
Glucogallin Attenuates RAW 264.7 Cells from Arsenic Trioxide Induced Toxicity via the NF-ҡB/NLRP3 Pathway


 , , ,
, , , {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Materials
2.2. Cell Culture
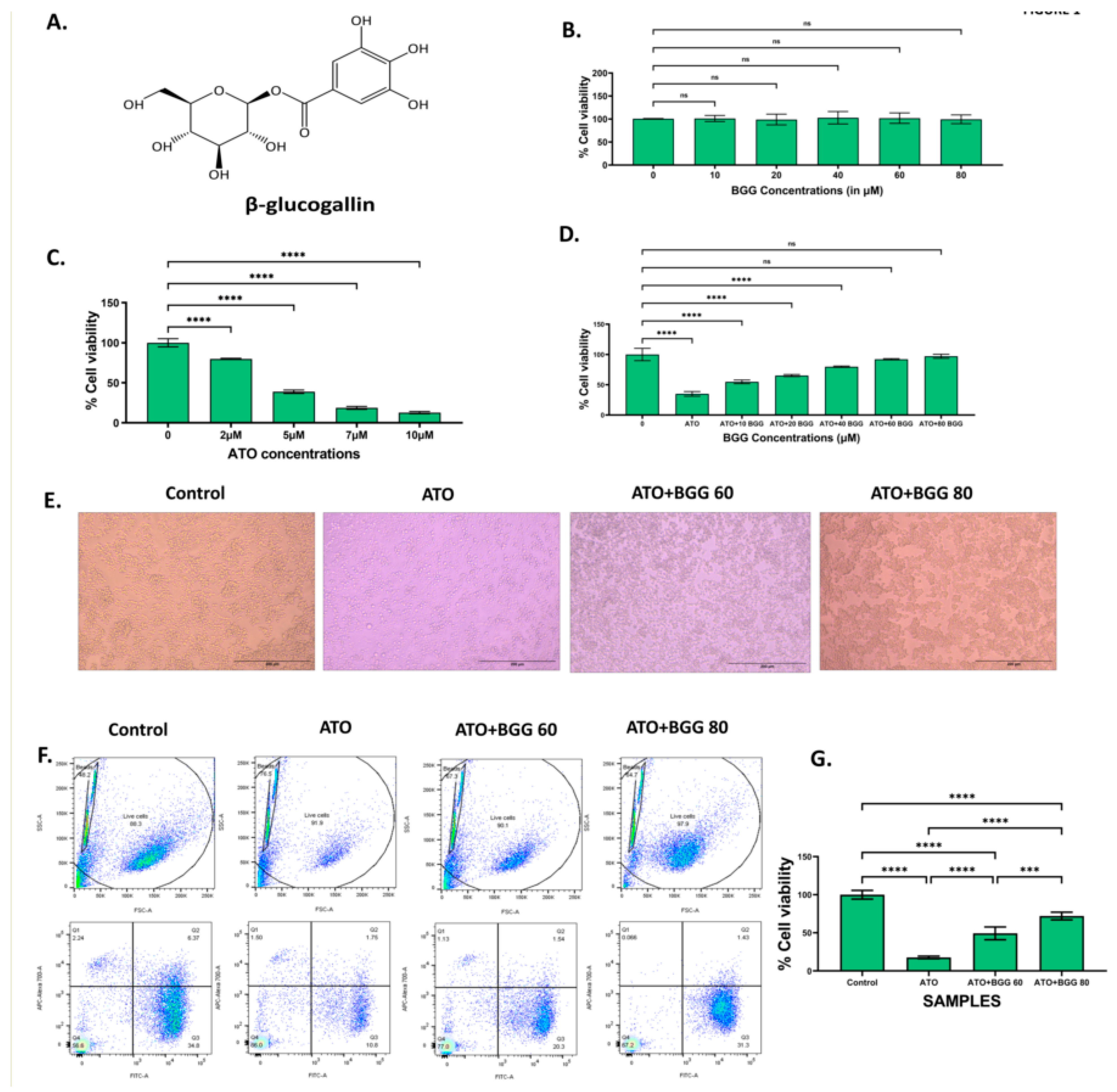
2.3. Cell Viability Assay
2.4. Calcine-AM and PI staining
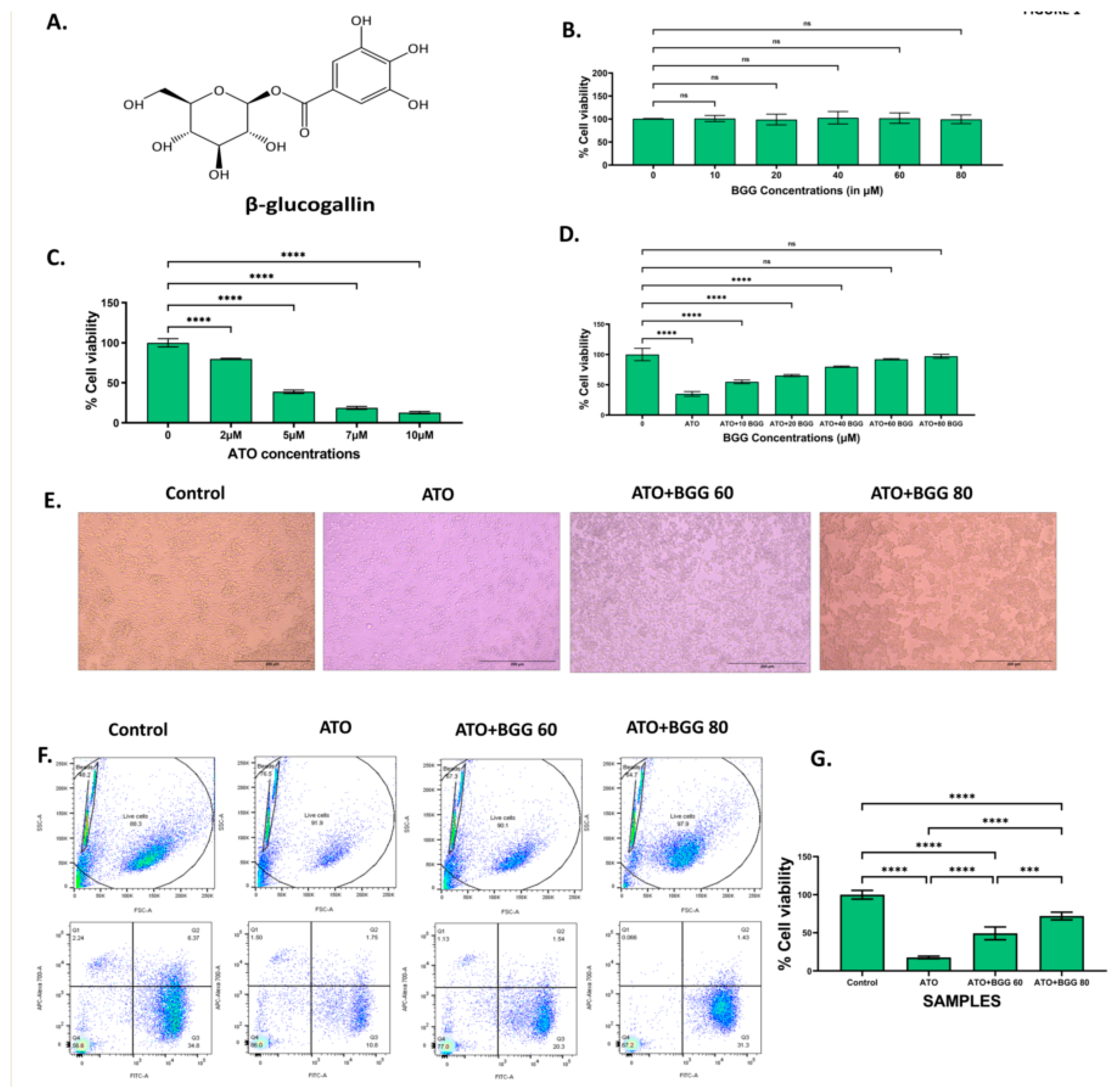
2.5. Quantification of Intracellular ROS Formation
2.6. Measurement of Mitochondrial Membrane Potential by JC-1
2.7. Measurement of Intracellular GSH Activity
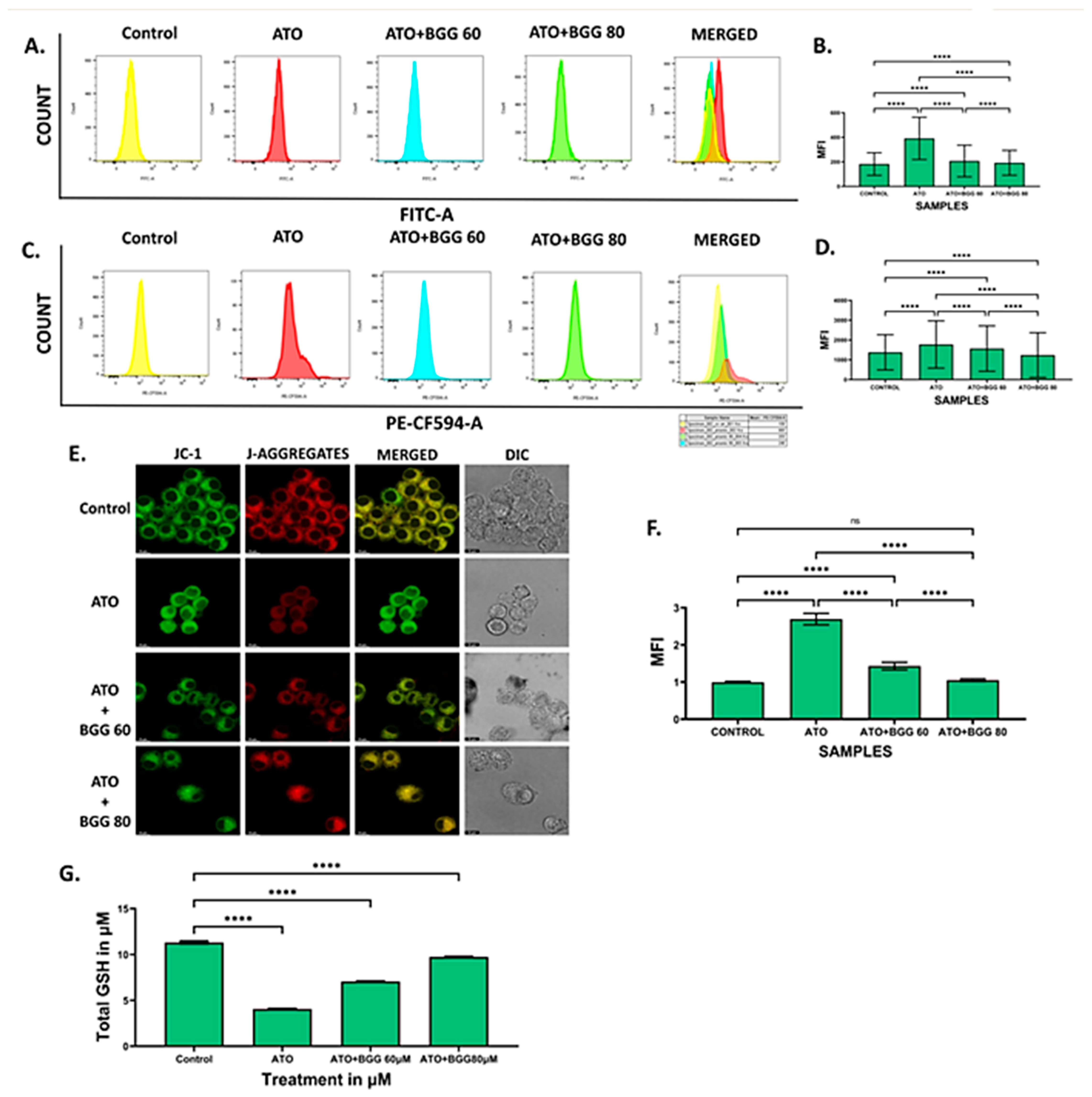
2.8. Cytometric Bead Array (CBA) for Extracellular Cytokines
2.9. Intracellular IL6 and IFN Production
2.10. Apoptotic Cell Quantification by Annexin V and PI
2.11. Western Blot Analysis
2.12. Study of PARP1/NFҡβ/NLRP3//IL-1β Expression by Confocal Microscopy
2.13. Statistical Analysis
3. Results
3.1. Effect of BGG against ATO Induced Cell Death
3.2. Effect of BGG against ATO-Induced Intracellular ROS Formation
3.3. Effect of BGG on ATO-Induced Mitochondrial Dysfunction
3.4. Effect of BGG on GSH Activity
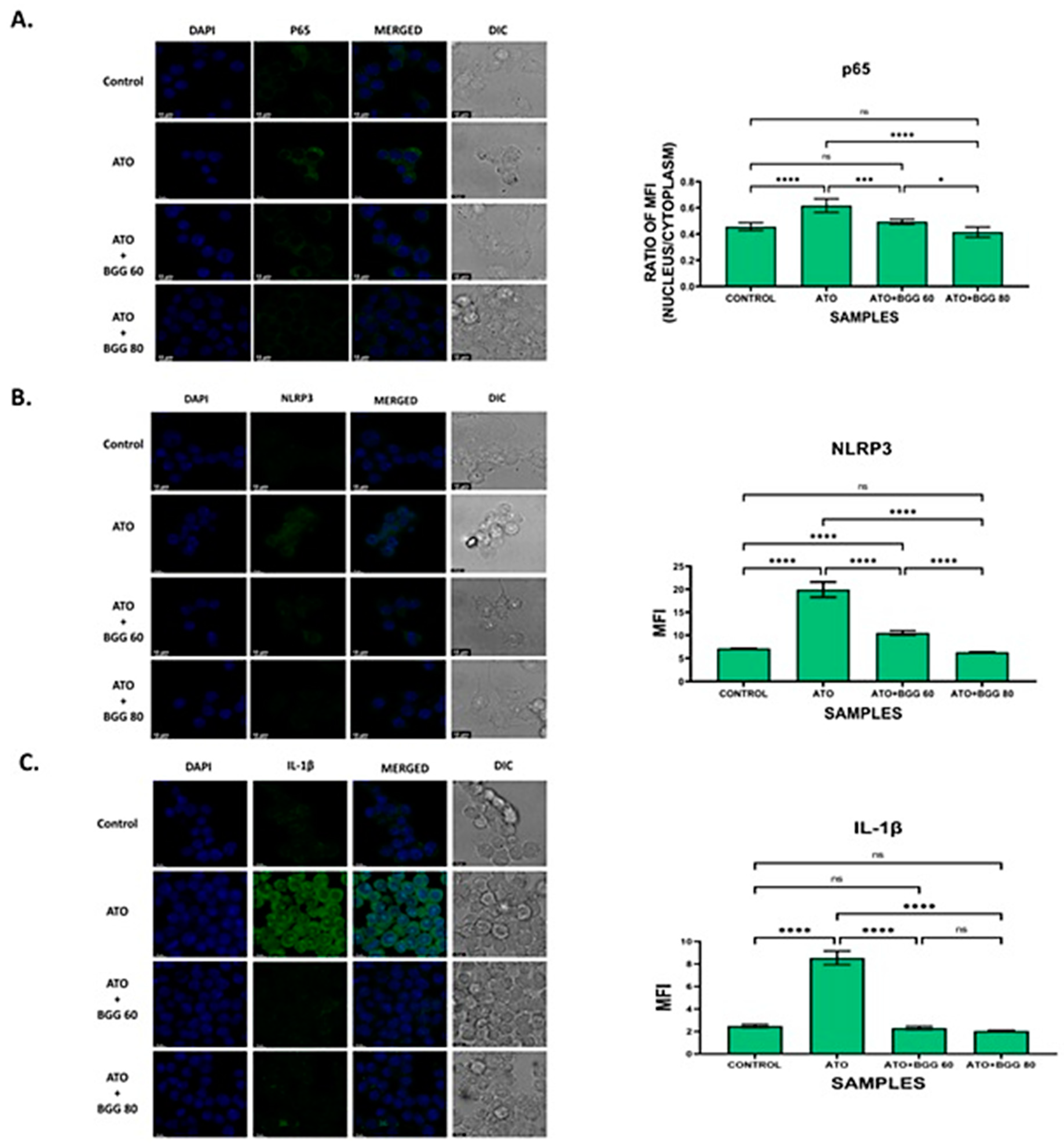
3.5. Effect of BGG against ATO-Induced Pro-Inflammatory Cytokines Productions and Inflammation in RAW 264.7 Macrophages Cells
3.5.1. Extracellular Screening of Interleukins
3.5.2. Intracellular Staining of Interleukins
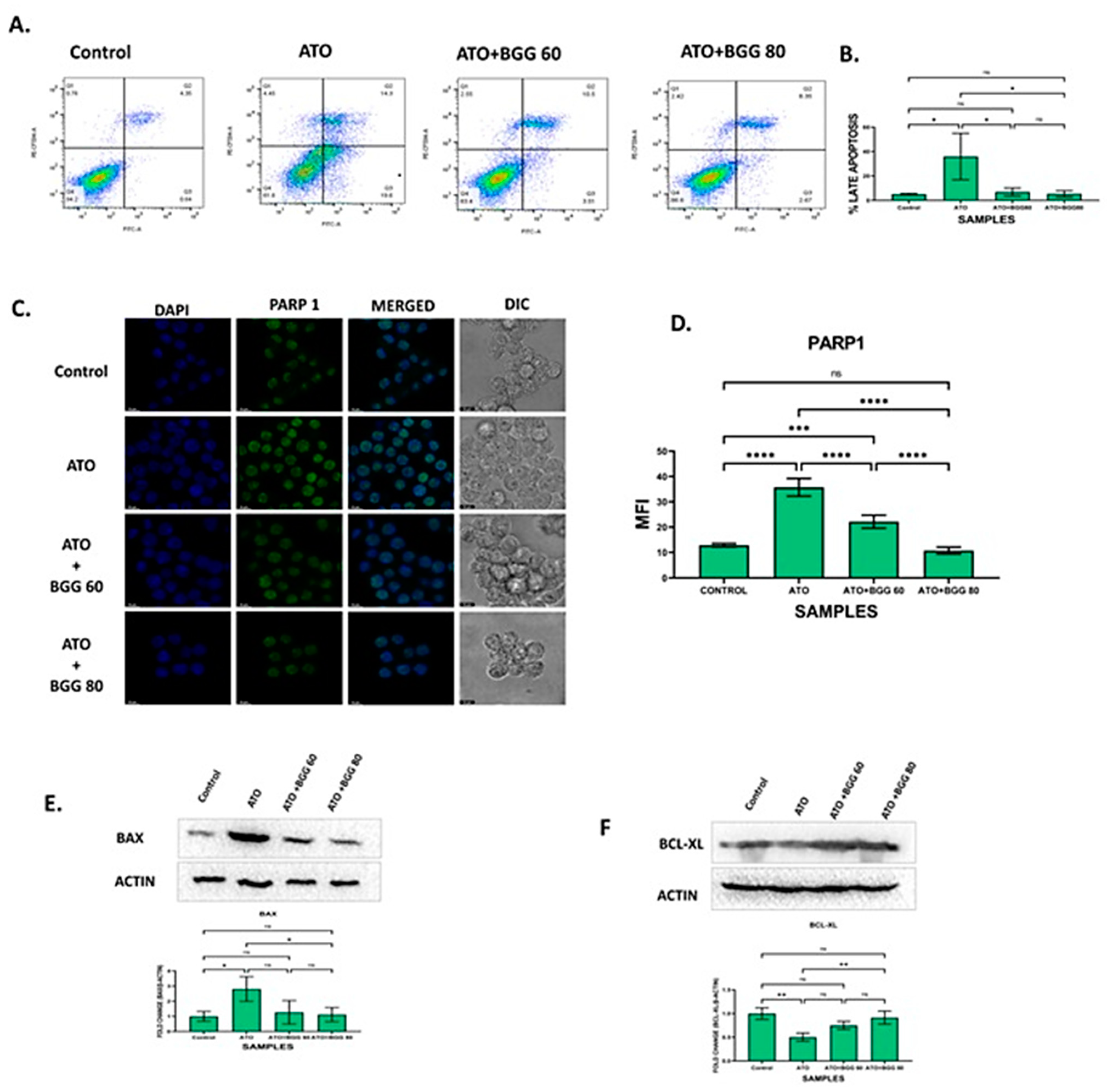
3.6. BGG Prevents ATO-Induced Apoptosis
3.7. Effect of BGG on ATO and Protein Expression of BAX and BCL-XL
3.8. Effect of BGG on ATO-Induced p-65/NLRP3/PARP1/IL-1β Pathway Activation in RAW 264.7 Macrophages
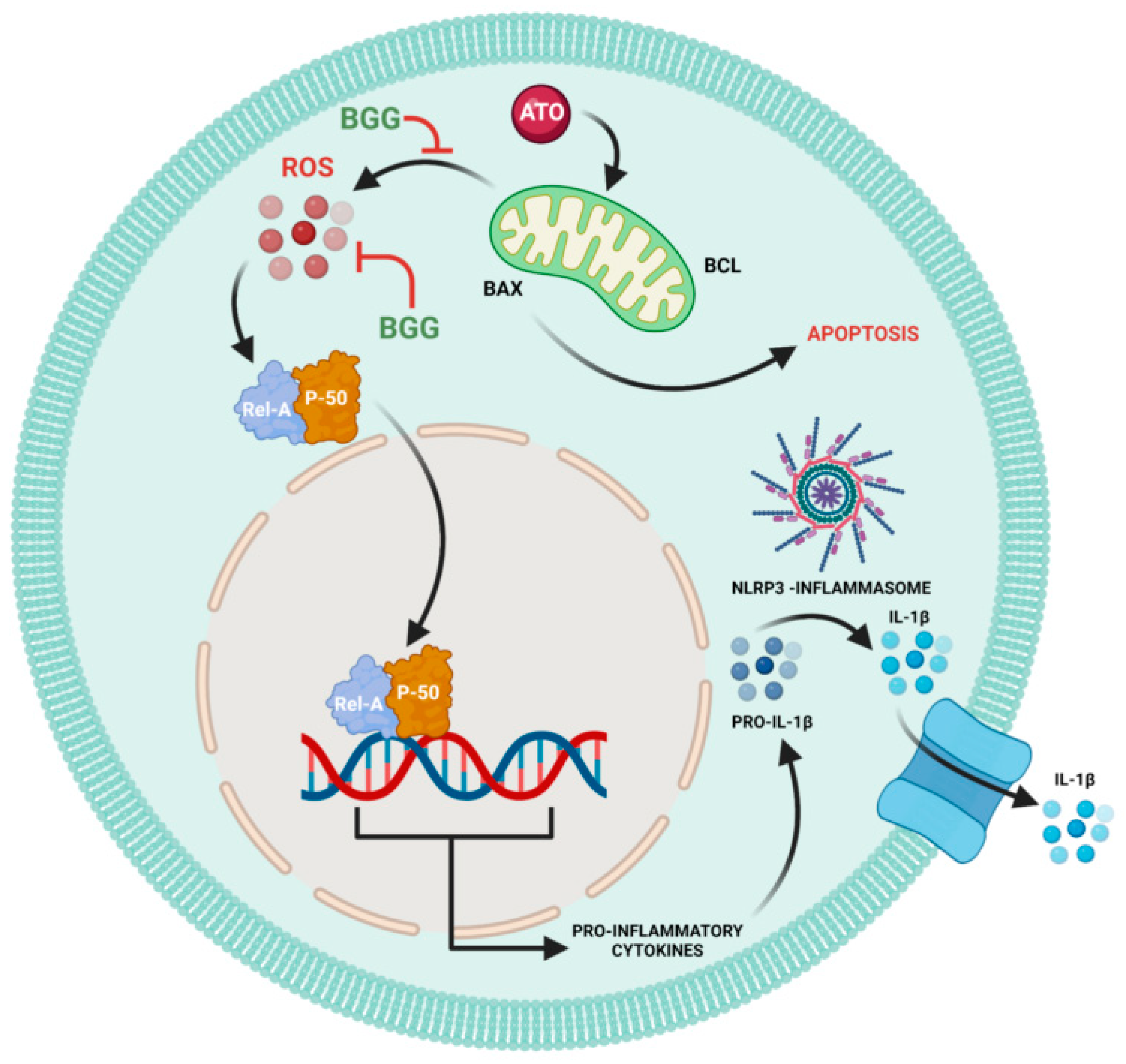
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Sample Availability
References
- World Health Organization. Arsenicosis case-detection, management and surveillance: Report of a regional consultation. In Proceedings of the Arsenicosis Case-Detection, Management and Surveillance: Report of a Regional Consultation, New Delhi, India, 5–9 November 2002. [Google Scholar]
- Saha, J.; Dikshit, A.; Bandyopadhyay, M.; Saha, K. A review of arsenic poisoning and its effects on human health. Crit. Rev. Environ. Sci. Technol. 1999, 29, 281–313. [Google Scholar] [CrossRef]
- Cohen, S.M.; Arnold, L.L.; Eldan, M.; Lewis, A.S.; Beck, B.D. Methylated arsenicals: The implications of metabolism and carcinogenicity studies in rodents to human risk assessment. Crit. Rev. Toxicol. 2006, 36, 99–133. [Google Scholar] [CrossRef]
- Pohl, C. Use of Ion Chromatography for Monitoring Ionic Contaminants in Water. In Chemistry and Water; Elsevier: Amsterdam, The Netherlands, 2017; pp. 353–391. [Google Scholar]
- Flora, S.; Bhadauria, S.; Kannan, G.; Singh, N. Arsenic induced oxidative stress and the role of antioxidant supplementation during chelation: A review. J. Environ. Biol. 2007, 28, 333. [Google Scholar]
- Picca, A.; Guerra, F.; Calvani, R.; Bucci, C.; Lo Monaco, M.R.; Bentivoglio, A.R.; Coelho-Júnior, H.J.; Landi, F.; Bernabei, R.; Marzetti, E. Mitochondrial dysfunction and aging: Insights from the analysis of extracellular vesicles. Int. J. Mol. Sci. 2019, 20, 805. [Google Scholar] [CrossRef]
- Zhang, W.; Liu, Y.; Ge, M.; Yao, C.; Xue, J.; Zhang, Z. Resveratrol reduces oxidative stress and improves arsenic efflux in rats exposed to arsenic trioxide. Pak. Vet. J. 2014, 34, 251–253. [Google Scholar]
- Parodi-Rullán, R.M.; Soto-Prado, J.; Vega-Lugo, J.; Chapa-Dubocq, X.; Díaz-Cordero, S.I.; Javadov, S. Divergent effects of cyclophilin-D inhibition on the female rat heart: Acute versus chronic post-myocardial infarction. Cell. Physiol. Biochem. 2018, 50, 288–303. [Google Scholar] [CrossRef]
- Valko, M.; Jomova, K.; Rhodes, C.J.; Kuča, K.; Musílek, K. Redox-and non-redox-metal-induced formation of free radicals and their role in human disease. Arch. Toxicol. 2016, 90, 1–37. [Google Scholar] [CrossRef]
- Jomova, K.; Jenisova, Z.; Feszterova, M.; Baros, S.; Liska, J.; Hudecova, D.; Rhodes, C.; Valko, M. Arsenic: Toxicity, oxidative stress and human disease. J. Appl. Toxicol. 2011, 31, 95–107. [Google Scholar] [CrossRef]
- Tseng, C.-H. An overview on peripheral vascular disease in blackfoot disease-hyperendemic villages in Taiwan. Angiology 2002, 53, 529–537. [Google Scholar] [CrossRef]
- Bhattacharya, S. Medicinal plants and natural products in amelioration of arsenic toxicity: A short review. Pharm. Biol. 2017, 55, 349–354. [Google Scholar] [CrossRef]
- Mohnen, D. Carbohydrates and Their Derivatives Including Tannins, Cellulose, and Related Lignins; Elsevier Science LTD: Oxford, UK, 1999; pp. 497–527. [Google Scholar]
- Khan, A.N.; Singh, R.; Bhattacharya, A.; Chakravarti, R.; Roy, S.; Ravichandiran, V.; Ghosh, D. A Short Review on Glucogallin and its Pharmacological Activities. Mini Rev. Med. Chem. 2022. [Google Scholar] [CrossRef] [PubMed]
- Ribeiro, S.M.R.; Schieber, A. Bioactive compounds in mango (Mangifera indica L.). In Bioactive Foods in Promoting Health; Elsevier: Amsterdam, The Netherlands, 2010; pp. 507–523. [Google Scholar]
- Gross, G. Synthesis of β-glucogallin from UDP-glucose and gallic acid by an enzyme preparation from oak leaves. FEBS Lett. 1982, 148, 67–70. [Google Scholar] [CrossRef]
- Singh, R.; Gautam, A.; Chandel, S.; Ghosh, A.; Dey, D.; Roy, S.; Ravichandiran, V.; Ghosh, D. Protease Inhibitory Effect of Natural Polyphenolic Compounds on SARS-CoV-2: An In Silico Study. Molecules 2020, 25, 4604. [Google Scholar] [CrossRef]
- Puppala, M.; Ponder, J.; Suryanarayana, P.; Reddy, G.B.; Petrash, J.M.; LaBarbera, D.V. The isolation and characterization of β-glucogallin as a novel aldose reductase inhibitor from Emblica officinalis. PLoS ONE 2012, 7, e31399. [Google Scholar] [CrossRef]
- Niimi, N.; Yako, H.; Takaku, S.; Chung, S.K.; Sango, K. Aldose reductase and the polyol pathway in schwann cells: Old and new problems. Int. J. Mol. Sci. 2021, 22, 1031. [Google Scholar] [CrossRef]
- Cao, T.; Wang, J.; Wu, Y.; Wang, L.; Zhang, H. Antiglaucoma Potential of β-Glucogallin Is Mediated by Modulating Mitochondrial Responses in Experimentally Induced Glaucoma. Neuroimmunomodulation 2020, 27, 142–151. [Google Scholar] [CrossRef]
- Majeed, M.; Majeed, S.; Pande, A.; Karri, S.K. Amelioration of carbon tetrachloride induced hepatotoxicity by Β-glucogallin, a gallic acid derivative of emblica officinalis gaertn.(euphorbiaceae). Science 2015, 4, 696–701. [Google Scholar]
- Kim, H.-G.; Kim, K.S.; Kim, M.; Shin, S.-H.; Lee, Y.-G.; Bang, M.-H.; Lee, D.-G.; Baek, N.-I. β-Glucogallin isolated from Fusidium coccineum and its enhancement of skin barrier effects. Appl. Biol. Chem. 2020, 63, 77. [Google Scholar] [CrossRef]
- Majeed, M.; Bhat, B.; Anand, T.S. Inhibition of UV induced adversaries by β-glucogallin from Amla (Emblica officinalis Gaertn.) fruits. Indian J. Nat. Prod. Resour. 2010, 1, 462–465. [Google Scholar]
- Grela, E.; Kozłowska, J.; Grabowiecka, A. Current methodology of MTT assay in bacteria–A review. Acta Histochem. 2018, 120, 303–311. [Google Scholar] [CrossRef]
- Gatti, R.; Belletti, S.; Orlandini, G.; Bussolati, O.; Dall’Asta, V.; Gazzola, G.C.J. Comparison of annexin V and calcein-AM as early vital markers of apoptosis in adherent cells by confocal laser microscopy. J. Histochem. Cytochem. 1998, 46, 895–900. [Google Scholar] [CrossRef]
- Menon, V.P.; Sudheer, A.R. Antioxidant and anti-inflammatory properties of curcumin. Mol. Targets Ther. Uses Curcumin Health Dis. 2007, 105–125. [Google Scholar]
- Liu, T.; Zhang, L.; Joo, D.; Sun, S.-C. NF-κB signaling in inflammation. Signal Transduct. Target. Ther. 2017, 2, 17023. [Google Scholar] [CrossRef]
- Giridharan, S.; Srinivasan, M. Mechanisms of NF-κB p65 and strategies for therapeutic manipulation. J. Inflamm. Res. 2018, 11, 407. [Google Scholar] [CrossRef]
- He, Y.; Hara, H.; Núñez, G. Mechanism and regulation of NLRP3 inflammasome activation. Trends Biochem. Sci. 2016, 41, 1012–1021. [Google Scholar] [CrossRef]
- Shaji, E.; Santosh, M.; Sarath, K.; Prakash, P.; Deepchand, V.; Divya, B. Arsenic contamination of groundwater: A global synopsis with focus on the Indian Peninsula. Geosci. Front. 2021, 12, 101079. [Google Scholar] [CrossRef]
- Loker, E.S.; Adema, C.M.; Zhang, S.M.; Kepler, T.B. Invertebrate immune systems–not homogeneous, not simple, not well understood. Immunol. Rev. 2004, 198, 10–24. [Google Scholar] [CrossRef]
- Cohen, S.M.; Ohnishi, T.; Arnold, L.L.; Le, X.C. Arsenic-induced bladder cancer in an animal model. Toxicol. Appl. Pharmacol. 2007, 222, 258–263. [Google Scholar] [CrossRef]
- Huang, H.-W.; Lee, C.-H.; Yu, H.-S. Arsenic-induced carcinogenesis and immune dysregulation. Int. J. Environ. Res. Public Health 2019, 16, 2746. [Google Scholar] [CrossRef]
- Linden, G.; Janga, H.; Franz, M.; Nist, A.; Stiewe, T.; Schmeck, B.; Vázquez, O.; Schulte, L.N. Efficient antisense inhibition reveals microRNA-155 to restrain a late-myeloid inflammatory programme in primary human phagocytes. RNA Biol. 2021, 18, 604–618. [Google Scholar] [CrossRef]
- Eghbaliferiz, S.; Iranshahi, M. Prooxidant activity of polyphenols, flavonoids, anthocyanins and carotenoids: Updated review of mechanisms and catalyzing metals. Phytother. Res. 2016, 30, 1379–1391. [Google Scholar] [CrossRef]
- Sener, U.; Uygur, R.; Aktas, C.; Uygur, E.; Erboga, M.; Balkas, G.; Caglar, V.; Kumral, B.; Gurel, A.; Erdogan, H. Protective effects of thymoquinone against apoptosis and oxidative stress by arsenic in rat kidney. Ren. Fail. 2016, 38, 117–123. [Google Scholar] [CrossRef]
- Hu, Y.; Li, J.; Lou, B.; Wu, R.; Wang, G.; Lu, C.; Wang, H.; Pi, J.; Xu, Y. The role of reactive oxygen species in arsenic toxicity. Biomolecules 2020, 10, 240. [Google Scholar] [CrossRef]
- Lambeth, J.D. Nox/Duox family of nicotinamide adenine dinucleotide (phosphate) oxidases. Curr. Opin. Hematol. 2002, 9, 11–17. [Google Scholar] [CrossRef]
- Gill, S.S.; Tuteja, N. Reactive oxygen species and antioxidant machinery in abiotic stress tolerance in crop plants. Plant Physiol. Biochem. 2010, 48, 909–930. [Google Scholar] [CrossRef]
- Khan, S.S.; Sharma, A.; Flora, S.J. Phytochemicals in the Management of Arsenic Toxicity. Chem. Res. Toxicol. 2022, 35, 916–934. [Google Scholar] [CrossRef]
- Tam, L.M.; Price, N.E.; Wang, Y. Molecular mechanisms of arsenic-induced disruption of DNA repair. Chem. Res. Toxicol. 2020, 33, 709–726. [Google Scholar] [CrossRef]
- Prakash, C.; Soni, M.; Kumar, V. Mitochondrial oxidative stress and dysfunction in arsenic neurotoxicity: A review. J. Appl. Toxicol. 2016, 36, 179–188. [Google Scholar] [CrossRef]
- Chatterjee, A.; Chatterji, U. All-trans retinoic acid ameliorates arsenic-induced oxidative stress and apoptosis in the rat uterus by modulating MAPK signaling proteins. J. Cell. Biochem. 2017, 118, 3796–3809. [Google Scholar] [CrossRef]
- Barchowsky, A.; Dudek, E.J.; Treadwell, M.D.; Wetterhahn, K.E. Arsenic induces oxidant stress and NF-kB activation in cultured aortic endothelial cells. Free Radic. Biol. Med. 1996, 21, 783–790. [Google Scholar] [CrossRef]
- Li, W.; Cao, T.; Luo, C.; Cai, J.; Zhou, X.; Xiao, X.; Liu, S. Crosstalk between ER stress, NLRP3 inflammasome, and inflammation. Appl. Microbiol. Biotechnol. 2020, 104, 6129–6140. [Google Scholar] [CrossRef] [PubMed]
- Renu, K.; Chakraborty, R.; Myakala, H.; Koti, R.; Famurewa, A.C.; Madhyastha, H.; Vellingiri, B.; George, A.; Gopalakrishnan, A.V. Molecular mechanism of heavy metals (Lead, Chromium, Arsenic, Mercury, Nickel and Cadmium)-induced hepatotoxicity–A review. Chemosphere 2021, 271, 129735. [Google Scholar] [CrossRef] [PubMed]
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Khan, A.N.; Singh, R.; Bhattacharya, A.; Kumar, S.; Ghosh, A.; Nag, D.; Ravichandiran, V.; Ghosh, D. Glucogallin Attenuates RAW 264.7 Cells from Arsenic Trioxide Induced Toxicity via the NF-ҡB/NLRP3 Pathway. Molecules 2022, 27, 5263. https://doi.org/10.3390/molecules27165263
Khan AN, Singh R, Bhattacharya A, Kumar S, Ghosh A, Nag D, Ravichandiran V, Ghosh D. Glucogallin Attenuates RAW 264.7 Cells from Arsenic Trioxide Induced Toxicity via the NF-ҡB/NLRP3 Pathway. Molecules. 2022; 27(16):5263. https://doi.org/10.3390/molecules27165263
Chicago/Turabian StyleKhan, Anam Najib, Rajveer Singh, Arka Bhattacharya, Sonu Kumar, Arijit Ghosh, Debasish Nag, Velayutham Ravichandiran, and Dipanjan Ghosh. 2022. "Glucogallin Attenuates RAW 264.7 Cells from Arsenic Trioxide Induced Toxicity via the NF-ҡB/NLRP3 Pathway" Molecules 27, no. 16: 5263. https://doi.org/10.3390/molecules27165263
APA StyleKhan, A. N., Singh, R., Bhattacharya, A., Kumar, S., Ghosh, A., Nag, D., Ravichandiran, V., & Ghosh, D. (2022). Glucogallin Attenuates RAW 264.7 Cells from Arsenic Trioxide Induced Toxicity via the NF-ҡB/NLRP3 Pathway. Molecules, 27(16), 5263. https://doi.org/10.3390/molecules27165263