
Precise Quantification of Molybdate In Vitro by the FRET-Based Nanosensor ‘MolyProbe’



Abstract
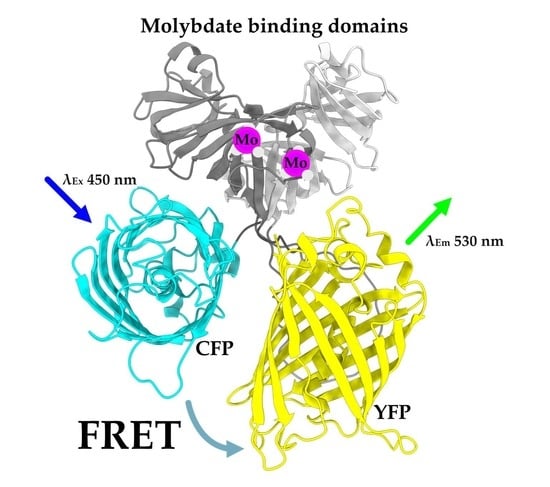
:
1. Introduction
2. Results
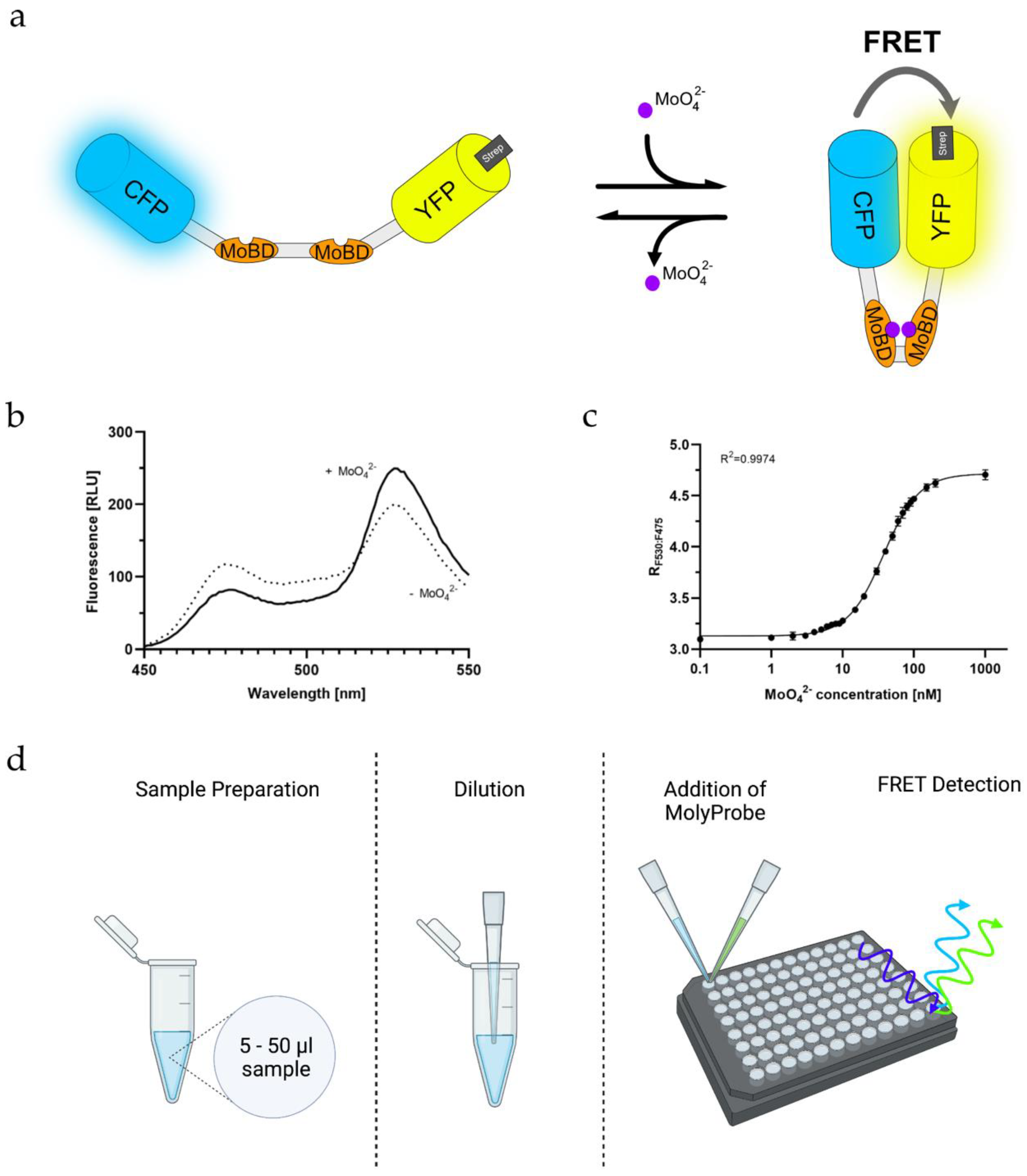
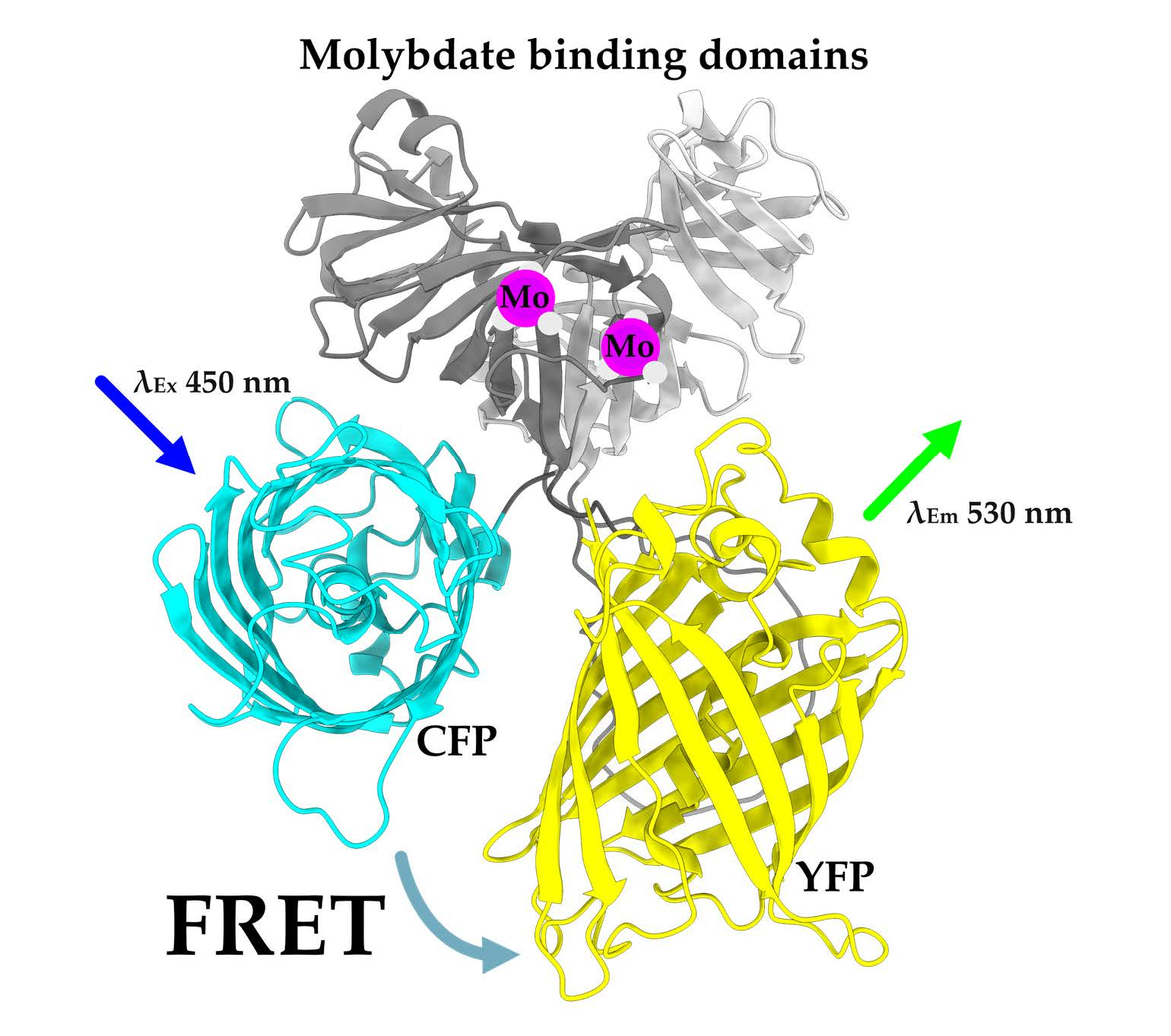
2.1. Creating a Strep-Tagged MolyProbe
2.2. Purification of MolyProbe-Strep
2.3. Quantifying Molybdate Using an In Vitro FRET Assay
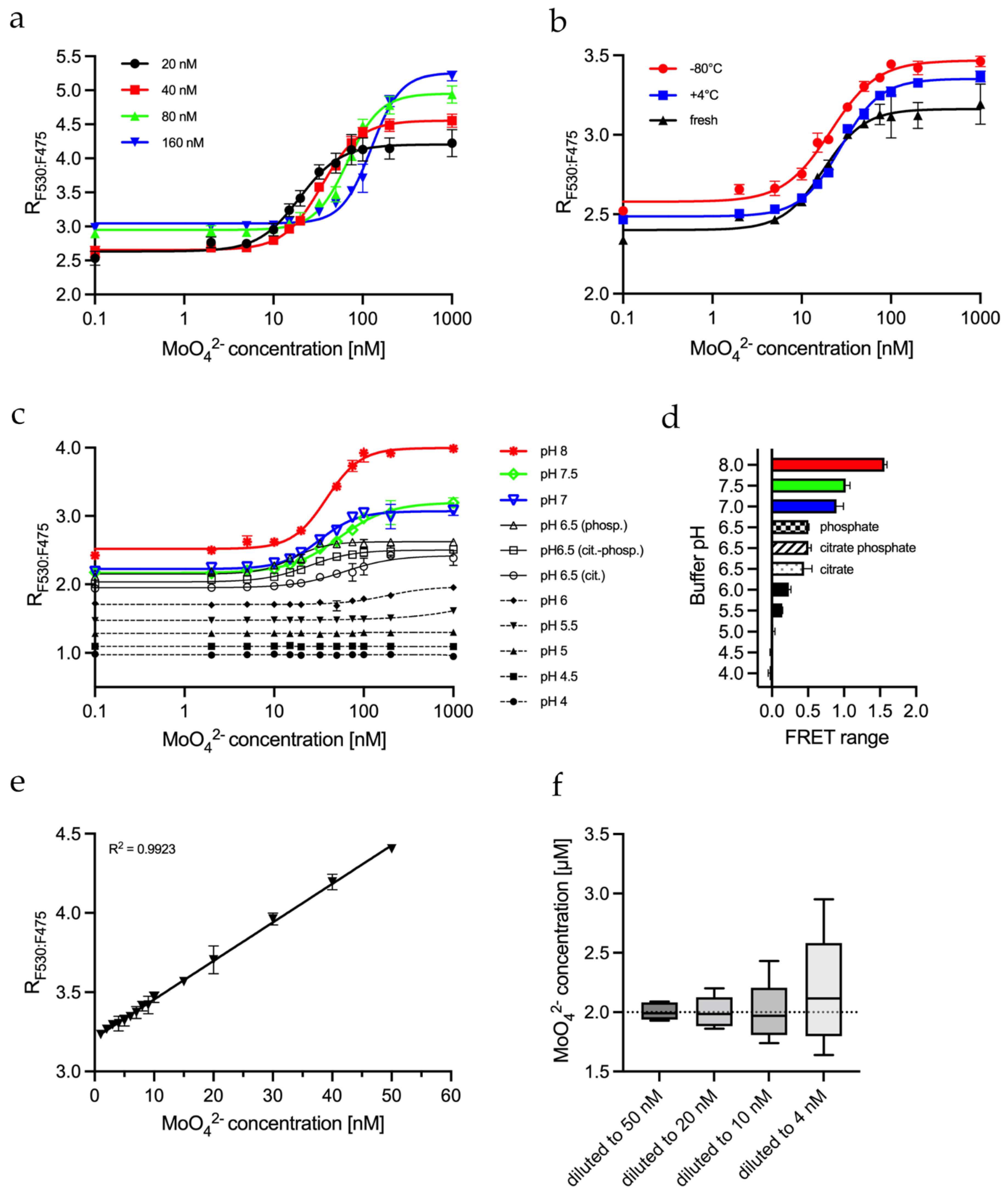
2.4. Biochemical Parameters Influencing the FRET Assay
2.5. Determining MolyProbe’s Detection Range
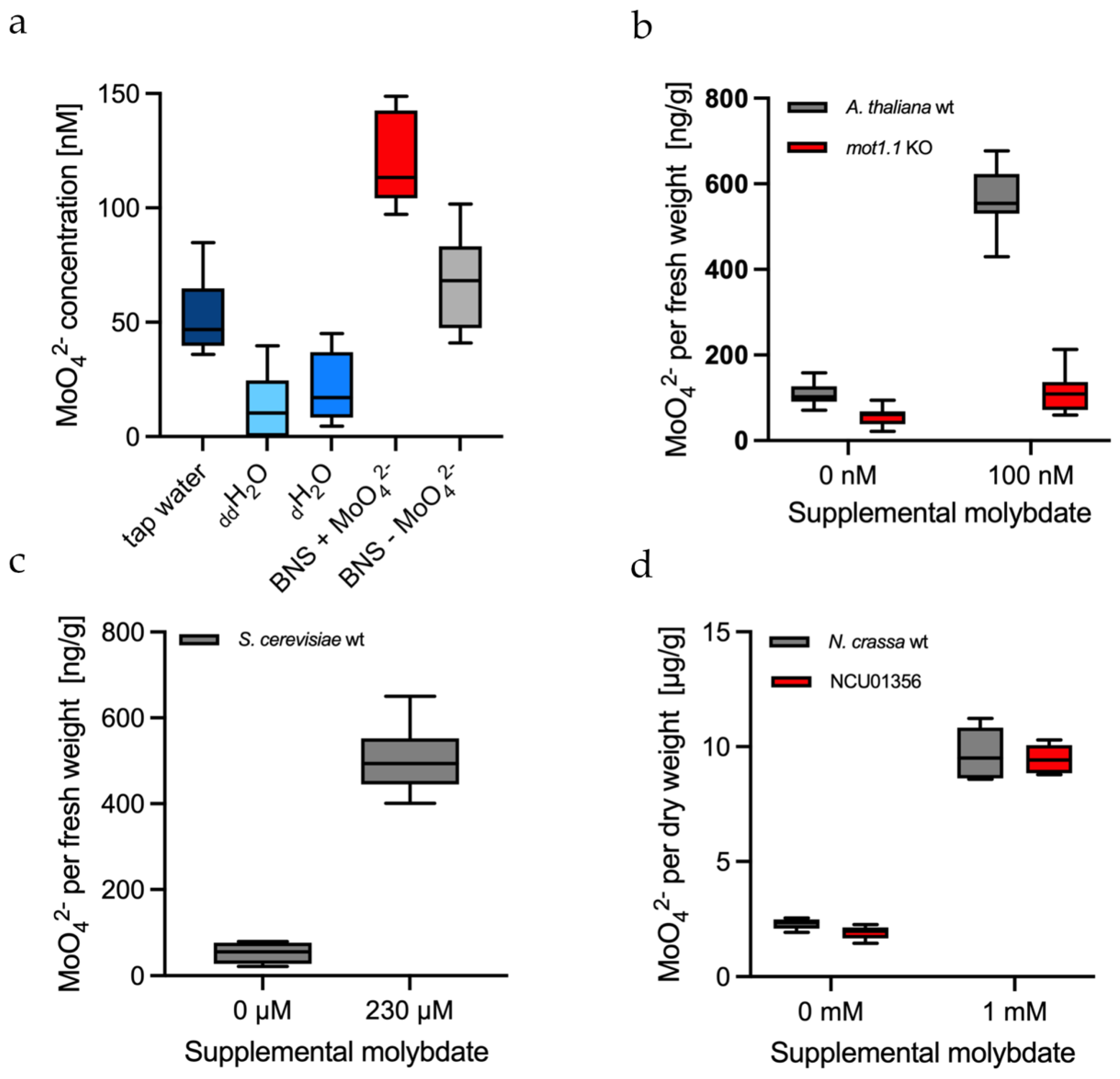
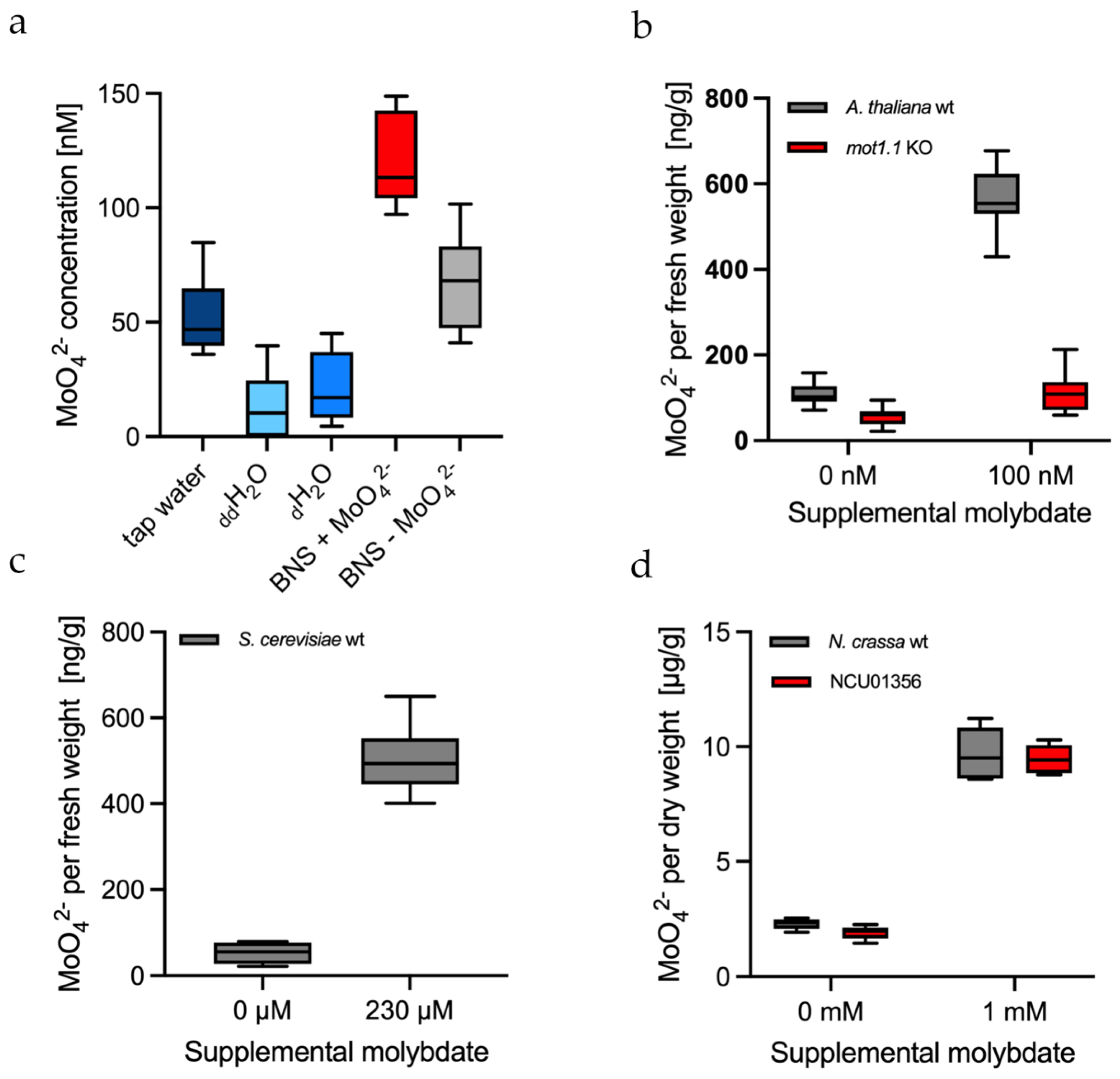
2.6. Measuring the Background of Molybdate in Water and Culture Media
2.7. In Vitro Molybdate Quantification in the Plant Arabidopsis Thaliana
2.8. In Vitro Molybdate Quantification in the Yeast Saccharomyces Cerevisiae
2.9. In Vitro Molybdate Quantification in the Fungus Neurospora Crassa
3. Discussion
4. Materials and Methods
4.1. Strains and Materials
4.2. Genetical Construction of MolyProbe-Strep
4.3. E. coli Expression and Purification of Recombinant MolyProbe-Strep
4.4. Purification Analysis of MolyProbe-Strep
4.5. MolyProbe In Vitro Assay
4.6. pH Dependency of MolyProbe In Vitro Assay
4.7. Neurospora Crassa Preparation
4.8. Saccharomyces Cerevisiae Preparation
4.9. Arabidopsis Thaliana Preparation
4.10. Data Analysis
4.11. Structural Prediction Using ColabFold
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Sample Availability
References
- Schwarz, G.; Mendel, R.R.; Ribbe, M.W. Molybdenum cofactors, enzymes and pathways. Nature 2009, 460, 839–847. [Google Scholar] [CrossRef] [PubMed]
- Mendel, R.R.; Bittner, F. Cell biology of molybdenum. Biochim. Biophys. Acta 2006, 1763, 621–635. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mendel, R.R.; Hercher, T.W.; Zupok, A.; Hasnat, M.A.; Leimkühler, S. The requirement of inorganic Fe-S clusters for the biosynthesis of the organometallic molybdenum cofactor. Inorganics 2020, 8, 43. [Google Scholar] [CrossRef]
- Stiefel, E.I. The biogeochemistry of molybdenum and tungsten. Met. Ions Biol. Syst. 2002, 39, 1–29. [Google Scholar] [CrossRef]
- Pau, R.N.; Lawson, D.M. Transport, homeostasis, regulation, and binding of molybdate and Tungstate to proteins. Met. Ions. Biol. Syst. 2002, 39, 31–74. [Google Scholar] [CrossRef]
- Lindsay, W. Chemical Equilibria in Soils; The Blackburn Press: Caldwell, NJ, USA, 1979; p. 449. [Google Scholar]
- Gupta, U.C. Molybdenum in Agriculture; Cambridge University Press: New York, NY, USA, 2009. [Google Scholar]
- Schiavon, M.; Pittarello, M.; Pilon-Smits, E.A.H.; Wirtz, M.; Hell, R.; Malagoli, M. Selenate and molybdate alter sulfate transport and assimilation in Brassica juncea L. Czern.: Implications for phytoremediation. Environ. Exp. Bot. 2012, 75, 41–51. [Google Scholar] [CrossRef]
- Allaway, W.H. The effect of soils and fertilizers on human and animal nutrition. Agric. Inf. Bull. 1975, 378, 21. [Google Scholar] [CrossRef]
- Kovacs, B.; Puskas-Preszner, A.; Huzsvai, L.; Levai, L.; Bodi, E. Effect of molybdenum treatment on molybdenum concentration and nitrate reduction in maize seedlings. Plant Physiol. Biochem. 2015, 96, 38–44. [Google Scholar] [CrossRef] [Green Version]
- Medicine, I.O. Dietary Reference Intakes for Vitamin A, Vitamin K, Arsenic, Boron, Chromium, Copper, Iodine, Iron, Manganese, Molybdenum, Nickel, Silicon, Vanadium, and Zinc, Dietary Reference Intakes for Vitamin A, Vitamin K, Arsenic, Boron, Chromium, Copper, Iodine, Iron, Manganese, Molybdenum, Nickel, Silicon, Vanadium, and Zinc; National Academies Press (US): Washington, DC, USA, 2001; Volume 11, pp. 420–439. [Google Scholar] [CrossRef] [Green Version]
- Kar, P.; Seel, M.; Weidemann, T.; Hofinger, S. Theoretical mimicry of biomembranes. FEBS Lett. 2009, 583, 1909–1915. [Google Scholar] [CrossRef] [Green Version]
- Li, X.; Luu, D.T.; Maurel, C.; Lin, J. Probing plasma membrane dynamics at the single-molecule level. Trends Plant Sci. 2013, 18, 617–624. [Google Scholar] [CrossRef]
- Brown, B. Biological Membranes; The Biochemical Society: London, UK, 1996. [Google Scholar]
- Poppe, J.; Brunle, S.; Hail, R.; Wiesemann, K.; Schneider, K.; Ermler, U. The molybdenum storage protein: A soluble ATP hydrolysis-dependent molybdate pump. FEBS J. 2018, 285, 4602–4616. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tejada-Jimenez, M.; Llamas, A.; Sanz-Luque, E.; Galvan, A.; Fernandez, E. A high-affinity molybdate transporter in eukaryotes. Proc. Natl. Acad. Sci. USA 2007, 104, 20126–20130. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tejada-Jimenez, M.; Chamizo-Ampudia, A.; Galvan, A.; Fernandez, E.; Llamas, A. Molybdenum metabolism in plants. Metallomics 2013, 5, 1191–1203. [Google Scholar] [CrossRef] [PubMed]
- Tomatsu, H.; Takano, J.; Takahashi, H.; Watanabe-Takahashi, A.; Shibagaki, N.; Fujiwara, T. An Arabidopsis thaliana high-affinity molybdate transporter required for efficient uptake of molybdate from soil. Proc. Natl. Acad. Sci. USA 2007, 104, 18807–18812. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gao, J.S.; Wu, F.F.; Shen, Z.L.; Meng, Y.; Cai, Y.P.; Lin, Y. A putative molybdate transporter LjMOT1 is required for molybdenum transport in Lotus japonicus. Physiol. Plant 2016, 158, 331–340. [Google Scholar] [CrossRef]
- Quistgaard, E.M.; Low, C.; Guettou, F.; Nordlund, P. Understanding transport by the major facilitator superfamily (MFS): Structures pave the way. Nat. Rev. Mol. Cell Biol. 2016, 17, 123–132. [Google Scholar] [CrossRef]
- Gonzalez-Guerrero, M.; Escudero, V.; Saez, A.; Tejada-Jimenez, M. Transition metal transport in plants and associated endosymbionts: Arbuscular mycorrhizal fungi and rhizobia. Front. Plant Sci. 2016, 7, 1088. [Google Scholar] [CrossRef] [Green Version]
- Nakanishi, Y.; Iida, S.; Ueoka-Nakanishi, H.; Niimi, T.; Tomioka, R.; Maeshima, M. Exploring dynamics of molybdate in living animal cells by a genetically encoded FRET nanosensor. PLoS ONE 2013, 8, e58175. [Google Scholar] [CrossRef]
- Tejada-Jimenez, M.; Galvan, A.; Fernandez, E. Algae and humans share a molybdate transporter. Proc. Natl. Acad. Sci. USA 2011, 108, 6420–6425. [Google Scholar] [CrossRef] [Green Version]
- Schüttelkopf, A.W.; Boxer, D.H.; Hunter, W.N. Crystal structure of activated mode reveals conformational changes involving both oxyanion and DNA-binding domains. J. Mol. Biol. 2003, 326, 761–767. [Google Scholar] [CrossRef]
- Ormo, M.; Cubitt, A.B.; Kallio, K.; Gross, L.A.; Tsien, R.Y.; Remington, S.J. Crystal structure of the aequorea victoria green fluorescent protein. Science 1996, 273, 1392–1395. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Anderson, L.A.; Palmer, T.; Price, N.C.; Bornemann, S.; Boxer, D.H.; Pau, R.N. Characterisation of the molybdenum-responsive ModE regulatory protein and its binding to the promoter region of the modABCD (molybdenum transport) operon of Escherichia coli. Eur. J. Biochem. 1997, 246, 119–126. [Google Scholar] [CrossRef] [PubMed]
- Gasber, A.; Klaumann, S.; Trentmann, O.; Trampczynska, A.; Clemens, S.; Schneider, S.; Sauer, N.; Feifer, I.; Bittner, F.; Mendel, R.R.; et al. Identification of an arabidopsis solute carrier critical for intracellular transport and inter-organ allocation of molybdate. Plant Biol. 2011, 13, 710–718. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Gladyshev, V.N. Molybdoproteomes and evolution of molybdenum utilization. J. Mol. Biol. 2008, 379, 881–899. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Webster, J.; Weber, R.W.S. Introduction to Fungi; Cambridge University Press: Cambridge, UK, 2007. [Google Scholar]
- Bischof, H.; Burgstaller, S.; Waldeck-Weiermair, M.; Rauter, T.; Schinagl, M.; Ramadani-Muja, J.; Graier, W.F.; Malli, R. Live-Cell imaging of physiologically relevant metal ions using genetically encoded FRET-based probes. Cells 2019, 8, 492. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maslanka, R.; Kwolek-Mirek, M.; Zadrag-Tecza, R. Autofluorescence of yeast saccharomyces cerevisiae cells caused by glucose metabolism products and its methodological implications. J. Microbiol. Methods 2018, 146, 55–60. [Google Scholar] [CrossRef]
- Donaldson, L. Autofluorescence in plants. Molecules 2020, 25, 2393. [Google Scholar] [CrossRef] [PubMed]
- Ide, Y.; Kusano, M.; Oikawa, A.; Fukushima, A.; Tomatsu, H.; Saito, K.; Hirai, M.Y.; Fujiwara, T. Effects of molybdenum deficiency and defects in molybdate transporter MOT1 on transcript accumulation and nitrogen/sulphur metabolism in Arabidopsis thaliana. J. Exp. Bot. 2011, 62, 1483–1497. [Google Scholar] [CrossRef]
- Betolngar, D.B.; Erard, M.; Pasquier, H.; Bousmah, Y.; Diop-Sy, A.; Guiot, E.; Vincent, P.; Merola, F. pH sensitivity of FRET reporters based on cyan and yellow fluorescent proteins. Anal. Bioanal. Chem. 2015, 407, 4183–4193. [Google Scholar] [CrossRef]
- Sikorski, R.S.; Hieter, P. A system of shuttle vectors and yeast host strains designed for efficient manipulation of DNA in Saccharomyces ceratisia. Genet. Soc. Am. 1989, 122, 19–27. [Google Scholar]
- Ringel, P.; Probst, C.; Dammeyer, T.; Buchmeier, S.; Jansch, L.; Wissing, J.; Tinnefeld, P.; Mendel, R.R.; Jockusch, B.M.; Kruse, T. Enzymatic characterization of recombinant nitrate reductase expressed and purified from Neurospora crassa. Fungal Genet. Biol. 2015, 80, 10–18. [Google Scholar] [CrossRef]
- Wajmann, S.; Hercher, T.W.; Buchmeier, S.; Hansch, R.; Mendel, R.R.; Kruse, T. The first step of neurospora crassa molybdenum cofactor biosynthesis: Regulatory aspects under N-Derepressing and nitrate-inducing conditions. Microorganisms 2020, 8, 534. [Google Scholar] [CrossRef] [Green Version]
- Vogel, H.J. A convenient growth medium for neurospora (Medium N). Microb. Genet. Bull. 1956, 13, 42–43. [Google Scholar]
- Conn, S.J.; Hocking, B.; Dayod, M.; Xu, B.; Athman, A.; Henderson, S.; Aukett, L.; Conn, V.; Shearer, M.K.; Fuentes, S.; et al. Protocol: Optimising hydroponic growth systems for nutritional and physiological analysis of Arabidopsis thaliana and other plants. Plant Methods 2013, 9, 4. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mirdita, M.; Schütze, K.; Moriwaki, Y.; Heo, L.; Ovchinnikov, S.; Steinegger, M. ColabFold—Making protein folding accessible to all. bioRxiv 2022. [Google Scholar] [CrossRef] [PubMed]
- Pettersen, E.F.; Goddard, T.D.; Huang, C.C.; Meng, E.C.; Couch, G.S.; Croll, T.I.; Morris, J.H.; Ferrin, T.E. UCSF ChimeraX: Structure visualization for researchers, educators, and developers. Protein Sci. 2021, 30, 70–82. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Organism | Strain Name | Genotype | Origin |
|---|---|---|---|
| Saccharomyces cerevisiae | YPH499 | ura3-52, lys2-801(amber), ade2-101(ochre), trp1-delta63, his3-delta200, leu2-delta1 | Sikorski and Hieter, 1989 [35] |
| Neurospora crassa | wild type NCU01356 | 74-OR23-1V; mat A ncu01356::hph; mat a | FGSC |
| Arabidopsis thaliana | wild type mot1.1 KO | Columbia-0 (Col-0) SALK_118311 | TU Braunschweig NASC |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Oliphant, K.D.; Karger, M.; Nakanishi, Y.; Mendel, R.R. Precise Quantification of Molybdate In Vitro by the FRET-Based Nanosensor ‘MolyProbe’. Molecules 2022, 27, 3691. https://doi.org/10.3390/molecules27123691
Oliphant KD, Karger M, Nakanishi Y, Mendel RR. Precise Quantification of Molybdate In Vitro by the FRET-Based Nanosensor ‘MolyProbe’. Molecules. 2022; 27(12):3691. https://doi.org/10.3390/molecules27123691
Chicago/Turabian StyleOliphant, Kevin D., Marius Karger, Yoichi Nakanishi, and Ralf R. Mendel. 2022. "Precise Quantification of Molybdate In Vitro by the FRET-Based Nanosensor ‘MolyProbe’" Molecules 27, no. 12: 3691. https://doi.org/10.3390/molecules27123691
APA StyleOliphant, K. D., Karger, M., Nakanishi, Y., & Mendel, R. R. (2022). Precise Quantification of Molybdate In Vitro by the FRET-Based Nanosensor ‘MolyProbe’. Molecules, 27(12), 3691. https://doi.org/10.3390/molecules27123691