
The Role of Saponins in the Treatment of Neuropathic Pain



Abstract
:1. Introduction
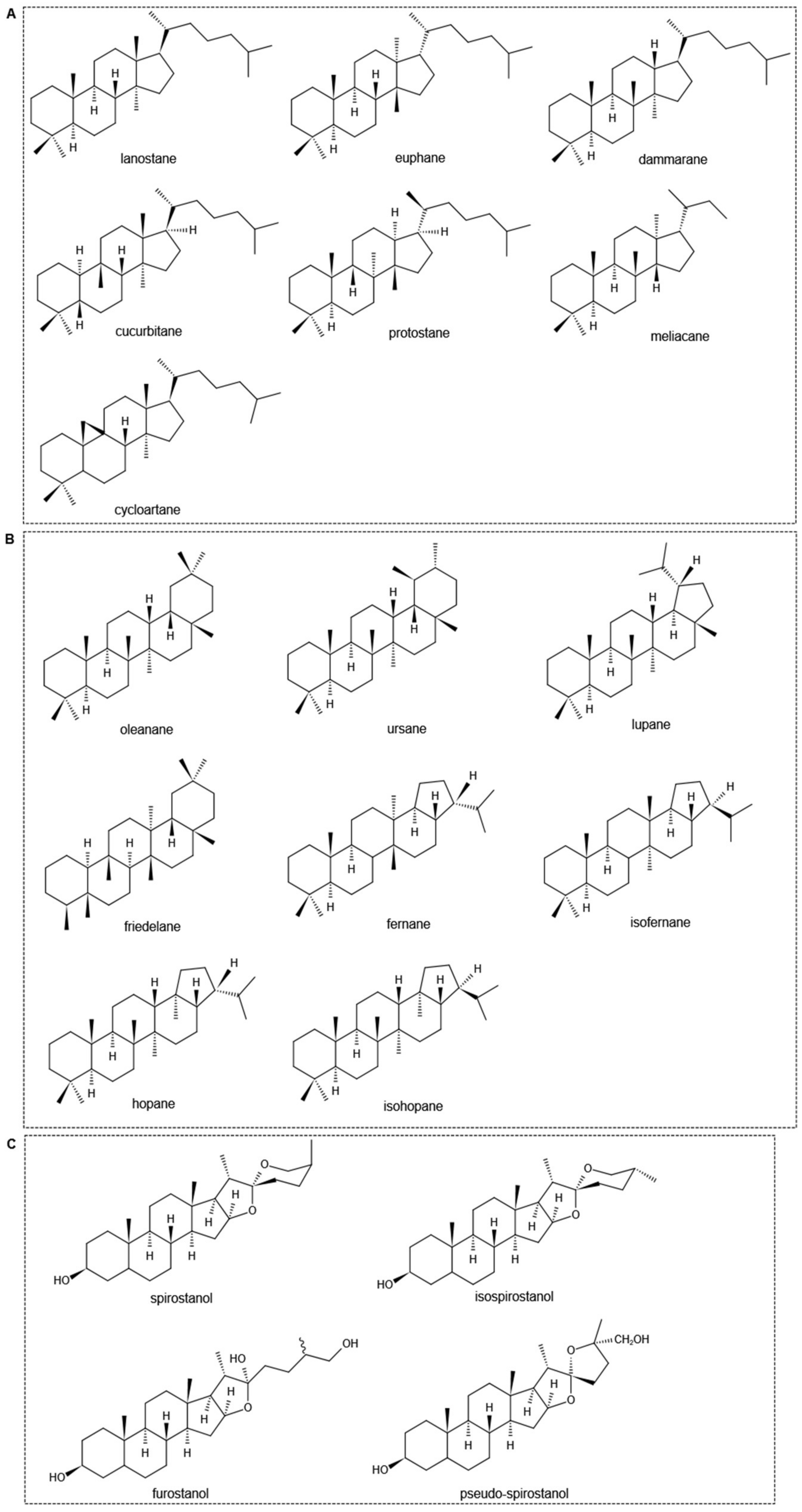
2. The Chemical Properties of Saponins
3. Preclinical Evidence for Saponins in Neuropathic Pain
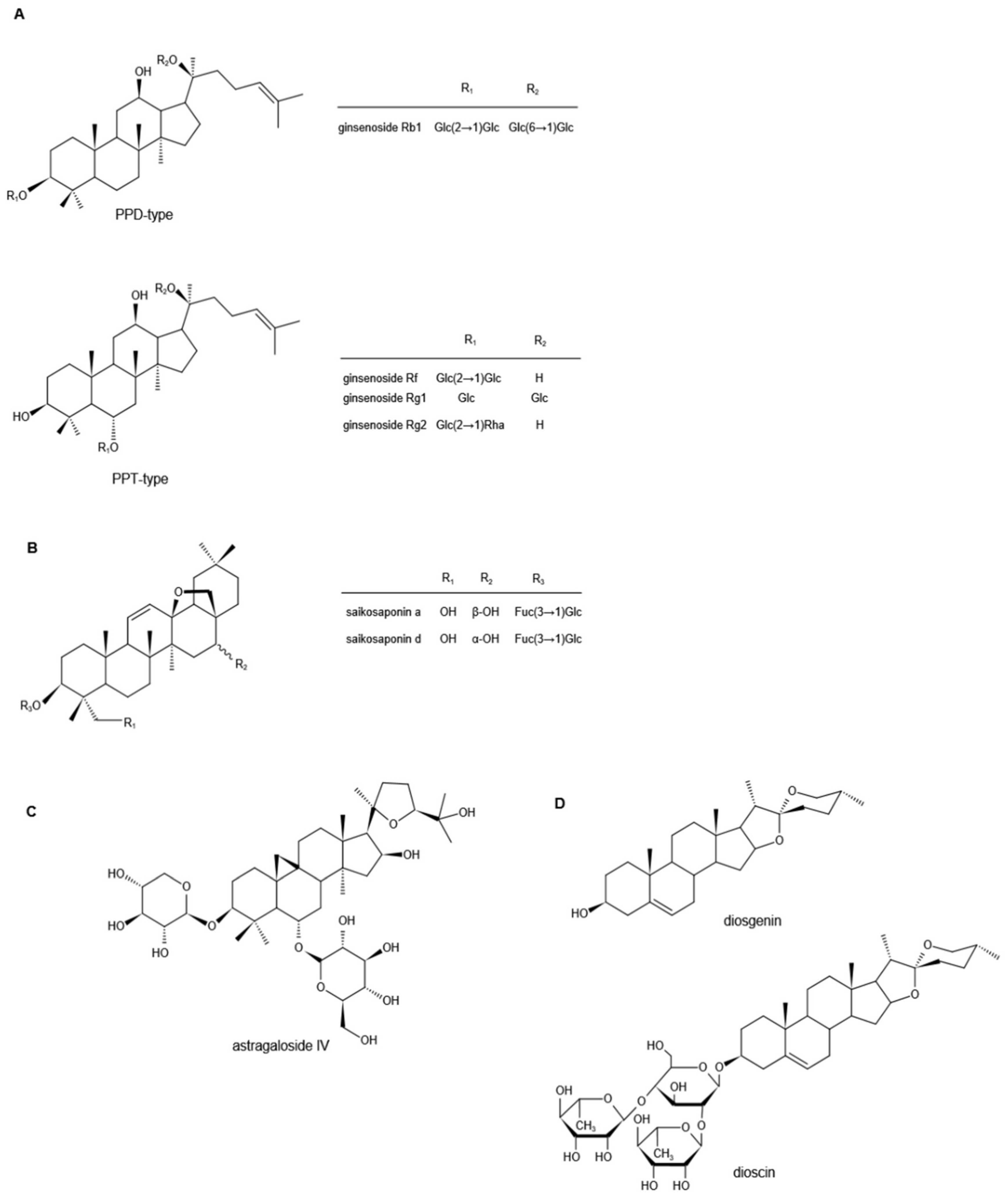
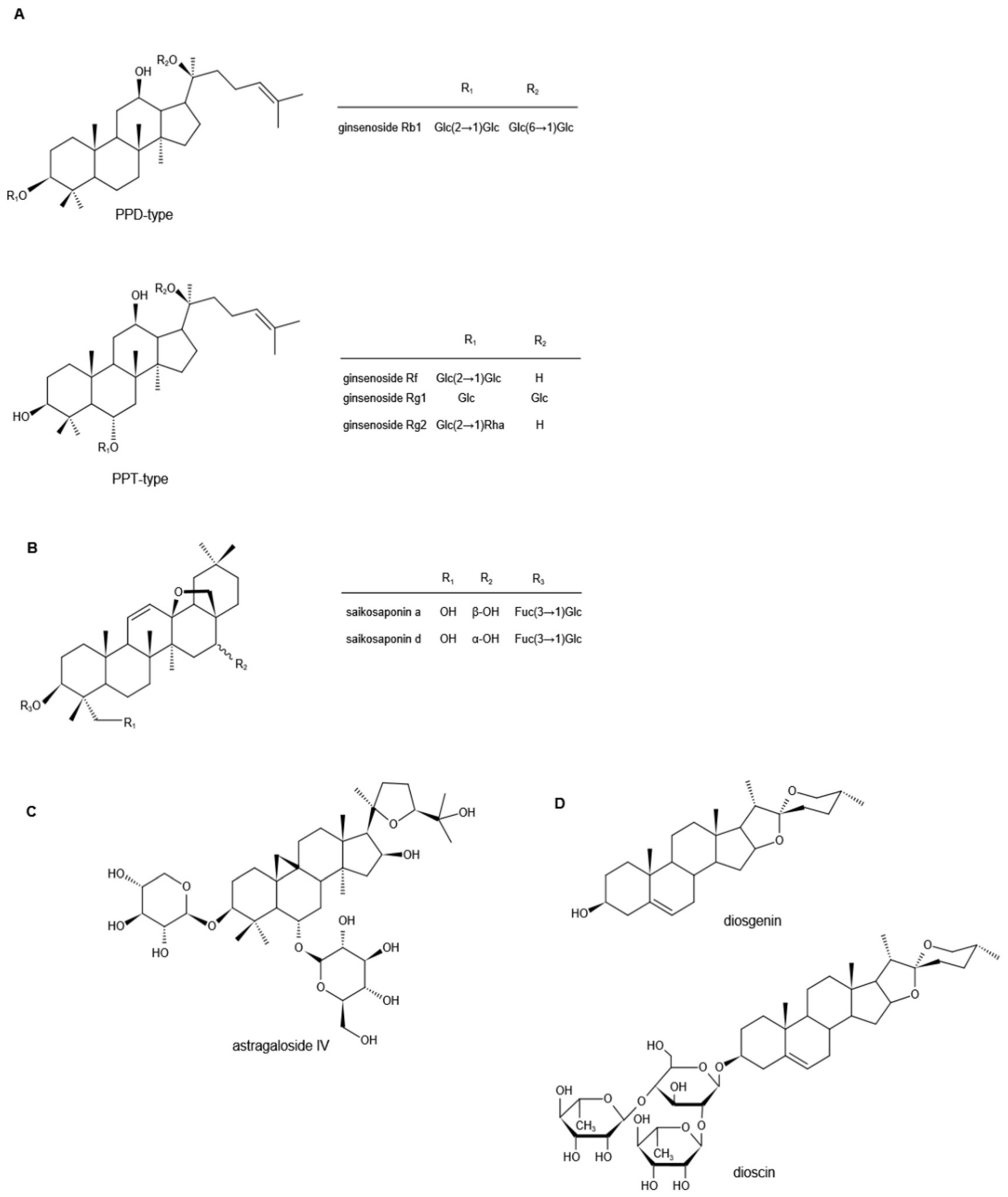
3.1. Ginsenosides
3.2. Saikosaponins
3.3. Astragalosides
3.4. Diosgenin
3.5. Saponin-Rich Extracts of O. sanctum
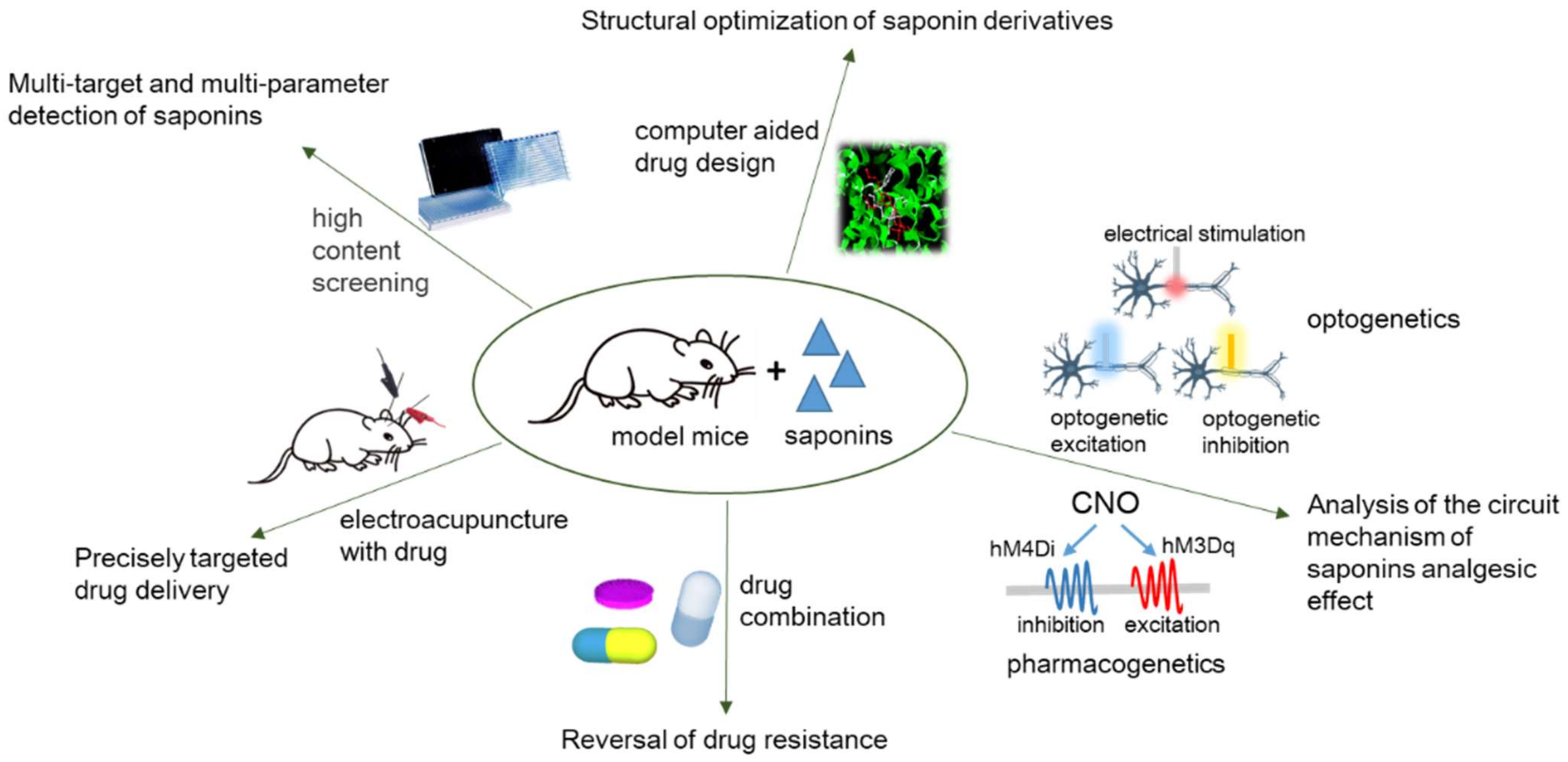
4. Conclusions and Perspective
5. Strengths and Limitations of the Review
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Sample Availability
References
- Raja, S.N.; Carr, D.B.; Cohen, M.; Finnerup, N.B.; Flor, H.; Gibson, S.; Keefe, F.J.; Mogil, J.S.; Ringkamp, M.; Sluka, K.A.; et al. The revised International Association for the Study of Pain definition of pain: Concepts, challenges, and compromises. Pain 2020, 161, 1976–1982. [Google Scholar] [CrossRef] [PubMed]
- St John Smith, E. Advances in understanding nociception and neuropathic pain. J. Neurol. 2018, 265, 231–238. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zilliox, L.A. Neuropathic Pain. Continuum 2017, 23, 512–532. [Google Scholar] [CrossRef] [PubMed]
- Meacham, K.; Shepherd, A.; Mohapatra, D.P.; Haroutounian, S. Neuropathic Pain: Central vs. Peripheral Mechanisms. Curr. Pain Headache Rep. 2017, 21, 28. [Google Scholar] [CrossRef] [PubMed]
- Van Hecke, O.; Austin, S.K.; Khan, R.A.; Smith, B.H.; Torrance, N. Neuropathic pain in the general population: A systematic review of epidemiological studies. Pain 2014, 155, 654–662. [Google Scholar] [CrossRef]
- Cohen, S.P.; Mao, J. Neuropathic pain: Mechanisms and their clinical implications. BMJ 2014, 348, f7656. [Google Scholar] [CrossRef] [Green Version]
- Finnerup, N.B.; Attal, N.; Haroutounian, S.; McNicol, E.; Baron, R.; Dworkin, R.H.; Gilron, I.; Haanpää, M.; Hansson, P.; Jensen, T.S.; et al. Pharmacotherapy for neuropathic pain in adults: A systematic review and meta-analysis. Lancet Neurol. 2015, 14, 162–173. [Google Scholar] [CrossRef] [Green Version]
- Jensen, T.S.; Finnerup, N.B. Allodynia and hyperalgesia in neuropathic pain: Clinical manifestations and mechanisms. Lancet Neurol. 2014, 13, 924–935. [Google Scholar] [CrossRef]
- Hansson, P.T.; Attal, N.; Baron, R.; Cruccu, G. Toward a definition of pharmacoresistant neuropathic pain. Eur. J. Pain 2009, 13, 439–440. [Google Scholar] [CrossRef]
- Papadopoulou, K.; Melton, R.E.; Leggett, M.; Daniels, M.J.; Osbourn, A.E. Compromised disease resistance in saponin-deficient plants. Proc. Natl. Acad. Sci. USA 1999, 96, 12923–12928. [Google Scholar] [CrossRef] [Green Version]
- Vincken, J.P.; Heng, L.; de Groot, A.; Gruppen, H. Saponins, classification and occurrence in the plant kingdom. Phytochemistry 2007, 68, 275–297. [Google Scholar] [CrossRef] [PubMed]
- Lin, M.; Yang, S.; Huang, J.; Zhou, L. Insecticidal Triterpenes in Meliaceae: Plant Species, Molecules and Activities: Part I; (Aphanamixis-Chukrasia). Int. J. Mol. Sci. 2021, 22, 13262. [Google Scholar] [CrossRef] [PubMed]
- Connolly, J.D.; Hill, R.A. Triterpenoids. Nat. Prod. Rep. 2008, 25, 794–830. [Google Scholar] [CrossRef] [PubMed]
- Luo, L.M.; Qin, L.; Pei, G.; Huang, S.G.; Zhou, X.J.; Chen, N.H. Advances in studies on steroidal saponins and their pharmacological activities in genus Lilium. Zhongguo Zhong Yao Za Zhi 2018, 43, 1416–1426. [Google Scholar] [CrossRef] [PubMed]
- Shi, J.; Arunasalam, K.; Yeung, D.; Kakuda, Y.; Mittal, G.; Jiang, Y. Saponins from edible legumes: Chemistry, processing, and health benefits. J. Med. Food 2004, 7, 67–78. [Google Scholar] [CrossRef] [PubMed]
- Man, S.; Gao, W.; Zhang, Y.; Huang, L.; Liu, C. Chemical study and medical application of saponins as anti-cancer agents. Fitoterapia 2010, 81, 703–714. [Google Scholar] [CrossRef]
- Hassan, H.S.; Sule, M.I.; Musa, A.M.; Musa, K.Y.; Abubakar, M.S.; Hassan, A.S. Anti-inflammatory activity of crude saponin extracts from five Nigerian medicinal plants. Afr. J. Tradit. Complementary Altern. Med. 2012, 9, 250–255. [Google Scholar] [CrossRef] [Green Version]
- Singh, D.; Chaudhuri, P.K. Structural characteristics, bioavailability and cardioprotective potential of saponins. Integr. Med. Res. 2018, 7, 33–43. [Google Scholar] [CrossRef]
- Xu, G.B.; Xiao, Y.H.; Zhang, Q.Y.; Zhou, M.; Liao, S.G. Hepatoprotective natural triterpenoids. Eur. J. Med. Chem. 2018, 145, 691–716. [Google Scholar] [CrossRef]
- Fang, Z.; Li, J.; Yang, R.; Fang, L.; Zhang, Y. A Review: The Triterpenoid Saponins and Biological Activities of Lonicera Linn. Molecules 2020, 25, 3773. [Google Scholar] [CrossRef]
- Lee, J.Y.; Choi, H.Y.; Park, C.S.; Kim, D.H.; Yune, T.Y. Total saponin extract, ginsenoside Rb1, and compound K alleviate peripheral and central neuropathic pain through estrogen receptors on rats. Phytother. Res. 2021, 35, 2119–2132. [Google Scholar] [CrossRef] [PubMed]
- Gao, C.; Guo, X.; Weng, L.; Zhou, W.; Shen, Y.; Yu, Y.; Han, Y. Effect of ginsenoside Rg1 on activity of spinal microglia and expression of p38 mitogen-activated protein kinase/nuclear transcription factor-kappa B in neuropathic pain rats. Int. J. Anesth. Resus 2017, 38, 1084. [Google Scholar] [CrossRef]
- Li, Y.; Chen, C.; Li, S.; Jiang, C. Ginsenoside Rf relieves mechanical hypersensitivity, depression-like behavior, and inflammatory reactions in chronic constriction injury rats. Phytother. Res. 2019, 33, 1095–1103. [Google Scholar] [CrossRef] [PubMed]
- Huang, F.; Li, Y.N.; Yin, F.; Wu, Y.T.; Zhao, D.X.; Li, Y.; Zhang, Y.F.; Zhu, Q.S. Ginsenoside Rb1 inhibits neuronal apoptosis and damage, enhances spinal aquaporin 4 expression and improves neurological deficits in rats with spinal cord ischemiareperfusion injury. Mol. Med. Rep. 2015, 11, 3565–3572. [Google Scholar] [CrossRef] [Green Version]
- Lee, G.; Choi, J.; Nam, Y.J.; Song, M.J.; Kim, J.K.; Kim, W.J.; Kim, P.; Lee, J.S.; Kim, S.; No, K.T.; et al. Identification and characterization of saikosaponins as antagonists of transient receptor potential A1 channel. Phytother. Res. 2020, 34, 788–795. [Google Scholar] [CrossRef]
- Lee, G.; Nam, Y.-J.; Kim, W.J.; Shin, B.H.; Lee, J.S.; Park, H.T.; Kim, P.; Lee, J.H.; Choi, Y. Saikosaponin D Ameliorates Mechanical Hypersensitivity in Animal Models of Neuropathic Pain. Planta Med. Int. Open 2020, 7, e145–e149. [Google Scholar] [CrossRef]
- Li, Z.Y.; Jiang, Y.M.; Liu, Y.M.; Guo, Z.; Shen, S.N.; Liu, X.M.; Pan, R.L. Saikosaponin D acts against corticosterone-induced apoptosis via regulation of mitochondrial GR translocation and a GR-dependent pathway. Prog. Neuropsychopharmacol. Biol. Psychiatry 2014, 53, 80–89. [Google Scholar] [CrossRef]
- Zhou, X.; Cheng, H.; Xu, D.; Yin, Q.; Cheng, L.; Wang, L.; Song, S.; Zhang, M. Attenuation of neuropathic pain by saikosaponin a in a rat model of chronic constriction injury. Neurochem. Res. 2014, 39, 2136–2142. [Google Scholar] [CrossRef]
- Lu, C.N.; Yuan, Z.G.; Zhang, X.L.; Yan, R.; Zhao, Y.Q.; Liao, M.; Chen, J.X. Saikosaponin a and its epimer saikosaponin d exhibit anti-inflammatory activity by suppressing activation of NF-kappaB signaling pathway. Int. Immunopharmacol. 2012, 14, 121–126. [Google Scholar] [CrossRef]
- Zhu, J.; Luo, C.; Wang, P.; He, Q.; Zhou, J.; Peng, H. Saikosaponin A mediates the inflammatory response by inhibiting the MAPK and NF-kappaB pathways in LPS-stimulated RAW 264.7 cells. Exp. Ther. Med. 2013, 5, 1345–1350. [Google Scholar] [CrossRef] [Green Version]
- Mao, X.; Miao, G.; Tao, X.; Hao, S.; Zhang, H.; Li, H.; Hou, Z.; Tian, R.; Lu, T.; Ma, J.; et al. Saikosaponin a protects TBI rats after controlled cortical impact and the underlying mechanism. Am. J. Transl. Res. 2016, 8, 133–141. [Google Scholar] [PubMed]
- Ahmadimoghaddam, D.; Zarei, M.; Mohammadi, S.; Izadidastenaei, Z.; Salehi, I. Bupleurum falcatum L. alleviates nociceptive and neuropathic pain: Potential mechanisms of action. J. Ethnopharmacol. 2021, 273, 113990. [Google Scholar] [CrossRef] [PubMed]
- Xu, Y.; Yu, Y.; Wang, Q.; Li, W.; Zhang, S.; Liao, X.; Liu, Y.; Su, Y.; Zhao, M.; Zhang, J. Active components of Bupleurum chinense and Angelica biserrata showed analgesic effects in formalin induced pain by acting on Nav1.7. J. Ethnopharmacol. 2021, 269, 113736. [Google Scholar] [CrossRef] [PubMed]
- Shi, G.B.; Fan, R.; Zhang, W.; Yang, C.; Wang, Q.; Song, J.; Gao, Y.; Hou, M.X.; Chen, Y.F.; Wang, T.C.; et al. Antinociceptive activity of astragaloside IV in the animal model of chronic constriction injury. Behav. Pharm. 2015, 26, 436–446. [Google Scholar] [CrossRef] [PubMed]
- Cheng, C.Y.; Yao, C.H.; Liu, B.S.; Liu, C.J.; Chen, G.W.; Chen, Y.S. The role of astragaloside in regeneration of the peripheral nerve system. J. Biomed. Mater. Res. A 2006, 76, 463–469. [Google Scholar] [CrossRef]
- Zhang, X.; Chen, J. The mechanism of astragaloside IV promoting sciatic nerve regeneration. Neural Regen. Res. 2013, 8, 2256–2265. [Google Scholar] [CrossRef] [PubMed]
- Yu, J.; Zhang, Y.; Sun, S.; Shen, J.; Qiu, J.; Yin, X.; Yin, H.; Jiang, S. Inhibitory effects of astragaloside IV on diabetic peripheral neuropathy in rats. Can. J. Physiol. Pharm. 2006, 84, 579–587. [Google Scholar] [CrossRef]
- Kiasalari, Z.; Rahmani, T.; Mahmoudi, N.; Baluchnejadmojarad, T.; Roghani, M. Diosgenin ameliorates development of neuropathic pain in diabetic rats: Involvement of oxidative stress and inflammation. Biomed. Pharm. 2017, 86, 654–661. [Google Scholar] [CrossRef]
- Kang, T.H.; Moon, E.; Hong, B.N.; Choi, S.Z.; Son, M.; Park, J.H.; Kim, S.Y. Diosgenin from Dioscorea nipponica ameliorates diabetic neuropathy by inducing nerve growth factor. Biol. Pharm. Bull. 2011, 34, 1493–1498. [Google Scholar] [CrossRef] [Green Version]
- Zhao, W.X.; Wang, P.F.; Song, H.G.; Sun, N. Diosgenin attenuates neuropathic pain in a rat model of chronic constriction injury. Mol. Med. Rep. 2017, 16, 1559–1564. [Google Scholar] [CrossRef]
- Lee, B.K.; Kim, C.J.; Shin, M.S.; Cho, Y.S. Diosgenin improves functional recovery from sciatic crushed nerve injury in rats. J. Exerc. Rehabil. 2018, 14, 566–572. [Google Scholar] [CrossRef] [PubMed]
- Muthuraman, A.; Diwan, V.; Jaggi, A.S.; Singh, N.; Singh, D. Ameliorative effects of Ocimum sanctum in sciatic nerve transection-induced neuropathy in rats. J. Ethnopharmacol. 2008, 120, 56–62. [Google Scholar] [CrossRef] [PubMed]
- Kaur, G.; Jaggi, A.S.; Singh, N. Exploring the potential effect of Ocimum sanctum in vincristine-induced neuropathic pain in rats. J. Brachial Plex. Peripher. Nerve Inj. 2010, 5, 3. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gautam, M.; Ramanathan, M. Saponins of Tribulus terrestris attenuated neuropathic pain induced with vincristine through central and peripheral mechanism. Inflammopharmacology 2019, 27, 761–772. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Chen, X.; Wu, L.; Li, Y.; Wang, L.; Zhao, X.; Zhao, T.; Zhang, L.; Yan, Z.; Wei, G. Ameliorative effects of escin on neuropathic pain induced by chronic constriction injury of sciatic nerve. J. Ethnopharmacol. 2021, 267, 113503. [Google Scholar] [CrossRef] [PubMed]
- Yan, F.; Chen, D.; Xie, J.; Zeng, W.; Li, Q. Escin alleviates chemotherapy-induced peripheral neuropathic pain by inducing autophagy in the spinal cord of rats. Nan Fang Yi Ke Da Xue Xue Bao 2020, 40, 1634–1638. [Google Scholar] [PubMed]
- Nemmani, K.V.; Ramarao, P. Ginsenoside Rf potentiates U-50,488H-induced analgesia and inhibits tolerance to its analgesia in mice. Life Sci. 2003, 72, 759–768. [Google Scholar] [CrossRef]
- Christensen, L.P. Ginsenosides chemistry, biosynthesis, analysis, and potential health effects. Adv. Food Nutr. Res. 2009, 55, 1–99. [Google Scholar] [CrossRef]
- Zheng, M.; Xin, Y.; Li, Y.; Xu, F.; Xi, X.; Guo, H.; Cui, X.; Cao, H.; Zhang, X.; Han, C. Ginsenosides: A Potential Neuroprotective Agent. BioMed Res. Int. 2018, 2018, 8174345. [Google Scholar] [CrossRef] [Green Version]
- Hou, M.; Wang, R.; Zhao, S.; Wang, Z. Ginsenosides in Panax genus and their biosynthesis. Acta Pharm. Sin. B 2021, 11, 1813–1834. [Google Scholar] [CrossRef]
- Lee, H.S.; Lee, H.J.; Yu, H.J.; Ju do, W.; Kim, Y.; Kim, C.T.; Kim, C.J.; Cho, Y.J.; Kim, N.; Choi, S.Y.; et al. A comparison between high hydrostatic pressure extraction and heat extraction of ginsenosides from ginseng (Panax ginseng CA Meyer). J. Sci. Food Agric. 2011, 91, 1466–1473. [Google Scholar] [CrossRef] [PubMed]
- Nag, S.A.; Qin, J.J.; Wang, W.; Wang, M.H.; Wang, H.; Zhang, R. Ginsenosides as Anticancer Agents: In vitro and in vivo Activities, Structure-Activity Relationships, and Molecular Mechanisms of Action. Front. Pharmacol. 2012, 3, 25. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wolf, G.; Yirmiya, R.; Goshen, I.; Iverfeldt, K.; Holmlund, L.; Takeda, K.; Shavit, Y. Impairment of interleukin-1 (IL-1) signaling reduces basal pain sensitivity in mice: Genetic, pharmacological and developmental aspects. Pain 2003, 104, 471–480. [Google Scholar] [CrossRef]
- Miyoshi, K.; Obata, K.; Kondo, T.; Okamura, H.; Noguchi, K. Interleukin-18-mediated microglia/astrocyte interaction in the spinal cord enhances neuropathic pain processing after nerve injury. J. Neurosci. 2008, 28, 12775–12787. [Google Scholar] [CrossRef]
- Ji, R.R.; Chamessian, A.; Zhang, Y.Q. Pain regulation by non-neuronal cells and inflammation. Science 2016, 354, 572–577. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sommer, C.; Leinders, M.; Üçeyler, N. Inflammation in the pathophysiology of neuropathic pain. Pain 2018, 159, 595–602. [Google Scholar] [CrossRef]
- Gao, X.-Q.; Yang, C.-X.; Chen, G.-J.; Wang, G.-Y.; Chen, B.; Tan, S.-K.; Liu, J.; Yuan, Q.-L. Ginsenoside Rb1 regulates the expressions of brain-derived neurotrophic factor and caspase-3 and induces neurogenesis in rats with experimental cerebral ischemia. J. Ethnopharmacol. 2010, 132, 393–399. [Google Scholar] [CrossRef]
- Adamo, D.; Calabria, E.; Coppola, N.; Pecoraro, G.; Mignogna, M.D. Vortioxetine as a new frontier in the treatment of chronic neuropathic pain: A review and update. Ther. Adv. Psychopharmacol. 2021, 11, 1–19. [Google Scholar] [CrossRef]
- Zhang, Q.L.; Li, S.Y.; Li, P. Effects of ginsenoside-Rg2 on mechanical allodynia, heat hyperalgeia, depressive state of rats with chronic sciatic nerve constriction injury. Zhongguo Ying Yong Sheng Li Xue Za Zhi 2019, 35, 228–231. [Google Scholar] [CrossRef]
- Ruizhen, C.; Chunyan, Z. Research progress on pharmacological activities of Saikosaponins from Radix Bupleurum. Occup. Health 2021, 37, 568–576. [Google Scholar] [CrossRef]
- He, Y.; Hu, Z.; Li, A.; Zhu, Z.; Yang, N.; Ying, Z.; He, J.; Wang, C.; Yin, S.; Cheng, S. Recent Advances in Biotransformation of Saponins. Molecules 2019, 24, 2365. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, X.; Li, X.; Huang, N.; Liu, R.; Sun, R. A comprehensive review and perspectives on pharmacology and toxicology of saikosaponins. Phytomedicine 2018, 50, 73–87. [Google Scholar] [CrossRef] [PubMed]
- Ashour, M.L.; Wink, M. Genus Bupleurum: A review of its phytochemistry, pharmacology and modes of action. J. Pharm. Pharm. 2011, 63, 305–321. [Google Scholar] [CrossRef] [PubMed]
- Ebata, N.; Nakajima, K.; Hayashi, K.; Okada, M.; Maruno, M. Saponins from the root of Bupleurum falcatum. Phytochemistry 1996, 41, 895–901. [Google Scholar] [CrossRef]
- Li, X.Q.; Song, Y.N.; Wang, S.J.; Rahman, K.; Zhu, J.Y.; Zhang, H. Saikosaponins: A review of pharmacological effects. J. Asian Nat. Prod. Res. 2018, 20, 399–411. [Google Scholar] [CrossRef]
- Wong, V.K.; Zhou, H.; Cheung, S.S.; Li, T.; Liu, L. Mechanistic study of saikosaponin-d (Ssd) on suppression of murine T lymphocyte activation. J Cell Biochem. 2009, 107, 303–315. [Google Scholar] [CrossRef]
- Wang, Y.L.; He, S.X.; Luo, J.Y. Progress in research on antitumor activity of saikosaponin and its mechanism. J. Chin. Integr. Med. 2006, 4, 98–100. [Google Scholar] [CrossRef]
- Gupta, R.; Saito, S.; Mori, Y.; Itoh, S.G.; Okumura, H.; Tominaga, M. Structural basis of TRPA1 inhibition by HC-030031 utilizing species-specific differences. Sci. Rep. 2016, 6, 37460. [Google Scholar] [CrossRef] [Green Version]
- Wang, S.; Kobayashi, K.; Kogure, Y.; Yamanaka, H.; Yamamoto, S.; Yagi, H.; Noguchi, K.; Dai, Y. Negative Regulation of TRPA1 by AMPK in Primary Sensory Neurons as a Potential Mechanism of Painful Diabetic Neuropathy. Diabetes 2018, 67, 98–109. [Google Scholar] [CrossRef] [Green Version]
- Nativi, C.; Gualdani, R.; Dragoni, E.; Di Cesare Mannelli, L.; Sostegni, S.; Norcini, M.; Gabrielli, G.; la Marca, G.; Richichi, B.; Francesconi, O.; et al. A TRPA1 antagonist reverts oxaliplatin-induced neuropathic pain. Sci. Rep. 2013, 3, 2005. [Google Scholar] [CrossRef] [Green Version]
- Nassini, R.; Gees, M.; Harrison, S.; De Siena, G.; Materazzi, S.; Moretto, N.; Failli, P.; Preti, D.; Marchetti, N.; Cavazzini, A.; et al. Oxaliplatin elicits mechanical and cold allodynia in rodents via TRPA1 receptor stimulation. Pain 2011, 152, 1621–1631. [Google Scholar] [CrossRef] [PubMed]
- Descoeur, J.; Pereira, V.; Pizzoccaro, A.; Francois, A.; Ling, B.; Maffre, V.; Couette, B.; Busserolles, J.; Courteix, C.; Noel, J.; et al. Oxaliplatin-induced cold hypersensitivity is due to remodelling of ion channel expression in nociceptors. EMBO Mol. Med. 2011, 3, 266–278. [Google Scholar] [CrossRef] [PubMed]
- Anderson, D.W.; Bradbury, K.A.; Schneider, J.S. Broad neuroprotective profile of nicotinamide in different mouse models of MPTP-induced parkinsonism. Eur. J. Neurosci. 2008, 28, 610–617. [Google Scholar] [CrossRef] [PubMed]
- Yuan, B.; Yang, R.; Ma, Y.; Zhou, S.; Zhang, X.; Liu, Y. A systematic review of the active saikosaponins and extracts isolated from Radix Bupleuri and their applications. Pharm. Biol. 2017, 55, 620–635. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lin, X.; Wu, S.; Wang, Q.; Shi, Y.; Liu, G.; Zhi, J.; Wang, F. Saikosaponin-D Reduces H2O2-Induced PC12 Cell Apoptosis by Removing ROS and Blocking MAPK-Dependent Oxidative Damage. Cell Mol. Neurobiol. 2016, 36, 1365–1375. [Google Scholar] [CrossRef]
- Sun, T.; Song, W.G.; Fu, Z.J.; Liu, Z.H.; Liu, Y.M.; Yao, S.L. Alleviation of neuropathic pain by intrathecal injection of antisense oligonucleotides to p65 subunit of NF-κB. Br. J. Anaesth. 2006, 97, 553–558. [Google Scholar] [CrossRef] [Green Version]
- Ramesh, G.; MacLean, A.G.; Philipp, M.T. Cytokines and chemokines at the crossroads of neuroinflammation, neurodegeneration, and neuropathic pain. Mediat. Inflamm. 2013, 2013, 480739. [Google Scholar] [CrossRef] [Green Version]
- Yoon, S.S.; Seo, J.W.; Ann, S.H.; Kim, H.Y.; Kim, H.S.; Cho, H.Y.; Yun, J.; Chung, E.Y.; Koo, J.S.; Yang, C.H. Effects of saikosaponin A on cocaine self-administration in rats. Neurosci. Lett. 2013, 555, 198–202. [Google Scholar] [CrossRef]
- Yoon, S.S.; Kim, H.S.; Cho, H.Y.; Yun, J.; Chung, E.Y.; Jang, C.G.; Kim, K.J.; Yang, C.H. Effect of saikosaponin A on maintenance of intravenous morphine self-administration. Neurosci. Lett. 2012, 529, 97–101. [Google Scholar] [CrossRef]
- Minett, M.S.; Nassar, M.A.; Clark, A.K.; Passmore, G.; Dickenson, A.H.; Wang, F.; Malcangio, M.; Wood, J.N. Distinct Nav1.7-dependent pain sensations require different sets of sensory and sympathetic neurons. Nat. Commun. 2012, 3, 791. [Google Scholar] [CrossRef] [Green Version]
- Chen, Z.; Liu, L.; Gao, C.; Chen, W.; Vong, C.T.; Yao, P.; Yang, Y.; Li, X.; Tang, X.; Wang, S.; et al. Astragali Radix (Huangqi): A promising edible immunomodulatory herbal medicine. J. Ethnopharmacol. 2020, 258, 112895. [Google Scholar] [CrossRef] [PubMed]
- Fu, J.; Wang, Z.; Huang, L.; Zheng, S.; Wang, D.; Chen, S.; Zhang, H.; Yang, S. Review of the botanical characteristics, phytochemistry, and pharmacology of Astragalus membranaceus (Huangqi). Phytother. Res. 2014, 28, 1275–1283. [Google Scholar] [CrossRef] [PubMed]
- Song, J.Z.; Mo, S.F.; Yip, Y.K.; Qiao, C.F.; Han, Q.B.; Xu, H.X. Development of microwave assisted extraction for the simultaneous determination of isoflavonoids and saponins in radix astragali by high performance liquid chromatography. J. Sep. Sci. 2007, 30, 819–824. [Google Scholar] [CrossRef] [PubMed]
- Ma, X.Q.; Shi, Q.; Duan, J.A.; Dong, T.T.; Tsim, K.W. Chemical analysis of Radix Astragali (Huangqi) in China: A comparison with its adulterants and seasonal variations. J. Agric. Food Chem. 2002, 50, 4861–4866. [Google Scholar] [CrossRef]
- Wang, Y.; Liu, L.; Ma, Y.; Guo, L.; Sun, Y.; Liu, Q.; Liu, J. Chemical Discrimination of Astragalus mongholicus and Astragalus membranaceus Based on Metabolomics Using UHPLC-ESI-Q-TOF-MS/MS Approach. Molecules 2019, 24, 4064. [Google Scholar] [CrossRef] [Green Version]
- Liu, D.-l.; Bao, H.-Y.; Liu, Y. Progress on Chemical Constituents and Pharmacological Effects of Astragali Radix in Recent Five Years. Food Drug 2014, 16, 68–70. [Google Scholar]
- Polat, E.; Bedir, E.; Perrone, A.; Piacente, S.; Alankus-Caliskan, O. Triterpenoid saponins from Astragalus wiedemannianus Fischer. Phytochemistry 2010, 71, 658–662. [Google Scholar] [CrossRef]
- Verotta, L.; Guerrini, M.; El-Sebakhy, N.A.; Assad, A.M.; Toaima, S.M.; Radwan, M.M.; Luo, Y.D.; Pezzuto, J.M. Cycloartane and oleanane saponins from egyptian astragalus spp. as modulators of lymphocyte proliferation. Planta Med. 2002, 68, 986–994. [Google Scholar] [CrossRef]
- Zhang, J.; Wu, C.; Gao, L.; Du, G.; Qin, X. Astragaloside IV derived from Astragalus membranaceus: A research review on the pharmacological effects. Adv. Pharm. 2020, 87, 89–112. [Google Scholar] [CrossRef]
- Yang, F.; Xiao, X.; Lee, B.H.; Vu, S.; Yang, W.; Yarov-Yarovoy, V.; Zheng, J. The conformational wave in capsaicin activation of transient receptor potential vanilloid 1 ion channel. Nat. Commun. 2018, 9, 2879. [Google Scholar] [CrossRef]
- Tominaga, M.; Caterina, M.J.; Malmberg, A.B.; Rosen, T.A.; Gilbert, H.; Skinner, K.; Raumann, B.E.; Basbaum, A.I.; Julius, D. The cloned capsaicin receptor integrates multiple pain-producing stimuli. Neuron 1998, 21, 531–543. [Google Scholar] [CrossRef] [Green Version]
- Mitchell, K.; Bates, B.D.; Keller, J.M.; Lopez, M.; Scholl, L.; Navarro, J.; Madian, N.; Haspel, G.; Nemenov, M.I.; Iadarola, M.J. Ablation of rat TRPV1-expressing Adelta/C-fibers with resiniferatoxin: Analysis of withdrawal behaviors, recovery of function and molecular correlates. Mol. Pain 2010, 6, 94. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, X.; Kang, L.; Li, G.; Zeng, H.; Zhang, L.; Ling, X.; Dong, H.; Liang, S.; Chen, H. Intrathecal leptin inhibits expression of the P2X2/3 receptors and alleviates neuropathic pain induced by chronic constriction sciatic nerve injury. Mol. Pain 2013, 9, 65. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lin, J.; Li, G.; Den, X.; Xu, C.; Liu, S.; Gao, Y.; Liu, H.; Zhang, J.; Li, X.; Liang, S. VEGF and its receptor-2 involved in neuropathic pain transmission mediated by P2X2/3 receptor of primary sensory neurons. Brain Res. Bull. 2010, 83, 284–291. [Google Scholar] [CrossRef] [PubMed]
- Dong, Z.Q.; Ma, F.; Xie, H.; Wang, Y.Q.; Wu, G.C. Down-regulation of GFRalpha-1 expression by antisense oligodeoxynucleotide attenuates electroacupuncture analgesia on heat hyperalgesia in a rat model of neuropathic pain. Brain Res. Bull. 2006, 69, 30–36. [Google Scholar] [CrossRef]
- Dong, Z.Q.; Ma, F.; Xie, H.; Wang, Y.Q.; Wu, G.C. Changes of expression of glial cell line-derived neurotrophic factor and its receptor in dorsal root ganglions and spinal dorsal horn during electroacupuncture treatment in neuropathic pain rats. Neurosci. Lett. 2005, 376, 143–148. [Google Scholar] [CrossRef]
- Shen, Y.; Meiri, K. GAP-43 dependency defines distinct effects of netrin-1 on cortical and spinal neurite outgrowth and directional guidance. Int. J. Dev. Neurosci. 2012, 31, 11–20. [Google Scholar] [CrossRef]
- Mendonca, H.R.; Araujo, S.E.; Gomes, A.L.; Sholl-Franco, A.; da Cunha Faria Melibeu, A.; Serfaty, C.A.; Campello-Costa, P. Expression of GAP-43 during development and after monocular enucleation in the rat superior colliculus. Neurosci. Lett. 2010, 477, 23–27. [Google Scholar] [CrossRef]
- Davidson, J.A. Treatment of the patient with diabetes: Importance of maintaining target HbA(1c) levels. Curr. Med. Res. Opin. 2004, 20, 1919–1927. [Google Scholar] [CrossRef]
- Yi, T.; Fan, L.L.; Chen, H.L.; Zhu, G.Y.; Suen, H.M.; Tang, Y.N.; Zhu, L.; Chu, C.; Zhao, Z.Z.; Chen, H.B. Comparative analysis of diosgenin in Dioscorea species and related medicinal plants by UPLC-DAD-MS. BMC Biochem. 2014, 15, 19. [Google Scholar] [CrossRef] [Green Version]
- Arya, P.; Kumar, P. Diosgenin a steroidal compound: An emerging way to cancer management. J. Food Biochem. 2021, 45, e14005. [Google Scholar] [CrossRef]
- Chen, Y.; Tang, Y.-M.; Yu, S.-L.; Han, Y.-W.; Kou, J.-P.; Liu, B.-L.; Yu, B.-Y. Advances in the pharmacological activities and mechanisms of diosgenin. Chin. J. Nat. Med. 2015, 13, 578–587. [Google Scholar] [CrossRef]
- Al-Habori, M.; Raman, A.; Lawrence, M.J.; Skett, P. In vitro effect of fenugreek extracts on intestinal sodium-dependent glucose uptake and hepatic glycogen phosphorylase A. Int. J. Exp. Diabetes Res. 2001, 2, 91–99. [Google Scholar] [CrossRef] [Green Version]
- Fan, R.; He, W.; Fan, Y.; Xu, W.; Xu, W.; Yan, G.; Xu, S. Recent advances in chemical synthesis, biocatalysis, and biological evaluation of diosgenin derivatives—A review. Steroids 2022, 180, 108991. [Google Scholar] [CrossRef]
- Fernandes, P.; Cruz, A.; Angelova, B.; Pinheiro, H.M.; Cabral, J.M.S. Microbial conversion of steroid compounds: Recent developments. Enzym. Microb. Tech. 2003, 32, 688–705. [Google Scholar] [CrossRef]
- Obrosova, I.G. Update on the pathogenesis of diabetic neuropathy. Curr. Diabetes Rep. 2003, 3, 439–445. [Google Scholar] [CrossRef]
- Kasznicki, J.; Kosmalski, M.; Sliwinska, A.; Mrowicka, M.; Stanczyk, M.; Majsterek, I.; Drzewoski, J. Evaluation of oxidative stress markers in pathogenesis of diabetic neuropathy. Mol. Biol. Rep. 2012, 39, 8669–8678. [Google Scholar] [CrossRef] [Green Version]
- Vincent, A.M.; Russell, J.W.; Low, P.; Feldman, E.L. Oxidative stress in the pathogenesis of diabetic neuropathy. Endocr. Rev. 2004, 25, 612–628. [Google Scholar] [CrossRef]
- Wang, C.; Ning, L.P.; Wang, Y.H.; Zhang, Y.; Ding, X.L.; Ge, H.Y.; Arendt-Nielsen, L.; Yue, S.W. Nuclear factor-kappa B mediates TRPV4-NO pathway involved in thermal hyperalgesia following chronic compression of the dorsal root ganglion in rats. Behav. Brain Res. 2011, 221, 19–24. [Google Scholar] [CrossRef]
- Zhang, Y.P.; Song, C.Y.; Yuan, Y.; Eber, A.; Rodriguez, Y.; Levitt, R.C.; Takacs, P.; Yang, Z.; Goldberg, R.; Candiotti, K.A. Diabetic neuropathic pain development in type 2 diabetic mouse model and the prophylactic and therapeutic effects of coenzyme Q10. Neurobiol. Dis. 2013, 58, 169–178. [Google Scholar] [CrossRef]
- Kumar, A.; Negi, G.; Sharma, S.S. Suppression of NF-κB and NF-κB regulated oxidative stress and neuroinflammation by BAY 11-7082 (IκB phosphorylation inhibitor) in experimental diabetic neuropathy. Biochimie 2012, 94, 1158–1165. [Google Scholar] [CrossRef]
- Kawamura, N.; Dyck, P.J.; Schmeichel, A.M.; Engelstad, J.K.; Low, P.A.; Dyck, P.J. Inflammatory mediators in diabetic and non-diabetic lumbosacral radiculoplexus neuropathy. Acta Neuropathol. 2008, 115, 231–239. [Google Scholar] [CrossRef]
- Gao, M.; Chen, L.; Yu, H.; Sun, Q.; Kou, J.; Yu, B. Diosgenin down-regulates NF-kappaB p65/p50 and p38MAPK pathways and attenuates acute lung injury induced by lipopolysaccharide in mice. Int. Immunopharmacol. 2013, 15, 240–245. [Google Scholar] [CrossRef]
- Gao, Z.; Feng, Y.; Ju, H. The Different Dynamic Changes of Nerve Growth Factor in the Dorsal Horn and Dorsal Root Ganglion Leads to Hyperalgesia and Allodynia in Diabetic Neuropathic Pain. Pain Physician 2017, 20, E551–E561. [Google Scholar]
- Calissano, P.; Amadoro, G.; Matrone, C.; Ciafrè, S.; Marolda, R.; Corsetti, V.; Ciotti, M.T.; Mercanti, D.; Di Luzio, A.; Severini, C.; et al. Does the term ‘trophic’ actually mean anti-amyloidogenic? The case of NGF. Cell Death Differ. 2010, 17, 1126–1133. [Google Scholar] [CrossRef]
- Byun, Y.H.; Lee, M.H.; Kim, S.S.; Kim, H.; Chang, H.K.; Lee, T.H.; Jang, M.H.; Shin, M.C.; Shin, M.S.; Kim, C.J. Treadmill running promotes functional recovery and decreases brain-derived neurotrophic factor mRNA expression following sciatic crushed nerve injury in rats. J. Sports Med. Phys. Fit. 2005, 45, 222–228. [Google Scholar]
- Liu, M.; Luo, F.; Qing, Z.; Yang, H.; Liu, X.; Yang, Z.; Zeng, J. Chemical Composition and Bioactivity of Essential Oil of Ten Labiatae Species. Molecules 2020, 25, 4862. [Google Scholar] [CrossRef]
- Palida, A.; Mi, R.; Cong, Y.; Yi, B.; Wang, X. Isolation and identification of chemical constituents in Ocimum bacilicum. West China J. Pharm. Sci. 2007, 22, 489–490. [Google Scholar] [CrossRef]
- Kaur, G.; Bali, A.; Singh, N.; Jaggi, A.S. Ameliorative potential of Ocimum sanctum in chronic constriction injury-induced neuropathic pain in rats. An. Acad. Bras. Ciênc. 2015, 87, 417–429. [Google Scholar] [CrossRef] [Green Version]
- Anandjiwala, S.; Kalola, J.; Rajani, M. Quantification of eugenol, luteolin, ursolic acid, and oleanolic acid in black (Krishna Tulasi) and green (Sri Tulasi) varieties of Ocimum sanctum Linn. using high-performance thin-layer chromatography. J. AOAC Int. 2006, 89, 1467–1474. [Google Scholar] [CrossRef] [Green Version]
- Rao, A.R.; Veeresham, C.; Asres, K. In vitro and in vivo inhibitory activities of four Indian medicinal plant extracts and their major components on rat aldose reductase and generation of advanced glycation endproducts. Phytother. Res. 2013, 27, 753–760. [Google Scholar] [CrossRef]
- Carrasco, C.; Naziroǧlu, M.; Rodríguez, A.B.; Pariente, J.A. Neuropathic Pain: Delving into the Oxidative Origin and the Possible Implication of Transient Receptor Potential Channels. Front. Physiol. 2018, 9, 95. [Google Scholar] [CrossRef]
- Bourinet, E.; Altier, C.; Hildebrand, M.E.; Trang, T.; Salter, M.W.; Zamponi, G.W. Calcium-permeable ion channels in pain signaling. Physiol. Rev. 2014, 94, 81–140. [Google Scholar] [CrossRef]
- Katsuyama, Y.; Sato, Y.; Okano, Y.; Masaki, H. Intracellular oxidative stress induced by calcium influx initiates the activation of phagocytosis in keratinocytes accumulating at S-phase of the cell cycle after UVB irradiation. J. Dermatol. Sci. 2021, 103, 41–48. [Google Scholar] [CrossRef]
- Gibson, G.E. Interactions of oxidative stress with cellular calcium dynamics and glucose metabolism in Alzheimer’s disease. Free Radic. Biol. Med. 2002, 32, 1061–1070. [Google Scholar] [CrossRef]
- Goodwin, J.; Nath, S.; Engelborghs, Y.; Pountney, D.L. Raised calcium and oxidative stress cooperatively promote alpha-synuclein aggregate formation. Neurochem. Int. 2013, 62, 703–711. [Google Scholar] [CrossRef]
- Carbonera, D.; Azzone, G.F. Permeability of inner mitochondrial membrane and oxidative stress. Biochim. Biophys. Acta 1988, 943, 245–255. [Google Scholar] [CrossRef]
- George, S.; Chaturvedi, P. Protective role of Ocimum canum plant extract in alcohol-induced oxidative stress in albino rats. Br. J. Biomed. Sci. 2008, 65, 80–85. [Google Scholar] [CrossRef]
- Kelm, M.A.; Nair, M.G.; Strasburg, G.M.; DeWitt, D.L. Antioxidant and cyclooxygenase inhibitory phenolic compounds from Ocimum sanctum Linn. Phytomedicine 2000, 7, 7–13. [Google Scholar] [CrossRef]
- Balanehru, S.; Nagarajan, B. Protective effect of oleanolic acid and ursolic acid against lipid peroxidation. Biochem. Int. 1991, 24, 981–990. [Google Scholar]
- Hu, S.; Wu, Y.; Zhao, B.; Hu, H.; Zhu, B.; Sun, Z.; Li, P.; Du, S. Panax notoginseng Saponins Protect Cerebral Microvascular Endothelial Cells against Oxygen-Glucose Deprivation/Reperfusion-Induced Barrier Dysfunction via Activation of PI3K/Akt/Nrf2 Antioxidant Signaling Pathway. Molecules 2018, 23, 2781. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Neco, P.; Rose, B.; Huynh, N.; Zhang, R.; Bridge, J.H.; Philipson, K.D.; Goldhaber, J.I. Sodium-calcium exchange is essential for effective triggering of calcium release in mouse heart. Biophys. J. 2010, 99, 755–764. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sarkar, A.; Pandey, D.N.; Pant, M.C. A report on the effects of Ocimum sanctum (Tulsi) leaves and seeds on blood and urinary uric acid, urea and urine volume in normal albino rabbits. Indian J. Physiol. Pharmacol. 1990, 34, 61–62. [Google Scholar] [PubMed]
- Dalbeth, N.; Choi, H.K.; Joosten, L.A.B.; Khanna, P.P.; Matsuo, H.; Perez-Ruiz, F.; Stamp, L.K. Gout. Nat. Rev. Dis. Prim. 2019, 5, 69. [Google Scholar] [CrossRef] [PubMed]
- Khanna, N.; Bhatia, J. Antinociceptive action of Ocimum sanctum (Tulsi) in mice: Possible mechanisms involved. J. Ethnopharmacol. 2003, 88, 293–296. [Google Scholar] [CrossRef]
- Zhang, X.H.; Feng, C.C.; Pei, L.J.; Zhang, Y.N.; Chen, L.; Wei, X.Q.; Zhou, J.; Yong, Y.; Wang, K. Electroacupuncture attenuates neuropathic pain and comorbid negative behavior: The involvement of the dopamine system in the amygdala. Front. Neurosci. 2021, 15, 657507. [Google Scholar] [CrossRef]
- Shao, F.; Du, J.; Wang, S.; Cerne, R.; Fang, J.; Shao, X.; Jin, X.; Fang, J. Electroacupuncture alleviates anxiety-like behavior in pain aversion rats by attenuating the expression of neuropeptide Y in anterior cingulate cortex. Clin. Complementary Med. Pharmacol. 2022, 2, 100019. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
{kind=link}
| Saponins | Animals/Cells | Dose mg/kg | Effects/Behavioral Evaluation | Mechanisms of Action | Reference |
|---|---|---|---|---|---|
| total saponin extract (TSE), ginsenoside Rb1, Rb1 metabolite compound K | male Sprague–Dawley rats | TSE 50 mg/kg, Rb1 12.5 mg/kg, compound K 7 mg/kg (p.o.) | ↓TNI-induced mechanical, cold, and warm allodynia; ↓SCI-induced mechanical allodynia and thermal hyperalgesia; →Basso–Beattie–Bresnahan locomotor scale | ↓IL-1β, IL-6, iNOS, COX-2; ↓microglialand astrocyte activation | Jee Youn Lee [21] |
| ginsenoside Rg1 | male Sprague–Dawley rats | 2, 5, 10 ug/uL (intrathecal injection); 4 days | ↓CCI-induced thermal hyperalgesia; | ↓IBA-1, OX-42; ↓microglialover-activation; ↓p38MAPK/NF-κB signaling pathway | Gao Chao [22] |
| ginsenoside Rf | male Sprague–Dawley rats | 0.5, 1.5, 3 mg/kg (i.p.); 1 day, 7 days, 14 days, 21 days | ↓CCI-induced mechanical hyperalgesia; ↓CCI-induced immobility in the forced swimming test | ↓IL-1β and IL-6 in both the spinal cord and the DRG; ↑IL-10 in the DRG but not in the spinal cord | Yangyi Li [23] |
| ginsenoside Rb1 | male and female Sprague-Dawley rats | 10 mg/kg (i.p.); 1–7 days | ↑SCI-induced reduction of Basso–Beattie–Bresnahan locomotor scale | ↓neuronal damage; ↓apoptotic rate in spinal cord neurons. ↑ AQP4 expression | Fei Huang [24] |
| saikosaponin A; saikosaponin B1; saikosaponin B2; saikosaponin C; saikosaponin D; saikosaponin F; B. falcatum extract | male ICR mice/HEK293 cells | B. falcatum extract 50 mg/kg, saikosaponin D 20 mg/kg (p.o.) | ↓AITC-induced nociceptive behaviors; ↓vincristine-induced mechanical hypersensitivity | saikosaponins are TRPA1 antagonists | Gyeongbeen Lee [25] |
| saikosaponin D | male ICR mice | 10 mg/kg (p.o.), 1 day and 15 days | ↓ STZ-induced mechanical hypersensitivity and paclitaxel-induced mechanical allodynia | ____ | Gyeongbeen Lee [26] |
| saikosaponin D | PC12 cells | 200, 300, 400 μg/mL | ↓H2O2-induced decrease in cell viability; ↓apoptosis rate; ↓caspase-3 activation and poly-ADP-ribose polymerase cleavage; improved the nuclear morphology | ↓H2O2-induced release of malonic dialdehyde MDA and lactate dehy-drogenase; ↑SOD; ↓apoptotic rate; ↓ H2O2-induced p-ERK, p-c-JNK, p-p38MAPK | Xuemei Lin [27] |
| saikosaponin A | male Sprague–Dawley rats | 6.25, 12.50, 25.00 mg/kg (i.p.), 14 days | ↓CCI-induced mechanical allodynia and thermal hyperalgesia | ↓TNF-α, IL-1β, IL-2 in spinal cord; ↓p38MAPK/NF-κB signaling pathway | Xin Zhou [28] |
| saikosaponin A; saikosaponin D | male BALB/c mice; male Sprague–Dawley rats; Raw264.7 cells | 20, 10, 5 mg/kg (p.o.); | ↓carrageenan-induced rat paw edema; ↓acetic acid-induced evans blue dye leakage | ↓NO, PGE2, IL-6, TNF-α, iNOS, COX-2 in LPS-induced RAW264.7 cells; ↓NF-κB signaling pathway | Chun-Ni Lu [29] |
| saikosaponin A | Raw264.7 cells | 3.125, 6.25, 12.5, 25 μM | ____ | ↓IL-1β, IL-6 TNF-α, iNOS, COX-2 in LPS-induced RAW264.7 cells; ↓MAPK/NF-κB signaling pathway | Jie Zhu [30] |
| saikosaponin A | male Sprague–Dawley rats | 20 mg/kg (i.v.); 3 days | ↑neurological functions andcognition; ↓brain edema and blood brain barrier permeability after controlled cortical impact | ↓AQP-4, MMP-9, MAPK, c-JNK, TNF-α, IL-6; ↓MAPK signaling pathway | Xiang Mao [31] |
| Bupleurum falcatum L. roots essential oil (BFEO); Saikosaponin A | male Swiss mice | BFEO 25, 50, 100 mg/kg (p.o.); SA 6, 12, 25 mg/kg (p.o.) | ↑the antinociceptive activity in formalin-induced paw licking test, ↓mechanical allodynia, →locomotor action | ↑the L-arginine–NO–cGMP-KATP channel pathway | Davoud Ahmadimoghaddam [32] |
| saikosaponin A | male Kunming mice; Nav1.7-CHO cells | 2.5, 5, 10 mg/kg (i.g); 100 nM; | ↓thermal pain and formalin-induced nociceptive responses | inhibitory effect on Nav1.7 | Yijia Xu [33] |
| astragaloside IV | male Sprague–Dawley rats | 15, 30, 60 mg/kg (i.p.), 23 days | ↓CCI-induced mechanical allodynia and thermal hyperalgesia; ↑CCI-induced reduction of nerve conduction velocity; →locomotor action | ↓P2 × 3, TRPA1 and TRPV1 in the DRG; restoring the histological structure of the damaged sciatic nerve by accumulating GFRα1 | Guo-Bing Shi [34] |
| astragaloside IV | Sprague–Dawley rats | 0, 50, 100, 200μM | ↑regeneration rate across the wide gap; ↑myelinated axons; ↑evoked action potential; ↓nerve regeneration | plays a dual role in anastomosis | Chun-Yuan Cheng [35] |
| astragaloside IV | BALB/c mice | 2.5, 5, 10 mg/kg (i.p.) | ↑denervating the left sciatic nerve-induced the number and diameter of myelinated nerve fibers; ↑motor nerve conduction velocity and action potential amplitude in the sciatic nerve | ↑growth-associated protein-43 expression; ↑pheral nerve regeneration and functional reconstruction | Xiaohong Zhang [36] |
| astragaloside IV | male Sprague–Dawley rats | 3, 6, 12 mg/kg (p.o.), 12 days | ↑pain threshold in STZ-diabetic rats; ↑motor nerve conduction velocity | ↓blood glucose concentration and HbA1C levels; ↑plasma insulin levels, the activity of glutathione peroxidase in nerves; ↓ the activation of aldose reductase in erythrocytes and advanced glycation end products; ↑Na+,K+-ATPase activity | Junxian Yu [37] |
| diosgenin | male albino Wistar rats | 40 mg/kg (i.g), 35 days | ↓mechanical hyperalgesia and thermal hyperalgesia and pain score in STZ-diabetic rats; | ↓MDA, ↑SOD and catalase activity; ↓NF-κB and IL-1β | Zahra Kiasalari [38] |
| diosgenin | male ICR mice, male Sprague–Dawley rats; PC12 cells, C6 glioma cells | 10 mg/kg (p.o.); 0.1–10 mg/mL | ↑NGF levels in alloxan-diabetic rats; ↑nerveconduction velocities | reverses functional and ultrastructural changes and induces neural regeneration | Tong Ho Kang [39] |
| diosgenin | male Sprague–Dawley rats | 10, 20, 40 mg/kg (i.p.), 14 days | ↓CCI-induced mechanical allodynia and thermal hyperalgesia. | ↓TNF-α, IL-1β, IL-2, and oxidative stress; ↓p38MAPK/NF-κB signaling pathway | Wei-Xin Zhao [40] |
| diosgenin | male Sprague–Dawley rats | 25, 50, 100 mg/kg (p.o.), 7 days | ↑functional locomotor recovery following sciatic crushed nerve injury | ↓nerve injury-induced increase in BDNF, TrkB, COX-2, and iNOS expressions | Byung-Ki Lee [41] |
| ocimum sanctum, saponin-rich extracts | Wistar albino rats | 100 and 200 mg/kg (p.o.), 14 days | ↓CCI-induced cold-allodynia, heat-hyperalgesia, mechanical hyperalgesia and tail cold-hyperalgesia | ↓oxidative stress and calcium levels | Gurpreet Kaur [42] |
| ocimum sanctum, saponin-rich extracts | Wistar albino rats | 100 and 200 mg/kg (p.o.), 14 days | ↓ vincristine-induced cold-allodynia, heat-hyperalgesia, mechanical hyperalgesia and tail cold-hyperalgesia | ↓oxidative stress and calcium levels | Gurpreet Kaur [43] |
| saponins of Tribulus terrestris | Wistar rats of either sex | 25, 50, 100 mg/kg (p.o.) | ↓vincristine-induced mechanical hyperalgesia and allodynia; ↓chemical-induced nociception | ↓TNF-α, IL-1β, and IL-6; ↑ nerve conduction velocity, neurotransmitters, l-glutamic acid and l-aspartic acid | Mrinmoy Gautam [44] |
| escin | male Kunming mice, male Sprague–Dawley rats; PC12 cells | 7, 14, 28 mg/kg (i.g.), 14 days; 15, 25, 35 mg/kg (i.g.), 3 days; 2.5, 5, 10 μM | ↓CCI-induced thermal hyperalgesia; ↓ formalin-induced nociceptive responses | ↓TLR-4/NF-κB signal pathway; ↓GFAP, NGF | Liudai Zhang [45] |
| escin | male Sprague–Dawley rats | 4 mg/kg (i.p.), 7 days | ↓Paclitaxel-induced mechanical allodynia and thermal hyperalgesia | ↑LC3II expression, ↓p62expression levels | Yan Fang [46] |
| ginsenoside Rf | Swiss male mice | 10−14, 10−12, and 10−10 mg/kg, (i.p.), 6 days; 10−12–10−2 mg/mL | ↑U50-induced analgesia, ↓tolerance | nonopioid and non-dihydropyridine-sensitive Ca2+ channel mechanisms; non-benzodiazepine-GABAAergic mechanisms | Kumar V.S. Nemmani [47] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tan, B.; Wu, X.; Yu, J.; Chen, Z. The Role of Saponins in the Treatment of Neuropathic Pain. Molecules 2022, 27, 3956. https://doi.org/10.3390/molecules27123956
Tan B, Wu X, Yu J, Chen Z. The Role of Saponins in the Treatment of Neuropathic Pain. Molecules. 2022; 27(12):3956. https://doi.org/10.3390/molecules27123956
Chicago/Turabian StyleTan, Bei, Xueqing Wu, Jie Yu, and Zhong Chen. 2022. "The Role of Saponins in the Treatment of Neuropathic Pain" Molecules 27, no. 12: 3956. https://doi.org/10.3390/molecules27123956
APA StyleTan, B., Wu, X., Yu, J., & Chen, Z. (2022). The Role of Saponins in the Treatment of Neuropathic Pain. Molecules, 27(12), 3956. https://doi.org/10.3390/molecules27123956