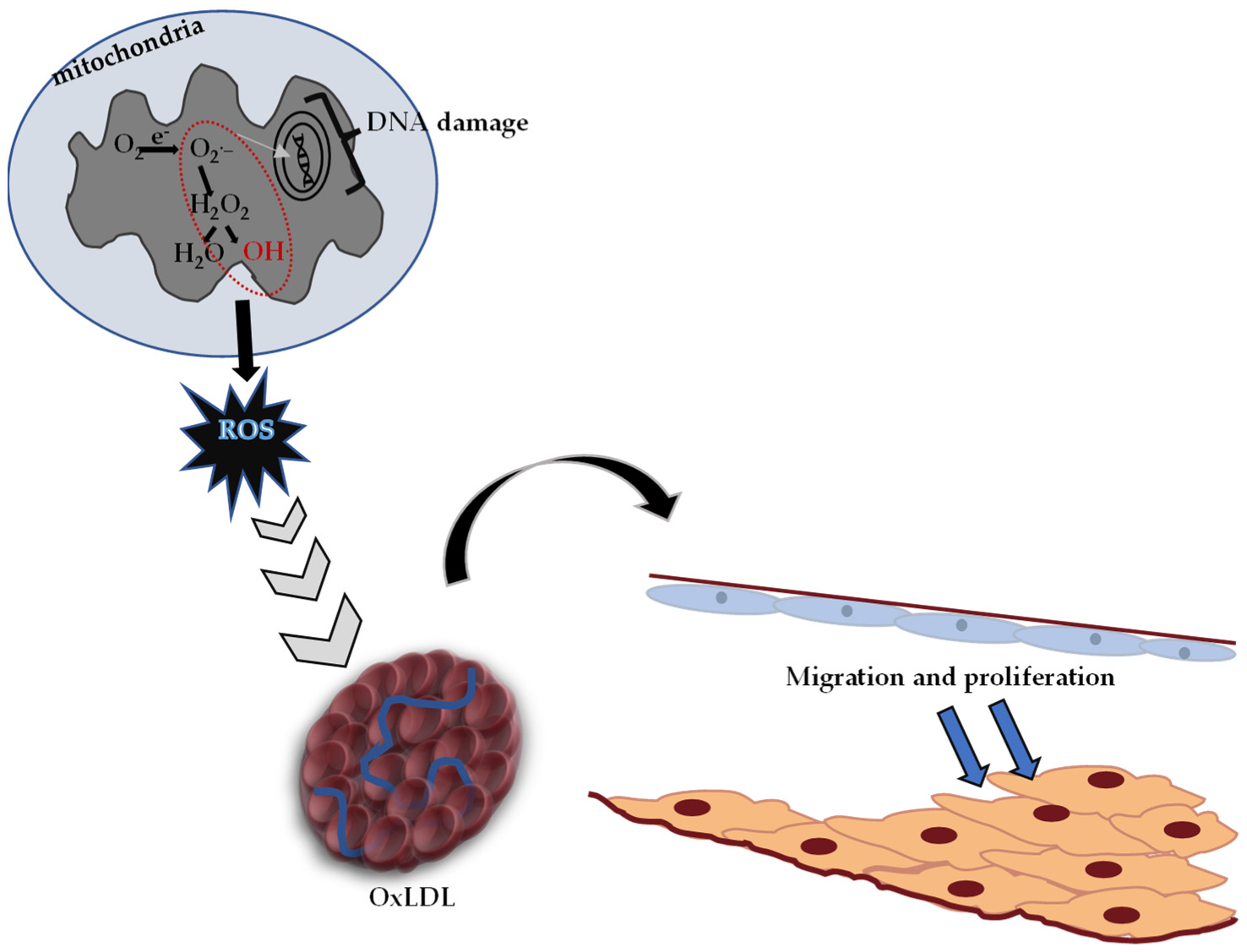
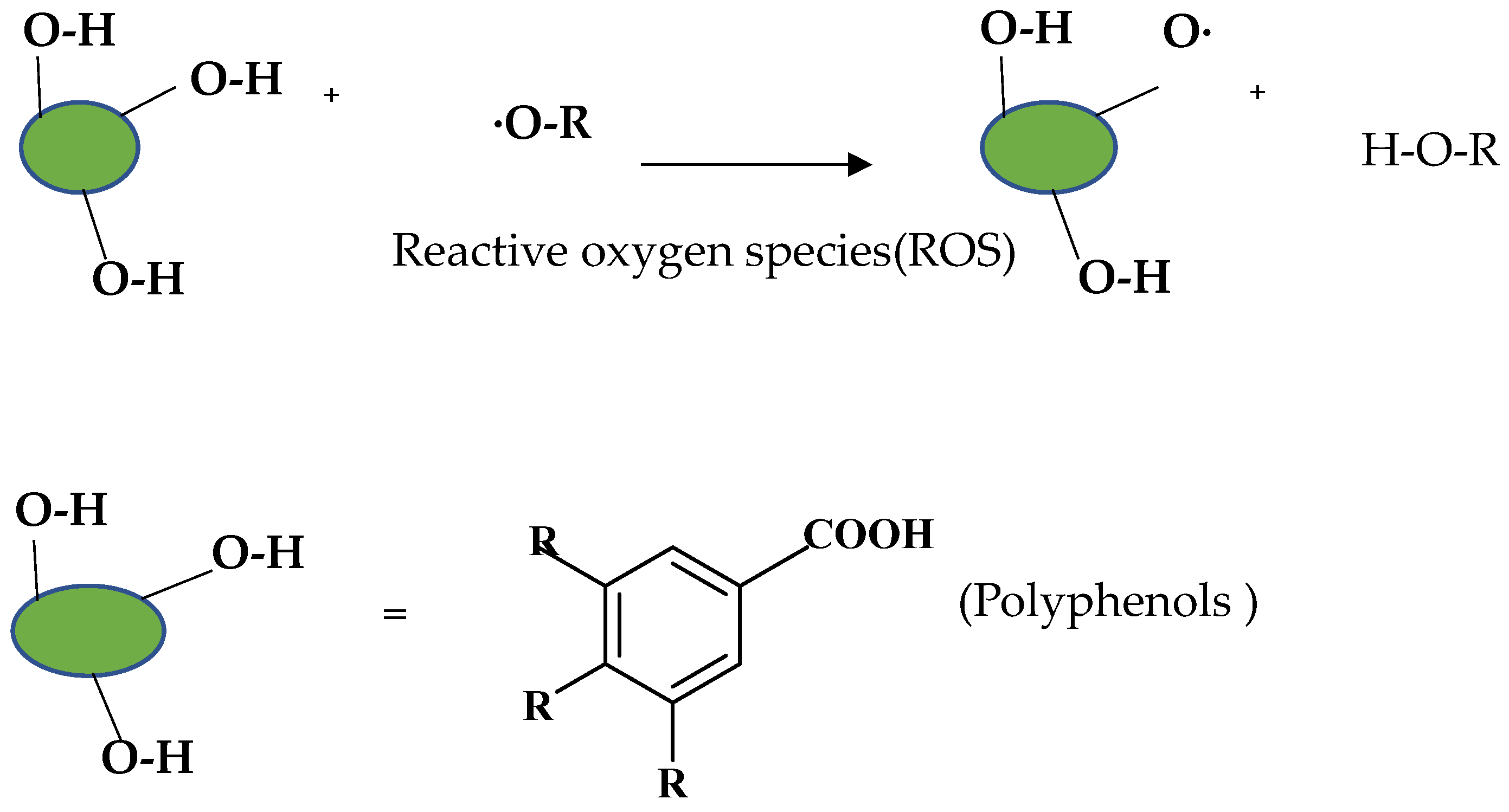
4. Protective Effects of Polyphenolic Compounds on ROS-Induced Oxidative Stress
4.1. Phenolic Acids
4.2. Flavonoids
4.3. Stilbenes
4.4. Lignans
5. Polyphenols on DNA and Cancer
5.1. Phenolic Acids
5.2. Flavonoids
5.3. Stilbenes
5.4. Lignans
| Study Design | Outcomes | Ref. |
|---|---|---|
| Naringin’s anti-tumor effects on thyroid cancer were studied. For 24, 48, or 72 h at 37 °C, Naringin (6, 12, and 25 g/mL) was applied to two cancer cells, TPC-1 and SW1736. | In MTT assays, naringin inhibited TPC-1 and SW1736 cell proliferation in a dose- and time-dependent manner. | [167] |
| Naringin was combined with the drug, atorvastatin. | The IC50 of naringin was determined as 196.2 μM in PC-3 and 117.2 μM in LNCaP cells. However, naringin and statin drug atorvastatin synergistically inhibited prostate cancer cells, PC-3, and LNCaP cells. | [168] |
| Catechin was studied on MC38 colon cancer cells. | The proliferation of cancer cells can be significantly inhibited by catechin concentrations between 250 and 1000 mg/mL, and the IC50 value is 250 mg/mL during a 24 h incubation period. Incubation time was 72 h, and IC50 was 142 mg/mL after triggering cancer cells. | [176] |
| An anticancer effect of gallic acid on non-small cell lung cancer cells A549. | The viability of A549 cells was determined by MTT assay after treatment with GA (0–52 g/mL) for 24 h. There was a dose-dependent decrease in viability after treatment with GA (0–52 g/mL). When compared with control cells, 12 g/mL GA significantly decreased cell viability. | [188] |
6. Polyphenols on Aging
6.1. Phenolic Acids
6.2. Flavonoids
6.3. Stilbenes
6.4. Lignans
7. Conclusions and Future Respects
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Rashidi, L. Different nano-delivery systems for delivery of nutraceuticals. Food Biosci. 2021, 43, 101258. [Google Scholar] [CrossRef]
- Lyu, X.; Lee, J.; Chen, W.N. Potential Natural Food Preservatives and Their Sustainable Production in Yeast: Terpenoids and Polyphenols. J. Agric. Food Chem. 2019, 67, 4397–4417. [Google Scholar] [CrossRef] [PubMed]
- Xie, G.; Zhao, A.; Zhao, L.; Chen, T.; Chen, H.; Qi, X.; Zheng, X.; Ni, Y.; Cheng, Y.; Lan, K.; et al. Metabolic Fate of Tea Polyphenols in Humans. J. Proteome Res. 2012, 11, 3449–3457. [Google Scholar] [CrossRef] [PubMed]
- Burns, J.; Yokota, T.; Ashihara, H.; Lean, M.E.J.; Crozier, A. Plant Foods and Herbal Sources of Resveratrol. J. Agric. Food Chem. 2002, 50, 3337–3340. [Google Scholar] [CrossRef] [PubMed]
- Tzanova, M.; Peeva, P. Rapid HPLC Method for Simultaneous Quantification of trans-Resveratrol and Quercetin in the Skin of Red Grapes. Food Anal. Methods 2018, 11, 514–521. [Google Scholar] [CrossRef]
- Sobolev, V.S.; Cole, R.J. trans-Resveratrol Content in Commercial Peanuts and Peanut Products. J. Agric. Food Chem. 1999, 47, 1435–1439. [Google Scholar] [CrossRef] [PubMed]
- Shrikanta, A.; Kumar, A.; Govindaswamy, V. Resveratrol content and antioxidant properties of underutilized fruits. J. Food Sci. Technol. 2015, 52, 383–390. [Google Scholar] [CrossRef] [PubMed]
- Xu, D.-P.; Li, Y.; Meng, X.; Zhou, T.; Zhou, Y.; Zheng, J.; Zhang, J.-J.; Li, H.-B. Natural Antioxidants in Foods and Medicinal Plants: Extraction, Assessment and Resources. Int. J. Mol. Sci. 2017, 18, 96. [Google Scholar] [CrossRef] [PubMed]
- Uddin, N.; Muhammad, N.; Nisar, M.; Aisha; Ali, N.; Ullah, R.; Ali, E.A.; Khan, A.A.; Rahman, I.U.; Khan, A.; et al. Distribution of polyphenolic compounds, antioxidant potential, and free amino acids in Ziziphus fruits extract; a study for determining the influence of wider geography. Food Sci. Nutr. 2022, 10, 1414–1430. [Google Scholar] [CrossRef]
- Besednova, N.N.; Andryukov, B.G.; Zaporozhets, T.S.; Kryzhanovsky, S.P.; Kuznetsova, T.A.; Fedyanina, L.N.; Makarenkova, I.D.; Zvyagintseva, T.N. Algae Polyphenolic Compounds and Modern Antibacterial Strategies: Current Achievements and Immediate Prospects. Biomedicines 2020, 8, 342. [Google Scholar] [CrossRef]
- Staszowska-Karkut, M.; Materska, M. Phenolic Composition, Mineral Content, and Beneficial Bioactivities of Leaf Extracts from Black Currant (Ribes nigrum L.), Raspberry (Rubus idaeus), and Aronia (Aronia melanocarpa). Nutrients 2020, 12, 463. [Google Scholar] [CrossRef] [PubMed]
- Oracz, J.; Zyzelewicz, D.; Nebesny, E. The Content of Polyphenolic Compounds in Cocoa Beans (Theobroma cacao L.), Depending on Variety, Growing Region, and Processing Operations: A Review. Crit. Rev. Food Sci. Nutr. 2015, 55, 1176–1192. [Google Scholar] [CrossRef] [PubMed]
- Alara, O.R.; Abdurahman, N.H.; Abdul Mudalip, S.K.; Olalere, O.A. Microwave-assisted extraction of Vernonia amygdalina leaf for optimal recovery of total phenolic content. J. Appl. Res. Med. Aromat. Plants 2018, 10, 16–24. [Google Scholar] [CrossRef]
- Pashazadeh, B.; Elhamirad, A.H.; Hajnajari, H.; Sharayei, P.; Armin, M. Optimization of the pulsed electric field-assisted extraction of functional compounds from cinnamon. Biocatal. Agric. Biotechnol. 2020, 23, 101461. [Google Scholar] [CrossRef]
- Sun, C.; Zhao, C.; Guven, E.C.; Paoli, P.; Simal-Gandara, J.; Ramkumar, K.M.; Wang, S.; Buleu, F.; Pah, A.; Turi, V.; et al. Dietary polyphenols as antidiabetic agents: Advances and opportunities. Food Front. 2020, 1, 18–44. [Google Scholar] [CrossRef]
- Niedzwiecki, A.; Roomi, M.; Kalinovsky, T.; Rath, M. Anticancer Efficacy of Polyphenols and Their Combinations. Nutrients 2016, 8, 552. [Google Scholar] [CrossRef]
- Xing, L.; Zhang, H.; Qi, R.; Tsao, R.; Mine, Y. Recent Advances in the Understanding of the Health Benefits and Molecular Mechanisms Associated with Green Tea Polyphenols. J. Agric. Food Chem. 2019, 67, 1029–1043. [Google Scholar] [CrossRef]
- Etxeberria, U.; Fernández-Quintela, A.; Milagro, F.I.; Aguirre, L.; Martínez, J.A.; Portillo, M.P. Impact of Polyphenols and Polyphenol-Rich Dietary Sources on Gut Microbiota Composition. J. Agric. Food Chem. 2013, 61, 9517–9533. [Google Scholar] [CrossRef]
- Duda-Chodak, A.; Tarko, T.; Satora, P.; Sroka, P. Interaction of dietary compounds, especially polyphenols, with the intestinal microbiota: A review. Eur. J. Nutr. 2015, 54, 325–341. [Google Scholar] [CrossRef]
- Singla, R.K.; Dubey, A.K.; Garg, A.; Sharma, R.K.; Fiorino, M.; Ameen, S.M.; Haddad, M.A.; Al-Hiary, M. Natural Polyphenols: Chemical Classification, Definition of Classes, Subcategories, and Structures. J. AOAC Int. 2019, 102, 1397–1400. [Google Scholar] [CrossRef]
- Sahiner, M.; Sahiner, N.; Sagbas, S.; Fullerton, M.L.; Blake, D.A. Fabrication of Biodegradable Poly(naringin) Particles with Antioxidant Activity and Low Toxicity. ACS Omega 2018, 3, 17359–17367. [Google Scholar] [CrossRef]
- Kumar, V.; Sharma, A.; Kohli, S.K.; Bali, S.; Sharma, M.; Kumar, R.; Bhardwaj, R.; Thukral, A.K. Differential distribution of polyphenols in plants using multivariate techniques. Biotechnol. Res. Innov. 2019, 3, 1–21. [Google Scholar] [CrossRef]
- Rashmi, H.B.; Negi, P.S. Phenolic acids from vegetables: A review on processing stability and health benefits. Food Res. Int. 2020, 136, 109298. [Google Scholar] [CrossRef]
- Kumar, N.; Goel, N. Phenolic acids: Natural versatile molecules with promising therapeutic applications. Biotechnol. Rep. 2019, 24, e00370. [Google Scholar] [CrossRef]
- Sahiner, N.; Sagbas, S.; Sahiner, M.; Blake, D.A.; Reed, W.F. Polydopamine particles as nontoxic, blood compatible, antioxidant and drug delivery materials. Colloids Surf. B Biointerfaces 2018, 172, 618–626. [Google Scholar] [CrossRef]
- Nakamura, K.; Yamada, Y.; Ikai, H.; Kanno, T.; Sasaki, K.; Niwano, Y. Bactericidal Action of Photoirradiated Gallic Acid via Reactive Oxygen Species Formation. J. Agric. Food Chem. 2012, 60, 10048–10054. [Google Scholar] [CrossRef]
- De Cristo Soares Alves, A.; Mainardes, R.M.; Khalil, N.M. Nanoencapsulation of gallic acid and evaluation of its cytotoxicity and antioxidant activity. Mater. Sci. Eng. C 2016, 60, 126–134. [Google Scholar] [CrossRef]
- Santos, E.M.S.; da Rocha, R.G.; Santos, H.O.; Guimarães, T.A.; de Carvalho Fraga, C.A.; da Silveira, L.H.; Batista, P.R.; de Oliveira, P.S.L.; Melo, G.A.; Santos, S.H.; et al. Gallic acid modulates phenotypic behavior and gene expression in oral squamous cell carcinoma cells by interfering with leptin pathway. Pathol.-Res. Pract. 2018, 214, 30–37. [Google Scholar] [CrossRef]
- Garud, M.S.; Kulkarni, Y.A. Gallic acid attenuates type I diabetic nephropathy in rats. Chem. Biol. Interact. 2018, 282, 69–76. [Google Scholar] [CrossRef]
- Sahiner, N.; Sagbas, S.; Sahiner, M.; Silan, C. P(TA) macro-, micro-, nanoparticle-embedded super porous p(HEMA) cryogels as wound dressing material. Mater. Sci. Eng. C 2017, 70, 317–326. [Google Scholar] [CrossRef]
- Sahiner, M.; Kurt, S.B.; Sahiner, N. Biodiverse Properties of Tannic Acid-Based Fibers. Fibers Polym. 2021, 22, 2986–2994. [Google Scholar] [CrossRef]
- Sahiner, M.; Blake, D.A.; Fullerton, M.L.; Suner, S.S.; Sunol, A.K.; Sahiner, N. Enhancement of biocompatibility and carbohydrate absorption control potential of rosmarinic acid through crosslinking into microparticles. Int. J. Biol. Macromol. 2019, 137, 836–843. [Google Scholar] [CrossRef]
- Noor, S.; Mohammad, T.; Rub, M.A.; Raza, A.; Azum, N.; Yadav, D.K.; Hassan, M.I.; Asiri, A.M. Biomedical features and therapeutic potential of rosmarinic acid. Arch. Pharm. Res. 2022, 45, 205–228. [Google Scholar] [CrossRef]
- Sahiner, M. Hydrolytic nondegradable bioactive rosmarinic acid particles. Polym. Adv. Technol. 2021, 32, 4891–4901. [Google Scholar] [CrossRef]
- Friedman, M.; Levin, C.E. Analysis and Biological Activities of Potato Glycoalkaloids, Calystegine Alkaloids, Phenolic Compounds, and Anthocyanins. In Advances in Potato Chemistry and Technology; Elsevier: Amsterdam, The Netherlands, 2009; pp. 127–161. [Google Scholar]
- Sahiner, N.; Sagbas, S.; Sahiner, M.; Aktas, N. Degradable Natural Phenolic Based Particles with Micro- and Nano-Size Range. Recent Pat. Mater. Sci. 2018, 11, 33–40. [Google Scholar] [CrossRef]
- Chen, R.; Qi, Q.-L.; Wang, M.-T.; Li, Q.-Y. Therapeutic potential of naringin: An overview. Pharm. Biol. 2016, 54, 3203–3210. [Google Scholar] [CrossRef]
- Alpaslan, D.; Dudu, T.E.; Şahiner, N.; Aktas, N. Synthesis and preparation of responsive poly(Dimethyl acrylamide/gelatin and pomegranate extract) as a novel food packaging material. Mater. Sci. Eng. C 2020, 108, 110339. [Google Scholar] [CrossRef]
- El Khawand, T.; Courtois, A.; Valls, J.; Richard, T.; Krisa, S. A review of dietary stilbenes: Sources and bioavailability. Phytochem. Rev. 2018, 17, 1007–1029. [Google Scholar] [CrossRef]
- Lu, H.-P.; Jia, Y.-N.; Peng, Y.-L.; Yu, Y.; Sun, S.-L.; Yue, M.-T.; Pan, M.-H.; Zeng, L.-S.; Xu, L. Oxyresveratrol, a Stilbene Compound from Morus alba L. Twig Extract Active against Trichophyton rubrum. Phyther. Res. 2017, 31, 1842–1848. [Google Scholar] [CrossRef]
- Giuliano, C.; Siani, F.; Mus, L.; Ghezzi, C.; Cerri, S.; Pacchetti, B.; Bigogno, C.; Blandini, F. Neuroprotective effects of lignan 7-hydroxymatairesinol (HMR/lignan) in a rodent model of Parkinson’s disease. Nutrition 2020, 69, 110494. [Google Scholar] [CrossRef] [PubMed]
- Bylund, A.; Saarinen, N.; Zhang, J.; Bergh, A.; Widmark, A.; Johansson, A.; Lundin, E.; Adlercreutz, H.; Hallmans, G.; Stattin, P.; et al. Anticancer Effects of a Plant Lignan 7-Hydroxymatairesinol on a Prostate Cancer Model In Vivo. Exp. Biol. Med. 2005, 230, 217–223. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.-Y.; Kim, J.-K.; Choi, J.-H.; Jung, J.-Y.; Oh, W.-Y.; Kim, D.C.; Lee, H.S.; Kim, Y.S.; Kang, S.S.; Lee, S.-H.; et al. Hepatoprotective Effect of Pinoresinol on Carbon Tetrachloride–Induced Hepatic Damage in Mice. J. Pharmacol. Sci. 2010, 112, 105–112. [Google Scholar] [CrossRef]
- Zheng, J.; Zhou, Y.; Li, S.; Zhang, P.; Zhou, T.; Xu, D.-P.; Li, H.-B. Effects and Mechanisms of Fruit and Vegetable Juices on Cardiovascular Diseases. Int. J. Mol. Sci. 2017, 18, 555. [Google Scholar] [CrossRef] [PubMed]
- Cardiovascular Diseases. Available online: https://www.who.int/health-topics/cardiovascular-diseases#tab=tab_2 (accessed on 16 April 2021).
- Jamee Shahwan, A.; Abed, Y.; Desormais, I.; Magne, J.; Preux, P.M.; Aboyans, V.; Lacroix, P. Epidemiology of coronary artery disease and stroke and associated risk factors in Gaza community–Palestine. PLoS ONE 2019, 14, e0211131. [Google Scholar] [CrossRef]
- Braunwald, E. Cardiovascular Medicine at the Turn of the Millennium: Triumphs, Concerns, and Opportunities. N. Engl. J. Med. 1997, 337, 1360–1369. [Google Scholar] [CrossRef]
- Chiva-Blanch, G.; Arranz, S.; Lamuela-Raventos, R.M.; Estruch, R. Effects of Wine, Alcohol and Polyphenols on Cardiovascular Disease Risk Factors: Evidences from Human Studies. Alcohol Alcohol. 2013, 48, 270–277. [Google Scholar] [CrossRef]
- Jneid, H.; Bhatt, D.L.; Corti, R.; Badimon, J.J.; Fuster, V.; Francis, G.S. Aspirin and Clopidogrel in Acute Coronary Syndromes. Arch. Intern. Med. 2003, 163, 1145. [Google Scholar] [CrossRef]
- Frei, B. Natural Antioxidants in Human Health and Disease; Elsevier: Amsterdam, The Netherlands, 1994; ISBN 9780080571683. [Google Scholar]
- Davignon, J. Role of Endothelial Dysfunction in Atherosclerosis. Circulation 2004, 109, III-27–III-32. [Google Scholar] [CrossRef]
- Papadogiannis, D.E.; Protogerou, A.D. Blood pressure variability: A confounder and a cardiovascular risk factor. Hypertens. Res. 2011, 34, 162–163. [Google Scholar] [CrossRef]
- Polonikov, A.V.; Vialykh, E.K.; Churnosov, M.I.; Illig, T.; Freidin, M.B.; Vasil’eva, O.V.; Bushueva, O.Y.; Ryzhaeva, V.N.; Bulgakova, I.V.; Solodilova, M.A. The C718T polymorphism in the 3′-untranslated region of glutathione peroxidase-4 gene is a predictor of cerebral stroke in patients with essential hypertension. Hypertens. Res. 2012, 35, 507–512. [Google Scholar] [CrossRef]
- Leifert, W.R.; Abeywardena, M.Y. Cardioprotective actions of grape polyphenols. Nutr. Res. 2008, 28, 729–737. [Google Scholar] [CrossRef] [PubMed]
- Rasines-Perea, Z.; Teissedre, P.-L. Grape Polyphenols’ Effects in Human Cardiovascular Diseases and Diabetes. Molecules 2017, 22, 68. [Google Scholar] [CrossRef]
- Bozkurt, B.; Aguilar, D.; Deswal, A.; Dunbar, S.B.; Francis, G.S.; Horwich, T.; Jessup, M.; Kosiborod, M.; Pritchett, A.M.; Ramasubbu, K.; et al. Contributory Risk and Management of Comorbidities of Hypertension, Obesity, Diabetes Mellitus, Hyperlipidemia, and Metabolic Syndrome in Chronic Heart Failure: A Scientific Statement From the American Heart Association. Circulation 2016, 134, e535–e578. [Google Scholar] [CrossRef]
- Pirola, L.; Fröjdö, S. Resveratrol: One molecule, many targets. IUBMB Life 2008, 60, 323–332. [Google Scholar] [CrossRef]
- Xia, E.-Q.; Deng, G.-F.; Guo, Y.-J.; Li, H.-B. Biological Activities of Polyphenols from Grapes. Int. J. Mol. Sci. 2010, 11, 622–646. [Google Scholar] [CrossRef]
- Georgiev, V.; Ananga, A.; Tsolova, V. Recent Advances and Uses of Grape Flavonoids as Nutraceuticals. Nutrients 2014, 6, 391–415. [Google Scholar] [CrossRef]
- Pagliaro, B.; Santolamazza, C.; Simonelli, F.; Rubattu, S. Phytochemical Compounds and Protection from Cardiovascular Diseases: A State of the Art. BioMed Res. Int. 2015, 2015, 918069. [Google Scholar] [CrossRef]
- Loffredo, L.; Perri, L.; Nocella, C.; Violi, F. Antioxidant and antiplatelet activity by polyphenol-rich nutrients: Focus on extra virgin olive oil and cocoa. Br. J. Clin. Pharmacol. 2017, 83, 96–102. [Google Scholar] [CrossRef]
- Akinwumi, B.; Bordun, K.-A.; Anderson, H. Biological Activities of Stilbenoids. Int. J. Mol. Sci. 2018, 19, 792. [Google Scholar] [CrossRef]
- Romani, A.; Ieri, F.; Urciuoli, S.; Noce, A.; Marrone, G.; Nediani, C.; Bernini, R. Health Effects of Phenolic Compounds Found in Extra-Virgin Olive Oil, by-Products, and Leaf of Olea europaea L. Nutrients 2019, 11, 1776. [Google Scholar] [CrossRef]
- Tresserra-Rimbau, A.; Rimm, E.B.; Medina-Remón, A.; Martínez-González, M.A.; de la Torre, R.; Corella, D.; Salas-Salvadó, J.; Gómez-Gracia, E.; Lapetra, J.; Arós, F.; et al. Inverse association between habitual polyphenol intake and incidence of cardiovascular events in the PREDIMED study. Nutr. Metab. Cardiovasc. Dis. 2014, 24, 639–647. [Google Scholar] [CrossRef] [PubMed]
- Castaldo, L.; Narváez, A.; Izzo, L.; Graziani, G.; Gaspari, A.; Di Minno, G.; Ritieni, A. Red Wine Consumption and Cardiovascular Health. Molecules 2019, 24, 3626. [Google Scholar] [CrossRef] [PubMed]
- Myasoedova, V.; Kirichenko, T.; Melnichenko, A.; Orekhova, V.; Ravani, A.; Poggio, P.; Sobenin, I.; Bobryshev, Y.; Orekhov, A. Anti-Atherosclerotic Effects of a Phytoestrogen-Rich Herbal Preparation in Postmenopausal Women. Int. J. Mol. Sci. 2016, 17, 1318. [Google Scholar] [CrossRef] [PubMed]
- Zafar, F.; Jahan, N.; Khalil-Ur-Rahman; Khan, A.; Akram, W. Cardioprotective Potential of Polyphenolic Rich Green Combination in Catecholamine Induced Myocardial Necrosis in Rabbits. Evid.-Based Complement. Altern. Med. 2015, 2015, 734903. [Google Scholar] [CrossRef]
- El-Bassossy, H.; Badawy, D.; Neamatallah, T.; Fahmy, A. Ferulic acid, a natural polyphenol, alleviates insulin resistance and hypertension in fructose fed rats: Effect on endothelial-dependent relaxation. Chem. Biol. Interact. 2016, 254, 191–197. [Google Scholar] [CrossRef]
- Ola-Davies, O.E.; Olukole, S.G. Gallic acid protects against bisphenol A-induced alterations in the cardio-renal system of Wistar rats through the antioxidant defense mechanism. Biomed. Pharmacother. 2018, 107, 1786–1794. [Google Scholar] [CrossRef]
- Hu, X.; Wang, H.; Lv, X.; Chu, L.; Liu, Z.; Wei, X.; Chen, Q.; Zhu, L.; Cui, W. Cardioprotective Effects of Tannic Acid on Isoproterenol-Induced Myocardial Injury in Rats: Further Insight into ‘French Paradox’. Phyther. Res. 2015, 29, 1295–1303. [Google Scholar] [CrossRef]
- Ma, D.; Zheng, B.; Du, H.; Han, X.; Zhang, X.; Zhang, J.; Gao, Y.; Sun, S.; Chu, L. The Mechanism Underlying the Protective Effects of Tannic Acid against Isoproterenol-Induced Myocardial Fibrosis in Mice. Front. Pharmacol. 2020, 11, 716. [Google Scholar] [CrossRef]
- Isenburg, J.C.; Simionescu, D.T.; Vyavahare, N.R. Elastin stabilization in cardiovascular implants: Improved resistance to enzymatic degradation by treatment with tannic acid. Biomaterials 2004, 25, 3293–3302. [Google Scholar] [CrossRef]
- Calixto, J.; Nicolau, M.; Rae, G. Pharmacological Actions of Tannic Acid. I. Effects on Isolated Smooth and Cardiac Muscles and on Blood Pressure. Planta Med. 1986, 52, 32–35. [Google Scholar] [CrossRef]
- Zhang, H.; Zhu, S.; Wang, D.; Wei, Y.; Hu, S.-S. Intramyocardial injection of tannic acid attenuates postinfarction remodeling: A novel approach to stabilize the breaking extracellular matrix. J. Thorac. Cardiovasc. Surg. 2009, 137, 216–222.e2. [Google Scholar] [CrossRef] [PubMed]
- Migliori, M.; Cantaluppi, V.; Mannari, C.; Bertelli, A.A.E.; Medica, D.; Quercia, A.D.; Navarro, V.; Scatena, A.; Giovannini, L.; Biancone, L.; et al. Caffeic Acid, a Phenol Found in White Wine, Modulates Endothelial Nitric Oxide Production and Protects from Oxidative Stress-Associated Endothelial Cell Injury. PLoS ONE 2015, 10, e0117530. [Google Scholar] [CrossRef] [PubMed]
- Pataki, T.; Bak, I.; Kovacs, P.; Bagchi, D.; Das, D.K.; Tosaki, A. Grape seed proanthocyanidins improved cardiac recovery during reperfusion after ischemia in isolated rat hearts. Am. J. Clin. Nutr. 2002, 75, 894–899. [Google Scholar] [CrossRef]
- Sergazy, S.; Shulgau, Z.; Fedotovskikh, G.; Chulenbayeva, L.; Nurgozhina, A.; Nurgaziyev, M.; Krivyh, E.; Kamyshanskiy, Y.; Kushugulova, A.; Gulyayev, A.; et al. Cardioprotective effect of grape polyphenol extract against doxorubicin induced cardiotoxicity. Sci. Rep. 2020, 10, 14720. [Google Scholar] [CrossRef]
- Stote, K.S.; Sweeney, M.I.; Kean, T.; Baer, D.J.; Novotny, J.A.; Shakerley, N.L.; Chandrasekaran, A.; Carrico, P.M.; Melendez, J.A.; Gottschall-Pass, K.T. The effects of 100% wild blueberry (Vaccinium angustifolium) juice consumption on cardiometablic biomarkers: A randomized, placebo-controlled, crossover trial in adults with increased risk for type 2 diabetes. BMC Nutr. 2017, 3, 45. [Google Scholar] [CrossRef]
- Ballmann, C.; Denney, T.; Beyers, R.J.; Quindry, T.; Romero, M.; Selsby, J.T.; Quindry, J.C. Long-term dietary quercetin enrichment as a cardioprotective countermeasure in mdx mice. Exp. Physiol. 2017, 102, 635–649. [Google Scholar] [CrossRef]
- Ganapathy, R.; Ramachandran, A.; Shivalingaiah, S.B.; Bishir, M.; Bhojaraj, S.; Sridhar, S.; Mohan, S.K.; Veeraraghavan, V.P.; Chidambaram, S.B.; Essa, M.M.; et al. Cardioprotective potential of polyphenols rich Thraatchathi Chooranam against isoproterenol induced myocardial necrosis in experimental rats. BMC Complement. Med. Ther. 2020, 20, 356. [Google Scholar] [CrossRef]
- Sharifi-Rad, J.; Rodrigues, C.F.; Sharopov, F.; Docea, A.O.; Can Karaca, A.; Sharifi-Rad, M.; Kahveci Karıncaoglu, D.; Gülseren, G.; Şenol, E.; Demircan, E.; et al. Diet, Lifestyle and Cardiovascular Diseases: Linking Pathophysiology to Cardioprotective Effects of Natural Bioactive Compounds. Int. J. Environ. Res. Public Health 2020, 17, 2326. [Google Scholar] [CrossRef]
- Yochum, L.; Kushi, L.H.; Meyer, K.; Folsom, A.R. Dietary flavonoid intake and risk of cardiovascular disease in postmenopausal women. Climacteric 1999, 2, 237–238. [Google Scholar] [CrossRef]
- Qin, Y.; Shu, F.; Zeng, Y.; Meng, X.; Wang, B.; Diao, L.; Wang, L.; Wan, J.; Zhu, J.; Wang, J.; et al. Daidzein Supplementation Decreases Serum Triglyceride and Uric Acid Concentrations in Hypercholesterolemic Adults with the Effect on Triglycerides Being Greater in Those with the GA Compared with the GG Genotype of ESR-β RsaI. J. Nutr. 2014, 144, 49–54. [Google Scholar] [CrossRef]
- Hao, J.; Kim, C.-H.; Ha, T.-S.; Ahn, H.-Y. Epigallocatechin-3 gallate prevents cardiac hypertrophy induced by pressure overload in rats. J. Vet. Sci. 2007, 8, 121. [Google Scholar] [CrossRef] [PubMed]
- Xuan, F.; Jian, J. Epigallocatechin gallate exerts protective effects against myocardial ischemia/reperfusion injury through the PI3K/Akt pathway-mediated inhibition of apoptosis and the restoration of the autophagic flux. Int. J. Mol. Med. 2016, 38, 328–336. [Google Scholar] [CrossRef] [PubMed]
- Villarreal-Calderon, R.; Reed, W.; Palacios-Moreno, J.; Keefe, S.; Herritt, L.; Brooks, D.; Torres-Jardón, R.; Calderón-Garcidueñas, L. Urban air pollution produces up-regulation of myocardial inflammatory genes and dark chocolate provides cardioprotection. Exp. Toxicol. Pathol. 2012, 64, 297–306. [Google Scholar] [CrossRef] [PubMed]
- Davis, D.W.; Tallent, R.; Navalta, J.W.; Salazar, A.; Lyons, T.J.; Basu, A. Effects of Acute Cocoa Supplementation on Postprandial Apolipoproteins, Lipoprotein Subclasses, and Inflammatory Biomarkers in Adults with Type 2 Diabetes after a High-Fat Meal. Nutrients 2020, 12, 1902. [Google Scholar] [CrossRef]
- Poudyal, H.; Campbell, F.; Brown, L. Olive Leaf Extract Attenuates Cardiac, Hepatic, and Metabolic Changes in High Carbohydrate–, High Fat–Fed Rats. J. Nutr. 2010, 140, 946–953. [Google Scholar] [CrossRef]
- Kurowska, E.M.; Banh, C.; Hasegawa, S.; Manners, G.D. Regulation of Apo B Production in HepG2 Cells by Citrus Limonoids. In Citrus Limonoids—Functional Chemicals in Agriculture and Food; ACS Publications: Washington, DC, USA, 2000; pp. 175–184. [Google Scholar]
- Negrão, M.R.; Keating, E.; Faria, A.; Azevedo, I.; Martins, M.J. Acute Effect of Tea, Wine, Beer, and Polyphenols on ecto-Alkaline Phosphatase Activity in Human Vascular Smooth Muscle Cells. J. Agric. Food Chem. 2006, 54, 4982–4988. [Google Scholar] [CrossRef]
- Hu, Q.; Zhang, T.; Yi, L.; Zhou, X.; Mi, M. Dihydromyricetin inhibits NLRP3 inflammasome-dependent pyroptosis by activating the Nrf2 signaling pathway in vascular endothelial cells. BioFactors 2018, 44, 123–136. [Google Scholar] [CrossRef]
- Zeng, Y.; Peng, Y.; Tang, K.; Wang, Y.Q.; Zhao, Z.Y.; Wei, X.Y.; Xu, X. Le Dihydromyricetin ameliorates foam cell formation via LXRα-ABCA1/ABCG1-dependent cholesterol efflux in macrophages. Biomed. Pharmacother. 2018, 101, 543–552. [Google Scholar] [CrossRef]
- Wang, Z.; Huang, Y.; Zou, J.; Cao, K.; Xu, Y.; Wu, J.M. Effects of red wine and wine polyphenol resveratrol on platelet aggregation in vivo and in vitro. Int. J. Mol. Med. 2002, 9, 77–79. [Google Scholar] [CrossRef]
- Pal, S.; Ho, N.; Santos, C.; Dubois, P.; Mamo, J.; Croft, K.; Allister, E. Red Wine Polyphenolics Increase LDL Receptor Expression and Activity and Suppress the Secretion of ApoB100 from Human HepG2 Cells. J. Nutr. 2003, 133, 700–706. [Google Scholar] [CrossRef]
- Soares Filho, P.R.; Castro, I.; Stahlschmidt, A. Efeito do vinho tinto associado ao exercício físico no sistema cardiovascular de ratos espontaneamente hipertensos. Arq. Bras. Cardiol. 2011, 96, 277–283. [Google Scholar] [CrossRef]
- Ungvari, Z.; Orosz, Z.; Rivera, A.; Labinskyy, N.; Xiangmin, Z.; Olson, S.; Podlutsky, A.; Csiszar, A. Resveratrol increases vascular oxidative stress resistance. Am. J. Physiol. Circ. Physiol. 2007, 292, H2417–H2424. [Google Scholar] [CrossRef]
- Hung, L.-M.; Su, M.-J.; Chu, W.-K.; Chiao, C.-W.; Chan, W.-F.; Chen, J.-K. The protective effect of resveratrols on ischaemia-reperfusion injuries of rat hearts is correlated with antioxidant efficacy. Br. J. Pharmacol. 2002, 135, 1627–1633. [Google Scholar] [CrossRef]
- Yoshida, Y.; Shioi, T.; Izumi, T. Resveratrol Ameliorates Experimental Autoimmune Myocarditis. Circ. J. 2007, 71, 397–404. [Google Scholar] [CrossRef]
- Lou, Y.; Wang, Z.; Xu, Y.; Zhou, P.; Cao, J.; Li, Y.; Chen, Y.; Sun, J.; Fu, L. Resveratrol prevents doxorubicin-induced cardiotoxicity in H9c2 cells through the inhibition of endoplasmic reticulum stress and the activation of the Sirt1 pathway. Int. J. Mol. Med. 2015, 36, 873–880. [Google Scholar] [CrossRef]
- Al-Harthi, S.E.; Alarabi, O.M.; Ramadan, W.S.; Alaama, M.N.; Al-Kreathy, H.M.; Damanhouri, Z.A.; Khan, L.M.; Osman, A.-M.M. Amelioration of doxorubicin-induced cardiotoxicity by resveratrol. Mol. Med. Rep. 2014, 10, 1455–1460. [Google Scholar] [CrossRef]
- Sabe, A.A.; Elmadhun, N.Y.; Dalal, R.S.; Robich, M.P.; Sellke, F.W. Resveratrol regulates autophagy signaling in chronically ischemic myocardium. J. Thorac. Cardiovasc. Surg. 2014, 147, 792–799. [Google Scholar] [CrossRef]
- Carolo dos Santos, K.; Pereira Braga, C.; Octavio Barbanera, P.; Rodrigues Ferreira Seiva, F.; Fernandes Junior, A.; Fernandes, A.A.H. Cardiac Energy Metabolism and Oxidative Stress Biomarkers in Diabetic Rat Treated with Resveratrol. PLoS ONE 2014, 9, e102775. [Google Scholar] [CrossRef]
- Akinwumi, B.; Raj, P.; Lee, D.; Acosta, C.; Yu, L.; Thomas, S.; Nagabhushanam, K.; Majeed, M.; Davies, N.; Netticadan, T.; et al. Disparate Effects of Stilbenoid Polyphenols on Hypertrophic Cardiomyocytes In Vitro vs. in the Spontaneously Hypertensive Heart Failure Rat. Molecules 2017, 22, 204. [Google Scholar] [CrossRef]
- Fan, Y.; Liu, L.; Fang, K.; Huang, T.; Wan, L.; Liu, Y.; Zhang, S.; Yan, D.; Li, G.; Gao, Y.; et al. Resveratrol Ameliorates Cardiac Hypertrophy by Down-regulation of miR-155 Through Activation of Breast Cancer Type 1 Susceptibility Protein. J. Am. Heart Assoc. 2016, 5, e002648. [Google Scholar] [CrossRef]
- Chen, W.-P.; Hung, L.-M.; Hsueh, C.-H.; Lai, L.-P.; Su, M.-J. Piceatannol, a derivative of resveratrol, moderately slows INa inactivation and exerts antiarrhythmic action in ischaemia-reperfused rat hearts. Br. J. Pharmacol. 2009, 157, 381–391. [Google Scholar] [CrossRef]
- Wang, D.; Zhang, Y.; Zhang, C.; Gao, L.; Li, J. Piceatannol pretreatment alleviates acute cardiac injury via regulating PI3K-Akt-eNOS signaling in H9c2 cells. Biomed. Pharmacother. 2019, 109, 886–891. [Google Scholar] [CrossRef]
- Zanwar, A.; Hegde, M.; Bodhankar, S. Cardioprotective activity of flax lignan concentrate extracted from seeds of Linum usitatissimum in isoprenalin induced myocardial necrosis in rats. Interdiscip. Toxicol. 2011, 4, 90–97. [Google Scholar] [CrossRef]
- Parikh, M.; Kura, B.; O’Hara, K.A.; Dibrov, E.; Netticadan, T.; Slezak, J.; Pierce, G.N. Cardioprotective Effects of Dietary Flaxseed Post-Infarction Are Associated with Changes in MicroRNA Expression. Biomolecules 2020, 10, 1297. [Google Scholar] [CrossRef]
- Zanwar, A.A.; Hegde, M.V.; Bodhankar, S.L. Protective role of concomitant administration of flax lignan concentrate and omega-3-fatty acid on myocardial damage in doxorubicin-induced cardiotoxicity. Food Sci. Hum. Wellness 2013, 2, 29–38. [Google Scholar] [CrossRef][Green Version]
- Sawant, S.H.; Bodhankar, S.L. Flax lignan concentrate reverses alterations in blood pressure, left ventricular functions, lipid profile and antioxidant status in DOCA-salt induced renal hypertension in rats. Ren. Fail. 2016, 38, 411–423. [Google Scholar] [CrossRef]
- Bassett, C.M.C.; Rodriguez-Leyva, D.; Pierce, G.N. Experimental and clinical research findings on the cardiovascular benefits of consuming flaxseed. Appl. Physiol. Nutr. Metab. 2009, 34, 965–974. [Google Scholar] [CrossRef]
- Hassan, N.A.; El-Bassossy, H.M.; Mahmoud, M.F.; Fahmy, A. Caffeic acid phenethyl ester, a 5-lipoxygenase enzyme inhibitor, alleviates diabetic atherosclerotic manifestations: Effect on vascular reactivity and stiffness. Chem. Biol. Interact. 2014, 213, 28–36. [Google Scholar] [CrossRef]
- Gun, A.; Ozer, M.K.; Bilgic, S.; Kocaman, N.; Ozan, G. Effect of Caffeic Acid Phenethyl Ester on Vascular Damage Caused by Consumption of High Fructose Corn Syrup in Rats. Oxid. Med. Cell. Longev. 2016, 2016, 3419479. [Google Scholar] [CrossRef]
- Wang, Y.; Kaur, G.; Kumar, M.; Kushwah, A.S.; Kabra, A.; Kainth, R. Caffeic Acid Prevents Vascular Oxidative Stress and Atherosclerosis against Atherosclerogenic Diet in Rats. Evid.-Based Complement. Altern. Med. 2022, 2022, 8913926. [Google Scholar] [CrossRef]
- Dupasquier, C.M.C.; Dibrov, E.; Kneesh, A.L.; Cheung, P.K.M.; Lee, K.G.Y.; Alexander, H.K.; Yeganeh, B.K.; Moghadasian, M.H.; Pierce, G.N. Dietary flaxseed inhibits atherosclerosis in the LDL receptor-deficient mouse in part through antiproliferative and anti-inflammatory actions. Am. J. Physiol. Circ. Physiol. 2007, 293, H2394–H2402. [Google Scholar] [CrossRef] [PubMed]
- Bloedon, L.T.; Balikai, S.; Chittams, J.; Cunnane, S.C.; Berlin, J.A.; Rader, D.J.; Szapary, P.O. Flaxseed and Cardiovascular Risk Factors: Results from a Double Blind, Randomized, Controlled Clinical Trial. J. Am. Coll. Nutr. 2008, 27, 65–74. [Google Scholar] [CrossRef]
- Narasimhulu, C.A.; Selvarajan, K.; Litvinov, D.; Parthasarathy, S. Anti-Atherosclerotic and Anti-Inflammatory Actions of Sesame Oil. J. Med. Food 2015, 18, 11–20. [Google Scholar] [CrossRef] [PubMed]
- Bhaskaran, S.; Santanam, N.; Penumetcha, M.; Parthasarathy, S. Inhibition of Atherosclerosis in Low-Density Lipoprotein Receptor-Negative Mice by Sesame Oil. J. Med. Food 2006, 9, 487–490. [Google Scholar] [CrossRef] [PubMed]
- Hemalatha, G.; Pugalendi, K.V.; Saravanan, R. Modulatory effect of sesamol on DOCA-salt-induced oxidative stress in uninephrectomized hypertensive rats. Mol. Cell. Biochem. 2013, 379, 255–265. [Google Scholar] [CrossRef] [PubMed]
- Stanely Mainzen Prince, P.; Priscilla, H.; Devika, P.T. Gallic acid prevents lysosomal damage in isoproterenol induced cardiotoxicity in Wistar rats. Eur. J. Pharmacol. 2009, 615, 139–143. [Google Scholar] [CrossRef]
- Umadevi, S.; Gopi, V.; Elangovan, V. Regulatory mechanism of gallic acid against advanced glycation end products induced cardiac remodeling in experimental rats. Chem. Biol. Interact. 2014, 208, 28–36. [Google Scholar] [CrossRef] [PubMed]
- Jin, L.; Lin, M.Q.; Piao, Z.H.; Cho, J.Y.; Kim, G.R.; Choi, S.Y.; Ryu, Y.; Sun, S.; Kee, H.J.; Jeong, M.H. Gallic acid attenuates hypertension, cardiac remodeling, and fibrosis in mice with N G-nitro-L-arginine methyl ester-induced hypertension via regulation of histone deacetylase 1 or histone deacetylase 2. J. Hypertens. 2017, 35, 1502–1512. [Google Scholar] [CrossRef]
- Sies, H. Oxidative stress: A concept in redox biology and medicine. Redox Biol. 2015, 4, 180–183. [Google Scholar] [CrossRef]
- Zhao, X.; Wu, Y. Correlations of Silent Information Regulator of Transcription 1 (SIRT1) Expression, Inflammatory Factors, and Oxidative Stress with Pulmonary Function in Patients with Acute Exacerbation of Chronic Obstructive Pulmonary Disease (AECOPD). Med. Sci. Monit. 2021, 27, e929046. [Google Scholar] [CrossRef]
- Kirkham, P.A.; Barnes, P.J. Oxidative Stress in COPD. Chest 2013, 144, 266–273. [Google Scholar] [CrossRef]
- Rivero, D.; Pérez-Magariño, S.; González-Sanjosé, M.L.; Valls-Belles, V.; Codoñer, P.; Muñiz, P. Inhibition of Induced DNA Oxidative Damage by Beers: Correlation with the Content of Polyphenols and Melanoidins. J. Agric. Food Chem. 2005, 53, 3637–3642. [Google Scholar] [CrossRef] [PubMed]
- Perron, N.R.; Hodges, J.N.; Jenkins, M.; Brumaghim, J.L. Predicting How Polyphenol Antioxidants Prevent DNA Damage by Binding to Iron. Inorg. Chem. 2008, 47, 6153–6161. [Google Scholar] [CrossRef]
- Bellion, P.; Digles, J.; Will, F.; Dietrich, H.; Baum, M.; Eisenbrand, G.; Janzowski, C. Polyphenolic Apple Extracts: Effects of Raw Material and Production Method on Antioxidant Effectiveness and Reduction of DNA Damage in Caco-2 Cells. J. Agric. Food Chem. 2010, 58, 6636–6642. [Google Scholar] [CrossRef]
- Loeb, L.A.; Wallace, D.C.; Martin, G.M. The mitochondrial theory of aging and its relationship to reactive oxygen species damage and somatic mtDNA mutations. Proc. Natl. Acad. Sci. USA 2005, 102, 18769–18770. [Google Scholar] [CrossRef]
- Barrero, A.F.; Herrador, M.M.; Arteaga, P.; Rosas-Romero, A.; Arteaga, J.F. Antioxidant Activity of Diterpenes and Polyphenols from Ophryosporus heptanthus. J. Agric. Food Chem. 2006, 54, 2537–2542. [Google Scholar] [CrossRef]
- Dulebohn, R.V.; Yi, W.; Srivastava, A.; Akoh, C.C.; Krewer, G.; Fischer, J.G. Effects of Blueberry (Vaccinium ashei) on DNA Damage, Lipid Peroxidation, and Phase II Enzyme Activities in Rats. J. Agric. Food Chem. 2008, 56, 11700–11706. [Google Scholar] [CrossRef]
- Natella, F.; Nardini, M.; Giannetti, I.; Dattilo, C.; Scaccini, C. Coffee Drinking Influences Plasma Antioxidant Capacity in Humans. J. Agric. Food Chem. 2002, 50, 6211–6216. [Google Scholar] [CrossRef]
- Rolt, A.; Cox, L.S. Structural basis of the anti-ageing effects of polyphenolics: Mitigation of oxidative stress. BMC Chem. 2020, 14, 50. [Google Scholar] [CrossRef] [PubMed]
- Li, D.; Rui, Y.; Guo, S.; Luan, F.; Liu, R.; Zeng, N. Ferulic acid: A review of its pharmacology, pharmacokinetics and derivatives. Life Sci. 2021, 284, 119921. [Google Scholar] [CrossRef]
- Hosseinzadeh, A.; Houshmand, G.; Goudarzi, M.; Sezavar, S.H.; Mehrzadi, S.; Mansouri, E.; Kalantar, M. Ameliorative effect of gallic acid on sodium arsenite-induced spleno-, cardio- and hemato-toxicity in rats. Life Sci. 2019, 217, 91–100. [Google Scholar] [CrossRef] [PubMed]
- Murakami, A.; Ashida, H.; Terao, J. Multitargeted cancer prevention by quercetin. Cancer Lett. 2008, 269, 315–325. [Google Scholar] [CrossRef] [PubMed]
- Kamada, C.; da Silva, E.L.; Ohnishi-Kameyama, M.; Moon, J.-H.; Terao, J. Attenuation of lipid peroxidation and hyperlipidemia by quercetin glucoside in the aorta of high cholesterol-fed rabbit. Free Radic. Res. 2005, 39, 185–194. [Google Scholar] [CrossRef] [PubMed]
- Youdim, K.A.; Shukitt-Hale, B.; Joseph, J.A. Flavonoids and the brain: Interactions at the blood–brain barrier and their physiological effects on the central nervous system. Free Radic. Biol. Med. 2004, 37, 1683–1693. [Google Scholar] [CrossRef]
- Ishisaka, A.; Ichikawa, S.; Sakakibara, H.; Piskula, M.K.; Nakamura, T.; Kato, Y.; Ito, M.; Miyamoto, K.; Tsuji, A.; Kawai, Y.; et al. Accumulation of orally administered quercetin in brain tissue and its antioxidative effects in rats. Free Radic. Biol. Med. 2011, 51, 1329–1336. [Google Scholar] [CrossRef]
- Sanbongi, C.; Osakabe, N.; Natsume, M.; Takizawa, T.; Gomi, S.; Osawa, T. Antioxidative Polyphenols Isolated from Theobroma cacao. J. Agric. Food Chem. 1998, 46, 454–457. [Google Scholar] [CrossRef]
- Burgos-Morón, E.; Calderón-Montaño, J.M.; Orta, M.L.; Pastor, N.; Pérez-Guerrero, C.; Austin, C.; Mateos, S.; López-Lázaro, M. The Coffee Constituent Chlorogenic Acid Induces Cellular DNA Damage and Formation of Topoisomerase I– and II–DNA Complexes in Cells. J. Agric. Food Chem. 2012, 60, 7384–7391. [Google Scholar] [CrossRef]
- Esselen, M.; Fritz, J.; Hutter, M.; Marko, D. Delphinidin Modulates the DNA-Damaging Properties of Topoisomerase II Poisons. Chem. Res. Toxicol. 2009, 22, 554–564. [Google Scholar] [CrossRef]
- Esselen, M.; Boettler, U.; Teller, N.; Bächler, S.; Hutter, M.; Rüfer, C.E.; Skrbek, S.; Marko, D. Anthocyanin-Rich Blackberry Extract Suppresses the DNA-Damaging Properties of Topoisomerase I and II Poisons in Colon Carcinoma Cells. J. Agric. Food Chem. 2011, 59, 6966–6973. [Google Scholar] [CrossRef]
- Shih, P.-H.; Yeh, C.-T.; Yen, G.-C. Anthocyanins Induce the Activation of Phase II Enzymes through the Antioxidant Response Element Pathway against Oxidative Stress-Induced Apoptosis. J. Agric. Food Chem. 2007, 55, 9427–9435. [Google Scholar] [CrossRef]
- Hwang, J.-T.; Kwon, D.Y.; Park, O.J.; Kim, M.S. Resveratrol protects ROS-induced cell death by activating AMPK in H9c2 cardiac muscle cells. Genes Nutr. 2008, 2, 323–326. [Google Scholar] [CrossRef]
- Haramizu, S.; Asano, S.; Butler, D.C.; Stanton, D.A.; Hajira, A.; Mohamed, J.S.; Alway, S.E. Dietary resveratrol confers apoptotic resistance to oxidative stress in myoblasts. J. Nutr. Biochem. 2017, 50, 103–115. [Google Scholar] [CrossRef]
- Du, L.; Wang, L.; Wang, B.; Wang, J.; Hao, M.; Chen, Y.; Li, X.; Li, Y.; Jiang, Y.; Li, C.; et al. A novel compound AB38b attenuates oxidative stress and ECM protein accumulation in kidneys of diabetic mice through modulation of Keap1/Nrf2 signaling. Acta Pharmacol. Sin. 2020, 41, 358–372. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Chen, X.; Li, J. Resveratrol protects against oxidized low-density lipoprotein-induced human umbilical vein endothelial cell apoptosis via inhibition of mitochondrial-derived oxidative stress. Mol. Med. Rep. 2017, 15, 2457–2464. [Google Scholar] [CrossRef] [PubMed]
- Soylemez, S.; Sepici, A.; Akar, F. Resveratrol Supplementation Gender Independently Improves Endothelial Reactivity and Suppresses Superoxide Production in Healthy Rats. Cardiovasc. Drugs Ther. 2009, 23, 449–458. [Google Scholar] [CrossRef] [PubMed]
- Youssef, F.S.; Ashour, M.L.; El-Beshbishy, H.A.; Ahmed Hamza, A.; Singab, A.N.B.; Wink, M. Pinoresinol-4-O-β-D-glucopyranoside: A lignan from prunes (Prunus domestica) attenuates oxidative stress, hyperglycaemia and hepatic toxicity in vitro and in vivo. J. Pharm. Pharmacol. 2020, 72, 1830–1839. [Google Scholar] [CrossRef]
- Pilar, B.; Güllich, A.; Oliveira, P.; Ströher, D.; Piccoli, J.; Manfredini, V. Protective Role of Flaxseed Oil and Flaxseed Lignan Secoisolariciresinol Diglucoside against Oxidative Stress in Rats with Metabolic Syndrome. J. Food Sci. 2017, 82, 3029–3036. [Google Scholar] [CrossRef]
- Kim, J.W.; Yang, H.; Kim, H.W.; Kim, H.P.; Sung, S.H. Lignans from Opuntia ficus-indica seeds protect rat primary hepatocytes and HepG2 cells against ethanol-induced oxidative stress. Biosci. Biotechnol. Biochem. 2017, 81, 181–183. [Google Scholar] [CrossRef]
- Youssef, F.S.; Menze, E.T.; Ashour, M.L. A Potent Lignan from Prunes Alleviates Inflammation and Oxidative Stress in Lithium/Pilocarpine-Induced Epileptic Seizures in Rats. Antioxidants 2020, 9, 575. [Google Scholar] [CrossRef]
- Halazonetis, T.D.; Gorgoulis, V.G.; Bartek, J. An Oncogene-Induced DNA Damage Model for Cancer Development. Science 2008, 319, 1352–1355. [Google Scholar] [CrossRef]
- O’Connor, M.J. Targeting the DNA Damage Response in Cancer. Mol. Cell 2015, 60, 547–560. [Google Scholar] [CrossRef]
- Roos, W.P.; Thomas, A.D.; Kaina, B. DNA damage and the balance between survival and death in cancer biology. Nat. Rev. Cancer 2016, 16, 20–33. [Google Scholar] [CrossRef]
- Ali, R.; Rakha, E.A.; Madhusudan, S.; Bryant, H.E. DNA damage repair in breast cancer and its therapeutic implications. Pathology 2017, 49, 156–165. [Google Scholar] [CrossRef]
- Hosoya, N.; Miyagawa, K. Targeting DNA damage response in cancer therapy. Cancer Sci. 2014, 105, 370–388. [Google Scholar] [CrossRef]
- Panahi, Y.; Saadat, A.; Beiraghdar, F.; Sahebkar, A. Adjuvant Therapy with Bioavailability-Boosted Curcuminoids Suppresses Systemic Inflammation and Improves Quality of Life in Patients with Solid Tumors: A Randomized Double-Blind Placebo-Controlled Trial. Phyther. Res. 2014, 28, 1461–1467. [Google Scholar] [CrossRef]
- Azqueta, A.; Collins, A. Polyphenols and DNA Damage: A Mixed Blessing. Nutrients 2016, 8, 785. [Google Scholar] [CrossRef]
- Shukla, D.; Nandi, N.K.; Singh, B.; Singh, A.; Kumar, B.; Narang, R.K.; Singh, C. Ferulic acid-loaded drug delivery systems for biomedical applications. J. Drug Deliv. Sci. Technol. 2022, 75, 103621. [Google Scholar] [CrossRef]
- Agarwal, C.; Tyagi, A.; Agarwal, R. Gallic acid causes inactivating phosphorylation of cdc25A/cdc25C-cdc2 via ATM-Chk2 activation, leading to cell cycle arrest, and induces apoptosis in human prostate carcinoma DU145 cells. Mol. Cancer Ther. 2006, 5, 3294–3302. [Google Scholar] [CrossRef]
- Ji, B.-C.; Hsu, W.-H.; Yang, J.-S.; Hsia, T.-C.; Lu, C.-C.; Chiang, J.-H.; Yang, J.-L.; Lin, C.-H.; Lin, J.-J.; Suen, L.-J.W.; et al. Gallic Acid Induces Apoptosis via Caspase-3 and Mitochondrion-Dependent Pathways in Vitro and Suppresses Lung Xenograft Tumor Growth In Vivo. J. Agric. Food Chem. 2009, 57, 7596–7604. [Google Scholar] [CrossRef]
- Liu, K.-C.; Ho, H.-C.; Huang, A.-C.; Ji, B.-C.; Lin, H.-Y.; Chueh, F.-S.; Yang, J.-S.; Lu, C.-C.; Chiang, J.-H.; Meng, M.; et al. Gallic acid provokes DNA damage and suppresses DNA repair gene expression in human prostate cancer PC-3 cells. Environ. Toxicol. 2013, 28, 579–587. [Google Scholar] [CrossRef] [PubMed]
- Fujiki, H.; Suganuma, M.; Okabe, S.; Sueoka, E.; Suga, K.; Imai, K.; Nakachi, K.; Kimura, S. Mechanistic Findings of Green Tea as Cancer Preventive for Humans. Exp. Biol. Med. 1999, 220, 225–228. [Google Scholar] [CrossRef]
- Mertens-Talcott, S.U.; Lee, J.-H.; Percival, S.S.; Talcott, S.T. Induction of Cell Death in Caco-2 Human Colon Carcinoma Cells by Ellagic Acid Rich Fractions from Muscadine Grapes (Vitis rotundifolia). J. Agric. Food Chem. 2006, 54, 5336–5343. [Google Scholar] [CrossRef]
- Luo, K.L.; Luo, J.-H.; Yu, Y.P. (−)-Epigallocatechin-3-gallate induces Du145 prostate cancer cell death via downregulation of inhibitor of DNA binding 2, a dominant negative helix-loop-helix protein. Cancer Sci. 2010, 101, 707–712. [Google Scholar] [CrossRef] [PubMed]
- Khan, N.; Mukhtar, H. Modulation of signaling pathways in prostate cancer by green tea polyphenols. Biochem. Pharmacol. 2013, 85, 667–672. [Google Scholar] [CrossRef]
- Zhou, J.; Xia, L.; Zhang, Y. Naringin inhibits thyroid cancer cell proliferation and induces cell apoptosis through repressing PI3K/AKT pathway. Pathol.-Res. Pract. 2019, 215, 152707. [Google Scholar] [CrossRef]
- Wu, X.; Huang, Z.; Liu, J.; Chen, Y.; Huang, H.; He, Y.; Li, D.; Zhang, L.; Du, Z.; Zhang, K.; et al. Effects and mechanism of inhibition of naringin in combination with atorvastatin on prostate cancer cells in vitro and in vivo. Phytochem. Lett. 2019, 32, 168–176. [Google Scholar] [CrossRef]
- Li, N.; Sun, C.; Zhou, B.; Xing, H.; Ma, D.; Chen, G.; Weng, D. Low Concentration of Quercetin Antagonizes the Cytotoxic Effects of Anti-Neoplastic Drugs in Ovarian Cancer. PLoS ONE 2014, 9, e100314. [Google Scholar] [CrossRef]
- Parvaresh, A.; Razavi, R.; Rafie, N.; Ghiasvand, R.; Pourmasoumi, M.; Miraghajani, M. Quercetin and ovarian cancer: An evaluation based on a systematic review. J. Res. Med. Sci. 2016, 21, 34. [Google Scholar] [CrossRef]
- Chen, R.; Hollborn, M.; Grosche, A.; Reichenbach, A.; Wiedemann, P.; Bringmann, A.; Kohen, L. Effects of the vegetable polyphenols epigallocatechin-3-gallate, luteolin, apigenin, myricetin, quercetin, and cyanidin in primary cultures of human retinal pigment epithelial cells. Mol. Vis. 2014, 20, 242–258. [Google Scholar]
- Rauf, A.; Imran, M.; Khan, I.A.; Ur-Rehman, M.; Gilani, S.A.; Mehmood, Z.; Mubarak, M.S. Anticancer potential of quercetin: A comprehensive review. Phyther. Res. 2018, 32, 2109–2130. [Google Scholar] [CrossRef]
- Schantz, M.; Mohn, C.; Baum, M.; Richling, E. Antioxidative efficiency of an anthocyanin rich bilberry extract in the human colon tumor cell lines Caco-2 and HT-29. J. Berry Res. 2010, 1, 25–33. [Google Scholar] [CrossRef]
- Joshi, R.; Rana, A.; Kumar, V.; Kumar, D.; Padwad, Y.S.; Yadav, S.K.; Gulati, A. Anthocyanins enriched purple tea exhibits antioxidant, immunostimulatory and anticancer activities. J. Food Sci. Technol. 2017, 54, 1953–1963. [Google Scholar] [CrossRef] [PubMed]
- Faria, A.; Pestana, D.; Teixeira, D.; de Freitas, V.; Mateus, N.; Calhau, C. Blueberry anthocyanins and pyruvic acid adducts: Anticancer properties in breast cancer cell lines. Phyther. Res. 2010, 24, 1862–1869. [Google Scholar] [CrossRef] [PubMed]
- Suner, S.S.; Sahiner, M.; Mohapatra, S.; Ayyala, R.S.; Bhethanabotla, V.R.; Sahiner, N. Degradable poly(catechin) nanoparticles as a versatile therapeutic agent. Int. J. Polym. Mater. Polym. Biomater. 2022, 71, 1104–1115. [Google Scholar] [CrossRef]
- Igura, K.; Ohta, T.; Kuroda, Y.; Kaji, K. Resveratrol and quercetin inhibit angiogenesis in vitro. Cancer Lett. 2001, 171, 11–16. [Google Scholar] [CrossRef]
- Banerjee, S.; Bueso-Ramos, C.; Aggarwal, B.B. Suppression of 7,12-dimethylbenz(a)anthracene-induced mammary carcinogenesis in rats by resveratrol: Role of nuclear factor-kappaB, cyclooxygenase 2, and matrix metalloprotease 9. Cancer Res. 2002, 62, 4945–4954. [Google Scholar]
- Cao, Y.; Fu, Z.-D.; Wang, F.; Liu, H.-Y.; Han, R. Anti-angiogenic activity of resveratrol, a natural compound from medicinal plants. J. Asian Nat. Prod. Res. 2005, 7, 205–213. [Google Scholar] [CrossRef]
- Kim, T.; Park, J.; Woo, J. Resveratrol induces cell death through ROS-dependent downregulation of Notch1/PTEN/Akt signaling in ovarian cancer cells. Mol. Med. Rep. 2019, 19, 3353–3360. [Google Scholar] [CrossRef]
- Miki, H.; Uehara, N.; Kimura, A.; Sasaki, T.; Yuri, T.; Yoshizawa, K.; Tsubura, A. Resveratrol induces apoptosis via ROS-triggered autophagy in human colon cancer cells. Int. J. Oncol. 2012, 40, 1020–1028. [Google Scholar] [CrossRef]
- Luo, H.; Yang, A.; Schulte, B.A.; Wargovich, M.J.; Wang, G.Y. Resveratrol Induces Premature Senescence in Lung Cancer Cells via ROS-Mediated DNA Damage. PLoS ONE 2013, 8, e60065. [Google Scholar] [CrossRef]
- Xu, Y.; Liu, Z.; Sun, J.; Pan, Q.; Sun, F.; Yan, Z.; Hu, X. Schisandrin B Prevents Doxorubicin-Induced Chronic Cardiotoxicity and Enhances Its Anticancer Activity In Vivo. PLoS ONE 2011, 6, e28335. [Google Scholar] [CrossRef] [PubMed]
- Hsieh, C.-J.; Kuo, P.-L.; Hsu, Y.-C.; Huang, Y.-F.; Tsai, E.-M.; Hsu, Y.-L. Arctigenin, a dietary phytoestrogen, induces apoptosis of estrogen receptor-negative breast cancer cells through the ROS/p38 MAPK pathway and epigenetic regulation. Free Radic. Biol. Med. 2014, 67, 159–170. [Google Scholar] [CrossRef] [PubMed]
- Zingue, S.; Cisilotto, J.; Tueche, A.B.; Bishayee, A.; Mefegue, F.A.; Sandjo, L.P.; Magne Nde, C.B.; Winter, E.; Michel, T.; Ndinteh, D.T.; et al. Crateva adansonii DC, an African ethnomedicinal plant, exerts cytotoxicity in vitro and prevents experimental mammary tumorigenesis in vivo. J. Ethnopharmacol. 2016, 190, 183–199. [Google Scholar] [CrossRef]
- Zhang, T.; Ma, L.; Wu, P.; Li, W.; Li, T.; Gu, R.; Dan, X.; Li, Z.; Fan, X.; Xiao, Z. Gallic acid has anticancer activity and enhances the anticancer effects of cisplatin in non-small cell lung cancer A549 cells via the JAK/STAT3 signaling pathway. Oncol. Rep. 2019, 41, 1779–1788. [Google Scholar] [CrossRef]
- De Camargo, A.C.; Regitano-d’Arce, M.A.B.; Biasoto, A.C.T.; Shahidi, F. Low Molecular Weight Phenolics of Grape Juice and Winemaking Byproducts: Antioxidant Activities and Inhibition of Oxidation of Human Low-Density Lipoprotein Cholesterol and DNA Strand Breakage. J. Agric. Food Chem. 2014, 62, 12159–12171. [Google Scholar] [CrossRef] [PubMed]
- Pham-Huy, L.A.; He, H.; Pham-Huy, C. Free radicals, antioxidants in disease and health. Int. J. Biomed. Sci. 2008, 4, 89–96. [Google Scholar] [PubMed]
- Raha, S.; Robinson, B.H. Mitochondria, oxygen free radicals, disease and ageing. Trends Biochem. Sci. 2000, 25, 502–508. [Google Scholar] [CrossRef]
- Dudonné, S.; Poupard, P.; Coutière, P.; Woillez, M.; Richard, T.; Mérillon, J.-M.; Vitrac, X. Phenolic Composition and Antioxidant Properties of Poplar Bud (Populus nigra) Extract: Individual Antioxidant Contribution of Phenolics and Transcriptional Effect on Skin Aging. J. Agric. Food Chem. 2011, 59, 4527–4536. [Google Scholar] [CrossRef]
- De Camargo, A.C.; Vidal, C.M.M.; Canniatti-Brazaca, S.G.; Shahidi, F. Fortification of Cookies with Peanut Skins: Effects on the Composition, Polyphenols, Antioxidant Properties, and Sensory Quality. J. Agric. Food Chem. 2014, 62, 11228–11235. [Google Scholar] [CrossRef]
- Cherubim, D.J.; Martins, C.V.; Fariña, L.; Lucca, R.A. Polyphenols as natural antioxidants in cosmetics applications. J. Cosmet. Dermatol. 2020, 19, 33–37. [Google Scholar] [CrossRef]
- Wei, X.; Liu, Y.; Xiao, J.; Wang, Y. Protective Effects of Tea Polysaccharides and Polyphenols on Skin. J. Agric. Food Chem. 2009, 57, 7757–7762. [Google Scholar] [CrossRef]
- Russo, G.L.; Spagnuolo, C.; Russo, M.; Tedesco, I.; Moccia, S.; Cervellera, C. Mechanisms of aging and potential role of selected polyphenols in extending healthspan. Biochem. Pharmacol. 2020, 173, 113719. [Google Scholar] [CrossRef]
- Mohammad, I.S.; Naveed, M.; Ijaz, S.; Shumzaid, M.; Hassan, S.; Muhammad, K.S.; Rasool, F.; Akhtar, N.; Ishaq, H.M.; Khan, H.M.S. Phytocosmeceutical formulation development, characterization and its in-vivo investigations. Biomed. Pharmacother. 2018, 107, 806–817. [Google Scholar] [CrossRef]
- Kashif, M.; Akhtar, N. Dermocosmetic emulgels for anti-aging effects: Evidence from chromatographic and non-invasive biophysical techniques. Pak. J. Pharm. Sci. 2019, 32, 845–852. [Google Scholar]
- Chen, W.; Müller, D.; Richling, E.; Wink, M. Anthocyanin-rich Purple Wheat Prolongs the Life Span of Caenorhabditis elegans Probably by Activating the DAF-16/FOXO Transcription Factor. J. Agric. Food Chem. 2013, 61, 3047–3053. [Google Scholar] [CrossRef]
- Wang, H.; Cao, G.; Prior, R.L. Oxygen Radical Absorbing Capacity of Anthocyanins. J. Agric. Food Chem. 1997, 45, 304–309. [Google Scholar] [CrossRef]
- Peixoto, H.; Roxo, M.; Krstin, S.; Röhrig, T.; Richling, E.; Wink, M. An Anthocyanin-Rich Extract of Acai (Euterpe precatoria Mart.) Increases Stress Resistance and Retards Aging-Related Markers in Caenorhabditis elegans. J. Agric. Food Chem. 2016, 64, 1283–1290. [Google Scholar] [CrossRef]
- Wei, J.; Zhang, G.; Zhang, X.; Xu, D.; Gao, J.; Fan, J.; Zhou, Z. Anthocyanins from Black Chokeberry (Aroniamelanocarpa Elliot) Delayed Aging-Related Degenerative Changes of Brain. J. Agric. Food Chem. 2017, 65, 5973–5984. [Google Scholar] [CrossRef]
- Li, Y.-R.; Li, S.; Lin, C.-C. Effect of resveratrol and pterostilbene on aging and longevity. Biofactors 2018, 44, 69–82. [Google Scholar] [CrossRef]
- Boo, Y.C. Human Skin Lightening Efficacy of Resveratrol and Its Analogs: From in Vitro Studies to Cosmetic Applications. Antioxidants 2019, 8, 332. [Google Scholar] [CrossRef]
- Khan, M.; Park, S.; Kim, H.-J.; Lee, K.-J.; Kim, D.-H.; Baek, S.-H.; Hong, S.-T. The Resveratrol Rice DJ526 Callus Significantly Increases the Lifespan of Drosophila (Resveratrol Rice DJ526 Callus for Longevity). Nutrients 2019, 11, 983. [Google Scholar] [CrossRef] [PubMed]
- Aguilar-Alonso, P.; Vera-López, O.; Brambila-Colombres, E.; Segura-Badilla, O.; Avalos-López, R.; Lazcano-Hernández, M.; Navarro-Cruz, A.R. Evaluation of Oxidative Stress in Cardiomyocytes during the Aging Process in Rats Treated with Resveratrol. Oxid. Med. Cell. Longev. 2018, 2018, 1390483. [Google Scholar] [CrossRef] [PubMed]
- Buonocore, D.; Lazzeretti, A.; Tocabens, P.; Nobile, V.; Cestone, E.; Santin, G.; Bottone, M.G.; Marzatico, F. Resveratrol-procyanidin blend: Nutraceutical and antiaging efficacy evaluated in a placebo-controlled, double-blind study. Clin. Cosmet. Investig. Dermatol. 2012, 5, 159–165. [Google Scholar] [CrossRef] [PubMed]
- Diaz, M.; Degens, H.; Vanhees, L.; Austin, C.; Azzawi, M. The effects of resveratrol on aging vessels. Exp. Gerontol. 2016, 85, 41–47. [Google Scholar] [CrossRef]
- Sun, J.; Jing, S.; Jiang, R.; Wang, C.; Zhang, C.; Chen, J.; Li, H. Metabolomics study of the therapeutic mechanism of Schisandra chinensis lignans on aging rats induced by D-galactose. Clin. Interv. Aging 2018, 13, 829–841. [Google Scholar] [CrossRef]
- Lili, Z.; Jing, T.; Ying, C.; Ligang, J. Effects of Schisandra chinensis Extract on the Learning and Memory Ability of Mice with Learning and Memory Disorders. Nat. Prod. Commun. 2020, 15, 1–6. [Google Scholar] [CrossRef]





| Phenolic Acids | Flavonoids | Stilbenes | Lignans | |
|---|---|---|---|---|
Hydrobenzoic acids | Hydroxycinnamic acids |  |  |  |
gallic acid | caffeic acid | myricetin | resveratrol | podophyllotoxin |
syringic acid | ferulic acid | catechin | rhapontigenin | secoisolariciresinol |
tannic acid | coumaric acid | rutin | piceatannol | enterodiol |
sinapinic acid | naringin | rhaponticin | sesamin | |
rosmarinic acid | quercetin | oxyresveratrol | ||
| Study Design | Outcomes | Ref. |
|---|---|---|
| A stock solution of resveratrol at a concentration of 200 mg/mL in human umbilical vein endothelial cell (HUVEC); cell culture study | When treated with resveratrol following administration to HUVECs ox LDL, it has been shown that resveratrol inhibits increased ROS formation and lipid peroxidation caused by ox-LDL and reduces SOD activity. Resveratrol effectively inhibited endothelial cell apoptosis through inhibition of mitochondrial pathway and inhibition of oxidative damage. It has been reported that resveratrol may be effective in the treatment of atherosclerosis | [148] |
| Rosmarinic acid with antioxidant properties with different test methods | RA was determined 67.5 ± 1.68 (μg/mL). Total phenol content in terms of gallic acid equivalency, 1.62 ± 0.41 TEAC value (mM trolox equivalent/g), 806.7 ± 39.5 quercetin eq for total flavonoid test. It was specified 15 μg dry weight RA reduced 366.2 ± 9.9 μmol Fe (III) ions | [32,34] |
| Administration of resveratrol 50 mg/mL to healthy rats in drinking water for 3 weeks | It shows that resveratrol can improve the capacity of endothelial function and oxidative stress under physiological conditions | [149] |
| Prunan from prunes alleviates inflammation and oxidative stress during lithium/pilocarpine-induced epileptic seizures | Comparing 50 mg/kg b.w. pinoresinol-4-O-d-glucopyranoside to pilocarpine-treated animals, this compound reduced malondialdehyde levels by 24.2% and increased catalase activity by 44.4% | [150] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sahiner, M.; Yilmaz, A.S.; Gungor, B.; Ayoubi, Y.; Sahiner, N. Therapeutic and Nutraceutical Effects of Polyphenolics from Natural Sources. Molecules 2022, 27, 6225. https://doi.org/10.3390/molecules27196225
Sahiner M, Yilmaz AS, Gungor B, Ayoubi Y, Sahiner N. Therapeutic and Nutraceutical Effects of Polyphenolics from Natural Sources. Molecules. 2022; 27(19):6225. https://doi.org/10.3390/molecules27196225
Chicago/Turabian StyleSahiner, Mehtap, A. Sanem Yilmaz, Buket Gungor, Yasmin Ayoubi, and Nurettin Sahiner. 2022. "Therapeutic and Nutraceutical Effects of Polyphenolics from Natural Sources" Molecules 27, no. 19: 6225. https://doi.org/10.3390/molecules27196225
APA StyleSahiner, M., Yilmaz, A. S., Gungor, B., Ayoubi, Y., & Sahiner, N. (2022). Therapeutic and Nutraceutical Effects of Polyphenolics from Natural Sources. Molecules, 27(19), 6225. https://doi.org/10.3390/molecules27196225