
Comparison of Fatty Acid Contents and MMP-1 Inhibitory Effects of the Two Antarctic Fish, Notothenia rossii and Champsocephalus gunnari

 , , ,
, , ,  , ,
, ,
Abstract
:1. Introduction
2. Results and Discussion
2.1. GC–MS Analysis of the Fish Extracts
2.1.1. Brain
2.1.2. Liver
2.1.3. Stomach
2.1.4. Skin
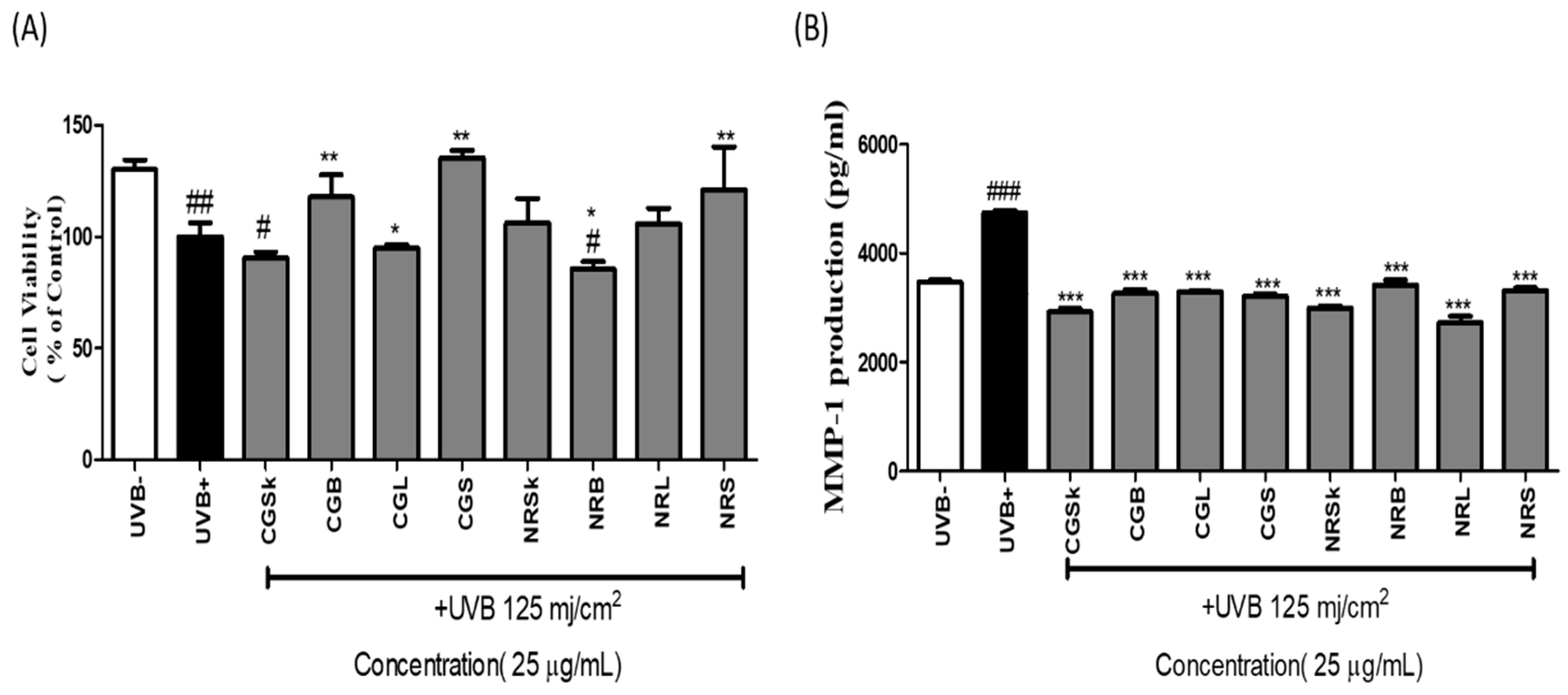
2.2. Cell Viability
2.3. MMP-1-Inhibitory Activity
3. Discussion
4. Materials and Methods
4.1. Animals
4.2. Chemicals and Reagents
4.3. Lipid Extraction
4.4. Fatty-Acid Analyses
4.5. Cell Culture
4.6. Cell Viability Assay
4.7. Measurement of MMP-1 in NHDF Cells
4.8. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Sample Availability
References
- Wong, Q.Y.A.; Chew, F.T. Defining Skin Aging and Its Risk Factors: A Systematic Review and Meta-Analysis. Sci. Rep. 2021, 11, 22075. [Google Scholar] [CrossRef] [PubMed]
- Kirkwood, K.L. Inflammaging. Immunol. Investig. 2018, 47, 770–773. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moon, H.J.; Lee, S.R.; Shim, S.N.; Jeong, S.H.; Stonik, V.A.; Rasskazov, V.A.; Zvyagintseva, T.; Lee, Y.H. Fucoidan Inhibits Uvb-Induced Mmp-1 Expression in Human Skin Fibroblasts. Biol. Pharm. Bull. 2008, 31, 284–289. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fisher, G.J.; Datta, S.C.; Talwar, H.S.; Wang, Z.-Q.; Varani, J.; Kang, S.; Voorhees, J.J. Molecular Basis of Sun-Induced Premature Skin Ageing and Retinoid Antagonism. Nature 1996, 379, 335–339. [Google Scholar] [CrossRef]
- Kohl, E.; Steinbauer, J.; Landthaler, M.; Szeimies, R.M. Skin Ageing. J. Eur. Acad. Dermatol. Venereol. 2011, 25, 873–884. [Google Scholar] [CrossRef]
- Dong, K.K.; Damaghi, N.; Picart, S.D.; Markova, N.G.; Obayashi, K.; Okano, Y.; Masaki, H.; Grether-Beck, S.; Krutmann, J.; Smiles, K.A. Uv-Induced DNA Damage Initiates Release of Mmp-1 in Human Skin. Exp. Dermatol. 2008, 17, 1037–1044. [Google Scholar] [CrossRef]
- Pilkington, S.M.; Watson, R.E.B.; Nicolaou, A.; Rhodes, L.E. Omega-3 Polyunsaturated Fatty Acids: Photoprotective Macronutrients. Exp. Dermatol. 2011, 20, 537–543. [Google Scholar] [CrossRef]
- Huang, T.-H.; Wang, P.-W.; Yang, S.-C.; Chou, W.-L.; Fang, J.-Y. Cosmetic and Therapeutic Applications of Fish Oil’s Fatty Acids on the Skin. Mar. Drugs 2018, 16, 256. [Google Scholar] [CrossRef] [Green Version]
- Engström, K.; Saldeen, A.-S.; Yang, B.; Mehta, J.L.; Saldeen, T. Effect of Fish Oils Containing Different Amounts of Epa, Dha, and Antioxidants on Plasma and Brain Fatty Acids and Brain Nitric Oxide Synthase Activity in Rats. Upsala J. Med. Sci. 2009, 114, 206–213. [Google Scholar] [CrossRef]
- Eastman, J.; Grande, L. Evolution of the Antarctic Fish Fauna with Emphasis on the Recent Notothenioids. Geol. Soc. Lond. Spec. Publ. 1989, 47, 241–252. [Google Scholar] [CrossRef]
- Eastman, J.T. The Nature of the Diversity of Antarctic Fishes. Polar Biol. 2005, 28, 93–107. [Google Scholar] [CrossRef]
- Stowasser, G.; Pond, D.W.; Collins, M.A. Fatty Acid Trophic Markers Elucidate Resource Partitioning within the Demersal Fish Community of South Georgia and Shag Rocks (Southern Ocean). Mar. Biol. 2012, 159, 2299–2310. [Google Scholar] [CrossRef]
- Machado, C.; Zaleski, T.; Rodrigues, E.; dos Santos Carvalho, C.; Cadena, S.M.S.C.; Gozzi, G.J.; Krebsbach, P.; Rios, F.S.A.; Donatti, L. Effect of Temperature Acclimation on the Liver Antioxidant Defence System of the Antarctic Nototheniids Notothenia Coriiceps and Notothenia Rossii. Comp. Biochem. Physiol. Part B Biochem. Mol. Biol. 2014, 172, 21–28. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rodrigues, E.; Feijó-Oliveira, M.; Vani, G.; Suda, C.; Carvalho, C.; Donatti, L.; Lavrado, H. Interaction of Warm Acclimation, Low Salinity, and Trophic Fluoride on Plasmatic Constituents of the Antarctic Fish Notothenia Rossii Richardson, 1844. Fish Physiol. Biochem. 2013, 39, 1591–1601. [Google Scholar] [CrossRef] [PubMed]
- De Souza, M.R.D.P.; Herrerias, T.; Zaleski, T.; Forgati, M.; Kandalski, P.K.; Machado, C.; Silva, D.T.; Piechnik, C.A.; Moura, M.O.; Donatti, L. Heat Stress in the Heart and Muscle of the Antarctic Fishes Notothenia Rossii and Notothenia Coriiceps: Carbohydrate Metabolism and Antioxidant Defence. Biochimie 2018, 146, 43–55. [Google Scholar] [CrossRef] [PubMed]
- Lund, E.; Sidell, B. Neutral Lipid Compositions of Antarctic Fish Tissues May Reflect Use of Fatty Acyl Substrates by Catabolic Systems. Mar. Biol. 1992, 112, 377–382. [Google Scholar] [CrossRef]
- Sidell, B.D.; Crockett, E.L.; Driedzic, W.R. Antarctic Fish Tissues Preferentially Catabolize Monoenoic Fatty Acids. J. Exp. Zool. 1995, 271, 73–81. [Google Scholar] [CrossRef]
- Hagen, W.; Kattner, G.; Friedrich, C. The Lipid Compositions of High-Antarctic Notothenioid Fish Species with Different Life Strategies. Polar Biol. 2000, 23, 785–791. [Google Scholar] [CrossRef]
- Benitez, L.V. Amino Acid and Fatty Acid Profiles in Aquaculture Nutrition Studies. In Fish Nutrtiton Research in Asia: Proceedings of the Third Asian Fish Nutrition Network Meeting; De Silva, S.S., Ed.; Asian Fishers Society: Manila, Philippines, 1989; pp. 23–35. [Google Scholar]
- Miller, R.G. History and Atlas of the Fishes of the Antarctic Ocean; Foresta Institute for Ocean and Mountain Studies: Carson City, NV, USA, 1993. [Google Scholar]
- Pagliarani, A.; Pirini, M.; Trigari, G.; Ventrella, V. Effect of Diets Containing Different Oils on Brain Fatty Acid Composition in Sea Bass (Dicentrarchus labrax L.). Comp. Biochem. Physiol. B Comp. Biochem. 1986, 83, 277–282. [Google Scholar]
- Sharma, P.; Kumar, V.; Sinha, A.K.; Ranjan, J.; Kithsiri, H.; Venkateshwarlu, G. Comparative Fatty Acid Profiles of Wild and Farmed Tropical Freshwater Fish Rohu (Labeo rohita). Fish Physiol. Biochem. 2010, 36, 411–417. [Google Scholar] [CrossRef]
- Stepanowska, K.; Nędzarek, A. Changes in the Body Chemical Composition and the Excretion of Nitrogen and Phosphorus During Long-Term Starvation of Antarctic Fish Notothenia Coriiceps and Notothenia Rossii. Eur. Zool. J. 2020, 87, 571–579. [Google Scholar] [CrossRef]
- Weber, K.; Goerke, H. Organochlorine Compounds in Fish Off the Antarctic Peninsula. Chemosphere 1996, 33, 377–392. [Google Scholar] [CrossRef]
- Osman, H.; Suriah, A.; Law, E. Fatty Acid Composition and Cholesterol Content of Selected Marine Fish in Malaysian Waters. Food Chem. 2001, 73, 55–60. [Google Scholar] [CrossRef]
- Magalhães, B.; Fiamoncini, J.; Deschamps, F.; Curi, R.; Silva, L. Comparison of Fatty Acid Composition in Nine Organs of the Sympatric Antarctic Teleost Fish Species Notothenia Coriiceps and Notothenia Rossii (Perciformes: Nototheniidae). Comp. Biochem. Physiol. Part B Biochem. Mol. Biol. 2010, 155, 132–137. [Google Scholar] [CrossRef] [PubMed]
- Sahena, F.; Zaidul, I.; Jinap, S.; Jahurul, M.; Khatib, A.; Norulaini, N. Extraction of Fish Oil from the Skin of Indian Mackerel Using Supercritical Fluids. J. Food Eng. 2010, 99, 63–69. [Google Scholar] [CrossRef]
- Zuta, C.P.; Simpson, B.K.; Chan, H.M.; Phillips, L. Concentrating Pufa from Mackerel Processing Waste. J. Am. Oil Chem. Soc. 2003, 80, 933–936. [Google Scholar] [CrossRef]
- Morse, N. Lipid-Lowering and Anti-Inflammatory Effects of Palmitoleic Acid: Evidence from Preclinical and Epidemiological Studies. Lipid Technol. 2015, 27, 107–111. [Google Scholar] [CrossRef]
- Souza, C.O.; Teixeira, A.A.; Biondo, L.A.; Silveira, L.S.; Calder, P.C.; Rosa Neto, J.C. Palmitoleic Acid Reduces the Inflammation in Lps-Stimulated Macrophages by Inhibition of NFΚB, Independently of PPARs. Clin. Exp. Pharmacol. Physiol. 2017, 44, 566–575. [Google Scholar] [CrossRef] [Green Version]
- Weimann, E.; Silva, M.B.B.; Murata, G.M.; Bortolon, J.R.; Dermargos, A.; Curi, R.; Hatanaka, E. Topical Anti-Inflammatory Activity of Palmitoleic Acid Improves Wound Healing. PLoS ONE 2018, 13, e0205338. [Google Scholar] [CrossRef]
- Chajès, V.; Thiébaut, A.C.; Rotival, M.; Gauthier, E.; Maillard, V.; Boutron-Ruault, M.-C.; Joulin, V.; Lenoir, G.M.; Clavel-Chapelon, F. Association between Serum Trans-Monounsaturated Fatty Acids and Breast Cancer Risk in the E3n-Epic Study. Am. J. Epidemiol. 2008, 167, 1312–1320. [Google Scholar] [CrossRef] [Green Version]
- Escrich, E.; Solanas, M.; Moral, R.; Grau, L.; Costa, I.; Vela, E.; Escrich, R. Dietary Lipids and Breast Cancer: Scientific Clinical, Anatomopathological and Molecular Evidences. Rev. Esp. De Obes. 2008, 6, 129–138. [Google Scholar]
- Martin-Moreno, J.M.; Willett, W.C.; Gorgojo, L.; Banegas, J.R.; Rodriguez-Artalejo, F.; Fernandez-Rodriguez, J.C.; Maisonneuve, P.; Boyle, P. Dietary Fat, Olive Oil Intake and Breast Cancer Risk. Int. J. Cancer 1994, 58, 774–780. [Google Scholar] [CrossRef] [PubMed]
- Macquart-Moulin, G.; Riboli, E.; Cornée, J.; Charnay, B.; Berthezene, P.; Day, N. Case-Control Study on Colorectal Cancer and Diet in Marseilles. Int. J. Cancer 1986, 38, 183–191. [Google Scholar] [CrossRef] [PubMed]
- Stoneham, M.; Goldacre, M.; Seagroatt, V.; Gill, L. Olive Oil, Diet and Colorectal Cancer: An Ecological Study and a Hypothesis. J. Epidemiol. Community Health 2000, 54, 756–760. [Google Scholar] [CrossRef] [Green Version]
- Carrillo Pérez, C.; Cavia Camarero, M.d.M.; Alonso de la Torre, S. Antitumor Effect of Oleic Acid; Mechanisms of Action. A Review. Nutr. Hosp. 2012, 27, 1860–1865. [Google Scholar]
- Carrillo Pérez, C.; Cavia Camarero, M.d.M.; Alonso de la Torre, S. Role of Oleic Acid in Immune System; Mechanism of Action; a Review. Nutr. Hosp. 2012, 27, 978–990. [Google Scholar]
- Teres, S.; Barceló-Coblijn, G.; Benet, M.; Alvarez, R.; Bressani, R.; Halver, J.E.; Escriba, P. Oleic Acid Content Is Responsible for the Reduction in Blood Pressure Induced by Olive Oil. Proc. Natl. Acad. Sci. USA 2008, 105, 13811–13816. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dunstan, J.A.; Mitoulas, L.R.; Dixon, G.; Doherty, D.A.; Hartmann, P.E.; Simmer, K.; Prescott, S.L. The Effects of Fish Oil Supplementation in Pregnancy on Breast Milk Fatty Acid Composition over the Course of Lactation: A Randomized Controlled Trial. Pediatric Res. 2007, 62, 689–694. [Google Scholar] [CrossRef] [Green Version]
- Ramakrishnan, U.; Stein, A.D.; Parra-Cabrera, S.; Wang, M.; Imhoff-Kunsch, B.; Juárez-Márquez, S.; Rivera, J.; Martorell, R. Effects of Docosahexaenoic Acid Supplementation During Pregnancy on Gestational Age and Size at Birth: Randomized, Double-Blind, Placebo-Controlled Trial in Mexico. Food Nutr. Bull. 2010, 31, S108–S116. [Google Scholar] [CrossRef]
- Bloomer, R.; Larson, D.; Galpin, A.; Fisher-Wellman, K.; Schilling, B. Effect of Eicosapentaenoic and Docosahexaenoic Acid on Resting and Exercise-Induced Inflammation and Oxidative Stress. J. Int. Soc. Sports Nutr. 2009, 6, 1. [Google Scholar] [CrossRef] [Green Version]
- Ebrahimi, M.; Ghayour-Mobarhan, M.; Rezaiean, S.; Hoseini, M.; Parizade, S.M.R.; Farhoudi, F.; Hosseininezhad, S.J.; Tavallaei, S.; Vejdani, A.; Azimi-Nezhad, M. Omega-3 Fatty Acid Supplements Improve the Cardiovascular Risk Profile of Subjects with Metabolic Syndrome, Including Markers of Inflammation and Auto-Immunity. Acta Cardiol. 2009, 64, 321–327. [Google Scholar] [CrossRef] [Green Version]
- Cottin, S.; Sanders, T.; Hall, W. The Differential Effects of Epa and Dha on Cardiovascular Risk Factors. Proc. Nutr. Soc. 2011, 70, 215–231. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Swanson, D.; Block, R.; Mousa, S.A. Omega-3 Fatty Acids Epa and Dha: Health Benefits Throughout Life. Adv. Nutr. 2012, 3, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Subedi, L.; Lee, T.H.; Wahedi, H.M.; Baek, S.-H.; Kim, S.Y. Resveratrol-Enriched Rice Attenuates Uvb-Ros-Induced Skin Aging Via Downregulation of Inflammatory Cascades. Oxidative Med. Cell. Longev. 2017, 2017, 8379539. [Google Scholar] [CrossRef]
- Hwang, E.; Kim, S.H.; Lee, S.; Lee, C.H.; Do, S.G.; Kim, J.; Kim, S.Y. A comparative study of baby immature and adult shoots of Aloe vera on UVB-induced skin photoaging in vitro. Phyther. Res. 2013, 27, 1874–1882. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
{kind=link}
| Species | Part | SFA | MUFA | PUFA | Cholesterol | |
|---|---|---|---|---|---|---|
| Omega-3 | Other PUFA | |||||
| NR | Brain | 20.04 | 30.86 | 32.21 | 0.64 | 16.25 |
| Liver | 27.10 | 47.02 | 21.81 | 4.16 | 0.00 | |
| Stomach | 28.51 | 29.89 | 30.76 | 10.85 | 0.00 | |
| Skin | 20.35 | 36.76 | 30.99 | 5.99 | 5.89 | |
| CG | Brain | 25.95 | 35.01 | 0.00 | 0.00 | 39.04 |
| Liver | 34.48 | 48.16 | 1.34 | 0.95 | 15.07 | |
| Stomach | 31.44 | 32.94 | 32.66 | 2.96 | 0.00 | |
| Skin | 29.96 | 42.21 | 25.90 | 1.93 | 0.00 | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lee, S.; Koo, M.H.; Han, D.-W.; Kim, I.-C.; Lee, J.H.; Kim, J.-H.; Sultana, R.; Kim, S.Y.; Youn, U.J.; Kim, J.-H. Comparison of Fatty Acid Contents and MMP-1 Inhibitory Effects of the Two Antarctic Fish, Notothenia rossii and Champsocephalus gunnari. Molecules 2022, 27, 4554. https://doi.org/10.3390/molecules27144554
Lee S, Koo MH, Han D-W, Kim I-C, Lee JH, Kim J-H, Sultana R, Kim SY, Youn UJ, Kim J-H. Comparison of Fatty Acid Contents and MMP-1 Inhibitory Effects of the Two Antarctic Fish, Notothenia rossii and Champsocephalus gunnari. Molecules. 2022; 27(14):4554. https://doi.org/10.3390/molecules27144554
Chicago/Turabian StyleLee, Seulah, Man Hyung Koo, Dong-Won Han, Il-Chan Kim, Jun Hyuck Lee, Jeong-Hoon Kim, Razia Sultana, Sun Yeou Kim, Ui Joung Youn, and Jin-Hyoung Kim. 2022. "Comparison of Fatty Acid Contents and MMP-1 Inhibitory Effects of the Two Antarctic Fish, Notothenia rossii and Champsocephalus gunnari" Molecules 27, no. 14: 4554. https://doi.org/10.3390/molecules27144554
APA StyleLee, S., Koo, M. H., Han, D.-W., Kim, I.-C., Lee, J. H., Kim, J.-H., Sultana, R., Kim, S. Y., Youn, U. J., & Kim, J.-H. (2022). Comparison of Fatty Acid Contents and MMP-1 Inhibitory Effects of the Two Antarctic Fish, Notothenia rossii and Champsocephalus gunnari. Molecules, 27(14), 4554. https://doi.org/10.3390/molecules27144554