
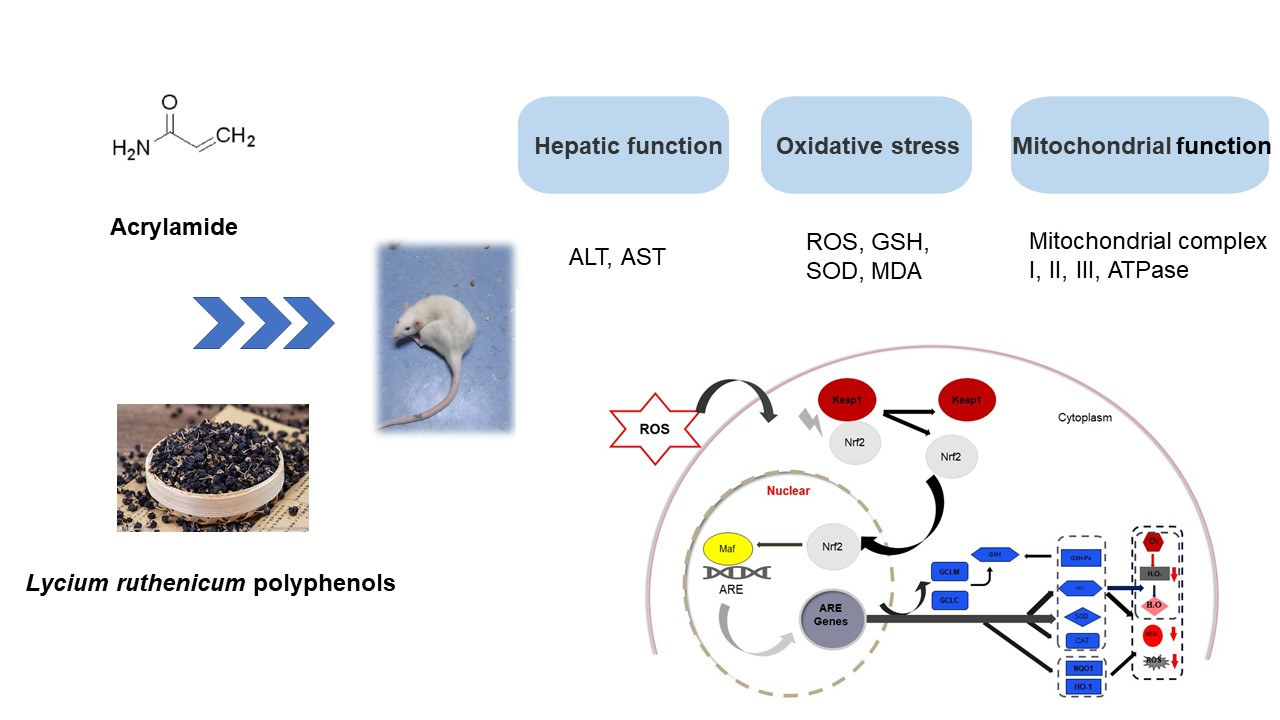
Protective Effect of Lycium ruthenicum Polyphenols on Oxidative Stress against Acrylamide Induced Liver Injury in Rats


Abstract
:
1. Introduction
2. Results
2.1. Effect of LRP on the ACR-Induced Serum Levels of ALT and AST
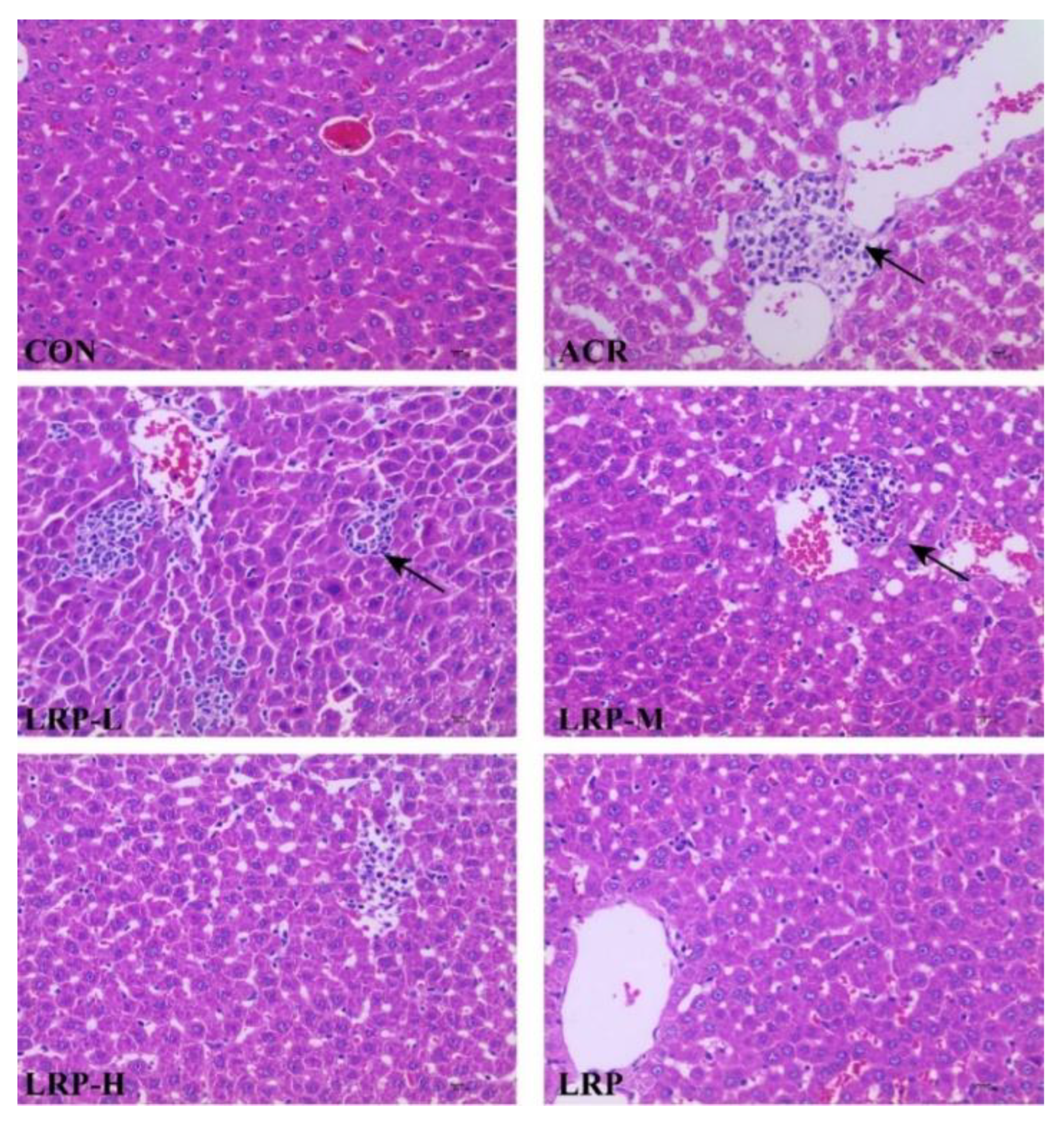
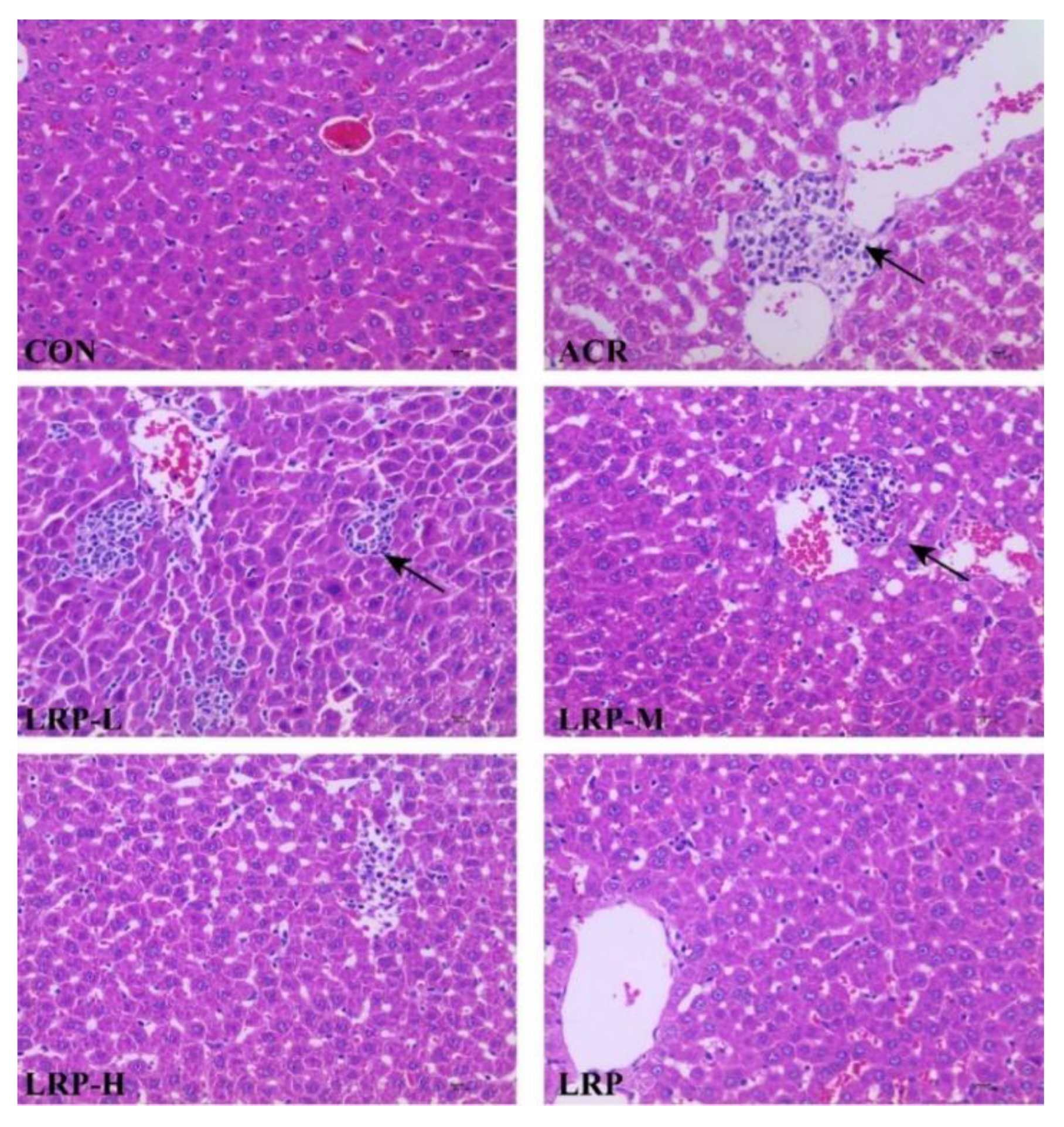
2.2. Effect of LRP on the ACR-Induced Histopathological Changes
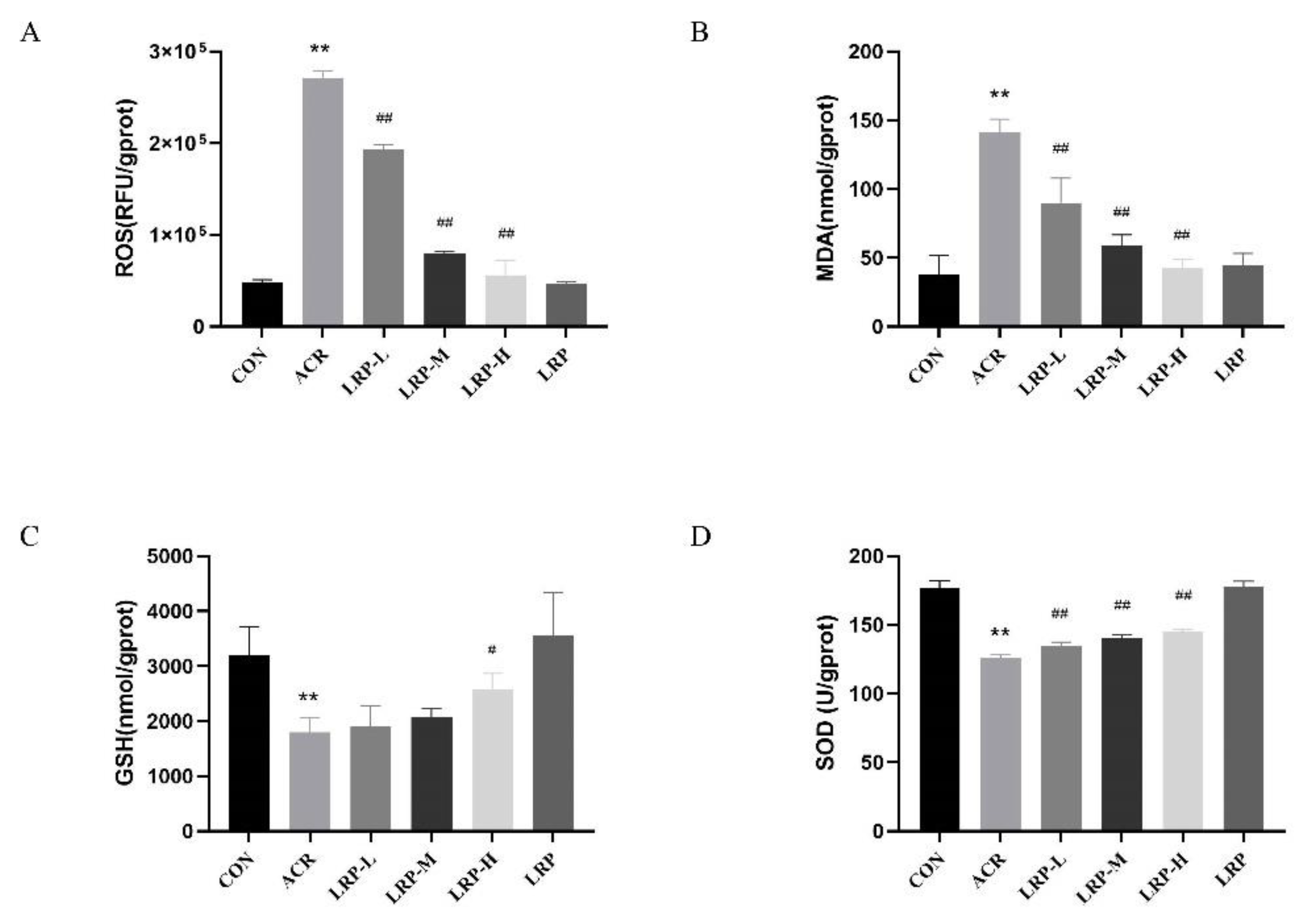
2.3. Effects of LRP on ROS in the Liver
2.4. Effects of LRP on SOD, GSH, and MDA in the Liver
2.5. Effect of LRP on ATPase Activities Induced by ACR in the Liver Mitochondrion
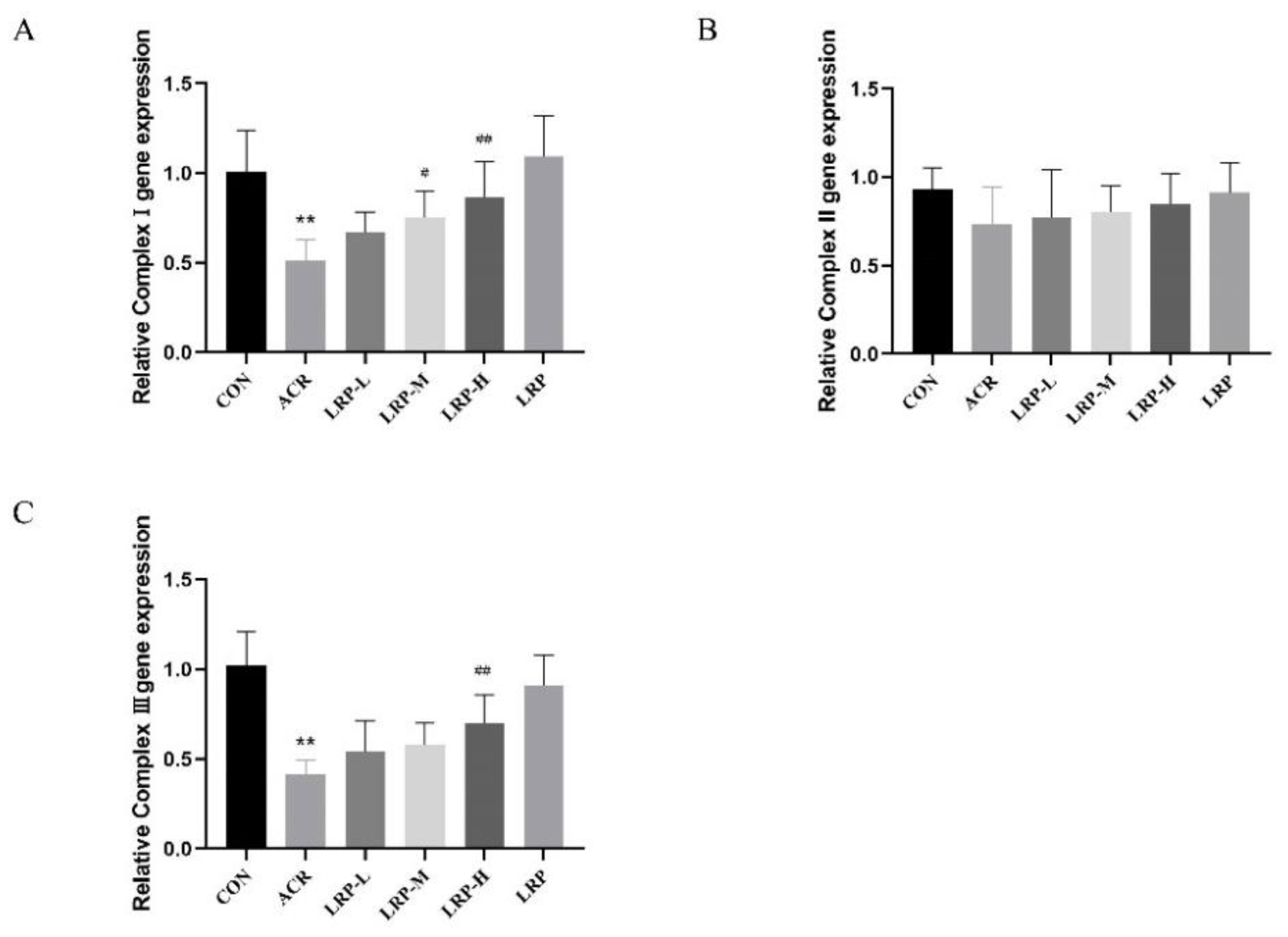
2.6. Effect of LRP on the mRNA Expression of Mitochondrial Complexes I–III Induced by ACR in Liver Tissue
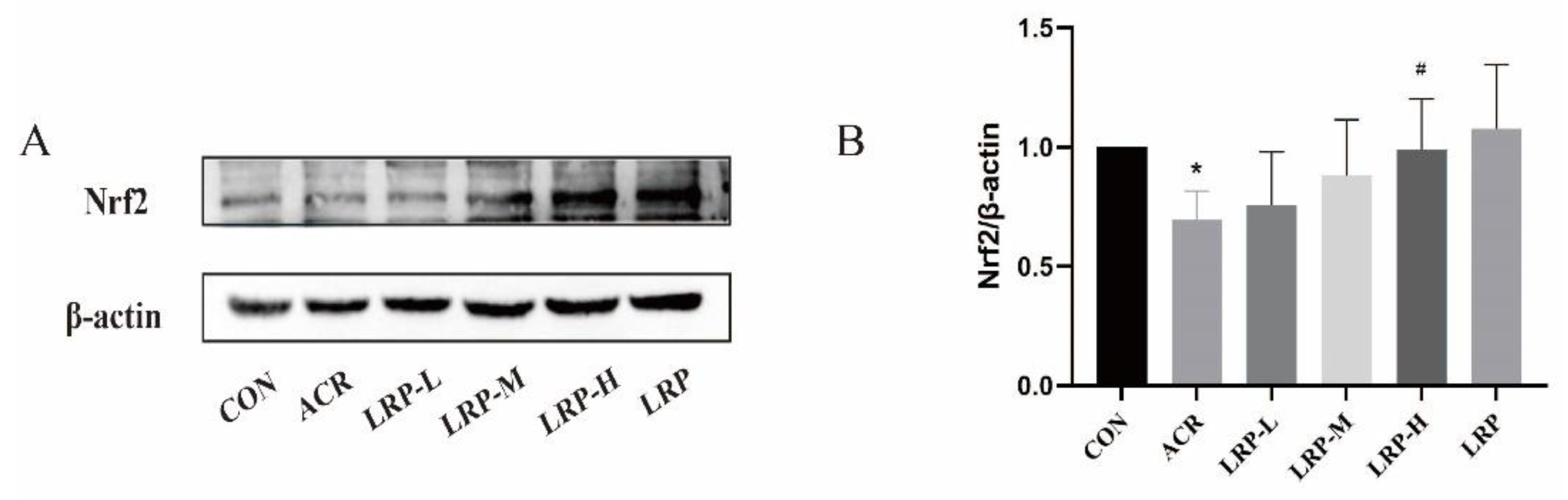
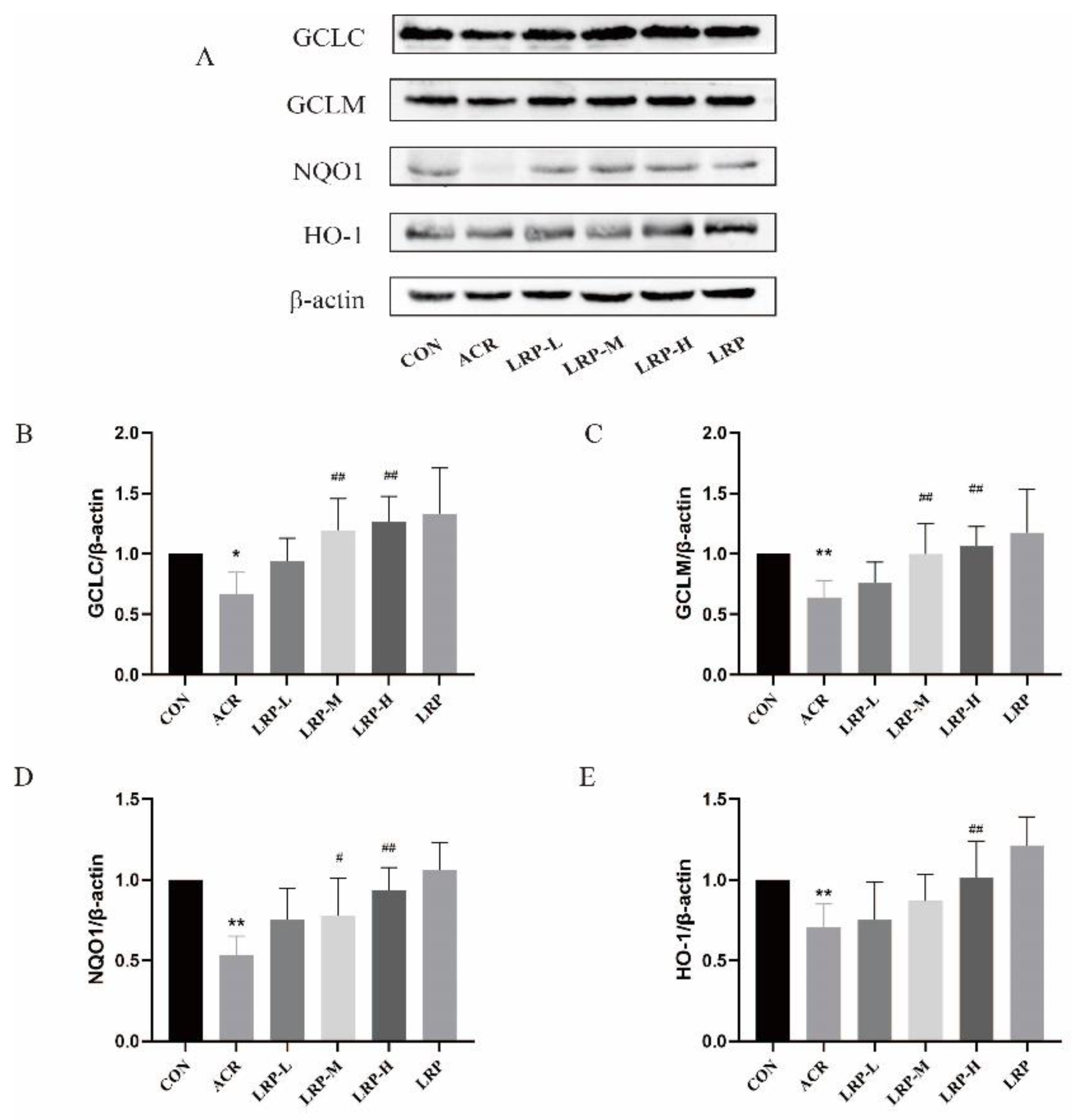
2.7. Effect of LRP on the Nrf2 Pathway Induced by ACR in Liver Tissue
3. Discussion
4. Materials and Methods
4.1. Chemicals and Reagents
4.2. Animals and Experimental Design
4.3. Sample Collection
4.4. Isolation of Liver Mitochondria
4.5. Histopathological Studies
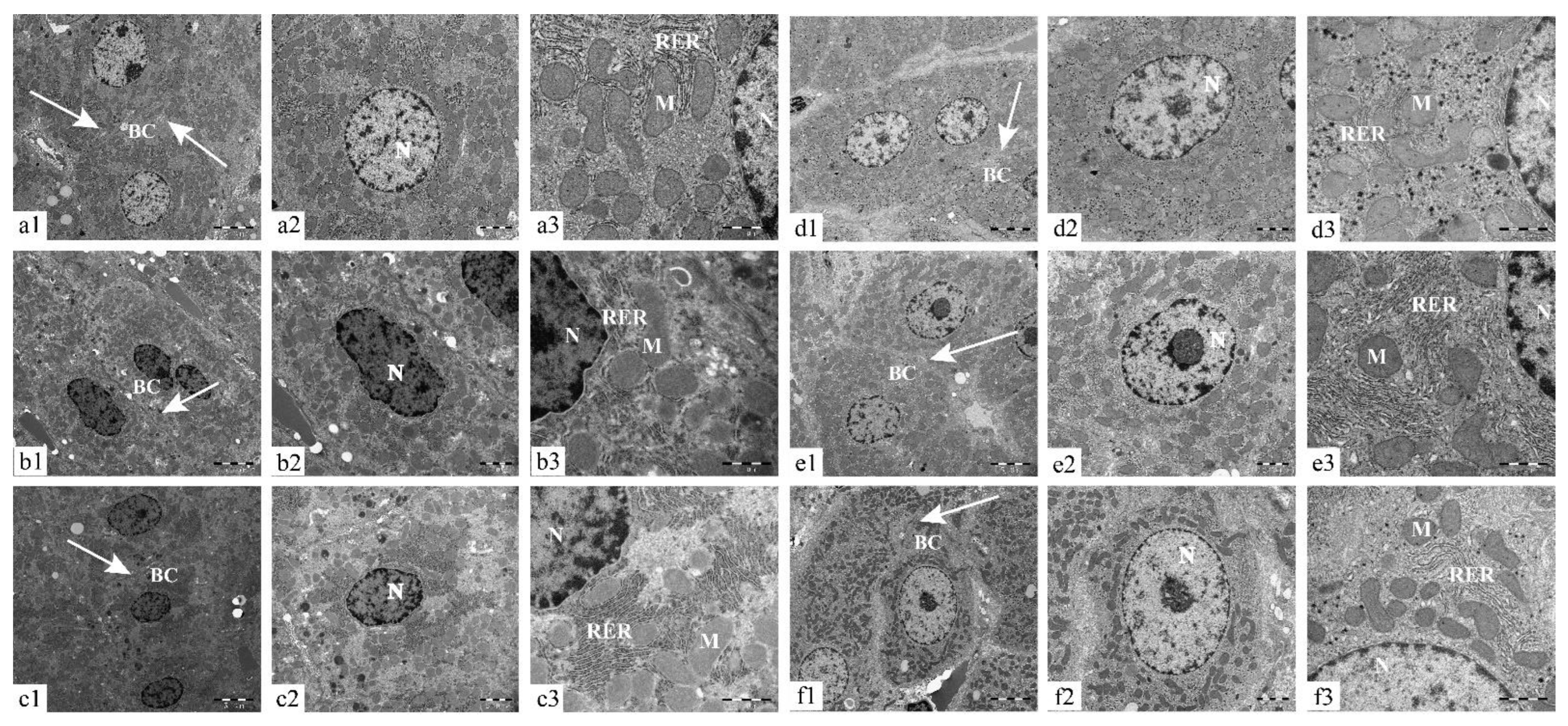
4.6. Transmission Electron Microscopy
4.7. Measurement of the AST, ALT Indexes in Serum
4.8. Assessment of Oxidative Stress in the Liver
4.9. Measurement of the Liver Mitochondrial ATPase
4.10. Total RNA Extraction, Reverse Transcription, and Quantitative Real-Time Polymerase Chain Reaction
4.11. Western Blot Analysis
4.12. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Sample Availability
References
- Semla, M.; Goc, Z.; Martiniakova, M.; Omelka, R.; Formicki, G. Acrylamide: A common food toxin related to physiological functions and health. Physiol. Res. 2017, 66, 205–217. [Google Scholar] [CrossRef] [PubMed]
- Goudarzi, M.; Mombeini, M.A.; Fatemi, I.; Aminzadeh, A.; Kalantari, H.; Nesari, A.; Najafzadehvarzi, H.; Mehrzadi, S. Neuroprotective effects of Ellagic acid against acrylamide-induced neurotoxicity in rats. Neurol. Res. 2019, 41, 419–428. [Google Scholar] [CrossRef]
- Abdel-Daim, M.M.; Abo El-Ela, F.I.; Alshahrani, F.K.; Bin-Jumah, M.; Al-Zharani, M.; Almutairi, B.; Alyousif, M.S.; Bungau, S.; Aleya, L.; Alkahtani, S. Protective effects of thymoquinone against acrylamide-induced liver, kidney and brain oxidative damage in rats. Environ. Sci. Pollut. Res. Int. 2020, 27, 37709–37717. [Google Scholar] [CrossRef] [PubMed]
- Belhadj Benziane, A.; Dilmi Bouras, A.; Mezaini, A.; Belhadri, A.; Benali, M. Effect of oral exposure to acrylamide on biochemical and hematologic parameters in Wistar rats. Drug Chem. Toxicol. 2019, 42, 157–166. [Google Scholar] [CrossRef]
- Friedman, M. Acrylamide: Inhibition of formation in processed food and mitigation of toxicity in cells, animals, and humans. Food Funct. 2015, 6, 1752–1772. [Google Scholar] [CrossRef]
- Rifai, L.; Saleh, F.A. A Review on Acrylamide in Food: Occurrence, Toxicity, and Mitigation Strategies. Int. J. Toxicol. 2020, 39, 93–102. [Google Scholar] [CrossRef]
- Kunnel, S.G.; Subramanya, S.; Satapathy, P.; Sahoo, I.; Zameer, F. Acrylamide Induced Toxicity and the Propensity of Phytochemicals in Amelioration: A Review. Cent. Nerv. Syst. Agents Med. Chem. 2019, 19, 100–113. [Google Scholar] [CrossRef] [PubMed]
- Uthra, C.; Shrivastava, S.; Jaswal, A.; Sinha, N.; Reshi, M.S.; Shukla, S. Therapeutic potential of quercetin against acrylamide induced toxicity in rats. Biomed. Pharmacother. 2017, 86, 705–714. [Google Scholar] [CrossRef] [PubMed]
- Mottram, D.S.; Wedzicha, B.L.; Dodson, A.T. Acrylamide is formed in the Maillard reaction. Nature 2002, 419, 448–449. [Google Scholar] [CrossRef] [PubMed]
- Zhou, P.P.; Zhao, Y.F.; Liu, H.L.; Ma, Y.J.; Li, X.W.; Yang, X.; Wu, Y.N. Dietary exposure of the Chinese population to acrylamide. Biomed. Environ. Sci. 2013, 26, 421–429. [Google Scholar] [CrossRef]
- Abt, E.; Robin, L.P.; McGrath, S.; Srinivasan, J.; DiNovi, M.; Adachi, Y.; Chirtel, S. Acrylamide levels and dietary exposure from foods in the United States, an update based on 2011-2015 data. Food Addit. Contam.Part. A Chem. Anal. Control. Expo. Risk Assess. 2019, 36, 1475–1490. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.; Kim, H.J. Dietary Exposure to Acrylamide and Associated Health Risks for the Korean Population. Int. J. Environ. Res. Public Health 2020, 17, 7619. [Google Scholar] [CrossRef] [PubMed]
- Pietropaoli, F.; Pantalone, S.; Cichelli, A.; d’Alessandro, N. Acrylamide in widely consumed foods-a review. Food Addit. Contam. Part. A Chem. Anal. Control. Expo. Risk Assess. 2022, 39, 1–35. [Google Scholar] [CrossRef] [PubMed]
- Foroutanfar, A.; Mehri, S.; Kamyar, M.; Tandisehpanah, Z.; Hosseinzadeh, H. Protective effect of punicalagin, the main polyphenol compound of pomegranate, against acrylamide-induced neurotoxicity and hepatotoxicity in rats. Phytother. Res. 2020, 34, 3262–3272. [Google Scholar] [CrossRef]
- Liu, Y.; Wang, R.; Zheng, K.; Xin, Y.; Jia, S.; Zhao, X. Metabonomics analysis of liver in rats administered with chronic low-dose acrylamide. Xenobiotica 2020, 50, 894–905. [Google Scholar] [CrossRef] [PubMed]
- Hou, L.; Liu, S.; Zhao, C.; Fan, L.; Hu, H.; Yin, S. The combination of T-2 toxin and acrylamide synergistically induces hepatotoxicity and nephrotoxicity via the activation of oxidative stress and the mitochondrial pathway. Toxicon 2021, 189, 65–72. [Google Scholar] [CrossRef] [PubMed]
- Altinoz, E.; Turkoz, Y.; Vardi, N. The protective effect of N-acetylcysteine against acrylamide toxicity in liver and small and large intestine tissues. Bratisl. Lek. Listy 2015, 116, 252–258. [Google Scholar] [CrossRef] [PubMed]
- Acaroz, U.; Ince, S.; Arslan-Acaroz, D.; Gurler, Z.; Kucukkurt, I.; Demirel, H.H.; Arslan, H.O.; Varol, N.; Zhu, K. The ameliorative effects of boron against acrylamide-induced oxidative stress, inflammatory response, and metabolic changes in rats. Food Chem. Toxicol. 2018, 118, 745–752. [Google Scholar] [CrossRef] [PubMed]
- Pan, X.; Zhu, L.; Lu, H.; Wang, D.; Lu, Q.; Yan, H. Melatonin Attenuates Oxidative Damage Induced by Acrylamide In Vitro and In Vivo. Oxid. Med. Cell. Longev. 2015, 2015, 703709. [Google Scholar] [CrossRef]
- Bin-Jumah, M.N.; Al-Huqail, A.A.; Abdelnaeim, N.; Kamel, M.; Fouda, M.M.A.; Abulmeaty, M.M.A.; Saadeldin, I.M.; Abdel-Daim, M.M. Potential protective effects of Spirulina platensis on liver, kidney, and brain acrylamide toxicity in rats. Environ. Sci. Pollut. Res. Int. 2021, 28, 26653–26663. [Google Scholar] [CrossRef] [PubMed]
- Seydi, E.; Rajabi, M.; Salimi, A.; Pourahmad, J. Involvement of mitochondrial-mediated caspase-3 activation and lysosomal labilization in acrylamide-induced liver toxicity. Toxicol. Environ. Chem. 2015, 97, 563–575. [Google Scholar] [CrossRef]
- Chen, J.; Chen, Y.; Zheng, Y.; Zhao, J.; Yu, H.; Zhu, J.; Li, D. Protective Effects and Mechanisms of Procyanidins on Parkinson’s Disease In Vivo and In Vitro. Molecules 2021, 26, 5558. [Google Scholar] [CrossRef] [PubMed]
- Kacar, S.; Sahinturk, V. The Protective Agents Used against Acrylamide Toxicity: An In Vitro Cell Culture Study-Based Review. Cell J. 2021, 23, 367–381. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Li, J.; Tao, W.; Zhang, X.; Gao, X.; Yong, J.; Zhao, J.; Zhang, L.; Li, Y.; Duan, J.A. Lycium ruthenicum studies: Molecular biology, Phytochemistry and pharmacology. Food Chem. 2018, 240, 759–766. [Google Scholar] [CrossRef]
- Gao, H.; Yuan, X.; Wang, Z.; Gao, Q.; Yang, J. Profiles and neuroprotective effects of Lycium ruthenicum polyphenols against oxidative stress-induced cytotoxicity in PC12 cells. J. Food Biochem. 2020, 44, e13112. [Google Scholar] [CrossRef]
- Chen, S.; Zhou, H.; Zhang, G.; Meng, J.; Deng, K.; Zhou, W.; Wang, H.; Wang, Z.; Hu, N.; Suo, Y. Anthocyanins from Lycium ruthenicum Murr. Ameliorated d-Galactose-Induced Memory Impairment, Oxidative Stress, and Neuroinflammation in Adult Rats. J. Agric. Food Chem. 2019, 67, 3140–3149. [Google Scholar] [CrossRef]
- Liu, Y.; Liu, M.; Chen, Q.; Liu, G.M.; Cao, M.J.; Sun, L.; Lu, Z.; Guo, C. Blueberry Polyphenols Ameliorate Visible Light and Lipid-Induced Injury of Retinal Pigment Epithelial Cells. J. Agric. Food Chem. 2018, 66, 12730–12740. [Google Scholar] [CrossRef]
- Lu, K.; Wang, J.; Yu, Y.; Wu, Y.; He, Z. Lycium ruthenicum Murr. alleviates nonalcoholic fatty liver in mice. Food Sci. Nutr. 2020, 8, 2588–2597. [Google Scholar] [CrossRef] [Green Version]
- Jiang, G.; Lei, A.; Chen, Y.; Yu, Q.; Xie, J.; Yang, Y.; Yuan, T.; Su, D. The protective effects of the Ganoderma atrum polysaccharide against acrylamide-induced inflammation and oxidative damage in rats. Food Funct. 2021, 12, 397–407. [Google Scholar] [CrossRef]
- Ibrahim, M.A.; Ibrahem, M.D. Acrylamide-induced hematotoxicity, oxidative stress, and DNA damage in liver, kidney, and brain of catfish (Clarias gariepinus). Environ. Toxicol. 2020, 35, 300–308. [Google Scholar] [CrossRef]
- Zhang, L.; Wang, E.; Chen, F.; Yan, H.; Yuan, Y. Potential protective effects of oral administration of allicin on acrylamide-induced toxicity in male mice. Food Funct. 2013, 4, 1229–1236. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.W.; Liu, J.; Wei, S.S.; Zhao, W.P.; Zhu, S.Q.; Zhou, B.H. Mitochondrial respiratory chain damage and mitochondrial fusion disorder are involved in liver dysfunction of fluoride-induced mice. Chemosphere 2020, 241, 125099. [Google Scholar] [CrossRef] [PubMed]
- Kianfar, M.; Nezami, A.; Mehri, S.; Hosseinzadeh, H.; Hayes, A.W.; Karimi, G. The protective effect of fasudil against acrylamide-induced cytotoxicity in PC12 cells. Drug Chem. Toxicol. 2020, 43, 595–601. [Google Scholar] [CrossRef] [PubMed]
- Liu, Z.; Tang, X.; Liu, C.; Dong, B.; Shao, Y.; Liu, B.; Yue, H. Ultrasonic extraction of anthocyanins from Lycium ruthenicum Murr. and its antioxidant activity. Food Sci. Nutr. 2020, 8, 2642–2651. [Google Scholar] [CrossRef]
- Tang, J.; Yan, Y.; Ran, L.; Mi, J.; Sun, Y.; Lu, L.; Gao, Y.; Zeng, X.; Cao, Y. Isolation, antioxidant property and protective effect on PC12 cell of the main anthocyanin in fruit of Lycium ruthenicum Murray. J. Funct. Foods 2017, 30, 97–107. [Google Scholar] [CrossRef]
- Tian, B.; Zhao, J.; Xie, X.; Chen, T.; Yin, Y.; Zhai, R.; Wang, X.; An, W.; Li, J. Anthocyanins from the fruits of Lycium ruthenicum Murray improve high-fat diet-induced insulin resistance by ameliorating inflammation and oxidative stress in mice. Food Funct. 2021, 12, 3855–3871. [Google Scholar] [CrossRef]
- Er, R.; Aydin, B.; Sekeroglu, V.; Atli Sekeroglu, Z. Protective effect of Argan oil on mitochondrial function and oxidative stress against acrylamide-induced liver and kidney injury in rats. Biomarkers 2020, 25, 458–467. [Google Scholar] [CrossRef]
- Zhao, M.; Wang, P.; Zhu, Y.; Liu, X.; Hu, X.; Chen, F. The chemoprotection of a blueberry anthocyanin extract against the acrylamide-induced oxidative stress in mitochondria: Unequivocal evidence in mice liver. Food Funct. 2015, 6, 3006–3012. [Google Scholar] [CrossRef]
- Prakash, C.; Kumar, V. Chronic Arsenic Exposure-Induced Oxidative Stress is Mediated by Decreased Mitochondrial Biogenesis in Rat Liver. Biol. Trace Elem. Res. 2016, 173, 87–95. [Google Scholar] [CrossRef]
- Zhao, M.; Lewis Wang, F.S.; Hu, X.; Chen, F.; Chan, H.M. Acrylamide-induced neurotoxicity in primary astrocytes and microglia: Roles of the Nrf2-ARE and NF-kappaB pathways. Food Chem. Toxicol. 2017, 106, 25–35. [Google Scholar] [CrossRef]
- Pan, X.; Wu, X.; Yan, D.; Peng, C.; Rao, C.; Yan, H. Acrylamide-induced oxidative stress and inflammatory response are alleviated by N-acetylcysteine in PC12 cells: Involvement of the crosstalk between Nrf2 and NF-kappaB pathways regulated by MAPKs. Toxicol. Lett. 2018, 288, 55–64. [Google Scholar] [CrossRef] [PubMed]
- Zhang, M.; Pan, H.; Xu, Y.; Wang, X.; Qiu, Z.; Jiang, L. Allicin Decreases Lipopolysaccharide-Induced Oxidative Stress and Inflammation in Human Umbilical Vein Endothelial Cells through Suppression of Mitochondrial Dysfunction and Activation of Nrf2. Cell Physiol. Biochem. 2017, 41, 2255–2267. [Google Scholar] [CrossRef]
- Liu, D.; Wang, H.; Zhang, Y.; Zhang, Z. Protective Effects of Chlorogenic Acid on Cerebral Ischemia/Reperfusion Injury Rats by Regulating Oxidative Stress-Related Nrf2 Pathway. Drug Des. Devel. Ther. 2020, 14, 51–60. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tan, X.; Li, L.; Wang, J.; Zhao, B.; Pan, J.; Wang, L.; Liu, X.; Liu, X.; Liu, Z. Resveratrol Prevents Acrylamide-Induced Mitochondrial Dysfunction and Inflammatory Responses via Targeting Circadian Regulator Bmal1 and Cry1 in Hepatocytes. J. Agric. Food Chem. 2019, 67, 8510–8519. [Google Scholar] [CrossRef] [PubMed]
- Ali, B.H.; Cahlikova, L.; Opletal, L.; Karaca, T.; Manoj, P.; Ramkumar, A.; Al Suleimani, Y.M.; Al Za’abi, M.; Nemmar, A.; Chocholousova-Havlikova, L.; et al. Effect of aqueous extract and anthocyanins of calyces of Hibiscus sabdariffa (Malvaceae) in rats with adenine-induced chronic kidney disease. J. Pharm. Pharmacol. 2017, 69, 1219–1229. [Google Scholar] [CrossRef]
- Wang, J.; He, J.; Fan, Y.; Xu, F.; Liu, Q.; He, R.; Yan, R. Extensive mitochondrial proteome disturbance occurs during the early stages of acute myocardial ischemia. Exp. Ther. Med. 2022, 23, 85. [Google Scholar] [CrossRef] [PubMed]
- Donmez, D.B.; Kacar, S.; Bagci, R.; Sahinturk, V. Protective effect of carnosic acid on acrylamide-induced liver toxicity in rats: Mechanistic approach over Nrf2-Keap1 pathway. J. Biochem. Mol. Toxicol 2020, 34, e22524. [Google Scholar] [CrossRef]
- Ghasemzadeh Rahbardar, M.; Cheraghi Farmad, H.; Hosseinzadeh, H.; Mehri, S. Protective effects of selenium on acrylamide-induced neurotoxicity and hepatotoxicity in rats. Iran. J. Basic Med. Sci. 2021, 24, 1041–1049. [Google Scholar] [CrossRef]
- Li, Z.; Miao, Z.; Ding, L.; Teng, X.; Bao, J. Energy metabolism disorder mediated ammonia gas-induced autophagy via AMPK/mTOR/ULK1-Beclin1 pathway in chicken livers. Ecotoxicol. Environ. Saf. 2021, 217, 112219. [Google Scholar] [CrossRef]
- Nasouti, R.; Khaksari, M.; Mirzaee, M.; Nazari-Robati, M. Trehalose protects against spinal cord injury through regulating heat shock proteins 27 and 70 and caspase-3 genes expression. J. Basic Clin. Physiol. Pharmacol. 2019, 31, 245–253. [Google Scholar] [CrossRef]
- Pan, X.; Yan, D.; Wang, D.; Wu, X.; Zhao, W.; Lu, Q.; Yan, H. Mitochondrion-Mediated Apoptosis Induced by Acrylamide is Regulated by a Balance Between Nrf2 Antioxidant and MAPK Signaling Pathways in PC12 Cells. Mol. Neurobiol. 2017, 54, 4781–4794. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Groups/ Parameters | CON | ACR | LRP-L | LRP-M | LRP-H | LRP |
|---|---|---|---|---|---|---|
| ALT (U/L) | 60.08 ± 9.22 | 124.18 ± 19.31 ** | 82.03 ± 15.76 # | 92.48 ± 22.17 # | 77.36 ± 26.19 # | 67.48 ± 6.03 |
| AST (U/L) | 100.45 ± 9.81 | 204.60 ± 33.93 ** | 165.19 ± 23.04 # | 160.52 ± 21.71 # | 146.51 ± 22.21 ## | 122.99 ± 10.36 |
| Groups/ Parameters | CON | ACR | LRP-L | LRP-M | LRP-H | LRP |
|---|---|---|---|---|---|---|
| Ca2+-ATPase | 2.28 ± 0.56 | 1.39 ± 0.17 ** | 1.60 ± 0.11 | 1.64 ± 0.16 | 1.90 ± 0.46 # | 2.31 ± 0.54 |
| Na+-K+-ATPase | 2.50 ± 0.49 | 1.40 ± 0.23 ** | 1.57 ± 0.14 | 1.76 ± 0.42 | 1.97 ± 0.45 | 2.48 ± 0.69 |
| Mg2+-ATPase | 2.43 ± 0.50 | 1.40 ± 0.22 ** | 1.61 ± 0.10 | 1.71 ± 0.41 | 1.92 ± 0.45 | 2.39 ± 0.57 |
| Total-ATPase | 7.29 ± 1.55 | 4.26 ± 0.63 ** | 4.78 ± 0.34 | 5.07 ± 0.98 | 5.89 ± 1.48 | 7.35 ± 1.64 |
| Groups | 0–7 d Treatment (i.g.) | 8–19 d Treatment (i.g.) |
|---|---|---|
| CON | normal saline | normal saline |
| ACR | normal saline | normal saline + ACR(40 mg/kg) |
| LRP-L | LRP(50 mg/kg) | LRP(50 mg/kg) + ACR(40 mg/kg) |
| LRP-M | LRP(100 mg/kg) | LRP(100 mg/kg) + ACR(40 mg/kg) |
| LRP-H | LRP(200 mg/kg) | LRP(200 mg/kg) + ACR(40 mg/kg) |
| LRP | LRP(200 mg/kg) | LRP(200 mg/kg) |
| Genes | Forward Primer | Reverse Primer |
|---|---|---|
| COX I | CTGCCCTCTGTACCCAAAGA | GACCCATCTTTCCAGAGGT |
| COX II | CCAGATGGCCAGAGGACTCA | TGTGAGTCCCGAGGGAATAGA |
| COX III | GCCACCACACCCCTATTGTA | TCCCGTTGCTATGAAGAATG |
| GAPDH | TGTTCCTACCCCCAATGTGT | CCCTGTTGCTGTAGCCGTAT |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gao, H.; Xue, Y.; Wu, L.; Huo, J.; Pang, Y.; Chen, J.; Gao, Q. Protective Effect of Lycium ruthenicum Polyphenols on Oxidative Stress against Acrylamide Induced Liver Injury in Rats. Molecules 2022, 27, 4100. https://doi.org/10.3390/molecules27134100
Gao H, Xue Y, Wu L, Huo J, Pang Y, Chen J, Gao Q. Protective Effect of Lycium ruthenicum Polyphenols on Oxidative Stress against Acrylamide Induced Liver Injury in Rats. Molecules. 2022; 27(13):4100. https://doi.org/10.3390/molecules27134100
Chicago/Turabian StyleGao, Hua, Yanzhong Xue, Lingyu Wu, Jinghong Huo, Yufei Pang, Jingxin Chen, and Qinghan Gao. 2022. "Protective Effect of Lycium ruthenicum Polyphenols on Oxidative Stress against Acrylamide Induced Liver Injury in Rats" Molecules 27, no. 13: 4100. https://doi.org/10.3390/molecules27134100
APA StyleGao, H., Xue, Y., Wu, L., Huo, J., Pang, Y., Chen, J., & Gao, Q. (2022). Protective Effect of Lycium ruthenicum Polyphenols on Oxidative Stress against Acrylamide Induced Liver Injury in Rats. Molecules, 27(13), 4100. https://doi.org/10.3390/molecules27134100