
Antibiofilm and Anti-Quorum Sensing Potential of Cycloartane-Type Triterpene Acids from Cameroonian Grassland Propolis: Phenolic Profile and Antioxidant Activity of Crude Extract

 ,
,  ,
, 
Abstract
:1. Introduction
2. Results
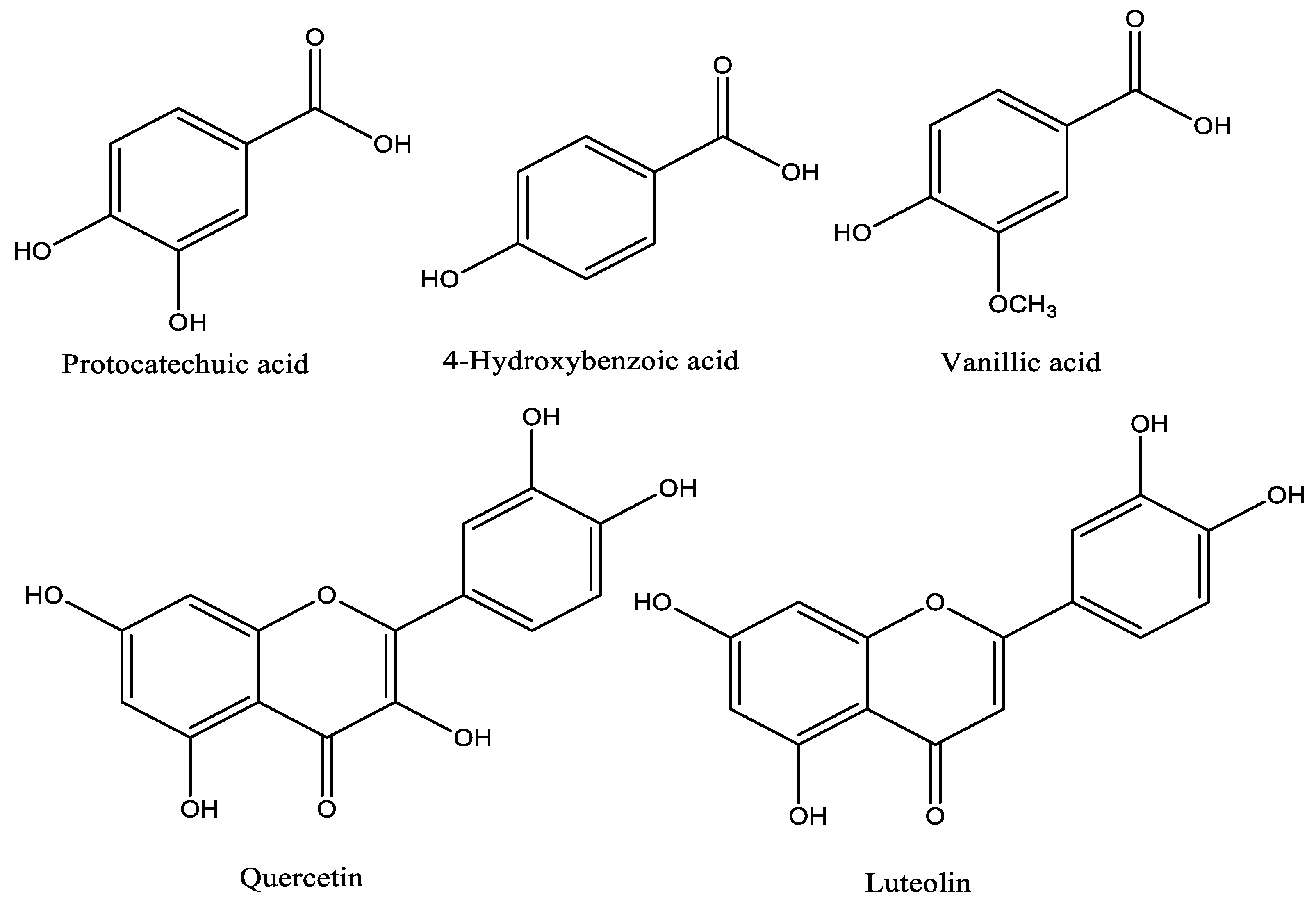
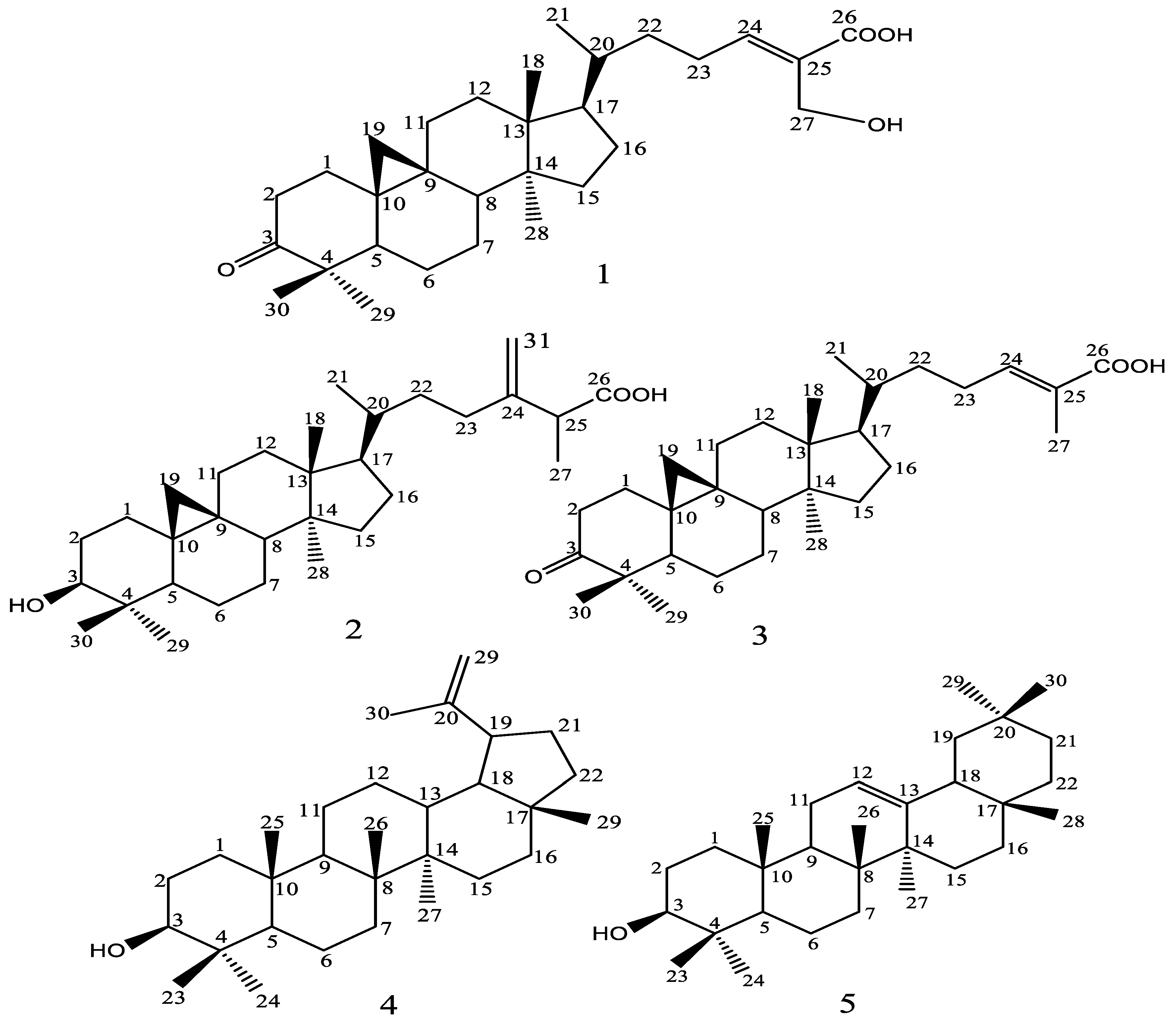
2.1. HPLC-DAD Phenolic Profile and NMR Data of Isolated Compounds
2.2. Antioxidant Activity
2.3. Violacein Inhibition and Anti-Quorum Sensing Activity
2.4. Swarming Motility Inhibition
2.5. Antimicrobial and Antibiofilm Activities
3. Discussion
4. Materials and Methods
4.1. Propolis Collection and Extraction
4.2. Isolation and Characterization of Compounds
4.3. HPLC-DAD Phenolic Profiling
4.4. Antioxidant Activity
4.5. Cyclic Voltammetry Measurements
4.6. Microbial Strains
4.7. Determination of Minimal Inhibitory Concentrations
4.8. Effect of Extract on Bacterial Biofilm Formation
4.9. Bioassay for Quorum-Sensing Inhibition (QSI) Activity Using C. violacium CV026
4.10. Violacein Inhibition Assay Using C. violacium CV12472
4.11. Swarming Motility Inhibition on P. aeruginosa PA01
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Sample Availability
References
- Fitzpatrick, M.C.; Bauch, C.T.; Townsend, J.P.; Galvani, A.P. Modelling microbial infection to address global health challenges. Nat. Microbiol. 2019, 4, 1612–1619. [Google Scholar] [CrossRef] [PubMed]
- Ngenge, A.; Ceylan, O.; Fru, G.; Arab, Y.; Emin, D.; Ozturk, M. Antimicrobial, antibiofilm, anti-quorum sensing and motility inhibition activities of essential oil from seeds of food spice Xylopia aethiopica (Dunal) A. Rich. on some pathogenic bacteria. Res. J. Biotechnol. 2021, 16, 68–76. [Google Scholar]
- Prestinaci, F.; Pezzotti, P.; Pantosti, A. Antimicrobial resistance: A global multifaceted phenomenon. Pathog. Glob. Health 2015, 109, 309–318. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guzzo, F.; Scognamiglio, M.; Fiorentino, A.; Buommino, E.; D’Abrosca, B. Plant derived natural products against Pseudomonas aeruginosa and Staphylococcus aureus: Antibiofilm activity and molecular mechanisms. Molecules 2020, 25, 5024. [Google Scholar] [CrossRef]
- Subramani, R.; Narayanasamy, M.; Feussner, K.D. Plant-derived antimicrobials to fight against multi-drug-resistant human pathogens. 3 Biotech 2017, 7, 172. [Google Scholar] [CrossRef]
- Mishra, B.; Wang, G. Individual and combined effects of engineered peptides and antibiotics on Pseudomonas aeruginosa biofilms. Pharmaceuticals 2017, 10, 58. [Google Scholar] [CrossRef] [Green Version]
- Lebeaux, D.; Ghigo, J.M.; Beloin, C. Biofilm-related infections: Bridging the gap between clinical management and fundamental aspects of recalcitrance toward antibiotics. Microbiol. Mol. Biol. Rev. 2014, 78, 510–543. [Google Scholar] [CrossRef] [Green Version]
- Lemos, A.; Campos, L.M.; Melo, L.; Guedes, M.; Oliveira, L.G.; Silva, T.P.; Melo, R.; Rocha, V.N.; Aguiar, J.; Apolônio, A.; et al. Antibacterial and Antibiofilm Activities of Psychorubrin, a Pyranonaphthoquinone Isolated From Mitracarpus frigidus (Rubiaceae). Front. Microbial. 2018, 9, 724. [Google Scholar] [CrossRef] [Green Version]
- Tamfu, A.N.; Ceylan, O.; Fru, G.C.; Ozturk, M.; Duru, M.E.; Shaheen, F. Antibiofilm, antiquorum sensing and antioxidant activity of secondary metabolites from seeds of Annona senegalensis, Persoon. Mic. Pathogen. 2020, 144, 104191. [Google Scholar] [CrossRef]
- González-Palma, I.; Escalona-Buendía, H.B.; Ponce-Alquicira, E.; Téllez-Téllez, M.; Gupta, V.K.; Díaz-Godínez, G.; Soriano-Santos, J. Evaluation of the antioxidant activity of aqueous and methanol extracts of Pleurotus ostreatus in different growth stages. Fronts. Microbial. 2016, 7, 1099. [Google Scholar] [CrossRef] [Green Version]
- Koh, C.L.; Sam, C.K.; Yin, W.F.; Tan, L.Y.; Krishnan, T.; Chong, Y.M.; Chan, K.G. Plant-derived natural products as sources of anti-quorum sensing compounds. Sensors 2013, 13, 6217–6228. [Google Scholar] [CrossRef] [Green Version]
- Bankova, V.; Bertelli, D.; Borba, R.S.; Conti, B.J.; Cunha, I.B.; Danert, C.; Eberlin, M.; Falcão, S.I.; Isla, M.; Moreno, M.I.; et al. Standard methods for Apis mellifera propolis research. J. Apic. Res. 2016, 58, 1–49. [Google Scholar] [CrossRef] [Green Version]
- Tamfu, A.N.; Domgnim, M.E.C.; Talla, E.; Tan, P.V.; Mbafor, T.J.; Popova, M.; Bankova, V. Chemical constituents and anti-ulcer activity of propolis from the North-West region of Cameroon. Res. J. Phytochem. 2016, 10, 45–57. [Google Scholar] [CrossRef]
- Tamfu, A.N.; Sawalda, M.; Fotsing, M.T.; Kouipou, R.M.T.; Talla, E.; Chi, G.F.; Epanda, J.J.E.; Mbafor, J.T.; Baig, T.A.; Jabeen, A.; et al. A new isoflavonol and other constituents from Cameroonian propolis and evaluation of their anti-inflammatory, antifungal and antioxidant potential. Saudi J. Biol. Sci. 2020, 27, 1659–1666. [Google Scholar] [CrossRef]
- Almuhayawi, M.S. Propolis as a novel antibacterial agent. Saudi J. Biol. Sci. 2020, 27, 3079–3086. [Google Scholar] [CrossRef]
- Siheri, W.; Alenezi, S.; Tusiimire, J.; Watson, D.G. The chemical and biological properties of propolis. In Bee Products—Chemical and Biological Properties; Springer: Cham, Switzerland, 2017; pp. 137–178. [Google Scholar] [CrossRef]
- Tamfu, A.N.; Tagatsing, M.F.; Talla, E.; Mbafor, J.T. Chemical profiles and bioactive molecules of propolis: A review. J. Nat. Prod. Resour. 2019, 5, 220–226. [Google Scholar] [CrossRef]
- Monzote, L.; Cuesta-Rubio, O.; Fernandez, M.C.; Hernandez, I.M.; Fraga, J.; Pérez, K.; Kerstens, M.; Maes, L.; Cos, P. In vitro antimicrobial assessment of Cuban propolis extracts. Mem. Inst. Oswaldo Cruz Rio De Jan. 2012, 107, 978–984. [Google Scholar] [CrossRef] [Green Version]
- Mahamat, A.A.; Nyemb, J.N.; Gade, I.S.; Ngenge, A.T.; Talla, E.; Céline, H.; Sophie, L.; Mbafor, J.T. A new fatty acid and some triterpenoids from propolis of Nkambe (North-West Region, Cameroon) and evaluation of the antiradical scavenging activity of their extracts. Open Chem. 2020, 18, 239–243. [Google Scholar] [CrossRef]
- Talla, E.; Tamfu, A.N.; Gade, I.S.; Yanda, L.; Mbafor, J.T.; Sophie, L.; Vander Elst, L.; Popova, M.; Bankova, V. New mono-ether of glycerol and triterpenes with DPPH radical scavenging activity from Cameroonian propolis. Nat. Prod. Res. 2017, 31, 1379–1389. [Google Scholar] [CrossRef]
- Tamfu, A.N.; Tagatsing, F.M.; Talla, E.; Ozturk, M.; Mbafor, J.T.; Duru, M.E.; Farzana, S. Chemical composition and evaluation of anticholinesterase activity of essential oil from Cameroonian propolis. Issues Biol. Sci. Pharm. Res. 2019, 7, 58–63. [Google Scholar] [CrossRef]
- Papachroni, D.; Graikou, K.; Kosalec, I.; Damianakos, H.; Ingram, V.; Chinou, I. Phytochemical analysis and biological evaluation of selected African propolis samples from Cameroon and Congo. Nat. Prod. Com. 2015, 10, 67–70. [Google Scholar] [CrossRef] [Green Version]
- Kardar, M.N.; Zhang, T.; Coxon, G.D.; Watson, D.G.; Fearnley, J.; Seidel, V. Characterisation of triterpenes and new phenolic lipids in Cameroonian propolis. Phytochemistry 2014, 106, 156–163. [Google Scholar] [CrossRef] [Green Version]
- Ingram, V.; Njikeu, J. Sweet, sticky, and sustainable social business. Ecol. Soc. 2011, 16, 3–7. [Google Scholar] [CrossRef]
- Miguel, M.G.; Nunes, S.; Dandlen, S.A.; Cavaco, A.M.; Antunes, M.D. Phenols and antioxidant activity of hydro-alcoholic extracts of propolis from Algarve, South of Portugal. Food Chem. Toxicol. 2010, 48, 3418–3423. [Google Scholar] [CrossRef]
- Sun, C.; Wu, Z.; Wang, Z.; Zhang, H. Effect of ethanol/water solvents on phenolic profiles and antioxidant properties of Beijing propolis extracts. Evi. Based Comp. Altern Med. 2015, 8, 1–9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Woźniak, M.; Mrówczyńska, L.; Waśkiewicz, A.; Rogoziński, T.; Ratajczak, I. Phenolic profile and antioxidant activity of propolis extracts from Poland. Nat. Prod. Com. 2019, 14, 1–7. [Google Scholar] [CrossRef]
- Neto, M.S.R.; Tintino, S.R.; da Silva, A.R.P.; Costa, M.S.; Boligon, A.A.; Matias, E.F.F.; de Queiroz Balbino, V.; Menezes, I.R.A.; Melo Coutinho, H.D. Seasonal variation of Brazilian red propolis: Antibacterial activity, synergistic effect and phytochemical screening. Food Chem. Toxicol. 2017, 107, 572–580. [Google Scholar] [CrossRef]
- Pablo, A.; Ulloa, J.V.; María, I.Á.; Mariela, L.; Seth, C.; Fernando, N.S. Effect of the addition of propolis extract on bioactive compounds and antioxidant activity of craft beer. J. Chem. 2017, 2017, 6716053. [Google Scholar] [CrossRef] [Green Version]
- Talla, E.; Tamfu, A.N.; Biyanzi, P.; Sakava, P.; Asobo, F.P.; Mbafor, J.T.; Tchuenguem, F.F.N.; Ndjouenkeu, R. Phytochemical screening, antioxidant activity, total polyphenols and flavonoids content of different extracts of propolis from Tekel (Ngaoundal, Adamawa region, Cameroon). J. Phytopharmacol. 2014, 3, 321–329. [Google Scholar] [CrossRef]
- De Marco, S.; Piccioni, M.; Pagiotti, R.; Pietrella, D. Antibiofilm and antioxidant activity of propolis and bud poplar resins versus Pseudomonas aeruginosa. Evid. Based Comp. Alt. Med. 2017, 2017, 5163575. [Google Scholar] [CrossRef] [Green Version]
- Martinello, M.; Mutinelli, F. Antioxidant activity in bee products: A Review. Antioxidants 2021, 10, 71. [Google Scholar] [CrossRef]
- Kocot, J.; Kiełczykowska, M.; Luchowska-Kocot, D.; Kurzepa, J.; Musik, I. Antioxidant potential of propolis, bee pollen, and royal jelly: Possible medical application. Oxid. Med. Cell. Longev. 2018, 2, 7074209. [Google Scholar] [CrossRef]
- Gülçin, I.; Bursal, E.; Sehitoglu, M.H.; Bilsel, M.; Gören, A.C. Polyphenol contents and antioxidant activity of lyophilized aqueous extract of propolis from Erzurum, Turkey. Food Chem. Toxicol. 2010, 48, 2227–2238. [Google Scholar] [CrossRef]
- Gülçin, I. Antioxidants and antioxidant methods: An updated overview. Arch. Toxicol. 2020, 94, 651–715. [Google Scholar] [CrossRef] [Green Version]
- Jiao, Y.; Kilmartin, P.A.; Fan, M.; Quek, S.Y. Assessment of phenolic contributors to antioxidant activity of new kiwifruit cultivars using cyclic voltammetry combined with HPLC. Food Chem. 2018, 268, 77–85. [Google Scholar] [CrossRef]
- Zielińska, D.; Turemko, M. Electroactive phenolic contributors and antioxidant capacity of flesh and peel of 11 apple cultivars measured by cyclic voltammetry and HPLC-DAD-MS/MS. Antioxidants 2020, 9, 1054. [Google Scholar] [CrossRef]
- Keyrouz, R.; Abasq, M.; Le Bourvellec, C.; Blanc, N.; Audibert, L.; ArGall, E.; Hauchard, D. Total phenolic contents, radical scavenging and cyclic voltammetry of seaweeds from Brittany. Food Chem. 2011, 126, 831–836. [Google Scholar] [CrossRef] [Green Version]
- Sforcin, J.M.; Bankova, V. Propolis: Is there a potential for the development of new drugs? J. Ethnopharmacol. 2011, 133, 253–260. [Google Scholar] [CrossRef]
- Petruzzi, L.; Rosaria Corbo, M.; Campaniello, D.; Speranza, B.; Sinigaglia, M.; Bevilacqua, A. Antifungal and antibacterial effect of propolis: A comparative hit for food-borne Pseudomonas, Enterobacteriaceae and Fungi. Foods 2020, 9, 559. [Google Scholar] [CrossRef]
- AL-Ani, I.; Zimmermann, S.; Reichling, J.; Wink, M. Antimicrobial activities of European propolis collected from various geographic origins alone and in combination with antibiotics. Medicines 2018, 5, 2. [Google Scholar] [CrossRef] [Green Version]
- Seidel, V.; Peyfoon, E.; Watson, D.G.; Fearnley, J. Comparative study of the antibacterial activity of propolis from different geographical and climatic zones. Phytother. Res. 2008, 22, 1256–1263. [Google Scholar] [CrossRef]
- Galletti, J.; Tobaldini-Valerio, F.K.; Silva, S.; Kioshima, É.S.; Trierveiler-Pereira, L.; Bruschi, M.; Negri, M.; Estivalet Svidzinski, T. Antibiofilm activity of propolis extract on Fusarium species from onychomycosis. Future Microbiol. 2017, 12, 1311–1321. [Google Scholar] [CrossRef] [Green Version]
- Grecka, K.; Kuś, P.M.; Okińczyc, P.; Worobo, R.W.; Walkusz, J.; Szweda, P. The anti-staphylococcal potential of ethanolic Polish propolis extracts. Molecules 2019, 24, 1732. [Google Scholar] [CrossRef] [Green Version]
- Fernández-Calderón, M.C.; Hernández-González, L.; Gómez-Navia, C.; Blanco-Blanco, M.T.; Sánchez-Silos, R.; Lucio, L.; Pérez-Giraldo, C. Antifungal and anti-biofilm activity of a new Spanish extract of propolis against Candida glabrata. BMC Complement. Med. Ther. 2021, 21, 147. [Google Scholar] [CrossRef]
- Djais, A.A.; Putri, N.; Jemmy, P.A.R.; Darwita, R.R.; Bachtiar, B.M. Effect of propolis on Streptococcus mutans biofilm formation. Pesqui. Bras. Odontopediatria Clin. Integr. 2019, 19, 1–9. [Google Scholar] [CrossRef]
- Santos, L.M.; Rodrigues, D.M.; Kalil, M.A.; Azevedo, V.; Meyer, R.; Umsza-Guez, M.A.; Machado, B.A.; Seyffert, N.; Portela, R.W. Activity of ethanolic and supercritical propolis extracts in Corynebacterium pseudotuberculosis and its associated biofilm. Front. Vet. Sci. 2021, 8, 700030. [Google Scholar] [CrossRef]
- Silva, G.N.S.D.; Primon-Barros, M.; Macedo, A.J.; Gnoatto, S.C.B. Triterpene derivatives as relevant scaffold for new antibiofilm drugs. Biomolecules 2019, 9, 58. [Google Scholar] [CrossRef] [Green Version]
- Evaristo, F.F.; Albuquerque, M.R.; dos Santos, H.S.; Bandeira, P.N.; Avila, F.N.; da Silva, B.R.; Vasconcelos, A.A.; Rabelo, E.M.; Nascimento-Neto, L.G.; Arruda, F.V.; et al. Antimicrobial effect of the triterpene 3β,6β,16β-trihydroxylup-20(29)-ene on planktonic cells and biofilms from Gram positive and Gram negative bacteria. Biomed. Res. Int. 2014, 2014, 729358. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Melchior, M.B.; Vaarkamp, H.; Fink-Gremmels, J. Biofilms: A role in recurrent mastitis infections? Vet. J. 2006, 171, 398–407. [Google Scholar] [CrossRef] [PubMed]
- Sycz, Z.; Tichaczek-Goska, D.; Wojnicz, D. Anti-planktonic and anti-biofilm properties of pentacyclic triterpenes-asiatic acid and ursolic acid as promising antibacterial future pharmaceuticals. Biomolecules 2022, 12, 98. [Google Scholar] [CrossRef]
- Tang, H.; Porras, G.; Brown, M.M.; Chassagne, F.; Lyles, J.T.; Bacsa, J.; Horswill, A.R.; Quav, C.L. Triterpenoid acids isolated from Schinus terebinthifolia fruits reduce Staphylococcus aureus virulence and abate dermonecrosis. Sci. Rep. 2020, 10, 8046. [Google Scholar] [CrossRef] [PubMed]
- Harley, B.K.; Neglo, D.; Tawiah, P.; Pipim, M.A.; Mireku-Gyimah, N.A.; Tettey, C.O.; Amengor, C.D.; Fleischer, T.C.; Waikhom, S.D. Bioactive triterpenoids from Solanum torvum fruits with antifungal, resistance modulatory and anti-biofilm formation activities against fluconazole-resistant candida albicans strains. PLoS ONE 2021, 16, e0260956. [Google Scholar] [CrossRef] [PubMed]
- Nagoor Meeran, M.F.; Goyal, S.N.; Suchal, K.; Sharma, C.; Patil, C.R.; Ojha, S.K. Pharmacological properties, molecular mechanisms, and pharmaceutical development of asiatic acid: A pentacyclic triterpenoid of therapeutic promise. Front. Pharm. 2018, 9, 892. [Google Scholar] [CrossRef] [PubMed]
- Wojnicz, D.; Tichaczek-Goska, D.; Kicia, M. Pentacyclic triterpenes combined with ciprofloxacin help to eradicate the biofilm formed in vitro by Escherichia coli. Indian J. Med. Res. 2015, 141, 343–353. [Google Scholar] [CrossRef] [Green Version]
- Lu, L.; Hu, W.; Tian, Z.; Yuan, D.; Yi, G.; Zhou, Y.; Cheng, Q.; Zhu, J.; Li, M. Developing natural products as potential anti-biofilm agents. Chin. Med. 2019, 14, 11. [Google Scholar] [CrossRef] [Green Version]
- Tamfu, A.N.; Ceylan, O.; Kucukaydin, S.; Duru, M.E. HPLC-DAD phenolic profiles, antibiofilm, anti-quorum sensing and enzyme inhibitory potentials of Camellia sinensis (L.) O. Kuntze and Curcuma longa L. LWT-Food Sci. Technol. 2020, 133, 110150. [Google Scholar] [CrossRef]
- Tamfu, A.N.; Ceylan, O.; Kucukaydin, S.; Ozturk, M.; Duru, M.E.; Dinica, R.M. Antibiofilm and enzyme inhibitory potentials of two Annonaceous food spices, African pepper (Xylopia aethiopica) and African nutmeg (Monodora myristica). Foods 2020, 9, 1768. [Google Scholar] [CrossRef]
- Alfred, T.N.; Ceylan, O.; Kucukaydin, S.; Olmez, O.T.; Godloves, C.F.; Sylvain, S.K.; Yeskaliyeva, B.; Duru, M.E.; Ozturk, M. HPLC-DAD and GC-MS characterization of Cameroonian honey samples and evaluation of their antibiofilm, anti-quorum sensing and antioxidant activities. Bull. Environ. Pharmacol. Life Sci. 2020, 9, 132–142. [Google Scholar]
- Alfred Ngenge, T.; Kucukaydin, S.; Ceylan, O.; Duru, M.E. Evaluation of enzyme inhibition and anti-quorum sensing potentials of Melaleuca alternifolia and Citrus sinensis essential oils. Nat. Prod. Com. 2021, 16, 1–8. [Google Scholar] [CrossRef]
- Tamfu, A.N.; Kucukaydin, S.; Quradha, M.M.; Ceylan, O.; Ugur, A.; Emin, D.M. Ultrasound-assisted extraction of Syringa vulgaris Mill., Citrus sinensis L. and Hypericum perforatum L.: Phenolic composition, enzyme inhibition and anti-quorum sensing activities. Chem. Afric. 2022, 5, 237–249. [Google Scholar] [CrossRef]
- Popova, M.; Gerginova, D.; Trusheva, B.; Simova, S.; Tamfu, A.N.; Ceylan, O.; Clark, K.; Bankova, V. A preliminary study of chemical profiles of honey, cerumen, and propolis of the African stingless bee Meliponula ferruginea. Foods 2021, 10, 997. [Google Scholar] [CrossRef]
- Mukherjee, S.; Bassler, B.L. Bacterial quorum sensing in complex and dynamically changing environments. Nat. Rev. Microbiol. 2019, 17, 371–382. [Google Scholar] [CrossRef]
- Boudiba, S.; Tamfu, A.N.; Berka, B.; Hanini, K.; Hioun, S.; Allaf, K.; Boudiba, L.; Ceylan, O. Anti-quorum sensing and antioxidant activity of essential oils extracted from Juniperus species, growing spontaneously in Tebessa Region (East of Algeria). Nat. Prod. Com. 2021, 16, 1–11. [Google Scholar] [CrossRef]
- Beddiar, H.; Boudiba, S.; Benahmed, M.; Tamfu, A.N.; Ceylan, Ö.; Hanini, K.; Kucukaydin, S.; Elomri, A.; Bensouici, C.; Laouer, H.; et al. Chemical composition, anti-quorum sensing, enzyme inhibitory, and antioxidant properties of phenolic extracts of Clinopodium nepeta L. Kuntze. Plants 2021, 10, 1955. [Google Scholar] [CrossRef]
- Savka, M.A.; Dailey, L.; Popova, M.; Mihaylova, R.; Merritt, B.; Masek, M.; Le, P.; Nor, S.R.; Ahmad, M.; Hudson, A.O.; et al. Chemical composition and disruption of quorum sensing signaling in geographically diverse United States propolis. Evid Based Complement. Alternat. Med. 2015, 15, 472593. [Google Scholar] [CrossRef]
- Lamberte, L.E.; Cabrera, E.C.; Rivera, W.L. Activity of the ethanolic extract of propolis (EEP) as a potential inhibitor of quorum sensing-mediated pigment production in Chromobacterium violaceum and virulence factor production in Pseudomonas aeruginosa. Philipp. Agric. Sci. 2011, 94, 14–22. [Google Scholar]
- Bulman, Z.; Hudson, A.; Savka, M.A. Propolis, a product of the bee hive, has an antagonistic affect on quorum-sensing regulated bioluminescence, gene transcription and motility. FASEB J. 2011, 25, 948. [Google Scholar] [CrossRef]
- Song, X.; Xia, Y.-X.; He, Z.-D.; Zhang, H.J. A review of natural products with anti-biofilm activity. Curr. Org. Chem. 2018, 22, 789–817. [Google Scholar] [CrossRef]
- Sorucu, A.; Ceylan, O. Determination of antimicrobial and anti-quorum sensing activities of water and ethanol extracts of propolis. Ankara Univ. Vet Fak. Derg. 2021, 68, 373–381. [Google Scholar] [CrossRef]
- Kocak, G.; Tamfu, A.N.; Bütün, V.; Ceylan, O. Synthesis of quaternary piperazine methacrylate homopolymers and their antibiofilm and anti-quorum sensing effects on pathogenic bacteria. J. Appl. Polym. Sci. 2021, 138, e50466. [Google Scholar] [CrossRef]
- Ceylan, O.; Tamfu, A.N.; Doğaç, Y.İ.; Teke, M. Antibiofilm and anti-quorum sensing activities of polyethylene imine coated magnetite and nickel ferrite nanoparticles. 3 Biotech 2020, 10, 513. [Google Scholar] [CrossRef]
- Cayan, F.; Deveci, E.; Tel-Cayan, G.; Duru, M.E. Identification and quantification of phenolic acid compounds of twenty-six mushrooms by HPLC–DAD. J. Food Meas. Charact. 2020, 14, 1690–1698. [Google Scholar] [CrossRef]
- Tamfu, A.N.; Kucukaydin, S.; Ceylan, O.; Sarac, N.; Duru, E.M. Phenolic composition, enzyme inhibitory and anti-quorum sensing activities of cinnamon (Cinnamomum zeylanicum Blume) and Basil (Ocimum basilicum Linn). Chem. Afric. 2021, 4, 759–767. [Google Scholar] [CrossRef]
- Tel-Cayan, G.; Duru, M.E. Chemical characterization and antioxidant activity of Eryngium pseudothoriifolium and E. thorifolium essential oils. J. Res. Pharm. 2019, 23, 1106–1114. [Google Scholar] [CrossRef] [Green Version]
- Apak, R.; Guclu, K.; Ozyurek, M.; Karademir, S.E. Novel total antioxidant capacity index for dietary polyphenols and vitamins C and E, using their cupric ion reducing capability in the presence of neocuproine: CUPRAC method. J. Agric. Food Chem. 2004, 52, 7970–7981. [Google Scholar] [CrossRef]
- Decker, E.A.; Welch, B. Role of ferritin as a lipid oxidation catalyst in muscle food. J. Agric. Food Chem. 1990, 38, 674–677. [Google Scholar] [CrossRef]
- Ana, P.L.; Wallans, T.P.S.; Edson, N.; Eduardo, M.R.; Rodrigo, A.A.M. Critical evaluation of voltammetric techniques for antioxidant capacity and activity: Presence of alumina on glassy-carbon electrodes alters the results. Electrochim. Acta 2020, 358, 136925. [Google Scholar] [CrossRef]
- CLSI (Clinical Laboratory Standards Institute). Quality Control Minimal Inhibitory Concentration (MIC) Limits for Broth Dilution and MIC Interpretative Breakpoints (M27-s2); CLSI: Wayne, PA, USA, 2006. [Google Scholar]
- Merritt, J.H.; Kadouri, D.E.; O’Toole, G.A. Growing and analyzing static biofilms. In Current Protocols in Microbiology; John Wiley & Sons, Inc.: Hoboken, NJ, USA, 2005. [Google Scholar] [CrossRef] [Green Version]
- Koh, K.M.; Tham, F.Y. Screening of traditional Chinese medicinal plants for quorum-sensing inhibitors activity. J. Microbiol. Immunol. Infect. 2011, 44, 144–148. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Packiavathy, I.A.S.V.; Agilandeswari, P.; Musthafa, K.S.; Pandian, S.K.; Ravi, A.V. Antibiofilm and quorum sensing inhibitory potential of Cuminum cyminum and its secondary metabolite methyl eugenol against Gram negative bacterial pathogens. Food Res. Int. 2012, 45, 85–92. [Google Scholar] [CrossRef]
- Kasote, D.; Bankova, V.; Viljoen, A.M. Propolis: Chemical diversity and challenges in quality control. Phytochem. Rev. 2022, 24, 1–25. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
| Compound Name | Linear Range (μg/mL) | LOQ (μg/mL) | µg/g Extract |
|---|---|---|---|
| Quercetin | 6.25–400 | 10.35 | 43.53 ± 1.10 |
| Protocatechic acid | 1.56–100 | 12.07 | 40.76 ± 0.82 |
| Vanillic acid | 12.5–100 | 4.79 | 29.90 ± 1.05 |
| 4-Hydroxybenzoic acid | 1.56–100 | 4.68 | 24.04 ± 0.21 |
| Luteolin | 3.00–30.0 | 3.99 | 4.44 ± 0.48 |
| 6,7-dihydroxycoumarin | 3.13–200 | 7.56 | TR |
| Catechin | 30.0–120 | 19.33 | TR |
| p-Coumaric acid | 1.56–100 | 3.06 | TR |
| transCinnamic acid | 1.56–100 | 7.56 | TR |
| Vanillin | 12.5–100 | 8.13 | TR |
| Position | 27-Hydroxymangiferonic Acid (1) | Ambolic Acid (2) | Mangiferonic Acid (3) | Lupeol (4) | β-Amyrin (5) | |||||
|---|---|---|---|---|---|---|---|---|---|---|
| 13C | 1H | 13C | 1H | 13C | 1H | 13C | 1H | 13C | 1H | |
| 1 | 33.6 | 1.85, 1.56 | 32.0 | 1.53, 1.22 | 33.4 | 1.85, 1.56 | 38.8 | / | 38.7 | 1.91 |
| 2 | 38.2 | 2.29, 2.75 | 30.4 | 1.76, 1.55 | 37.6 | 2.31, 2.71 | 27.5 | / | 23.6 | / |
| 3 | 218 | / | 77.9 | 3.30 | 214.8 | / | 79.0 | 3.19 | 79.01 | 3.24 |
| 4 | 55.0 | / | 40.5 | / | 50.3 | / | 39.9 | / | 37.2 | / |
| 5 | 42.8 | 2.15 | 47.1 | 1.29 | 48.4 | 1.69 | 55.3 | / | 55.3 | / |
| 6 | 21.2 | 1.56, 0.97 | 20.8 | 1.58, 0.79 | 21.5 | 1.53 | 19.3 | / | 18.0 | / |
| 7 | 28.2 | 1.90, 1.35 | 26.2 | 1.32, 1.08 | 25.7 | 1.34 | 34.2 | / | 32.8 | 1.57 |
| 8 | 47.8 | 1.61 | 47.9 | 1.51 | 47.9 | 1.50 | 41.1 | / | 41.5 | / |
| 9 | 21.0 | / | 19.7 | / | 21.1 | / | 50.5 | / | 47.6 | / |
| 10 | 25.7 | / | 26.0 | / | 25.9 | / | 37.2 | / | 36.8 | / |
| 11 | 26.7 | 2.25, 1.20 | 26.4 | 1.99, 1.12 | 26.7 | / | 21.2 | 1.40 | 28.1 | 1.94 |
| 12 | 32.8 | 1.65 | 32.8 | 1.60 | 32.8 | 1.64 | 25.3 | 1.88 | 121.8 | 5.18 |
| 13 | 45.4 | / | 45.3 | / | 45.4 | / | 38.5 | / | 145.3 | / |
| 14 | 48.8 | / | 48.6 | / | 48.8 | / | 42.8 | / | 42.1 | / |
| 15 | 35.5 | 1.29 | 35.5 | 1.30 | 35.5 | 1.35 | 27.2 | / | 26.6 | / |
| 16 | 25.5 | 1.35 | 27.9 | 1.86 | 28.2 | - | 35.6 | / | 31.1 | / |
| 17 | 52.2 | 1.61 | 52.5 | 1.61 | 52.2 | 1.61 | 43.0 | / | 40.8 | / |
| 18 | 18.2 | 0.99 | 18.0 | 0.98 | 18.1 | 1.08 | 48.3 | / | 50.5 | 1.94 |
| 19 | 29.7 | 0.61, 0.82 | 29.8 | 0.56, 0.34 | 29.6 | 0.58, 0.78 | 47.8 | 2.38 | 28.1 | / |
| 20 | 36.0 | 1.45 | 36.1 | 1.42 | 36.0 | / | 150.9 | / | 33.7 | / |
| 21 | 18.1 | 0.93 | 18.3 | 0.89 | 18.1 | 0.91 | 30.1 | / | 39.6 | / |
| 22 | 35.2 | 1.60 | 35.0 | 1.61 | 29.6 | / | 40.3 | / | 39.7 | / |
| 23 | 25.7 | 2.19 | 25.8 | 2.12, 1.93 | 34.8 | / | 28.4 | 1.04 | 28.1 | 0.80 |
| 24 | 130.0 | 7.02 | 148.9 | / | 145.8 | 6.9 | 15.6 | 0.97 | 16.7 | 0.91 |
| 25 | 149.2 | / | 45.6 | 3.18 | 126.6 | / | 16.1 | 1.40 | 15.6 | 0.77 |
| 26 | 171.2 | / | 178.9 | / | 173.0 | / | 16.0 | 0.84 | 16.8 | 0.94 |
| 27 | 57 | 4.37 | 16.3 | 1.31 | 11.9 | 1.85 | 14.5 | 0.79 | 23.2 | 1.15 |
| 28 | 22.2 | / | 24.9 | 0.97 | 22.2 | 1.04 | 18.1 | 1.26 | 17.5 | 1.08 |
| 29 | 20.8 | 1.08 | 14.1 | 0.81 | 20.8 | 1.10 | 109.3 | 4.56, 4.65 | 18.7 | 0.84 |
| 30 | 19.3 | 0.91 | 19.3 | 0.90 | 19.3 | 1.04 | 20.2 | 1.69 | 21.3 | 0.81 |
| 31 | / | / | 110.2 | 4.98, 4.92 | / | / | / | / | / | / |
| Test Sample | DPPH• Assay | ABTS•+ Assay | CUPRAC Assay | Metal Chelating Assay |
|---|---|---|---|---|
| IC50 (µg /mL) | IC50 (µg /mL) | A0.5 (µg/mL) | %Inh. (100 µg/mL) | |
| Extract | 14.90 ± 1.10 | 10.86 ± 0.91 | 6.50 ± 0.25 | 46.21 ± 0.71 |
| Standards | ||||
| α-Tocopherol | 12.26 ± 0.07 | 4.31 ± 0.10 | 10.20 ± 0.01 | NT |
| BHT | 25.37 ± 0.47 | 4.10 ± 0.06 | 3.80 ± 0.00 | NT |
| Quercetin | 2.07 ± 0.10 | 1.18 ± 0.03 | NT | 35.91 ± 0.82 |
| EDTA | NT | NT | NT | 85.40 ± 0.10 |
| Sample | MIC (mg/mL) | Violacein İnhibition (%) | |||||
|---|---|---|---|---|---|---|---|
| MIC | MIC/2 | MIC/4 | MIC/8 | MIC/16 | MIC/32 | ||
| Propolis extract | 0.5 | 100 ± 0.00 | 100 ± 0.00 | 85.26 ± 0.50 | 20.50 ± 0.10 | - | - |
| 27-Hydroxymangiferonic acid | 1.0 | 100 ± 0.00 | 100 ± 0.00 | 100 ± 0.00 | 60.49 ± 0.25 | 35.89 ± 0.64 | - |
| Ambolic acid | 1.0 | 100 ± 0.00 | 100 ± 0.00 | 100 ± 0.00 | 45.22 ± 1.00 | 24.90 ± 1.00 | - |
| Mangiferonic acid | 0.5 | 100 ± 0.00 | 100 ± 0.00 | 82.60 ± 0.21 | 35.77 ± 0.31 | - | - |
| Sample | Anti-Quorum Sensing İnhibition Zones (mm) | ||||
|---|---|---|---|---|---|
| MIC (mg/mL) | MIC | MIC/2 | MIC/4 | MIC/8 | |
| Propolis extract | 0.25 | 18.0 ± 1.0 | 14.5 ± 0.5 | - | - |
| 27-Hydroxymangiferonic acid | 0.5 | 12.0 ± 0.5 | - | - | - |
| Ambolic acid | 1.0 | 9.0 ± 1.0 | - | - | - |
| Mangiferonic acid | 0.5 | 12.3 ± 1.0 | - | - | - |
| Sample | Swarming İnhibition (%) | ||
|---|---|---|---|
| 100 µg/mL | 75 µg/mL | 50 µg/mL | |
| Propolis extract | 47.95 ± 1.11 | 25.65 ± 0.20 | - |
| 27-Hydroxymangiferonic acid | 45.50 ± 1.20 | 16.62 ± 0.25 | - |
| Ambolic acid | 58.72 ± 1.05 | 31.80 ± 0.91 | 06.68 ± 0.70 |
| Mangiferonic acid | 40.78 ± 0.50 | 29.64 ± 0.25 | 08.47 ± 0.49 |
| Sample | S. aureus ATCC 25923 | L. monocytogenes ATCC 7644 | E. faecalis ATCC 29212 | E. coli ATCC 25922 | P. aeruginosa ATCC 27853 | S. typhi ATCC 14028 | C. albicans ATCC 10239 | C. tropicalis ATCC 13803 |
|---|---|---|---|---|---|---|---|---|
| PR | 0.25 | >1 | 1 | 0.5 | 1 | 0.5 | 0.25 | 0.5 |
| 1 | 0.5 | 1 | 1 | >1 | >1 | >1 | 0.5 | 1 |
| 2 | 0.5 | >1 | 0.5 | >1 | >1 | 1 | 1 | 1 |
| 3 | 0.25 | 0.5 | 0.5 | 1 | >1 | 1 | 0.5 | 0.25 |
| PR | 1 | 2 | 3 | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| MIC | MIC/2 | MIC | MIC/2 | MIC/4 | MIC/8 | MIC | MIC/2 | MIC/4 | MIC/8 | MIC | MIC/2 | MIC/4 | MIC/8 | |
| Biofilm İnhibition (% İnhibition) | ||||||||||||||
| S. aureus | 18.3 ± 0.8 | 5.1± 0.2 | 40.5 ± 0.9 | 25.5 ± 0.2 | 8.8 ± 0.3 | - | 24.2 ± 0.3 | 5.5 ± 0.1 | - | - | 8.6 ± 0.2 | - | - | - |
| L. monocytogenes | - | - | 17.4 ± 0.1 | 5.5 ± 0.5 | - | - | - | - | - | - | 15.7 ± 0.2 | - | - | - |
| E. faecalis | - | - | 18.7 ± 0.4 | 5.4 ± 0.1 | - | - | 22.9 ± 1.0 | - | - | - | 32.4 ± 0.9 | 12.6 ± 0.2 | - | - |
| E. coli | 38.5 ± 1.2 | 20.5 ± 0.3 | 38.5 ± 1.1 | 25.8 ± 0.8 | 15.6 ± 0.6 | 8.3 ± 0.4 | 32.5 ± 0.5 | 21.8 ± 0.7 | 11.9 ± 0.2 | 3.3 ± 0.1 | 44.5 ± 1.0 | 28.9 ± 0.8 | 12.1 ± 0.2 | 5.2 ± 0.1 |
| P. aeruginosa | 20.6 ± 0.5 | - | - | - | - | - | - | - | - | - | - | - | - | - |
| S. typhi | - | - | - | - | - | - | 17.8 ± 0.3 | - | - | - | 14.2 ± 0.8 | - | - | - |
| C. albicans | 8.4 ± 0.3 | - | 13.5 ± 0.1 | - | - | - | 17.5 ± 0.6 | - | - | - | 44.5 ± 0.4 | 26.1 ± 0.5 | - | - |
| C. tropicalis | 10.9 ± 1.5 | - | 40.1 ± 1.0 | 21.8 ± 0.4 | 7.9 ± 0.1 | - | 39.8 ± 1.0 | 26.0 ± 0.9 | 10.2 ± 0.4 | - | 22.3 ± 0.8 | 5.8 ± 0.1 | - | - |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tamfu, A.N.; Ceylan, O.; Cârâc, G.; Talla, E.; Dinica, R.M. Antibiofilm and Anti-Quorum Sensing Potential of Cycloartane-Type Triterpene Acids from Cameroonian Grassland Propolis: Phenolic Profile and Antioxidant Activity of Crude Extract. Molecules 2022, 27, 4872. https://doi.org/10.3390/molecules27154872
Tamfu AN, Ceylan O, Cârâc G, Talla E, Dinica RM. Antibiofilm and Anti-Quorum Sensing Potential of Cycloartane-Type Triterpene Acids from Cameroonian Grassland Propolis: Phenolic Profile and Antioxidant Activity of Crude Extract. Molecules. 2022; 27(15):4872. https://doi.org/10.3390/molecules27154872
Chicago/Turabian StyleTamfu, Alfred Ngenge, Ozgur Ceylan, Geta Cârâc, Emmanuel Talla, and Rodica Mihaela Dinica. 2022. "Antibiofilm and Anti-Quorum Sensing Potential of Cycloartane-Type Triterpene Acids from Cameroonian Grassland Propolis: Phenolic Profile and Antioxidant Activity of Crude Extract" Molecules 27, no. 15: 4872. https://doi.org/10.3390/molecules27154872
APA StyleTamfu, A. N., Ceylan, O., Cârâc, G., Talla, E., & Dinica, R. M. (2022). Antibiofilm and Anti-Quorum Sensing Potential of Cycloartane-Type Triterpene Acids from Cameroonian Grassland Propolis: Phenolic Profile and Antioxidant Activity of Crude Extract. Molecules, 27(15), 4872. https://doi.org/10.3390/molecules27154872