
Structural Determinants of the Specific Activities of an L-Amino Acid Oxidase from Pseudoalteromonas luteoviolacea CPMOR-1 with Broad Substrate Specificity

 , , , , and
, , , , and
Abstract
1. Introduction
2. Results
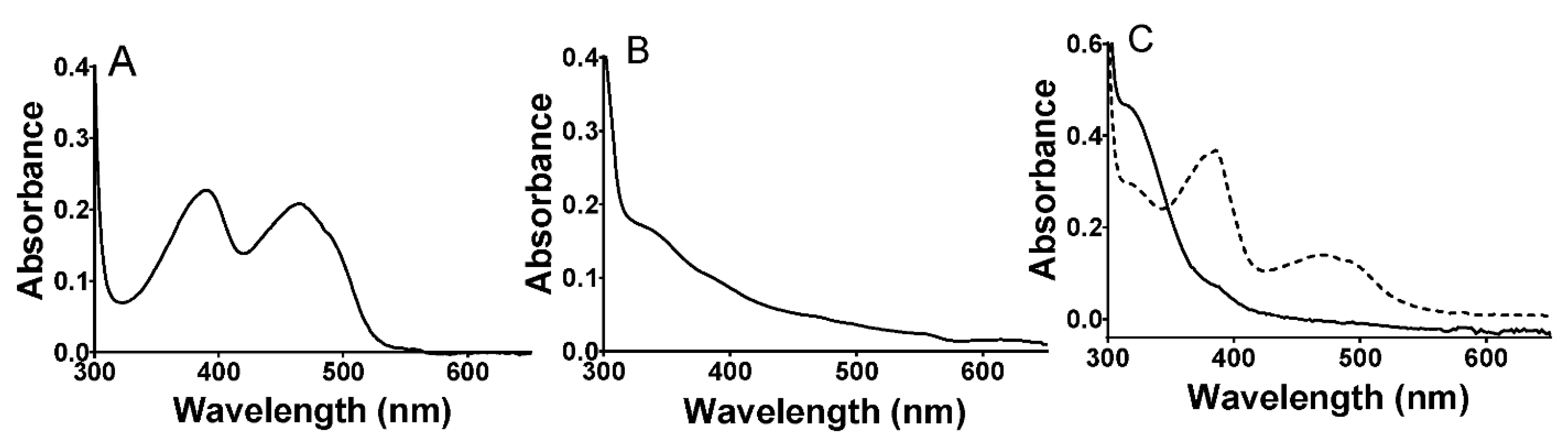
2.1. Properties of P. luteoviolacea CPMOR-1 LAAO
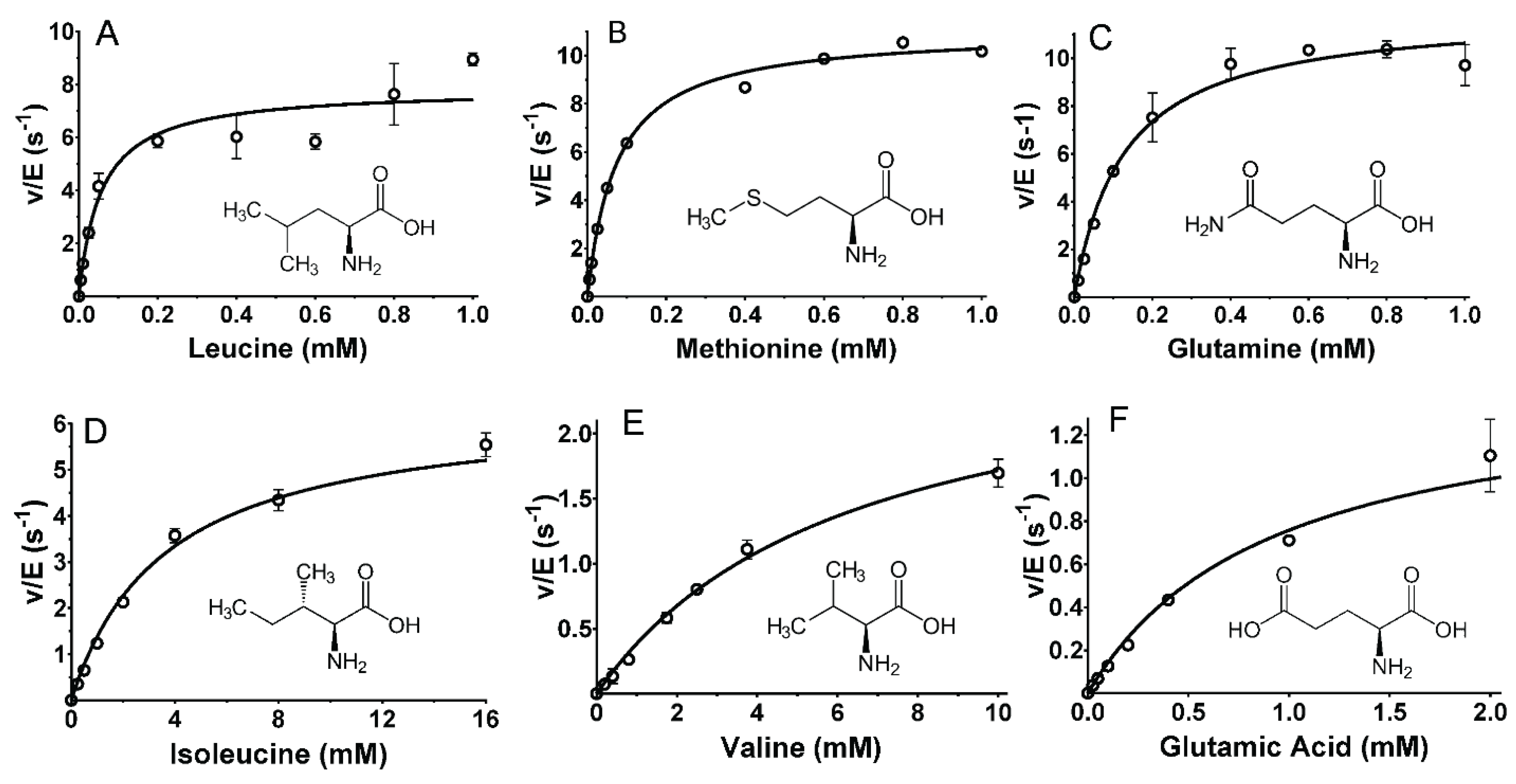
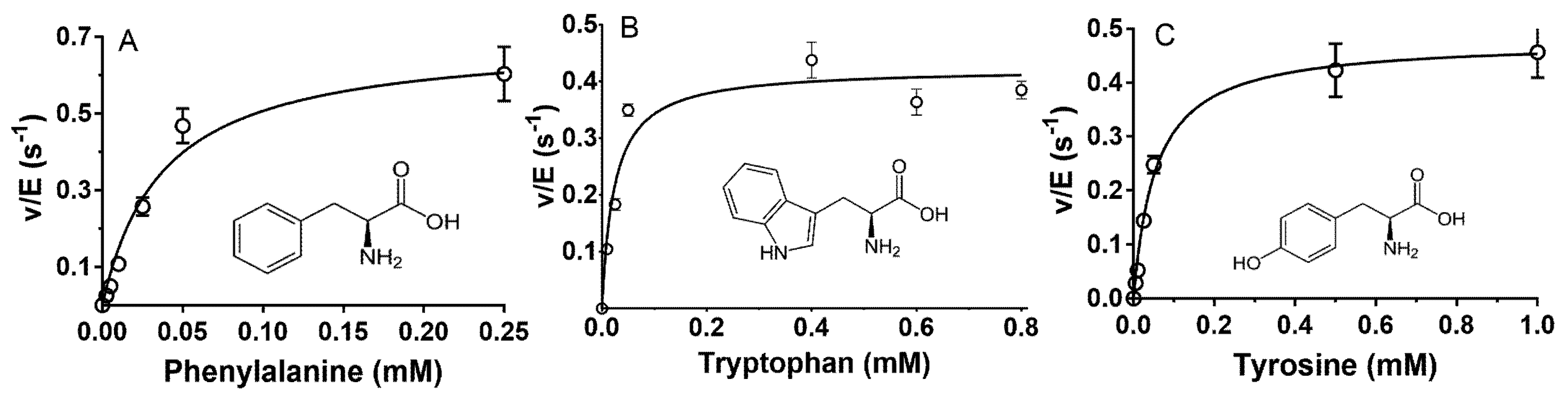
2.2. Steady-State Kinetic Analysis of Reactions of P. luteoviolacea CPMOR-1 LAAO with Amino Acids
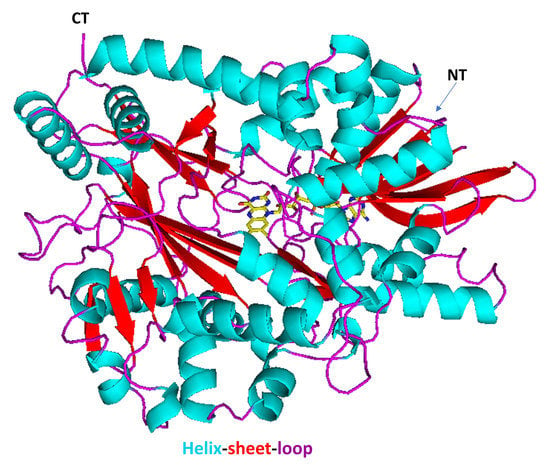
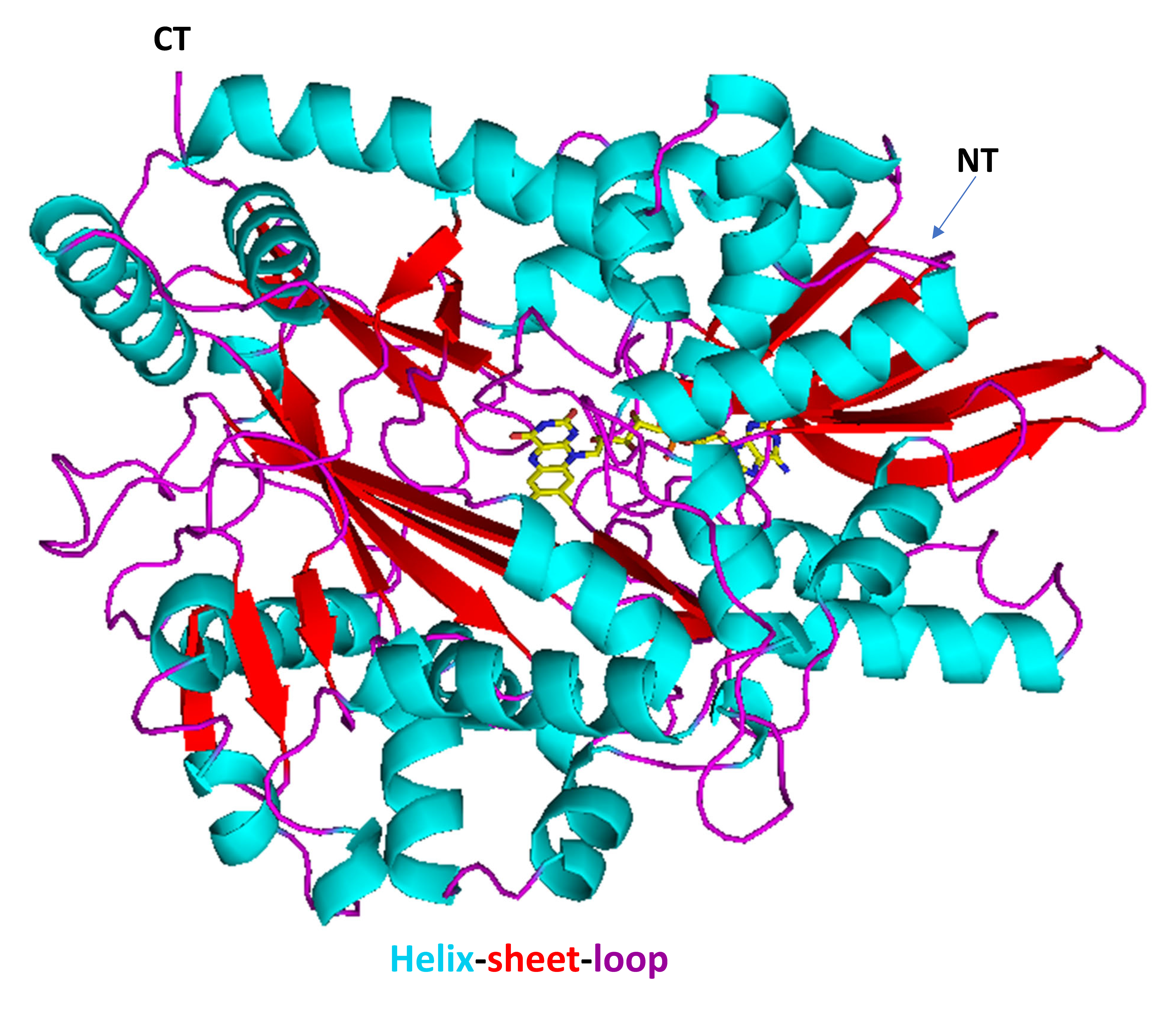
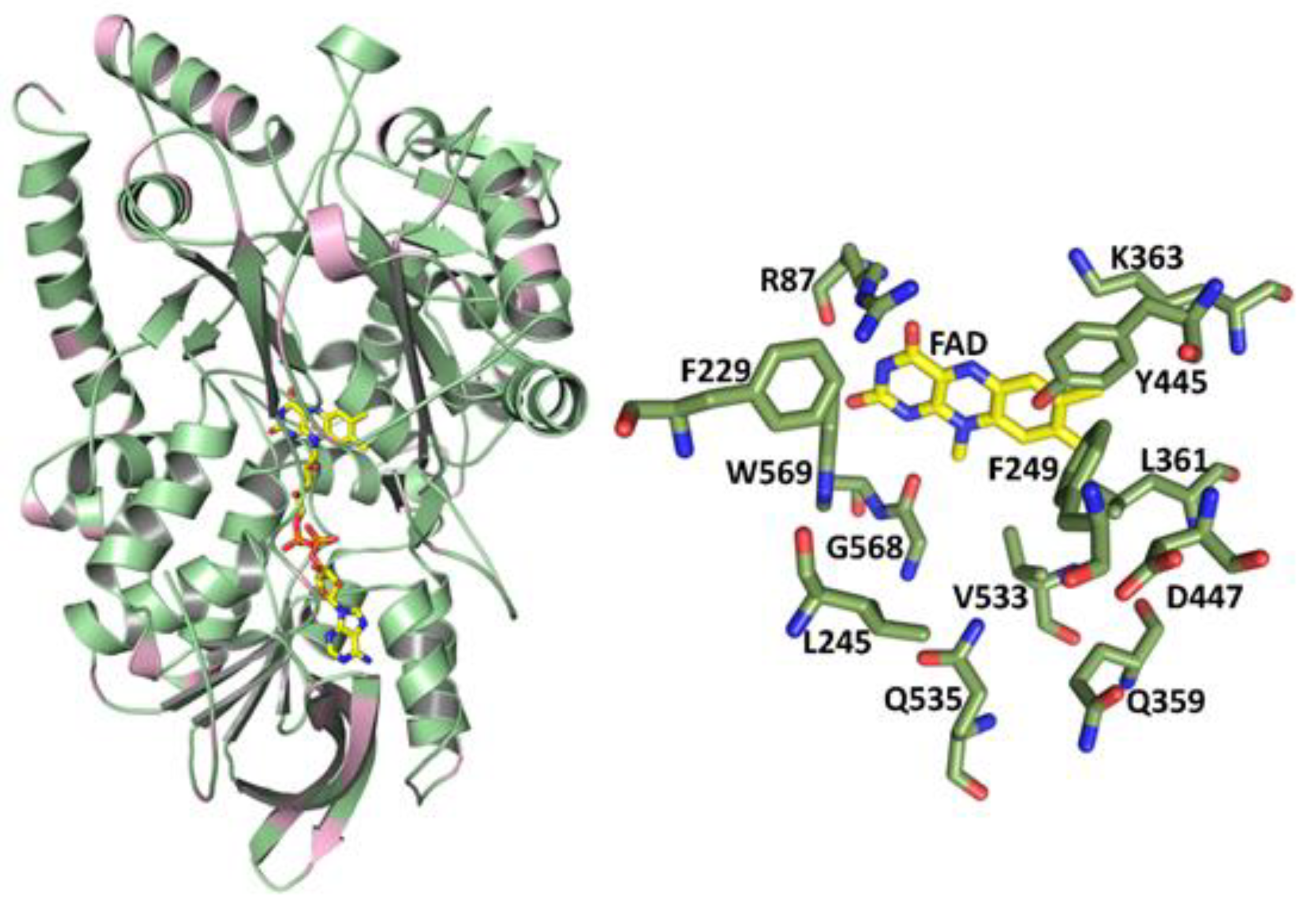
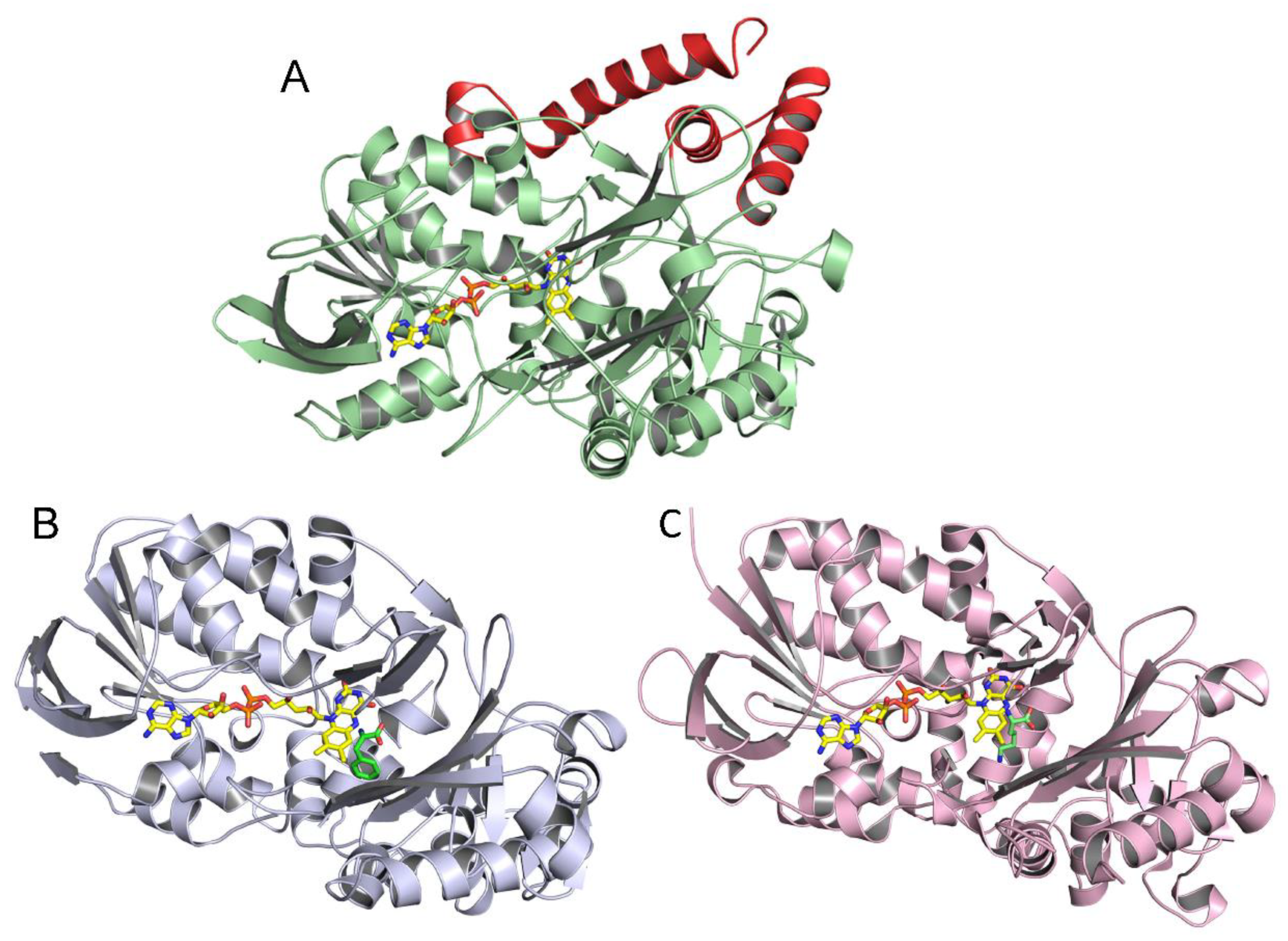
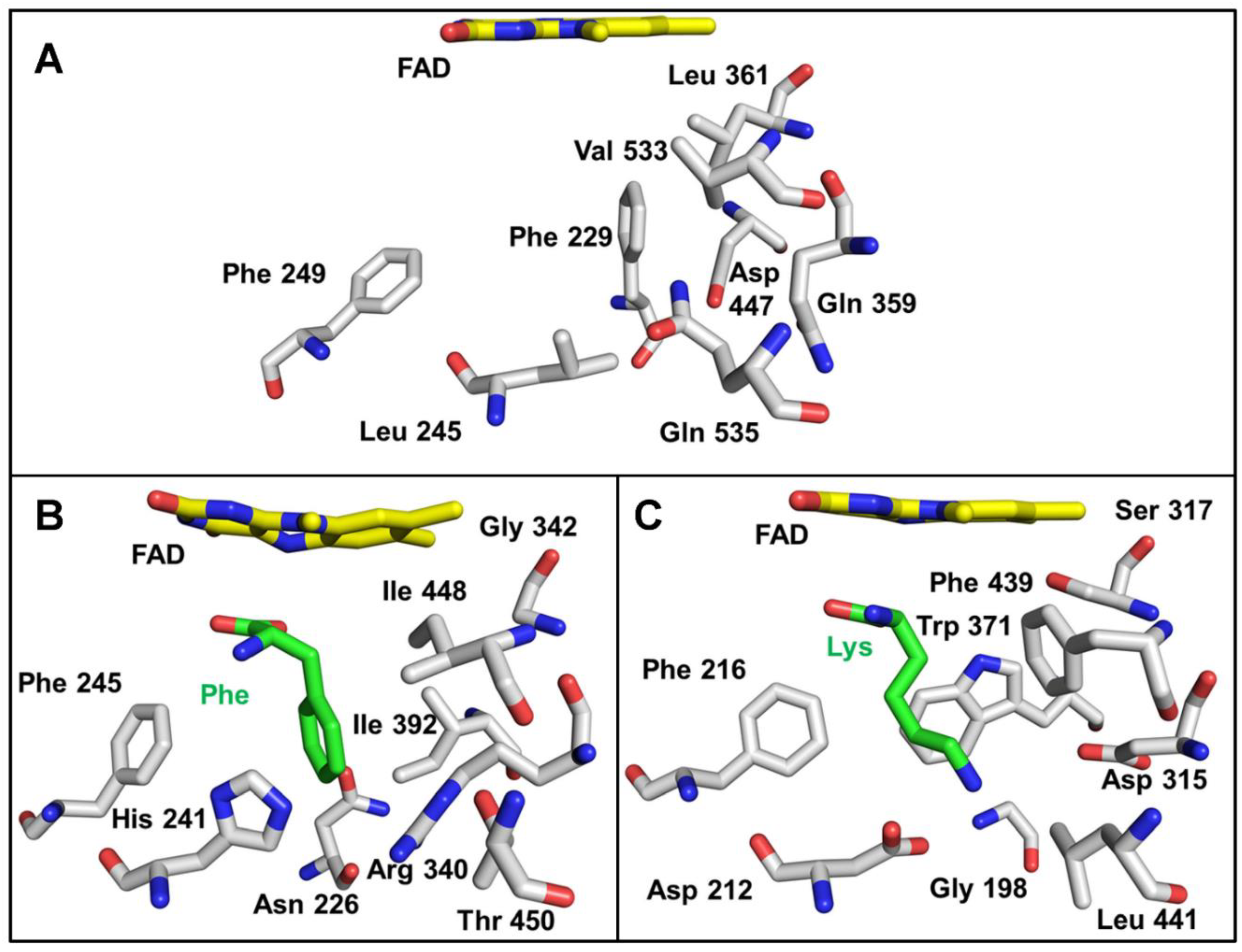
2.3. Correlation of LAAO Sequences and Structures with Substrate Specificity
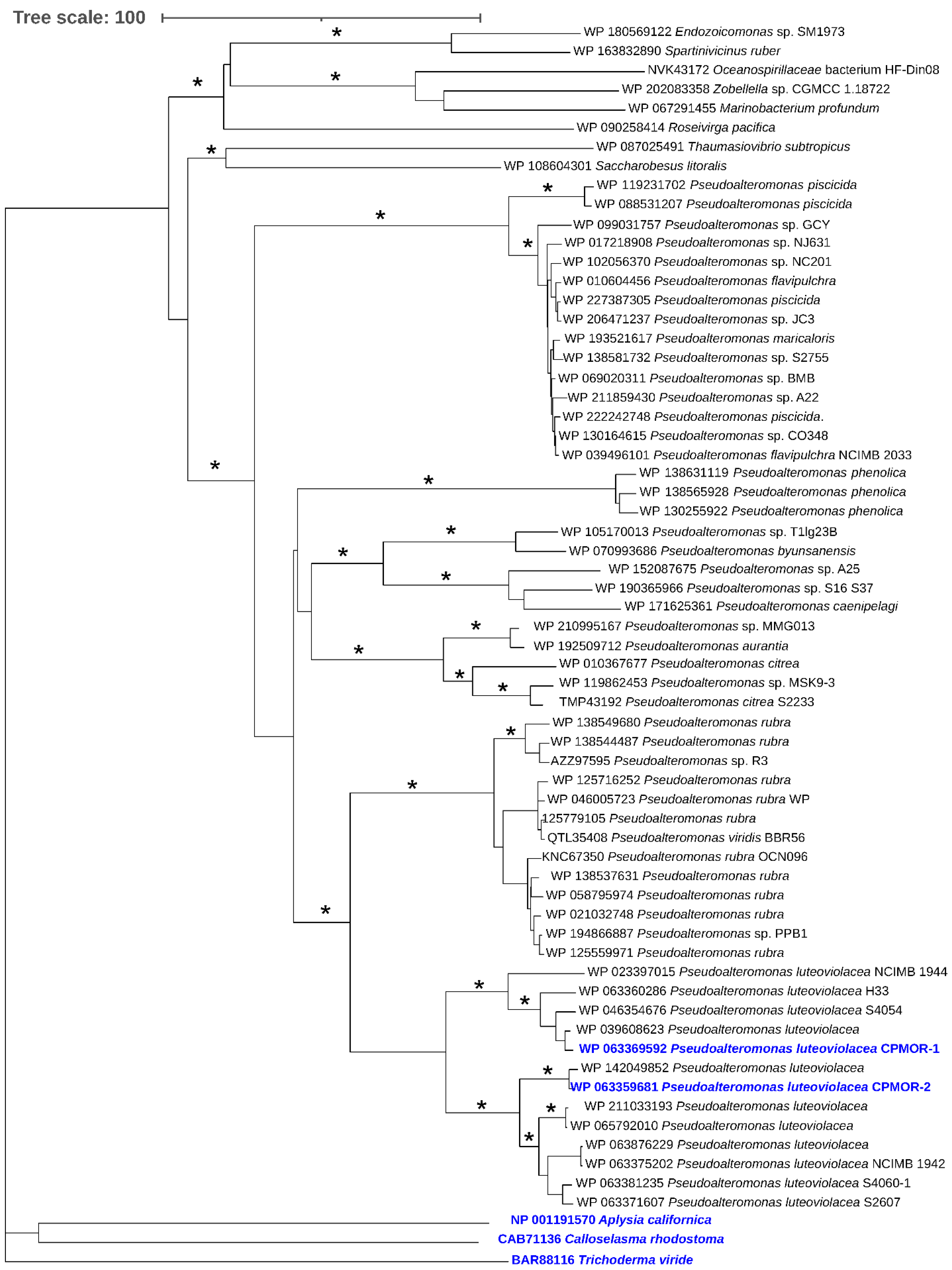
2.4. Phylogenetic Analysis of LAAOs
3. Discussion
3.1. Biological Roles of LAAOs
3.2. Structure–Function Correlations
3.3. Potential Applications of the P. luteoviolacea CPMOR-1 LAAO
4. Materials and Methods
4.1. Expression and Purification of P. luteoviolacea CPMOR-1 LAAO
4.2. Steady-State Kinetics
4.3. Phylogenetic Analysis of LAAOs
4.4. Structure Modeling and Sequence Alignment
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Sample Availability
References
- Pollegioni, L.; Motta, P.; Molla, G. L-Amino acid oxidase as biocatalyst: A dream too far? Appl. Environ. Microbiol. 2013, 97, 9323–9341. [Google Scholar] [CrossRef] [PubMed]
- Campillo-Brocal, J.C.; Chacón-Verdú, M.D.; Lucas-Elío, P.; Sánchez-Amat, A. Distribution in microbial genomes of genes similar to lodA and goxA which encode a novel family of quinoproteins with amino acid oxidase activity. BMC Genom. 2015, 16, 231. [Google Scholar] [CrossRef] [PubMed]
- Neubauer, O.; Gross, W. Zur Kenntnis des Tyrosinabbaus in der künstlich durchbluteten Leber. Zeitschrift Für Phys. Chem. 1910, 67, 219–229. [Google Scholar] [CrossRef][Green Version]
- Blanchard, M.; Green, D.E.; Nocito, V.; Ratner, S. L-Amino acid oxidase of animal tissue. J. Biol. Chem. 1944, 155, 421–440. [Google Scholar] [CrossRef]
- Zeller, E.A.; Maritz, A. Ueber eine neue l-Aminosaeure-Oxidase. Hel. Chim. Acta 1944, 27, 1888–1902. [Google Scholar] [CrossRef]
- Du, X.-Y.; Clemetson, K.J. Snake venom l-amino acid oxidases. Toxicon 2002, 40, 659–665. [Google Scholar] [CrossRef]
- Ali, S.A.; Stoeva, S.; Abbasi, A.; Alam, J.; Kayed, R.; Faigle, M.; Neumeister, B.; Voelter, W. Isolation, structural, and functional characterization of an apoptosis-Inducing -amino acid oxidase from Leaf-Nosed Viper (Eristocophis macmahoni) snake venom. Arch. Biochem. Biophys. 2001, 384, 216–226. [Google Scholar] [CrossRef] [PubMed]
- Ehara, T.; Kitajima, S.; Kanzawa, N.; Tamiya, T.; Tsuchiya, T. Antimicrobial action of achacin is mediated by L-amino acid oxidase activity. FEBS Lett. 2002, 531, 509–512. [Google Scholar] [CrossRef]
- Jimbo, M.; Nakanishi, F.; Sakai, R.; Muramoto, K.; Kamiya, H. Characterization of L-amino acid oxidase and antimicrobial activity of aplysianin A, a sea hare-derived antitumor-antimicrobial protein. Fish. Sci. 2003, 69, 1240–1246. [Google Scholar] [CrossRef]
- Yang, H.; Johnson, P.M.; Ko, K.-C.; Kamio, M.; Germann, M.W.; Derby, C.D.; Tai, P.C. Cloning, characterization and expression of escapin, a broadly antimicrobial FAD-containing l-amino acid oxidase from ink of the sea hare Aplysia californica. J. Exp. Biol. 2005, 208, 3609–3622. [Google Scholar] [CrossRef]
- Yu, Z.; Wang, J.; Lin, J.; Zhao, M.; Qiu, J. Exploring regulation genes involved in the expression of L-amino acid oxidase in Pseudoalteromonas sp. Rf-1. PLoS ONE 2015, 10, e0122741. [Google Scholar] [CrossRef] [PubMed]
- Mai-Prochnow, A.; Lucas-Elio, P.; Egan, S.; Thomas, T.; Webb, J.S.; Sanchez-Amat, A.; Kjelleberg, S. Hydrogen peroxide linked to lysine oxidase activity facilitates biofilm differentiation and dispersal in several Gram-negative bacteria. J. Bacteriol. 2008, 190, 5493–5501. [Google Scholar] [CrossRef]
- Sikora, L.; Marzluf, G.A. Regulation of L-amino acid oxidase and of D-amino acid oxidase in Neurospora crassa. Mol. Gen. Genet. 1982, 186, 33–39. [Google Scholar] [CrossRef] [PubMed]
- Puiffe, M.-L.; Lachaise, I.; Molinier-Frenkel, V.; Castellano, F. Antibacterial properties of the mammalian L-amino acid oxidase IL4I1. PLoS ONE 2013, 8, e54589. [Google Scholar] [CrossRef]
- Brearley, G.M.; Price, C.P.; Atkinson, T.; Hammond, P.M. Purification and partial characterisation of a broad-range L-amino acid oxidase from Bacillus carotarum 2Pfa isolated from soil. Appl. Environ. Microbiol. 1994, 41, 670–676. [Google Scholar] [CrossRef]
- Pilone, M.S. D-Amino acid oxidase: New findings. Cell. Mol. Life Sci. 2000, 57, 1732–1747. [Google Scholar] [CrossRef] [PubMed]
- Pollegioni, L.; Sacchi, S.; Murtas, G. Human D-amino acid oxidase: Structure, function, and regulation. Front. Mol. Biosci. 2018, 5, 107. [Google Scholar] [CrossRef]
- Campillo-Brocal, J.C.; Lucas-Elio, P.; Sanchez-Amat, A. Distribution in different organisms of amino acid oxidases with FAD or a quinone As cofactor and their role as antimicrobial proteins in marine bacteria. Mar. Drugs 2015, 13, 7403–7418. [Google Scholar] [CrossRef]
- Faust, A.; Niefind, K.; Hummel, W.; Schomburg, D. The structure of a bacterial L-amino acid oxidase from Rhodococcus opacus gives new evidence for the hydride mechanism for dehydrogenation. J. Mol. Biol. 2007, 367, 234–248. [Google Scholar] [CrossRef]
- Geueke, B.; Hummel, W. A new bacterial L-amino acid oxidase with a broad substrate specificity: Purification and characterization. Enzym. Microb. Technol. 2002, 31, 77–87. [Google Scholar] [CrossRef]
- Datta, S.; Mori, Y.; Takagi, K.; Kawaguchi, K.; Chen, Z.W.; Okajima, T.; Kuroda, S.; Ikeda, T.; Kano, K.; Tanizawa, K.; et al. Structure of a quinohemoprotein amine dehydrogenase with an uncommon redox cofactor and highly unusual crosslinking. Proc. Natl. Acad. Sci. USA 2001, 98, 14268–14273. [Google Scholar] [CrossRef] [PubMed]
- Gómez, D.; Lucas-Elío, P.; Sanchez-Amat, A.; Solano, F. A novel type of lysine oxidase: L-lysine-ε-oxidase. Biochim. Biophys. Acta 2006, 1764, 1577–1585. [Google Scholar] [CrossRef] [PubMed]
- Campillo-Brocal, J.C.; Lucas-Elio, P.; Sanchez-Amat, A. Identification in Marinomonas mediterranea of a novel quinoprotein with glycine oxidase activity. Microbiol. Open 2013, 2, 684–694. [Google Scholar] [CrossRef] [PubMed]
- Andreo-Vidal, A.; Mamounis, K.J.; Sehanobish, E.; Avalos, D.; Campillo-Brocal, J.C.; Sanchez-Amat, A.; Yukl, E.T.; Davidson, V.L. Structure and enzymatic properties of an unusual cysteine tryptophylquinone-dependentglycine oxidase from Pseudoalteromonas Luteoviolacea. Biochemistry 2018, 57, 1155–1165. [Google Scholar] [CrossRef]
- Gomez, D.; Espinosa, E.; Bertazzo, M.; Lucas-Elio, P.; Solano, F.; Sanchez-Amat, A. The macromolecule with antimicrobial activity synthesized by Pseudoalteromonas luteoviolacea strains is an L-amino acid oxidase. Appl. Environ. Microbiol. 2008, 79, 925–930. [Google Scholar] [CrossRef]
- Andreo-Vidal, A.; Sanchez-Amat, A.; Campillo-Brocal, J.C. The Pseudoalteromonas luteoviolacea L-amino acid oxidase with antimicrobial activity Is a flavoenzyme. Mar. Drugs 2018, 16, 499. [Google Scholar] [CrossRef]
- Savino, S.; Meijer, J.D.; Rozeboom, H.J.; van Beek, H.L.; Fraaije, M.W. Kinetic and structural properties of a robust bacterial L-amino acid oxidase. Catalysts 2021, 11, 1309. [Google Scholar] [CrossRef]
- Moustafa, I.M.; Foster, S.; Lyubimov, A.Y.; Vrielink, A. Crystal Structure of LAAO from Calloselasma rhodostoma with an l-phenylalanine substrate: Insights into structure and mechanism. J. Mol. Biol. 2006, 364, 991–1002. [Google Scholar] [CrossRef]
- Kondo, H.; Kitagawa, M.; Matsumoto, Y.; Saito, M.; Amano, M.; Sugiyama, S.; Tamura, T.; Kusakabe, H.; Inagaki, K.; Imada, K. Structural basis of strict substrate recognition of l-lysine α-oxidase from Trichoderma viride. Protein Sci. 2020, 29, 2213–2225. [Google Scholar] [CrossRef]
- Kabir, M.P.; Orozco-Gonzalez, Y.; Gozem, S. Electronic spectra of flavin in different redox and protonation states: A computational perspective on the effect of the electrostatic environment. Phys. Chem. Chem. Phys. 2019, 21, 16526–16537. [Google Scholar] [CrossRef]
- Pawelek, P.; Cheah, J.; Coulombe, R.; Macheroux, P.; Ghisla, S.; Vrielink, A. The structure of L-amino acid oxidase reveals the substrate trajectory into an enantiomerically conserved active site. EMBO J. 2000, 19, 4204–4215. [Google Scholar] [CrossRef] [PubMed]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular Evolutionary Genetics Analysis across Computing Platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef] [PubMed]
- Davis, M.A.; Askin, M.C.; Hynes, M.J. Amino acid catabolism by an areA-regulated gene encoding an L-amino acid oxidase with broad substrate specificity in Aspergillus nidulans. Appl. Environ. Microbiol. 2005, 71, 3551–3555. [Google Scholar] [CrossRef]
- Nuutinen, J.T.; Timonen, S. Identification of nitrogen mineralization enzymes, L-amino acid oxidases, from the ectomycorrhizal fungi Hebeloma spp. and Laccaria bicolor. Mycol. Res. 2008, 112, 1453–1464. [Google Scholar] [CrossRef] [PubMed]
- Mehta, P.K.; Hale, T.I.; Christen, P. Aminotransferases: Demonstration of homology and division into evolutionary subgroups. Eur. J. Biochem. 1993, 214, 549–561. [Google Scholar] [CrossRef]
- Nikodijević, D.D.; Jovankić, J.V.; Cvetković, D.M.; Anđelković, M.Z.; Nikezić, A.G.; Milutinović, M.G. L-amino acid oxidase from snake venom: Biotransformation and induction of apoptosis in human colon cancer cells. Eur. J. Pharmacol. 2021, 910, 174466. [Google Scholar] [CrossRef]
- Kitani, Y.; Fernandes, J.M.O.; Kiron, V. Identification of the Atlantic cod l-amino acid oxidase and its alterations following bacterial exposure. Dev. Comp. Immunol. 2015, 50, 116–120. [Google Scholar] [CrossRef]
- Huang, R.; Li, M.; Gregory, R.L. Bacterial interactions in dental biofilm. Virulence 2011, 2, 435–444. [Google Scholar] [CrossRef]
- Yang, L.; Liu, Y.; Wu, H.; Høiby, N.; Molin, S.; Song, Z.J. Current understanding of multi-species biofilms. Int. J. Oral. Sci. 2011, 3, 74–81. [Google Scholar] [CrossRef]
- Nakano, S.; Minamino, Y.; Hasebe, F.; Ito, S. Deracemization and stereoinversion to aromatic d-amino acid derivatives with ancestral l-amino acid oxidase. ACS Catal. 2019, 9, 10152–10158. [Google Scholar] [CrossRef]
- Sehanobish, E.; Shin, S.; Sanchez-Amat, A.; Davidson, V.L. Steady-state kinetic mechanism of LodA, a novel cysteine tryptophylquinone-dependent oxidase. FEBS Lett. 2014, 588, 752–756. [Google Scholar] [CrossRef] [PubMed]
- Letunic, I.; Bork, P. Interactive Tree Of Life (iTOL) v4: Recent updates and new developments. Nucleic Acids Res. 2019, 47, W256–W259. [Google Scholar] [CrossRef] [PubMed]
- Studer, G.; Tauriello, G.; Bienert, S.; Biasini, M.; Johner, N.; Schwede, T. ProMod3—A versatile homology modelling toolbox. PLOS Comput. Biol. 2021, 17, e1008667. [Google Scholar] [CrossRef] [PubMed]
- Studer, G.; Rempfer, C.; Waterhouse, A.M.; Gumienny, R.; Haas, J.; Schwede, T. QMEANDisCo—distance constraints applied on model quality estimation. Bioinformatics 2020, 36, 1765–1771. [Google Scholar] [CrossRef] [PubMed]
- Papadopoulos, J.S.; Agarwala, R. COBALT: Constraint-based alignment tool for multiple protein sequences. Bioinformatics 2007, 23, 1073–1079. [Google Scholar] [CrossRef]
- Schrödinger, L.L.C. The PyMOL Molecular Graphics System, Version 2.0; Schrödinger, LLC: New York, NY, USA, 2017. [Google Scholar]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Substrate | kcat (s−1) | Km (mM) | kcat/Km (M−1·s−1) | R2 |
|---|---|---|---|---|
| Leu | 9.38 ± 0.21 | 0.0567 ± 0.0051 | 1.65 × 105 | 0.995 |
| Met | 10.9 ± 0.4 | 0.0737 ± 0.0011 | 1.48 × 105 | 0.99 |
| Gln | 12.0 ± 0.7 | 0.126 ± 0.025 | 9.51 × 104 | 0.979 |
| Phe | 0.698 ± 0.052 | 0.0376 ± 0.0096 | 1.85 × 104 | 0.969 |
| Trp | 0.50 ± 0.023 | 0.0565 ± 0.0123 | 8.85 × 103 | 0.984 |
| Tyr | 0.478 ± 0.025 | 0.0552 ± 0.0109 | 8.67 × 103 | 0.972 |
| Ile | 6.39 ± 0.34 | 3.65 ± 0.61 | 1.75 × 103 | 0.986 |
| Glu | 1.45 ± 0.12 | 0.921 ± 0.235 | 1.58 × 103 | 0.967 |
| Arg | 1.07 ± 0.05 | 2.11 ± 0.38 | 5.08 × 102 | 0.967 |
| Val | 2.80 ± 0.28 | 6.28 ± 1.17 | 4.45 × 102 | 0.981 |
| Ala | 5.26 ± 1.13 | 21.7 ± 0.83 | 2.42 × 102 | 0.99 |
| Cys | 0.113 ± 0.012 | 0.467 ± 0.215 | 2.41 × 102 | 0.965 |
| Lys | 5.10 ± 0.27 | 43.1 ± 7.6 | 1.18 × 102 | 0.969 |
| His | 0.108 ± 0.008 | 1.01 ± 0.30 | 1.07 × 102 | 0.978 |
| Asn | 0.259 ± 0.022 | 7.96 ± 1.63 | 32.5 | 0.969 |
| Ser | 0.281 ± 0.032 | 40.3 ± 9.81 | 6.97 | 0.986 |
| Thr | 0.135 ± 0.008 | 173 ± 29 | 0.777 | 0.984 |
| Asp | 1.72 ± 0.98 × 10−3 | 14.3 ± 27.6 | 0.12 | 0.832 |
| Gly | 5.45 ± 0.55 × 10−3 | 71.1 ± 31.4 | 7.67 × 10−2 | 0.923 |
| Substrate | kcat (s−1) | Km (mM) | kcat/Km (M−1·s−1) | R2 |
|---|---|---|---|---|
| Trp | 0.50 ± 0.023 | 0.0565 ± 0.0123 | 8.85 × 103 | 0.984 |
| 5-OH Trp | 0.371 ± 0.056 | 0.462 ± 1.97 | 8.04 × 102 | 0.935 |
| Tyr | 0.478 ± 0.025 | 0.0552 ± 0.0109 | 8.67 × 103 | 0.972 |
| 3-Nitro Tyr | 0.545 ± 0.045 | 0.038.6 ± 10 | 1.41 × 104 | 0.981 |
| Cys | 0.113 ± 0.012 | 0.467 ± 0.215 | 2.41 × 102 | 0.965 |
| Homo Cys | 0.179 ± 0.062 | 0.306 ± 0.238 | 5.84 × 102 | 0.982 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mamounis, K.J.; Caldas Nogueira, M.L.; Marchi Salvador, D.P.; Andreo-Vidal, A.; Sanchez-Amat, A.; Davidson, V.L. Structural Determinants of the Specific Activities of an L-Amino Acid Oxidase from Pseudoalteromonas luteoviolacea CPMOR-1 with Broad Substrate Specificity. Molecules 2022, 27, 4726. https://doi.org/10.3390/molecules27154726
Mamounis KJ, Caldas Nogueira ML, Marchi Salvador DP, Andreo-Vidal A, Sanchez-Amat A, Davidson VL. Structural Determinants of the Specific Activities of an L-Amino Acid Oxidase from Pseudoalteromonas luteoviolacea CPMOR-1 with Broad Substrate Specificity. Molecules. 2022; 27(15):4726. https://doi.org/10.3390/molecules27154726
Chicago/Turabian StyleMamounis, Kyle J., Maria Luiza Caldas Nogueira, Daniela Priscila Marchi Salvador, Andres Andreo-Vidal, Antonio Sanchez-Amat, and Victor L. Davidson. 2022. "Structural Determinants of the Specific Activities of an L-Amino Acid Oxidase from Pseudoalteromonas luteoviolacea CPMOR-1 with Broad Substrate Specificity" Molecules 27, no. 15: 4726. https://doi.org/10.3390/molecules27154726
APA StyleMamounis, K. J., Caldas Nogueira, M. L., Marchi Salvador, D. P., Andreo-Vidal, A., Sanchez-Amat, A., & Davidson, V. L. (2022). Structural Determinants of the Specific Activities of an L-Amino Acid Oxidase from Pseudoalteromonas luteoviolacea CPMOR-1 with Broad Substrate Specificity. Molecules, 27(15), 4726. https://doi.org/10.3390/molecules27154726