
Ameliorative Effect of Citropten Isolated from Citrusaurantifolia Peel Extract as a Modulator of T Cell and Intestinal Epithelial Cell Activity in DSS-Induced Colitis

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Results
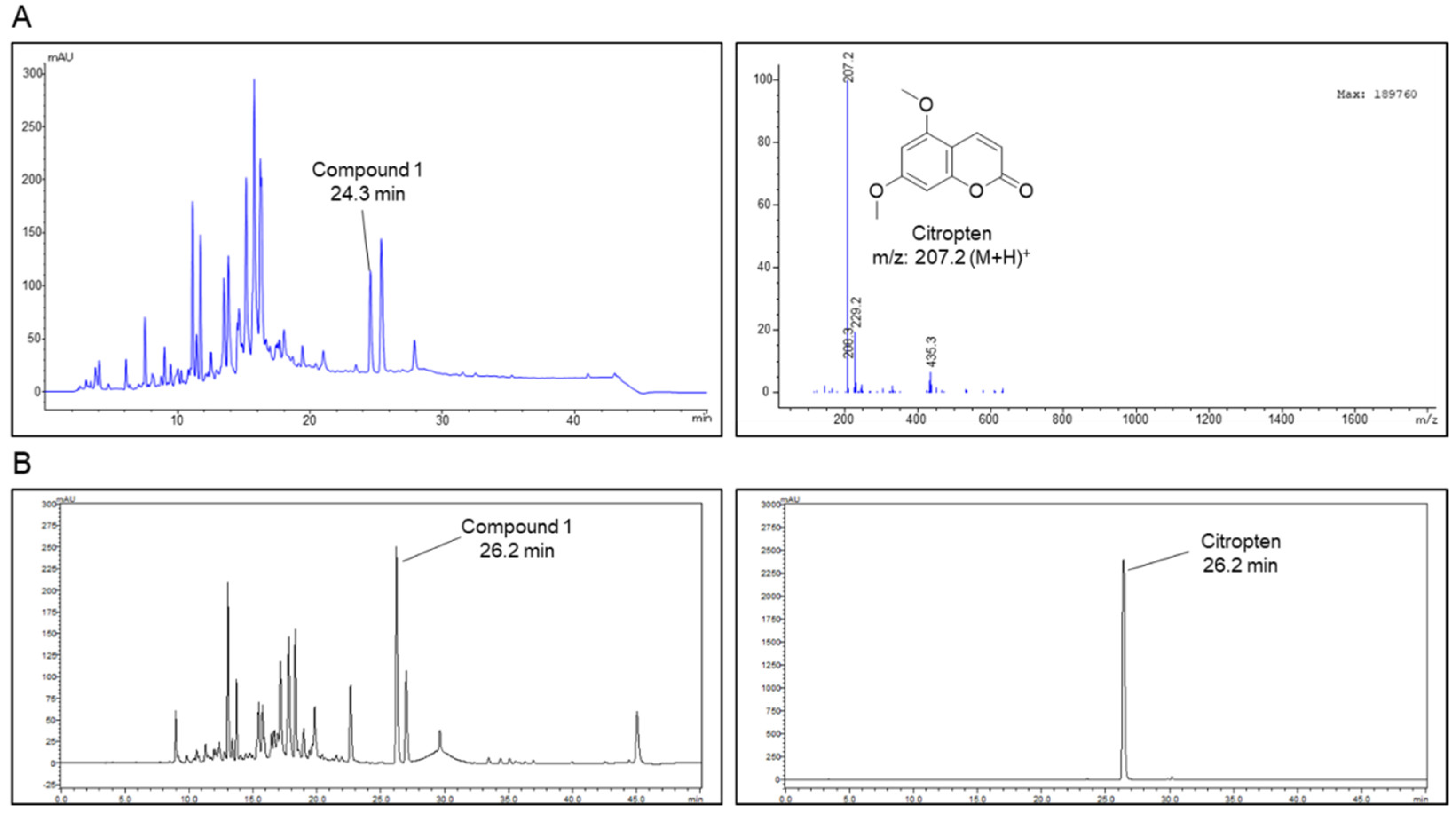
2.1. Isolation of Citropten from C. aurantifolia Peel Extract and Its Chemical Structure
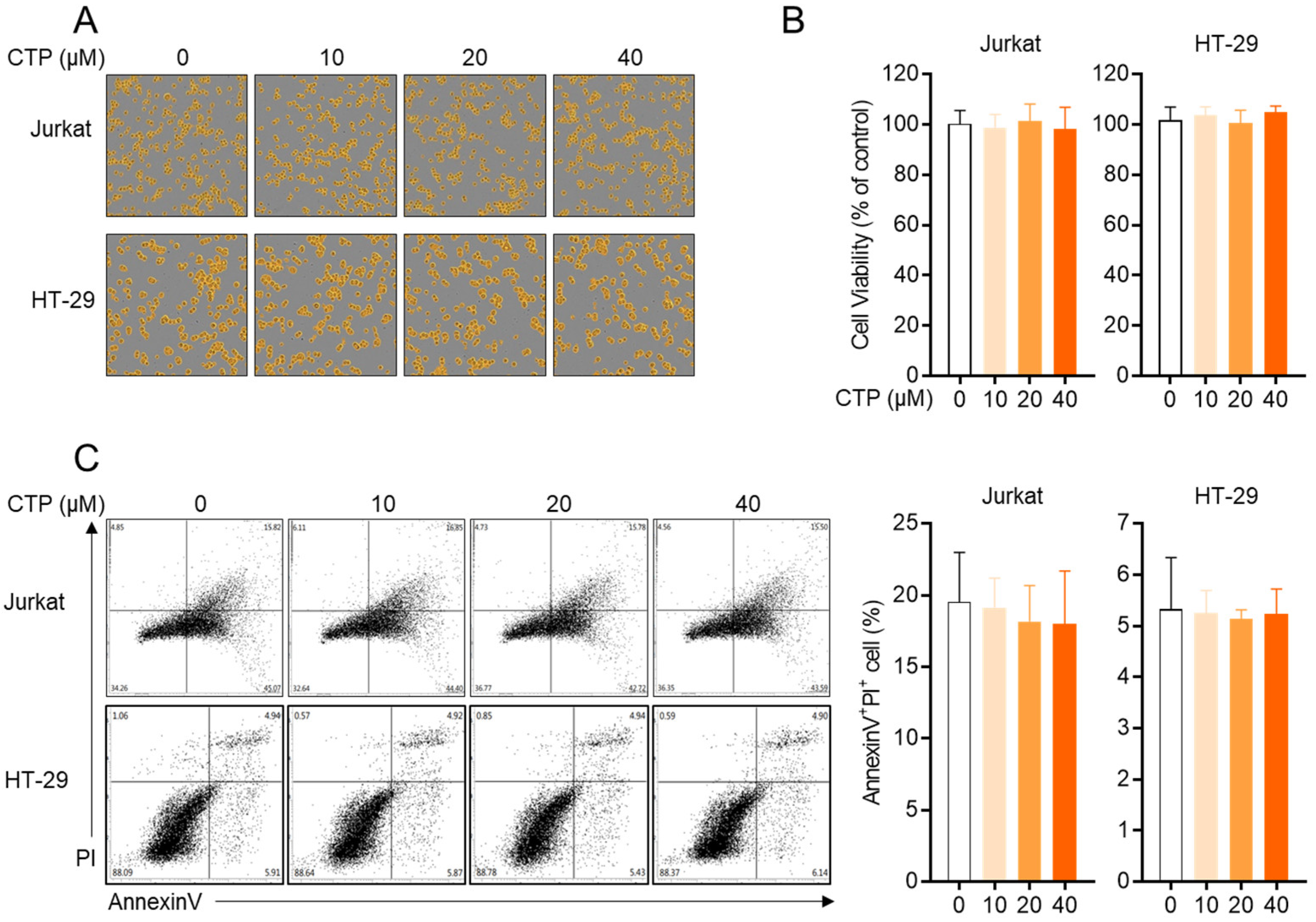
2.2. Citropten Has No Negative Effect on the Viability of Jurkat and HT-29 Cells
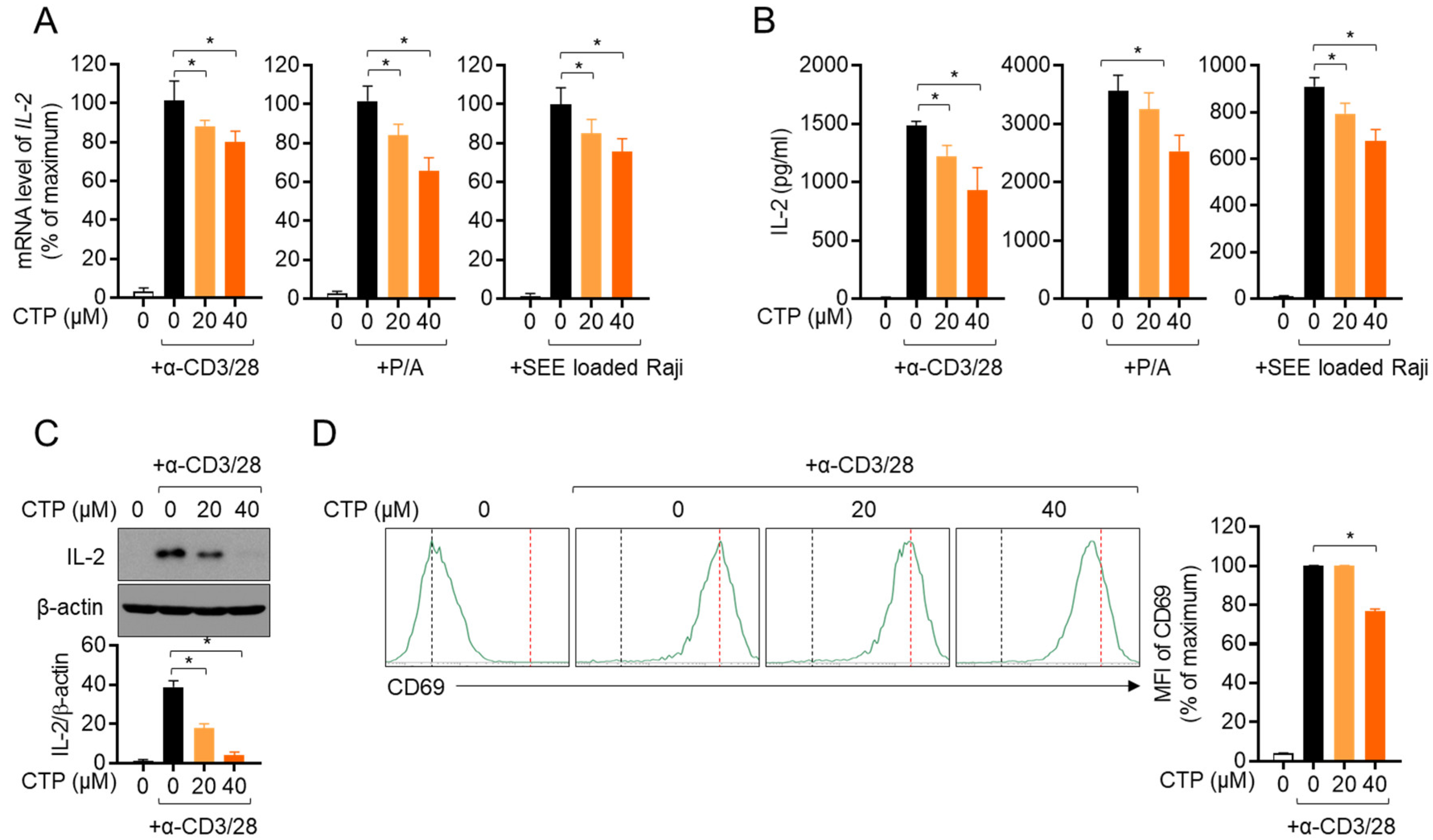
2.3. Activity of Jurkat T Cells Is Downregulated by Pre-Treatment with Citropten
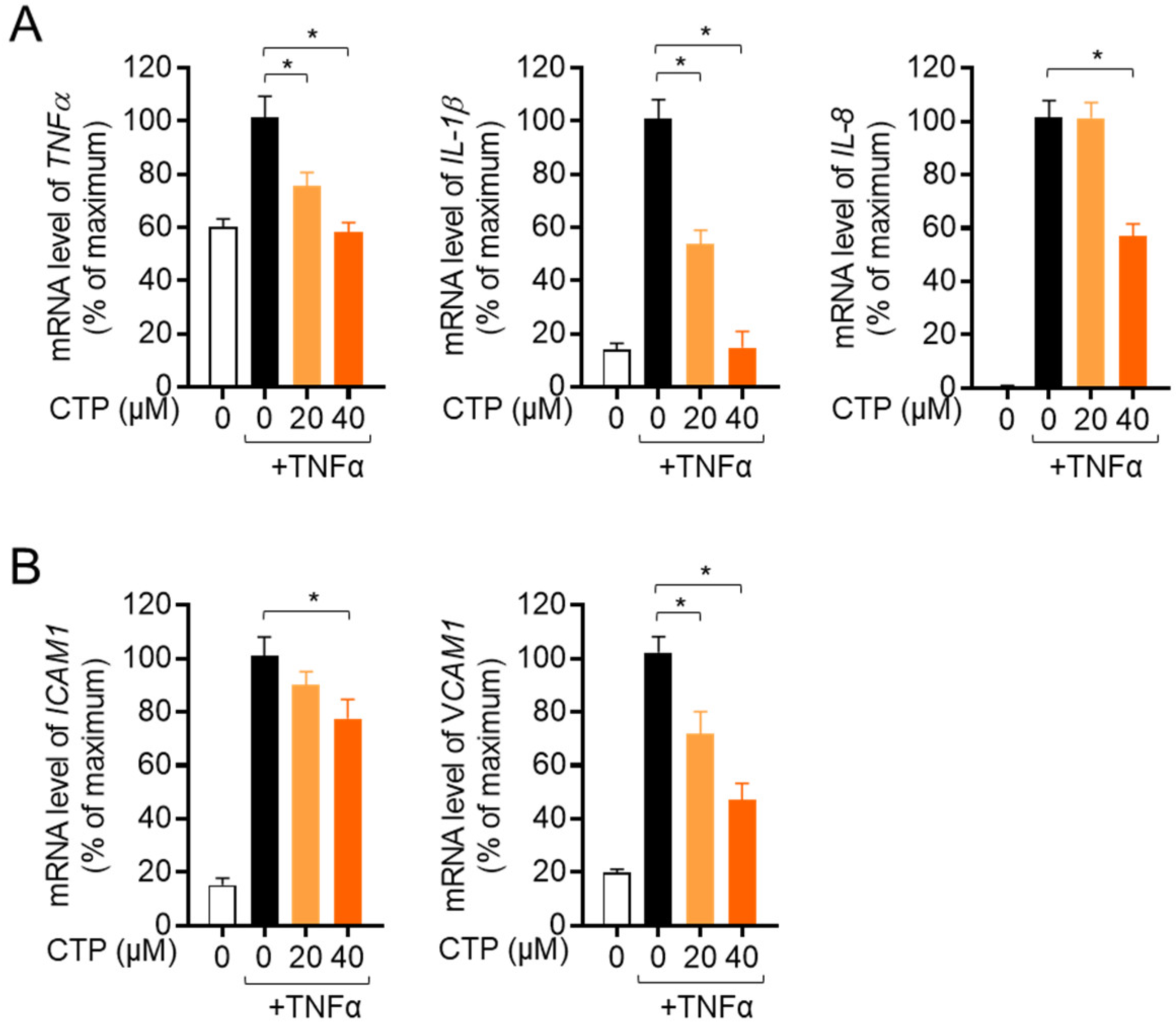
2.4. Citropten Suppresses the Production of Inflammatory Cytokines in Activated HT-29 Cells
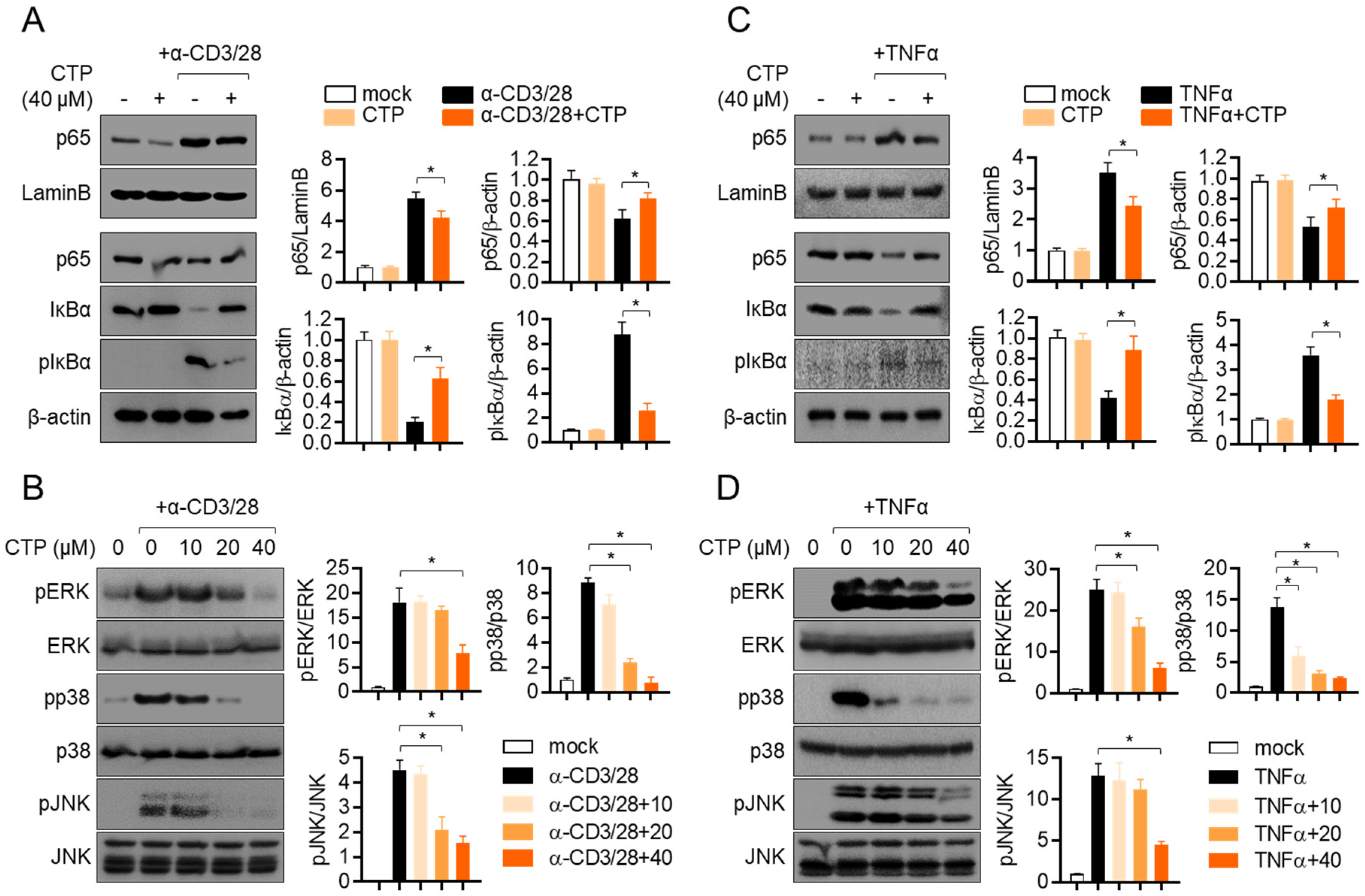
2.5. Pre-Treatment with Citropten Reduces NFκB and MAPK Signaling Pathway in Activated Jurkat and HT-29
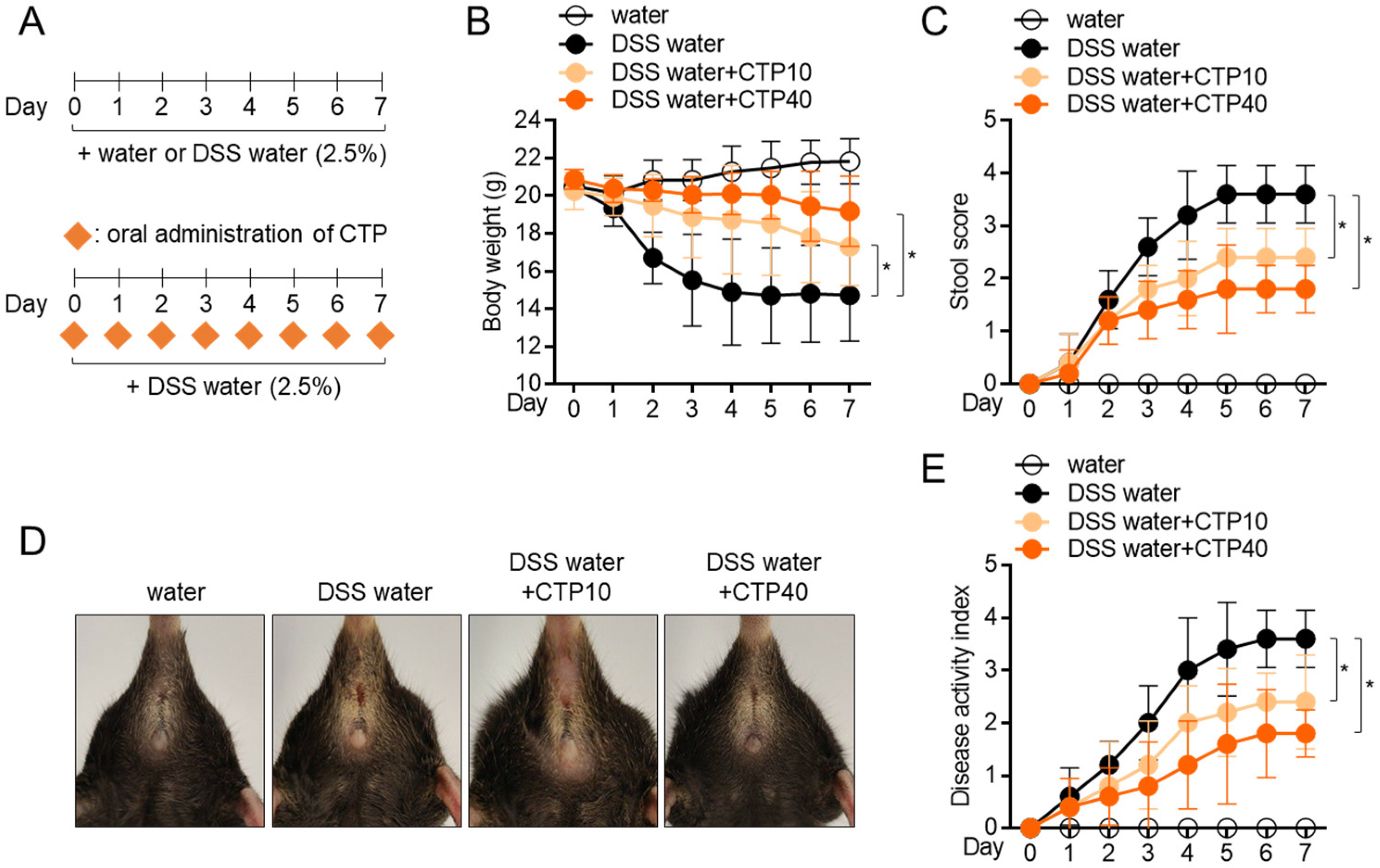
2.6. Oral Administration of Citropten Attenuates DSS-Induced Colitis in Mice Model
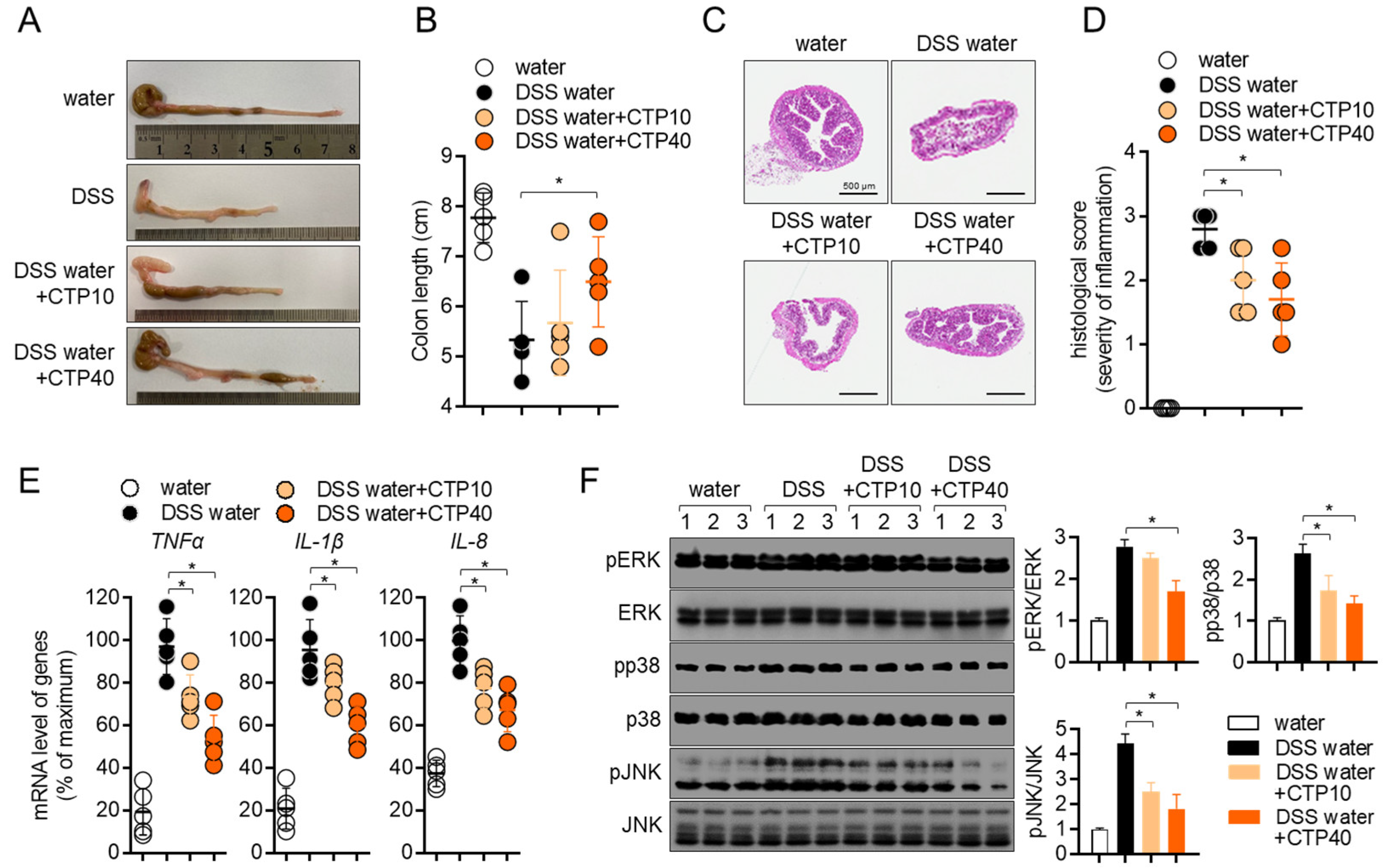
2.7. Oral Administration of Citropten Alleviates the Colonic Inflammation in DSS-Induced Colitis Model
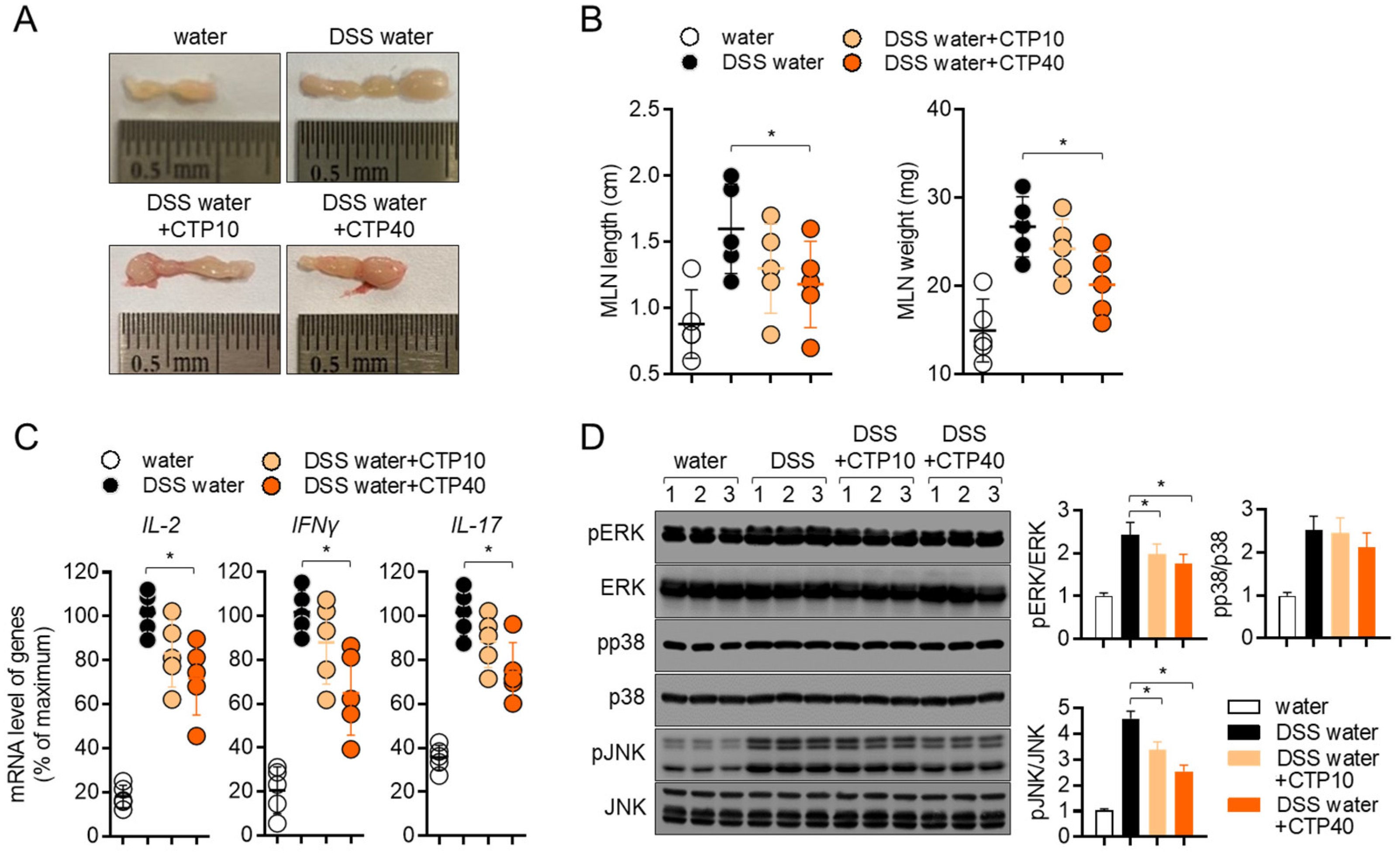
2.8. Oral Treatment with Citropten Ameliorates T Cell Activity in DSS-Induced Colitis Model
3. Discussion
4. Materials and Methods
4.1. Cell Culture
4.2. Animals
4.3. Plant Material
4.4. Isolation of Citropten from C. aurantifolia Peel Extract
4.5. Condition of Liquid Chromatography–Mass Spectrometry Analysis
4.6. Reagents and Antibodies
4.7. Cell Confluency Check by IncuCyte Imaging System
4.8. Cell Viability Check by MTT Assay
4.9. AnnexinV/PI Apoptosis Assay
4.10. T Cell Stimulation
4.11. Determination of mRNA Levels by Real-Time Quantitative PCR
4.12. ELISA
4.13. Western Blot Analysis
4.14. Determination of CD69 Expression by Flow Cytometry
4.15. Induction of Colitis by Using DSS
4.16. Determination of Stool Scoring
4.17. Determination of Disease Activity Index
4.18. H&E Staining and Determination of Histological Score
4.19. Statistics
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Wedro, B. Colitis: Symptoms, Causes, Diet, and Treatment. MedicineNet 2021. Available online: https://www.medicinenet.com/colitis/article.htm (accessed on 23 March 2022).
- Ordás, I.; Eckmann, L.; Talamini, M.; Baumgart, D.C.; Sandborn, W.J. Ulcerative colitis. Lancet 2012, 380, 1606–1619. [Google Scholar] [CrossRef]
- Fiocchi, C. Inflammatory bowel disease: Etiology and pathogenesis. Gastroenterology 1998, 115, 182–205. [Google Scholar] [CrossRef]
- Ko, J.; Auyeung, K. Inflammatory Bowel Disease: Etiology, Pathogenesis and Current Therapy. Curr. Pharm. Des. 2014, 20, 1082–1096. [Google Scholar] [CrossRef] [PubMed]
- Neurath, M.F.; Finotto, S.; Fuss, I.; Boirivant, M.; Galle, P.R.; Strober, W. Regulation of T-cell apoptosis in inflammatory bowel disease: To die or not to die, that is the mucosal question. Trends Immunol. 2001, 22, 21–26. [Google Scholar] [CrossRef]
- Coskun, M. Intestinal epithelium in inflammatory bowel disease. Front. Med. 2014, 1, 24. [Google Scholar] [CrossRef]
- Weiss, A. TCR Signal Transduction: Opening the Black Box. J. Immunol. 2009, 183, 4821–4827. [Google Scholar] [CrossRef]
- Shapiro, V.S.; Truitt, K.E.; Imboden, J.B.; Weiss, A. CD28 mediates transcriptional upregulation of the interleukin-2 (IL-2) promoter through a composite element containing the CD28RE and NF-IL-2B AP-1 sites. Mol. Cell. Biol. 1997, 17, 4051–4058. [Google Scholar] [CrossRef]
- Cheng, J.; Montecalvo, A.; Kane, L.P. Regulation of NF-κB induction by TCR/CD28. Immunol. Res. 2011, 50, 113–117. [Google Scholar] [CrossRef]
- Bonizzi, G.; Karin, M. The two NF-κB activation pathways and their role in innate and adaptive immunity. Trends Immunol. 2004, 25, 280–288. [Google Scholar] [CrossRef]
- Hayden, M.S.; Ghosh, S. Signaling to NF-κB. Genes Dev. 2004, 18, 2195–2224. [Google Scholar] [CrossRef]
- Hayes, J.D.; Pulford, D.J.; Ellis, E.M.; McLeod, R.; James, R.F.L.; Seidegård, J.; Mosialou, E.; Jernström, B.; Neal, G.E. Regulation of rat glutathione S-transferase A5 by cancer chemopreventive agents: Mechanisms of inducible resistance to aflatoxin B1. Chem. Biol. Interact. 1998, 111, 51–67. [Google Scholar] [CrossRef]
- Sharma, S.; Stutzman, J.D.; Kelloff, G.J.; Steele, V.E. Screening of Potential Chemopreventive Agents Using Biochemical Markers of Carcinogenesis. Cancer Res. 1994, 54, 5848–5855. [Google Scholar] [PubMed]
- Borgatti, M.; Mancini, I.; Bianchi, N.; Guerrini, A.; Lampronti, I.; Rossi, D.; Sacchetti, G.; Gambari, R. Bergamot (Citrus bergamia Risso) fruit extracts and identified components alter expression of interleukin 8 gene in cystic fibrosis bronchial epithelial cell lines. BMC Biochem. 2011, 12, 15. [Google Scholar] [CrossRef] [PubMed]
- Canini, A.; Alesiani, D.; D’Arcangelo, G.; Tagliatesta, P. Gas chromatography-mass spectrometry analysis of phenolic compounds from Carica papaya L. leaf. J. Food Compos. Anal. 2007, 20, 584–590. [Google Scholar] [CrossRef]
- Salvatore, A.; Borkosky, S.; Willink, E.; Bardón, A. Toxic effects of lemon peel constituents on Ceratitis capitata. J. Chem. Ecol. 2004, 30, 323–333. [Google Scholar] [CrossRef]
- Alesiani, D.; Cicconi, R.; Mattei, M.; Montesano, C.; Bei, R.; Canini, A. Cell cycle arrest and differentiation induction by 5,7-dimethoxycoumarin in melanoma cell lines. Int. J. Oncol. 2008, 32, 425–434. [Google Scholar] [CrossRef]
- Fang, J.Y.; Richardson, B.C. The MAPK signalling pathways and colorectal cancer. Lancet Oncol. 2005, 6, 322–327. [Google Scholar] [CrossRef]
- Yang, W.; Wang, H. 5,7-Dimethoxycoumarin prevents chronic mild stress induced depression in rats through increase in the expression of heat shock protein-70 and inhibition of monoamine oxidase-A levels. Saudi J. Biol. Sci. 2018, 25, 253–258. [Google Scholar] [CrossRef]
- Musumeci, L.; Maugeri, A.; Cirmi, S.; Lombardo, G.E.; Russo, C.; Gangemi, S.; Calapai, G.; Navarra, M. Citrus fruits and their flavonoids in inflammatory bowel disease: An overview. Nat. Prod. Res. 2020, 34, 122–136. [Google Scholar] [CrossRef]
- Patil, J.R.; Jayaprakasha, G.K.; Chidambara Murthy, K.N.; Chetti, M.B.; Patil, B.S. Characterization of Citrus aurantifolia bioactive compounds and their inhibition of human pancreatic cancer cells through apoptosis. Microchem. J. 2009, 94, 108–117. [Google Scholar] [CrossRef]
- Dunleavy, K.A.; Ungaro, R.C.; Manning, L.; Gold, S.; Novak, J.; Colombel, J.F. Vitamin C Deficiency: The Forgotten Micronutrient. Crohn’s Colitis 360 2021, 3, 1. [Google Scholar]
- Kondo, K.; Hiramoto, K.; Yamate, Y.; Goto, K.; Sekijima, H.; Ooi, K. Ameliorative Effect of High-Dose Vitamin C Administration on Dextran Sulfate Sodium-Induced Colitis Mouse Model. Biol. Pharm. Bull. 2019, 42, 954–959. [Google Scholar] [CrossRef] [PubMed]
- Toribio, M.L.; Gutiérrez-Ramos, J.C.; Pezzi, L.; Marcos, M.A.R.; Martínez-A, C. lnterleukin-2-dependent autocrine proliferation in T-cell development. Nature 1989, 342, 82–85. [Google Scholar] [CrossRef] [PubMed]
- Lacy, P.; Stow, J.L. Cytokine release from innate immune cells: Association with diverse membrane trafficking pathways. Blood 2011, 118, 9–18. [Google Scholar] [CrossRef] [PubMed]
- Stanley, A.C.; Lacy, P. Pathways for cytokine secretion. Physiology 2010, 25, 218–229. [Google Scholar] [CrossRef] [PubMed]
- Marsal, J.; Agace, W.W. Targeting T-cell migration in inflammatory bowel disease. J. Intern. Med. 2012, 272, 411–429. [Google Scholar] [CrossRef] [PubMed]
- Stephens, M.; Liao, S.; Von Der Weid, P.Y. Mesenteric lymphatic alterations observed during DSS induced intestinal inflammation are driven in a TLR4-PAMP/DAMP discriminative manner. Front. Immunol. 2019, 10, 557. [Google Scholar] [CrossRef] [PubMed]
- Heitman, B.; Irizarry, A. Infectious disease causes of lymphadenopathy: Localized versus diffuse. Lippincott’s Prim. Care Pract. 1999, 3, 19–38. [Google Scholar]
- Oh, H.; Ghosh, S. NF-κB: Roles and regulation in different CD4+ T-cell subsets. Immunol. Rev. 2013, 252, 41–51. [Google Scholar] [CrossRef]
- Park, S.-H.; Cho, G.; Park, S.-G. NF-κB Activation in T Helper 17 Cell Differentiation. Immune Netw. 2014, 14, 14. [Google Scholar] [CrossRef]
- Ruan, Q.; Kameswaran, V.; Zhang, Y.; Zheng, S.; Sun, J.; Wang, J.; DeVirgiliis, J.; Liou, H.C.; Beg, A.A.; Chen, Y.H. The Th17 immune response is controlled by the Rel-RORγ-RORγT transcriptional axis. J. Exp. Med. 2011, 208, 2321–2333. [Google Scholar] [CrossRef] [PubMed]
- Broom, O.J.; Widjaya, B.; Troelsen, J.; Olsen, J.; Nielsen, O.H. Mitogen Activated Protein Kinases: A Role in Inflammatory Bowel Disease? Clin. Exp. Immunol. 2009, 158, 272–280. [Google Scholar] [CrossRef] [PubMed]
- Kwon, K.H.; Ohigashi, H.; Murakami, A. Dextran Sulfate Sodium Enhances Interleukin-1β Release via Activation of P38 MAPK and ERK1/2 Pathways in Murine Peritoneal Macrophages. Life Sci. 2007, 81, 362–371. [Google Scholar] [CrossRef] [PubMed]
- Caruso, R.; Fina, D.; Peluso, I.; Stolfi, C.; Fantini, M.C.; Gioia, V.; Caprioli, F.; del Vecchio Blanco, G.; Paoluzi, O.A.; MacDonald, T.T.; et al. A Functional Role for Interleukin-21 in Promoting the Synthesis of the T-Cell Chemoattractant, MIP-3alpha, by Gut Epithelial Cells. Gastroenterology 2007, 132, 166–175. [Google Scholar] [CrossRef] [PubMed]
- Waetzig, G.H.; Seegert, D.; Rosenstiel, P.; Nikolaus, S.; Schreiber, S. P38 Mitogen-Activated Protein Kinase Is Activated and Linked to TNF-α Signaling in Inflammatory Bowel Disease. J. Immunol. 2002, 168, 5342–5351. [Google Scholar] [CrossRef] [PubMed]
- Dahan, S.; Roda, G.; Pinn, D.; Roth-Walter, F.; Kamalu, O.; Martin, A.P.; Mayer, L. Epithelial: Lamina Propria Lymphocyte Interactions Promote Epithelial Cell Differentiation. Gastroenterology 2008, 134, 192. [Google Scholar] [CrossRef]
- Assi, K.; Pillai, R.; Gómez-Muñoz, A.; Owen, D.; Salh, B. The Specific JNK Inhibitor SP600125 Targets Tumour Necrosis Factor-Alpha Production and Epithelial Cell Apoptosis in Acute Murine Colitis. Immunology 2006, 118, 112–121. [Google Scholar] [CrossRef]
- Mitsuyama, K.; Suzuki, A.; Tomiyasu, N.; Tsuruta, O.; Kitazaki, S.; Takeda, T.; Satoh, Y.; Bennett, B.L.; Toyonaga, A.; Sata, M. Pro-Inflammatory Signaling by Jun-N-Terminal Kinase in Inflammatory Bowel Disease. Int. J. Mol. Med. 2006, 17, 449–455. [Google Scholar] [CrossRef][Green Version]
- Reinisch, W.; Hung, K.; Hassan-Zahraee, M.; Cataldi, F. Targeting endothelial ligands: ICAM-1/alicaforsen, MAdCAM-1. J. Crohn’s Colitis 2018, 12, S669–S677. [Google Scholar] [CrossRef]
- Soriano, A.; Salas, A.; Sans, M.; Gironella, M.; Elena, M.; Anderson, D.C.; Piqué, J.M.; Panés, J. VCAM-1, but not ICAM-1 or MAdCAM-1, immunoblockade ameliorates DSS-induced colitis in mice. Lab. Investig. 2000, 80, 1541–1551. [Google Scholar] [CrossRef]
- Burns, R.C.; Rivera-Nieves, J.; Moskaluk, C.A.; Matsumoto, S.; Cominelli, F.; Ley, K. Antibody blockade of ICAM-1 and VCAM-1 ameliorates inflammation in the SAMP-1/Yit adoptive transfer model of Crohn’s disease in mice. Gastroenterology 2001, 121, 1428–1436. [Google Scholar] [CrossRef]
- Villablanca, E.J.; Cassani, B.; Von Andrian, U.H.; Mora, J.R. Blocking lymphocyte localization to the gastrointestinal mucosa as a therapeutic strategy for inflammatory bowel diseases. Gastroenterology 2011, 140, 1776–1784. [Google Scholar] [CrossRef] [PubMed]
- Henrique, D.M.; Ana, C.P.; Guilherme, T.P.B.; Helton, C.L.; Nelson, S.M.J.; Luciano, B.; Mark, W.; Jairo, K.B.; Gilberto, U.L.B. Furocoumarins and coumarins photoinactivate Colletotrichum acutatum and Aspergillus nidulans fungi under solar radiation. J. Photochem. Photobiol. B 2014, 131, 74–83. [Google Scholar]
- Koelink, P.J.; Wildenberg, M.E.; Stitt, L.W.; Feagan, B.G.; Koldijk, M.; van’t Wout, A.B.; Atreya, R.; Vieth, M.; Brandse, J.F.; Duijst, S.; et al. Development of reliable, valid and responsive scoring systems for endoscopy and histology in animal models for inflammatory bowel disease. J. Crohn’s Colitis 2018, 12, 794–803. [Google Scholar] [CrossRef] [PubMed]








Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lee, H.-S.; Kim, E.-N.; Jeong, G.-S. Ameliorative Effect of Citropten Isolated from Citrusaurantifolia Peel Extract as a Modulator of T Cell and Intestinal Epithelial Cell Activity in DSS-Induced Colitis. Molecules 2022, 27, 4633. https://doi.org/10.3390/molecules27144633
Lee H-S, Kim E-N, Jeong G-S. Ameliorative Effect of Citropten Isolated from Citrusaurantifolia Peel Extract as a Modulator of T Cell and Intestinal Epithelial Cell Activity in DSS-Induced Colitis. Molecules. 2022; 27(14):4633. https://doi.org/10.3390/molecules27144633
Chicago/Turabian StyleLee, Hyun-Su, Eun-Nam Kim, and Gil-Saeng Jeong. 2022. "Ameliorative Effect of Citropten Isolated from Citrusaurantifolia Peel Extract as a Modulator of T Cell and Intestinal Epithelial Cell Activity in DSS-Induced Colitis" Molecules 27, no. 14: 4633. https://doi.org/10.3390/molecules27144633
APA StyleLee, H.-S., Kim, E.-N., & Jeong, G.-S. (2022). Ameliorative Effect of Citropten Isolated from Citrusaurantifolia Peel Extract as a Modulator of T Cell and Intestinal Epithelial Cell Activity in DSS-Induced Colitis. Molecules, 27(14), 4633. https://doi.org/10.3390/molecules27144633