
Chemical Composition and Phytotoxic, Antibacterial and Antibiofilm Activity of the Essential Oils of Eucalyptus occidentalis, E. striaticalyx and E. stricklandii

 ,
,  ,
,  , and
, and 
Abstract
:1. Introduction
2. Results and Discussion
2.1. Essential Oil Yields
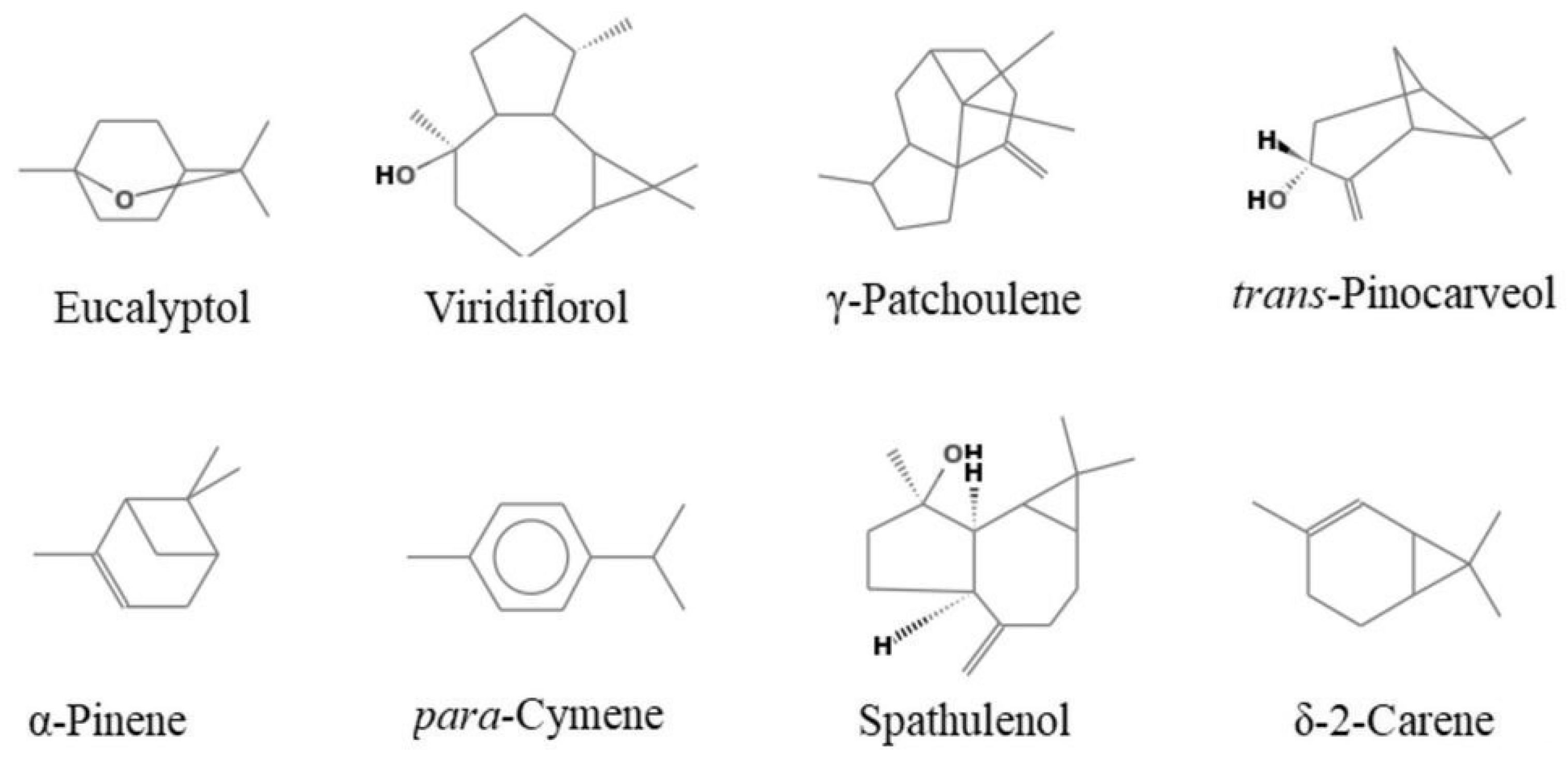
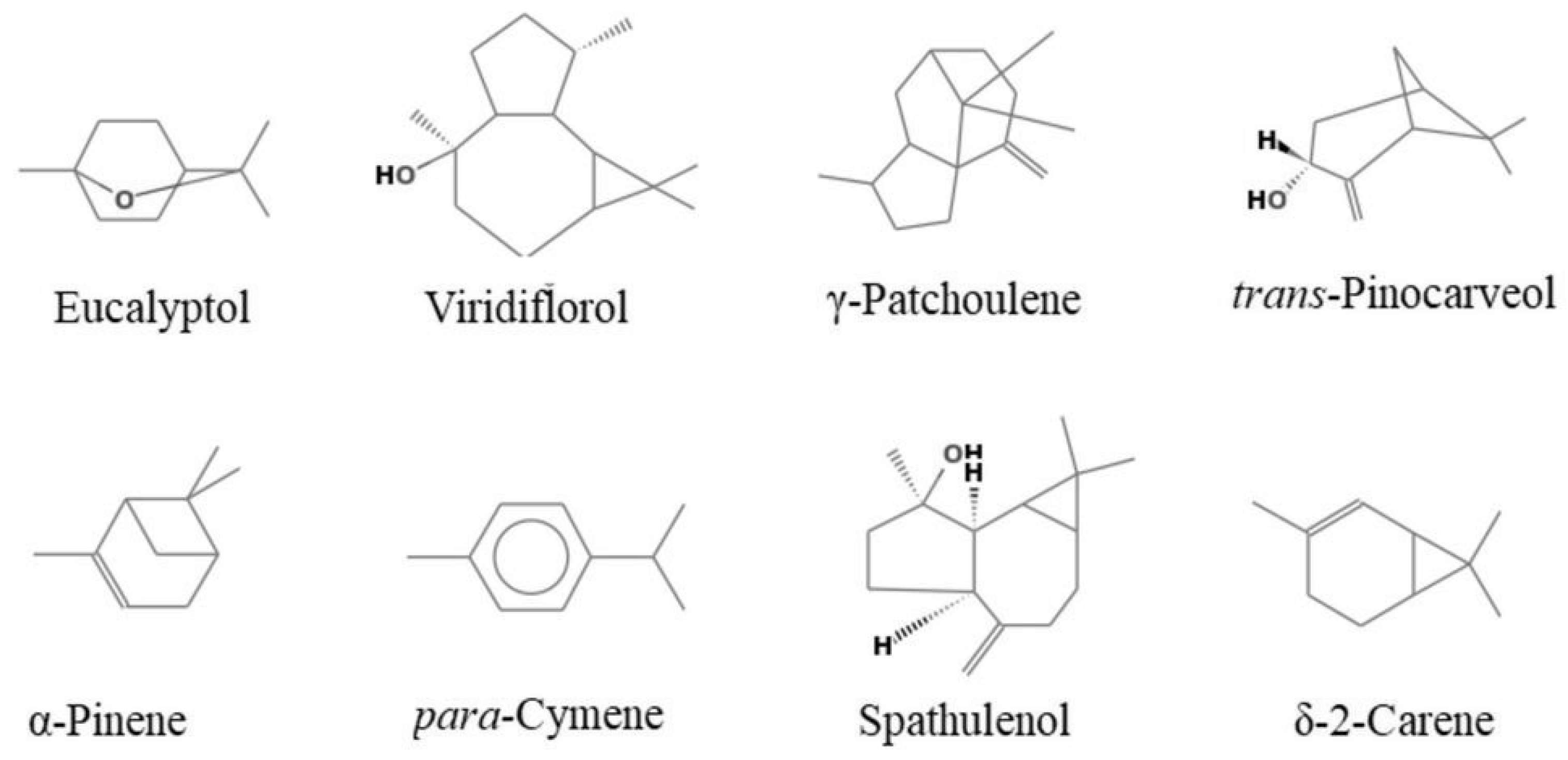
2.2. Chemical Composition
2.3. Phytotoxic Activity
2.4. Antimicrobial Activity
3. Materials and Methods
3.1. Plant Material
3.2. Isolation and Analysis of the Essential Oils
3.3. Phytotoxic Activity
3.4. Antimicrobial Activity
3.4.1. Microorganisms and Culture Conditions
3.4.2. Minimal Inhibitory Concentration (MIC)
3.5. Biofilm Inhibitory Activity
3.5.1. Activity on Mature Bacterial Biofilm
3.5.2. Effects of EOs on Cell Metabolic Activity within the Biofilm
3.6. Statistical Analysis
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Sample Availability
References
- Amri, I.; Khammassi, M.; Gargouri, S.; Hanana, M.; Jamoussi, B.; Hamrouni, L.; Mabrouk, Y. Tunisian pine essential oils: Chemical composition, herbicidal and antifungal properties. J. Essent. Oil-Bear. Plants 2022, 25, 430–443. [Google Scholar] [CrossRef]
- Cheng, S.S.; Huang, C.G.; Chen, Y.J.; Yu, J.J.; Chen, W.J.; Chang, S.T. Chemical compositions and larvicidal activities of leaf essential oils from two Eucalyptus species. Bioresour. Technol. 2009, 100, 452–456. [Google Scholar] [CrossRef] [PubMed]
- Zhou, L.; Li, J.; Kong, Q.; Luo, S.; Wang, J.; Feng, S.; Yuan, M.; Chen, T.; Yuan, S.; Ding, C. Chemical composition, antioxidant, antimicrobial, and phytotoxic potential of Eucalyptus grandis × E. urophylla leaves essential oils. Molecules 2021, 26, 1450. [Google Scholar] [CrossRef] [PubMed]
- Limam, H.; Ben Jemaa, M.; Tammar, S.; Ksibi, N.; Khammassi, N.; Jallouli, S.; Del Re, G.; Msaada, K. Variation in chemical profile of leaves essential oils from thirteen Tunisian Eucalyptus species and evaluation of their antioxidant and antibacterial properties. Ind. Crops Prod. 2020, 158, 112964. [Google Scholar] [CrossRef]
- Amakura, Y.; Yoshimura, M.; Sugimoto, N.; Yamazaki, T.; Yoshida, T. Marker constituents of the natural antioxidant Eucalyptus leaf extract for the evaluation of food additives. Biosci. Biotechnol. Biochem. 2009, 73, 1060–1065. [Google Scholar] [CrossRef] [PubMed]
- Asgary, S.; Sahebkar, A.; Afshani, M.R.; Keshvari, M.; Haghjooyjavanmard, S.; Rafieian Kopaei, M. Clinical evaluation of blood pressure lowering, endothelial function improving, hypolipidemic and anti-inflammatory effects of pomegranate juice in hypertensive subjects. Phytother. Res. 2014, 28, 193–199. [Google Scholar] [CrossRef]
- Msaada, K.; Salem, N.; Bachrouch, O.; Bousselmi, S.; Tammar, S.; Alfaify, A.; Al Sane, K.; Ben Ammar, W.; Azeiz, S.; Haj Brahim, A.; et al. Chemical composition and antioxidant and antimicrobial activities of wormwood (Artemisia absinthium L.) essential oils and phenolics. J. Chem. 2015, 2015, 804658. [Google Scholar] [CrossRef]
- Harkat-Madouria, L.; Asma, B.; Madania, K.; Bey-Ould Si Saida, Z.; Rigouc, P.; Grenierd, D.; Allaloua, H.; Reminia, H.; Adjaouda, A.; Boulekbache-Makhlouf, L. Chemical composition, antibacterial and antioxidant activities of essential oil of Eucalyptus globulus from Algeria. Ind. Crops Prod. 2015, 78, 148–153. [Google Scholar] [CrossRef]
- Batish, D.R.; Singh, H.P.; Kohli, R.K.; Kaur, S. Eucalyptus essential oil as a natural pesticide. For. Ecol. Manag. 2009, 256, 2166–2174. [Google Scholar] [CrossRef]
- Salehi, B.; Sharifi-Rad, J.; Quispe, C.; Llaique, H.; Villalobos, M.; Smeriglio, A.; Trombetta, D.; Ezzat, S.M.; Salem, M.A.; Zayed, A.; et al. Insights into Eucalyptus genus chemical constituents, biological activities and health-promoting effects. Trends Food Sci. Technol. 2019, 91, 609–624. [Google Scholar] [CrossRef]
- Ben Hassine, D.; Kammoun El Euch, S.; Souchard, J.P.; Cazaux, S.; Abderrabba, M.; Bouajila, J. Phytochemical study and pharmaceutical properties of essential oils and organic extracts of two Eucalyptus species: E. stricklandii Maiden and E. brevifolia F. Muell. J. Essent. Oil Res. 2022, 34, 339–351. [Google Scholar] [CrossRef]
- Bande-Borujeni, S.; Zandi-Sohani, N.; Ramezani, L. Chemical composition and bioactivity of essential oil from Eucalyptus occidentalis leaves against two stored product pests. Int. J. Trop. Insect Sci. 2008, 38, 216–223. [Google Scholar] [CrossRef]
- Ben Ghnaya, A.; Hamrouni, L.; Amri, I.; Ahoues, H.; Hanana, M.; Romane, A. Study of allelopathic effects of Eucalyptus erythrocorys L. crude extracts against germination and seedling growth of weeds and wheat. Nat. Prod. Res. 2016, 30, 2058–2064. [Google Scholar] [CrossRef] [PubMed]
- Ramezani, S.; Saharkhiz, M.J.; Ramezani, F.; Fotokian, M.H. Use of essential oils as bioherbicides. J. Essent. Oil-Bear. Plants 2008, 11, 319–327. [Google Scholar] [CrossRef]
- Elaissi, A.; Medini, H.; Marzouki, H.; Larbi Khouja, M.; Lynene, F.; Chemli, R.; Harzallah-Skhiri, F. Variation in volatile leaf oils of twelve eucalyptus species harvested from Hajeb Layoun arboreta (Tunisia). Chem. Biodivers. 2010, 27, 705–716. [Google Scholar] [CrossRef]
- Bignell, C.M.; Dunlop, P.J.; Brophy, J.J. Volatile leaf oils of some South-western and Southern Australian species of the genus Eucalyptus (series I). Part XVII: Subgenus Symphyomyrtus (i) Section Bisectaria, Series Calycogonae and (ii) Section Dumaria, Series Dumosae, Series Rigentes and Series Ovulares. Flavour Fragr. J. 1997, 12, 269–275. [Google Scholar] [CrossRef]
- Khammassi, M.; Mighri, H.; Ben Mansour, M.; Amri, I.; Jamoussi, B.; Khaldi, A. Metabolite profiling and potential antioxidant activity of sixteen fennel (Foeniculum vulgare Mill.) populations wild-growing in Tunisia. S. Afr. J. Bot. 2022, 148, 407–414. [Google Scholar] [CrossRef]
- Saoud, I.; Hamrouni, L.; Gargouri, S.; Amri, I.; Hanana, M.; Fezzani, T.; Bouzid, S.; Jamoussi, B. Chemical composition, weed killer and antifungal activities of Tunisian thyme (Thymus capitatus Hoff. et Link.) essential oils. Acta Aliment. 2013, 42, 417–427. [Google Scholar] [CrossRef]
- Elaissi, A.; Chraif, I.; Bannour, F.; Farhat, F.; Ben Salah, M.; Chemli, R.; Khouja, M.L. Contribution to the Qualitative and Quantitative Study of Seven Eucalyptus Species Essential Oil Harvested of Hajeb’s Layoun Arboreta (Tunisia). J. Essent. Oil-Bear. Plants 2007, 10, 15–25. [Google Scholar] [CrossRef]
- Bagheri, H.; Manap, M.Y.B.A.; Solati, Z. Antioxidant activity of Piper nigrum L. essential oil extracted by supercritical CO2 extraction and hydro-distillation. Talanta 2014, 121, 220–228. [Google Scholar] [CrossRef]
- Bignell, C.M.; Dunlop, P.J.; Brophy, J.J.; Jackson, J.F. Volatile leaf oils of some South-western and Southern Australian species of the genus Eucalyptus. Part XI. Subgenus Symphyomyrtus. A—Section Bisectaria. (a) Series Occidentales, (b) unpublished Series Annulatae, (c) Series Micromembranae, (d) Series Obliquae, (e) Series Dundasianae, (f) Series Cooperianae, (g) Series Halophilae, (h) Series Salmonophloiae, and (i) Series Pubescentes. B—Section Dumaria. (a) Series Merrickianae. Flavour Fragr. J. 1996, 11, 107–112. [Google Scholar] [CrossRef]
- Diäaz-Maroto, M.C.; Perez-Coello, M.S.; Esteban, J.; Sanz, J.S. Comparison of the volatile composition of wild fennel samples (Foeniculum vulgare Mill.) from central Spain. J. Agric. Food Chem. 2006, 54, 6814–6818. [Google Scholar] [CrossRef] [PubMed]
- Telci, I.; Toncer, O.G.; Sahbaz, N. Yield, essential oil content and composition of Coriandrum sativum varieties (var. vulgare Alef and var. microcarpum DC.) grown in two different locations. J. Essent. Oil Res. 2006, 18, 189–193. [Google Scholar] [CrossRef]
- Amri, I.; Mancini, E.; De Martino, L.; Marandino, A.; Lamia, H.; Mohsen, H.; Bassem, J.; Scognamiglio, M.R.; Reverchon, E.; De Feo, V. Chemical composition and biological activities of the essential oils from three Melaleuca species grown in Tunisia. Int. J. Mol. Sci. 2012, 13, 16580–16591. [Google Scholar] [CrossRef]
- Li, A.; Wu, H.; Feng, Y.; Deng, S.; Hou, A.; Che, F.; Liu, Y.; Geng, Q.; Ni, H.; Wei, Y. A strategy of rapidly screening out herbicidal chemicals from Eucalyptus essential oils. Pest. Manag. Sci. 2020, 76, 917–927. [Google Scholar] [CrossRef] [PubMed]
- De Martino, L.; Mancini, E.; Rolim de Almeida, L.F.; De Feo, V. The antigerminative activity of twenty-seven monoterpenes. Molecules 2010, 15, 6630–6637. [Google Scholar] [CrossRef]
- Qaderi, M.M.; Cavers, P.B.; Bernards, M.A. Isolation and structural characterization of a water-soluble germination inhibitor from scotch thistle (Onopordum acanthium) cypselas. J. Chem. Ecol. 2003, 29, 2425–2438. [Google Scholar] [CrossRef]
- Singh, H.P.; Batish, D.R.; Kohli, R.K. Allelopathic effect of two volatile monoterpenes against bill goat weed (Ageratum conyzoides L.). Crop. Prot. 2002, 21, 347–350. [Google Scholar] [CrossRef]
- Romagni, J.G.; Allen, S.N.; Dayan, F.E. Allelopathic effects of volatile cineoles on two weedy plant species. J. Chem. Ecol. 2000, 26, 303–313. [Google Scholar] [CrossRef]
- Kordali, S.; Cakir, A.; Ozer, H.; Cakmakci, R.; Kesdek, M.; Mete, E. Antifungal, phytotoxic and insecticidal properties of essential oil isolated from Turkish Origanum acutidens and its three components, carvacrol, thymol and p-cymene. Bioresour. Technol. 2008, 99, 8788–8795. [Google Scholar] [CrossRef]
- Araniti, F.; Sanchéz Moreiras, A.M.; Graña, E.; Reigosa, M.G.; Abenavoli, M. R: Terpenoid trans-caryophyllene inhibits weed germination and induces plant water status alteration and oxidative damage in adult Arabidopsis. Plant Biol. 2017, 19, 79–89. [Google Scholar] [CrossRef] [PubMed]
- Abrahim, D.; Francischini, A.C.; Pergo, E.M.; Kelmer-Bracht, A.M.; Ishii-Iwamoto, E.L. Effects of α-pinene on the mitochondrial respiration of maize seedlings. Plant Physiol. Biochem. 2003, 41, 985–991. [Google Scholar] [CrossRef]
- Singh, H.P.; Batish, D.R.; Kaur, S.; Arora, K.; Kohli, R.K. a-Pinene inhibits growth and induces oxidative stress in roots. Ann. Bot. 2006, 98, 1261–1269. [Google Scholar] [CrossRef] [PubMed]
- Chowhan, N.; Singh, H.P.; Kohl, R.K. Phytotoxic effects of β-pinene on early growth and associated biochemical changes in rice. Acta Physiol. Plant. 2011, 33, 2369–2376. [Google Scholar] [CrossRef]
- Podesta, F.E.; Plaxton, W.C. Regulation of cytosolic carbon metabolism in germinating Ricinus communis cotyledons: I. Developmental profiles for the activity, concentration, and molecular structure of the pyrophosphate-and ATP-dependent phosphofructokinase, phosphoenolpyruvate carboxylase and pyruvate kinase. Planta 1994, 194, 374–380. [Google Scholar]
- Nazzaro, F.; Fratianni, F.; De Martino, L.; Coppola, R.; De Feo, V. Effect of essential oils on pathogenic bacteria. Pharmaceuticals 2013, 6, 1451–1474. [Google Scholar] [CrossRef]
- Nazzaro, F.; Fratianni, F.; Coppola, R. Quorum sensing and phytochemicals. Int. J. Mol. Sci. 2013, 14, 12607–12619. [Google Scholar] [CrossRef]
- LaSarre, B.; Federle, M.J. Exploiting quorum sensing to confuse bacterial pathogens. Microbiol. Mol. Biol. Rev. 2013, 77, 73–111. [Google Scholar] [CrossRef]
- Mączka, W.; Duda-Madej, A.; Górny, A.; Grabarczyk, M.; Wińska, K. Can eucalyptol replace antibiotics? Molecules 2021, 26, 4933. [Google Scholar] [CrossRef]
- Jennings, W.; Shibamoto, T. Qualitative Analysis of Flavour and Fragrance Volatiles by Glass Capillary Gas Chromatography; Academic Press: New York, NY, USA, 1980. [Google Scholar]
- Davies, N.W. Gas chromatographic retention indices of monoterpenes and sesquiterpenes on methyl silicon and Carbowax 20M phases. J. Chromatogr. A 1990, 503, 1–24. [Google Scholar] [CrossRef]
- Adams, R.P. Identification of Essential Oil Components by Gas Chromatography/Mass Spectrometry; Allured Publishing Co.: Carol Stream, IL, USA, 2007. [Google Scholar]
- McLafferty, F.W. Wiley Registry of Mass Spectral Data, with NIST Spectral Data CD Rom, 7th ed.; John Wiley & Sons: New York, NY, USA, 1980. [Google Scholar]
- Sarker, S.D.; Nahar, L.; Kumarasamy, Y. Microtitre plate-based antibacterial assay incorporating resazurin as an indicator of cell growth, and its application in the in vitro antibacterial screening of phytochemicals. Methods 2007, 42, 321–324. [Google Scholar] [CrossRef] [PubMed]
- Nazzaro, F.; Polito, F.; Amato, G.; Caputo, L.; Francolino, R.; d’Acierno, A.; Fratianni, F.; Candido, V.; Coppola, R.; De Feo, V. Chemical composition of essential oils and bulbs and aerial parts of two cultivars of Allium sativum and their biofilm activity agaianst food and nosocomial pathogens. Antibiotics 2022, 11, 724. [Google Scholar] [CrossRef] [PubMed]
- Fratianni, F.; d’Acierno, A.; Ombra, M.N.; Amato, G.; De Feo, V.; Ayala-Zaval, J.F.; Coppola, R.; Nazzaro, F. Fatty acid composition and in vitro anti-inflammatory activity of five cold-pressed Prunus seed oils, and their anti-biofilm effect against pathogenic bacteria. Front. Nutr. 2021, 8, 775751. [Google Scholar] [CrossRef] [PubMed]
- Fratianni, F.; Ombra, M.N.; d’Acierno, A.; Caputo, L.; Amato, G.; De Feo, V.; Coppola, R.; Nazzaro, F. Polyphenols content and in vitro α-glycosidase activity of different italian monofloral honeys, and their effect on selected pathogenic and probiotic bacteria. Microorganisms 2021, 9, 1694. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
{kind=link}
| Compound | Ki a | Ki b | A% | B% | C% | Identification | |
|---|---|---|---|---|---|---|---|
| 1 | α-thujene | 930 | - | - | 2.1 | - | 1,2,3 |
| 2 | α-pinene | 939 | 1012 | 0.2 | 7.4 | 4.3 | 1,2,3 |
| 3 | Camphene | 954 | 1075 | - | - | 0.1 | 1,2,3 |
| 4 | β-pinene | 979 | 1110 | - | 1.8 | - | 1,2,3 |
| 5 | Myrcene | 990 | 1145 | - | 1.5 | - | 1,2,3 |
| 6 | δ-2-carene | 1002 | - | - | 8.5 | - | 1,2 |
| 7 | α-phellandrene | 1002 | 1177 | 0.1 | - | - | 1,2,3 |
| 8 | α-terpinene | 1017 | 1170 | - | 1.3 | - | 1,2,3 |
| 9 | p-cymene | 1024 | - | 0.8 | 19.4 | 0.6 | 1,2,3 |
| 10 | Eucalyptol | 1031 | 1031 | 40.8 | 29.6 | 73.6 | 1,2,3 |
| 11 | γ-terpinene | 1059 | 1221 | - | 0.6 | 1.0 | 1,2,3 |
| 12 | Cis-sabinene hydrate | 1070 | - | - | 0.1 | - | 1,2 |
| 13 | Terpinolene | 1088 | - | - | 0.9 | - | 1,2,3 |
| 14 | Exo-fenchol | 1121 | - | - | - | 0.2 | 1,2 |
| 15 | Cis-p-menth-2-en-1-ol | 1121 | - | - | 0.3 | - | 1,2 |
| 16 | α-camphonenal | 1126 | - | - | - | 0.1 | 1,2 |
| 17 | Trans-pinocarveol | 1139 | - | 0.2 | 0.3 | 6.0 | 1,2 |
| 18 | Isopulegol | 1149 | 1533 | - | 0.3 | - | 1,2 |
| 19 | Eucarvone | 1150 | - | - | - | 1.3 | 1,2 |
| 20 | Sabina ketone | 1159 | 1651 | - | 0.4 | - | 1,2 |
| 21 | Isoborneol | 1160 | 1642 | - | - | 0.4 | 1,2 |
| 22 | Pinocarvone | 1164 | 1586 | - | 0.1 | - | 1,2,3 |
| 23 | Neo-iso-pulegol | 1171 | - | - | - | 0.2 | 1,2 |
| 24 | Terpinen-4-ol | 1177 | 1590 | - | 3.4 | 0.7 | 1,2,3 |
| 25 | Cryptone | 1185 | 1659 | - | 0.4 | 0.2 | 1,2 |
| 26 | α-terpineol | 1188 | 1661 | - | 0.4 | 1.2 | 1,2,3 |
| 27 | Neo-dihydrocarveol | 1194 | - | - | - | 0.4 | 1,2 |
| 28 | Trans-pulegol | 1214 | - | - | 0.3 | - | 1,2 |
| 29 | Trans-carveol | 1216 | 1878 | - | 0.2 | 0.5 | 1,2 |
| 30 | Cis-pulegol | 1229 | - | - | 0.4 | - | 1,2 |
| 31 | Cumin aldehyde | 1241 | - | - | 0.1 | - | 1,2 |
| 32 | δ-elemene | 1338 | 1479 | 0.1 | 1.5 | - | 1,2 |
| 33 | β-longipinene | 1400 | - | 1.0 | - | - | 1,2 |
| 34 | Longifolene | 1407 | 1574 | 4.8 | - | - | 1,2 |
| 35 | Aromadendrene | 1441 | 1631 | 0.7 | 0.2 | 0.1 | 1,2 |
| 36 | α-himachalene | 1451 | - | 0.1 | 0.4 | - | 1,2 |
| 37 | Allo-aromadendrene | 1460 | 1660 | 0.1 | - | 0.1 | 1,2 |
| 38 | 9-epi-(E)-Caryophyllene | 1466 | - | 0.1 | 0.1 | - | 1,2 |
| 39 | γ-gurjunene | 1477 | - | 0.6 | 0.6 | - | 1,2 |
| 40 | Epi-cubebol | 1494 | - | 0.1 | 0.1 | - | 1,2 |
| 41 | Viridiflorene | 1496 | - | - | 0.4 | 0.1 | 1,2 |
| 42 | γ-patchoulene | 1502 | - | 5.5 | - | 0.3 | 1,2 |
| 43 | Trans-β-guaiene | 1502 | - | 2.0 | - | - | 1,2 |
| 44 | Spathulenol | 1578 | - | 3.0 | 8.7 | 0.4 | 1,2 |
| 45 | Globulol | 1590 | 2104 | - | 2.1 | 1.8 | 1,2 |
| 46 | Viridiflorol | 1592 | 2110 | 29.9 | - | - | 1,2 |
| 47 | Cubeban-11-ol | 1595 | - | 1.0 | 0.2 | - | 1,2 |
| 48 | Rosifoliol | 1600 | - | - | 0.4 | 0.1 | 1,2 |
| 49 | Guaiol | 1600 | 2094 | - | - | 0.1 | 1,2 |
| 50 | α-epi-7-epi-5-eudesmol | 1607 | - | 2.0 | - | - | 1,2 |
| 51 | Epi-cedrol | 1619 | - | 2.1 | - | - | 1,2 |
| 52 | γ-eudesmol | 1632 | 2178 | - | - | 0.1 | 1,2 |
| 53 | Allo-aromadendrene epoxide | 1641 | - | - | 0.2 | - | 1,2 |
| 54 | Cubenol | 1646 | 2080 | 1.1 | - | - | 1,2 |
| 56 | β-eudesmol | 1650 | - | - | 0.2 | 3.2 | 1,2 |
| 57 | Vulgarone B | 1651 | - | - | 0.9 | - | 1,2 |
| 58 | Cedr-8(15)-en-9-α-ol | 1651 | - | - | 0.2 | - | 1,2 |
| 59 | α-eudesmol | 1653 | 2247 | 0.8 | 0.8 | - | 1,2 |
| 60 | α-cadinol | 1654 | 2224 | - | 0.3 | - | 1,2 |
| 61 | 5-hydroxy-isobornyl isobutanoate | 1658 | - | - | 0.2 | - | 1,2 |
| Total | 97.1 | 96.5 | 97.1 | ||||
| Monoterpene hydrocarbons | 1.1 | 43.5 | 6 | ||||
| Oxygenated monoterpenes | 41.0 | 36.3 | 84.8 | ||||
| Sesquiterpene hydrocarbons | 15.0 | 3.2 | 0.6 | ||||
| Oxygenated sesquiterpenes | 40.0 | 13.5 | 5.7 | ||||
| Weeds | Doses (µL/mL) | Essential Oils | ||
|---|---|---|---|---|
| E. occidentalis | E. stricklandii | E. striaticalyx | ||
| L. rigidum | Control (water) | 86.66 ± 2.88 a | 86.66 ± 2.88 a | 86.66 ± 2.88 a |
| 0 | 81.7 ± 1.7 a | 81.7 ± 1.7 a | 81.7 ± 1.7 a | |
| 0.75 | 78.3 ± 1.7 a | 61.7 ± 3.3 b | 70.0 ± 2.9 b | |
| 1.50 | 55.0 ± 2.9 b | 45.0 ± 5 c | 46.7 ± 4.4 c | |
| 2.25 | 33.3 ± 4.4 c | 31.7 ± 6 d | 26.6 ± 3.3 d | |
| 3.00 | 33.3 ± 1.6 c | 6.7 ± 1.7 e | 10.0 ± 2.9 e | |
| S. arvensis | Control (water) | 96.66 ± 2.88 a | 96.66 ± 2.88 a | 96.66 ± 2.88 a |
| 0 | 96.7 ± 1.7 a | 96.7 ± 1.7 a | 96.7 ± 1.7 a | |
| 0.75 | 70.0 ± 2.9 b | 63.3 ± 3.3 b | 66.7 ± 4.4 b | |
| 1.50 | 46.7 ± 4.4 c | 31.7 ± 1.7 c | 26.7 ± 4.4 c | |
| 2.25 | 23.3 ± 1.7 d | 16.7 ± 4.4 d | 6.7 ± 1.7 d | |
| 3.00 | 1.7 ± 1.7 e | 0 ± 0 e | 0 ± 0 d | |
| T. campestre | Control (water) | 93.66 ± 2.9 a | 93.66 ± 2.9 a | 93.66 ± 2.9 a |
| 0 | 85 ± 2.9 a | 85.0 ± 2.9 a | 85.0 ± 2.9 a | |
| 0.75 | 75 ± 2.9 a | 63.3 ± 4.4 b | 46.7 ± 4.4 b | |
| 1.50 | 38.3 ± 4.4 b | 38.3 ± 1.7 c | 23.3 ± 1.7 c | |
| 2.25 | 35.0 ± 2.9 b | 21.7 ± 1.7 d | 8.3 ± 3.3 d | |
| 3.00 | 20.0 ± 2.9 c | 0 ± 0 e | 0 ± 0 d | |
| Weeds | Doses (µL/mL) | Essential Oils | ||
|---|---|---|---|---|
| E. occidentalis | E. stricklandii | E. striaticalyx | ||
| L. rigidum | Control (water) | 6.7 ± 0.5 a | 6.7 ± 0.5 a | 6.7 ± 0.5 a |
| 0 | 6.4 ± 0.6 a | 6.4 ± 0.6 a | 6.4 ± 0.6 a | |
| 0.75 | 3.6 ± 0.3 b | 4.8 ± 0.21 b | 5.7 ± 0.3 ab | |
| 1.50 | 2.7 ± 0.3 bc | 4.4 ± 0.23 b | 4.7 ± 0.3 b | |
| 2.25 | 1.4 ± 0.2 c | 2.7 ± 0.2 c | 3.4 ± 0.3 c | |
| 3.00 | 1.4 ± 0.1 c | 1.8 ± 0.3 c | 1.5 ± 0.2 d | |
| S. arvensis | Control (water) | 8 ± 0.3 a | 8 ± 0.3 a | 8 ± 0.3 a |
| 0 | 8 ± 0.46 a | 8.0 ± 0.46 a | 8 ± 0.46 a | |
| 0.75 | 5.9 ± 0.26 b | 5.0 ± 0.3 b | 6.7 ± 0.4 b | |
| 1.50 | 3.6 ± 0.23 c | 3.2 ± 0.1 c | 4.2 ± 0.2 c | |
| 2.25 | 2.1 ± 0.3 d | 2.3 ± 0.2 d | 1.9 ± 0.3 d | |
| 3.00 | 0.1 ± 0.1 e | 0 ± 0 e | 0 ± 0 e | |
| T. campestre | Control (water) | 7 ± 0.4 a | 7 ± 0.4 a | 7 ± 0.4 a |
| 0 | 6.9 ± 0.48 a | 6.9 ± 0.48 a | 6.9 ± 0.48 a | |
| 0.75 | 5.7 ± 0.2 b | 5.8 ± 0.4 b | 6.6 ± 0.3 a | |
| 1.50 | 4.3 ± 0.3 c | 3.9 ± 0.2 c | 4.3 ± 0.3 b | |
| 2.25 | 2.7 ± 0.2 d | 2.7 ± 0.4 d | 2.1 ± 0.2 c | |
| 3.00 | 1.3 ± 0.1 e | 0 ± 0 e | 0 ± 0 d | |
| Weeds | Doses (µL/mL) | Essential Oils | ||
|---|---|---|---|---|
| E. occidentalis | E. stricklandii | E. striaticalyx | ||
| L. rigidum | Control (water) | 5.5 ± 0.3 a | 5.5 ± 0.3 a | 5.5 ± 0.3 a |
| 0 | 5.7 ± 0.3 a | 5.7 ± 0.3 a | 5.7 ± 0.3 a | |
| 0.75 | 5.3 ± 0.3 a | 4.9 ± 0.2 a | 3.7 ± 0.2 b | |
| 1.50 | 3.2 ± 0.2 b | 3.5 ± 0.3 b | 2.2 ± 0.1 c | |
| 2.25 | 1.3 ± 0.1 c | 2.3 ± 0.3 c | 2.2 ± 0.1 c | |
| 3.00 | 0.5 ± 0.2 d | 1.4 ± 0.1 d | 0.3 ± 0.1 d | |
| S. arvensis | Control (water) | 6.9 ± 0.4 a | 6.9 ± 0.4 a | 6.9 ± 0.4 a |
| 0 | 6.8 ± 0.32 a | 6.8 ± 0.32 a | 6.8 ± 0.32 a | |
| 0.75 | 6.5 ± 0.3 a | 4.1 ± 0.1 b | 5.0 ± 0.2 b | |
| 1.50 | 4.7 ± 0.4 b | 2.3 ± 0.2 c | 2.4 ± 0.2 c | |
| 2.25 | 2.9 ± 0.2 c | 1.5 ± 0.2 d | 2 ± 0.1 c | |
| 3.00 | 0.1 ± 0.1 d | 0 ± 0 e | 0 ± 0 d | |
| T. campestre | Control (water) | 6 ± 0.2 a | 6 ± 0.2 a | 6 ± 0.2 a |
| 0 | 5.6 ± 0.3 a | 5.6 ± 0.3 a | 5.6 ± 0.3 a | |
| 0.75 | 5.1 ± 0.2 a | 4.9 ± 0.1 b | 3.7 ± 0.5 b | |
| 1.50 | 3.2 ± 0.2 b | 2.7 ± 0.3 c | 1.8 ± 0.2 c | |
| 2.25 | 2.3 ± 0.4 c | 2.5 ± 0.2 c | 1.3 ± 0.1 c | |
| 3.00 | 1.4 ± 0.1 d | 0 ± 0 d | 0 ± 0 d | |
| EO | A. baumannii | E. coli | L. monocytogenes | P. aeruginosa | S. aureus |
|---|---|---|---|---|---|
| E. occidentalis | 25 ± 2 * | 30 ± 3 * | 35 ± 2 | 45 ± 2 **** | 40 ± 2 |
| E. striaticalyx | 30 ± 3 | 40 ± 3 **** | 40 ± 2 * | 40 ± 3 * | 35 ± 3 |
| E. stricklandii | 35 ± 3 | 40 ± 3 **** | 35 ± 2 | 35 ± 3 | 35 ± 3 |
| Tetracycline | 31 ± 1 | 24 ± 3 | 33 ± 2 | 34 ± 2 | 38 ± 1 |
| Time 0 | A. baumannii | E. coli | L. monocytogenes | P. aeruginosa | S. aureus |
|---|---|---|---|---|---|
| E. occidentalis 10 μL/mL | 83.92 ± 2,17 **** | 20.54 ± 0.07 **** | 0 ± 0 | 0 ± 0 | 2.02 ± 0.74 |
| E. occidentalis 20 μL/mL | 86.73 ± 1.16 **** | 46.93 ± 0.87 **** | 79.48 ± 0.36 **** | 13.79 ± 1.00 **** | 31.08 ± 0.73 **** |
| E. striaticalyx 10 μL/mL | 35.57 ± 6.98 **** | 0 ± 0 | 8.23 ± 0.69 **** | 0.84 ± 0.20 | 48.25 ± 2.12 **** |
| E. striaticalyx 20 μL/mL | 66.66 ± 0.94 **** | 10.36 ± 4.61 **** | 27.16 ± 4.11 **** | 30.67 ± 2.49 **** | 60.46 ± 0.66 **** |
| E. stricklandii 10 μL/mL | 5.24 ± 0.65 ** | 1.12 ± 0.40 | 0 ± 0 | 0 ± 0 | 45.20 ± 1.92 **** |
| E. stricklandii 20 μL/mL | 46.49 ± 3.62 **** | 1.48 ± 0.27 | 36.86 0.78 **** | 42.66 ± 1.33 **** | 45.16 ± 2.01 **** |
| 24 h | A. baumannii | E. coli | L. monocytogenes | P. aeruginosa | S. aureus |
| E. occidentalis 10 μL/mL | 9.71 ± 0.67 **** | 28.60 ± 1.04 **** | 0 ± 0 | 47.97 ± 0.58 **** | 49.93 ± 0.82 **** |
| E. occidentalis 20 μL/mL | 32.45 ± 0.62 **** | 40.35 ± 0.87 **** | 67.75 ± 0.46 **** | 56.20 ± 0.57 **** | 52.42 ± 0.85 **** |
| E. striaticalyx 10 μL/mL | 39.11 ± 0.38 **** | 6.35 ± 0.73 **** | 26.38 ± 1.35 **** | 8.18 ± 0.57 **** | 19.40 ± 0.79 **** |
| E. striaticalyx 20 μL/mL | 45.23 ± 0.91 **** | 31.21 ± 1.21 **** | 34.14 ± 0.62 **** | 17.43 ± 0.58 **** | 23.29 ± 1.15 **** |
| E. stricklandii 10 μL/mL | 25.72 ± 0.83 **** | 0 ± 0 | 0 ± 0 | 9.23 ± 1.37 **** | 29.10 ± 0.89 **** |
| E. stricklandii 20 μL/mL | 58.83 ± 0.38 **** | 29.22 ± 0.81 **** | 42.28 ± 0.62 **** | 51.60 ± 0.37 **** | 36.04 ± 0.89 **** |
| Time 0 | A. baumannii | E. coli | L. monocytogenes | P. aeruginosa | S. aureus |
|---|---|---|---|---|---|
| E. occidentalis 10 μL/mL | 41.05 ± 3.27 **** | 0 ± 0 | 0 ± 0 | 0 ± 0 | 57.44 ± 4.98 **** |
| E. occidentalis 20 μL/mL | 55.18 ± 1.30 **** | 7.70 ± 0.80 **** | 44.69 ± 0.40 **** | 14.33 ± 0.20 **** | 72.03 ± 2.30 **** |
| E. striaticalyx 10 μL/mL | 0 ± 0 | 12.50 ± 0.57 **** | 96.01 ± 1.11 **** | 44.35 ± 0.85 **** | 67.18 ± 4.25 **** |
| E. striaticalyx 20 μL/mL | 27.19 ± 1.19 **** | 27.11 ± 1.32 **** | 96.81 ± 1.31 **** | 39.38 ± 0.85 **** | 76.36 ± 0.99 **** |
| E. stricklandii 10 μL/mL | 0.00 ± 0.00 | 0.00 ± 0.00 | 20.90 ± 0.67 **** | 0 ± 0 | 79.44 ± 0.78 **** |
| E. stricklandii 20 μL/mL | 42.06 ± 1.30 **** | 21.28 ± 1.87 **** | 97.67 ± 1.81 **** | 60.65 ± 1.87 **** | 83.02 ± 1.04 **** |
| 24 h | A. baumannii | E. coli | L. monocytogenes | P. aeruginosa | S. aureus |
| E. occidentalis 10 μL/mL | 0 ± 0 | 10.32 ± 0.57 **** | 0 ± 0 | 0 ± 0 | 18.43 ± 0.24 **** |
| E. occidentalis 20 μL/mL | 88.39 ± 1.50 **** | 27.08 ± 0.68 **** | 0 ± 0 | 9.73 ± 0.47 **** | 30.54 ± 0.63 **** |
| E. striaticalyx 10 μL/mL | 62.22 ± 1.26 **** | 23.15 ± 0.76 **** | 26.28 0.15 **** | 44.40 ± 0.69 **** | 0 ± 0 |
| E. striaticalyx 20 μL/mL | 67.91 ± 0.72 **** | 24.05 ± 0.92 **** | 26.55 ± 2.44 **** | 45.11 ± 0.84 **** | 0.68 ± 0.02 |
| E. stricklandii 10 μL/mL | 6.26 ± 0.22 **** | 0 ± 0 | 27.15 ± 0.80 **** | 42.53 ± 0.35 **** | 0 ± 0 |
| E. stricklandii 20 μL/mL | 67.14 ± 0.15 **** | 13.29 ± 0.34 **** | 29.77 ± 0.53 **** | 45.31 ± 0.88 **** | 0 ± 0 |
| Species/Part Collected | Date of Harvest | Harvest Site | Specimen | Bioclimatic Stage |
|---|---|---|---|---|
| E. occidentalis/leaves | July 2021 | Djebel Mansour, Zaghouan | EOC2143 | Semi-arid |
| E. striaticalyx/leaves | ES2145 | |||
| E. stricklandii/leaves | ESI2145 | |||
| L. rigidum/seeds | June 2021 | Sidi Ismail, Beja | LR2105 | Subhumid with mild winter |
| S. arvensis/seeds | SA2103 | |||
| T. campestre/seeds | TC2101 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Khammassi, M.; Polito, F.; Amri, I.; Khedhri, S.; Hamrouni, L.; Nazzaro, F.; Fratianni, F.; De Feo, V. Chemical Composition and Phytotoxic, Antibacterial and Antibiofilm Activity of the Essential Oils of Eucalyptus occidentalis, E. striaticalyx and E. stricklandii. Molecules 2022, 27, 5820. https://doi.org/10.3390/molecules27185820
Khammassi M, Polito F, Amri I, Khedhri S, Hamrouni L, Nazzaro F, Fratianni F, De Feo V. Chemical Composition and Phytotoxic, Antibacterial and Antibiofilm Activity of the Essential Oils of Eucalyptus occidentalis, E. striaticalyx and E. stricklandii. Molecules. 2022; 27(18):5820. https://doi.org/10.3390/molecules27185820
Chicago/Turabian StyleKhammassi, Marwa, Flavio Polito, Ismail Amri, Sana Khedhri, Lamia Hamrouni, Filomena Nazzaro, Florinda Fratianni, and Vincenzo De Feo. 2022. "Chemical Composition and Phytotoxic, Antibacterial and Antibiofilm Activity of the Essential Oils of Eucalyptus occidentalis, E. striaticalyx and E. stricklandii" Molecules 27, no. 18: 5820. https://doi.org/10.3390/molecules27185820
APA StyleKhammassi, M., Polito, F., Amri, I., Khedhri, S., Hamrouni, L., Nazzaro, F., Fratianni, F., & De Feo, V. (2022). Chemical Composition and Phytotoxic, Antibacterial and Antibiofilm Activity of the Essential Oils of Eucalyptus occidentalis, E. striaticalyx and E. stricklandii. Molecules, 27(18), 5820. https://doi.org/10.3390/molecules27185820