
Tetrabromobisphenol A Disturbs Brain Development in Both Thyroid Hormone-Dependent and -Independent Manners in Xenopus laevis

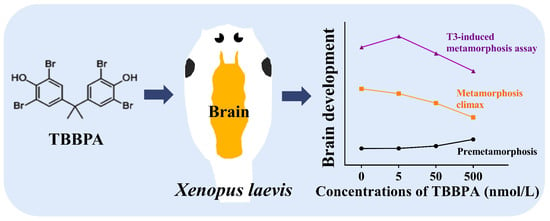
Abstract
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Results
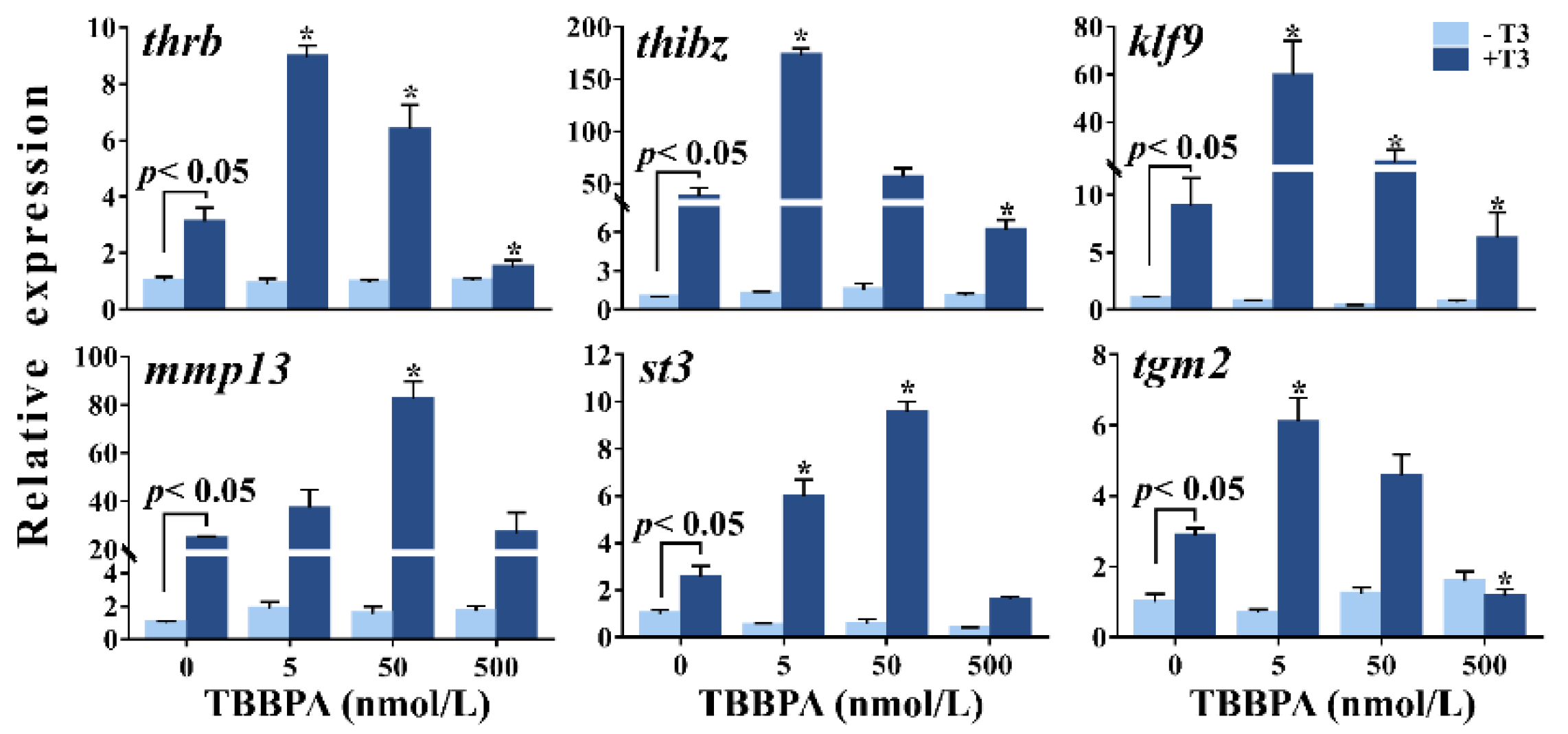
2.1. TBBPA Affects T3-Induced TH-Response Gene Expression in Xenopus laevis Brains
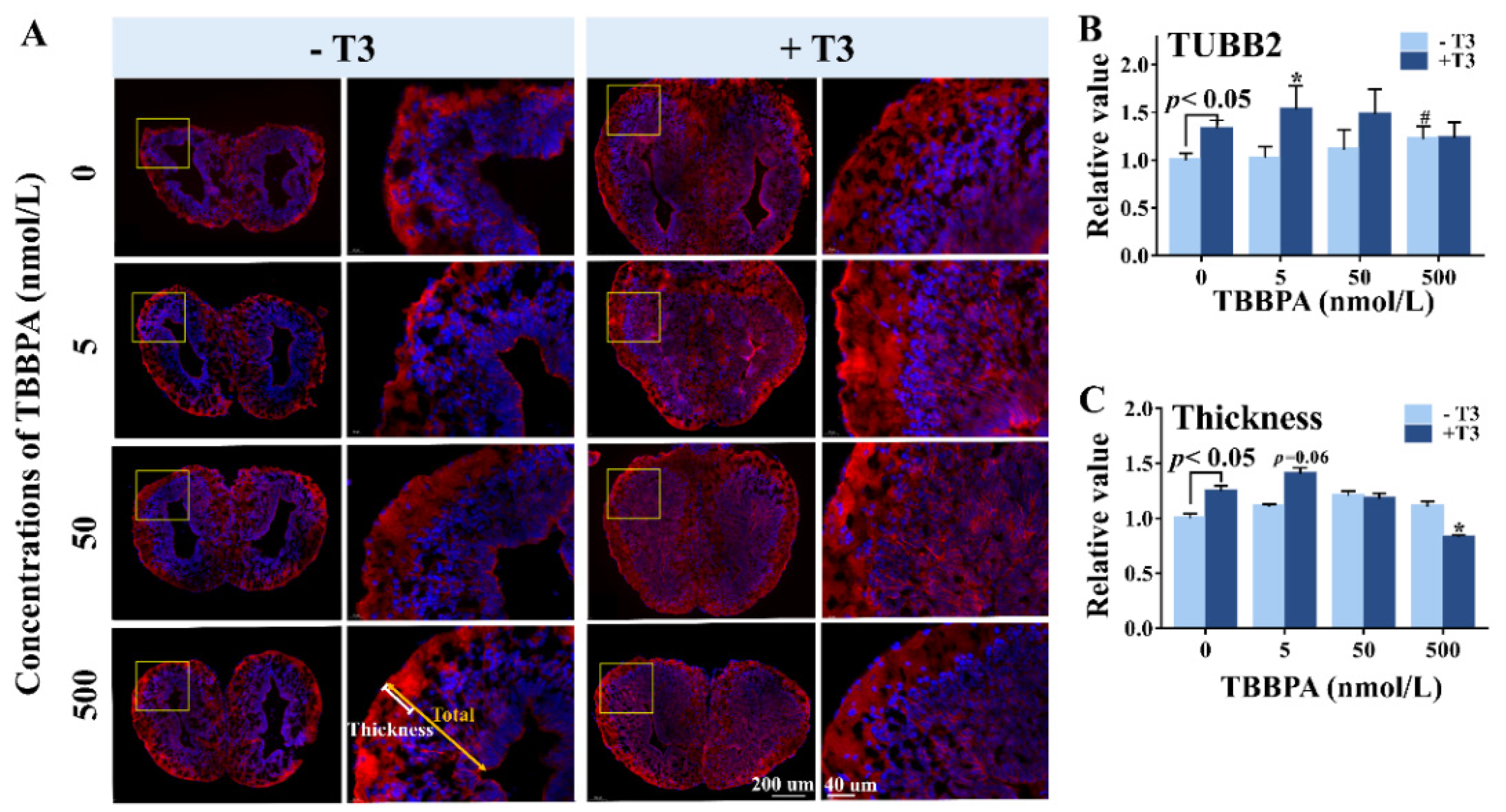
2.2. TBBPA Alters Brain Morphology in the Absence and Presence of T3
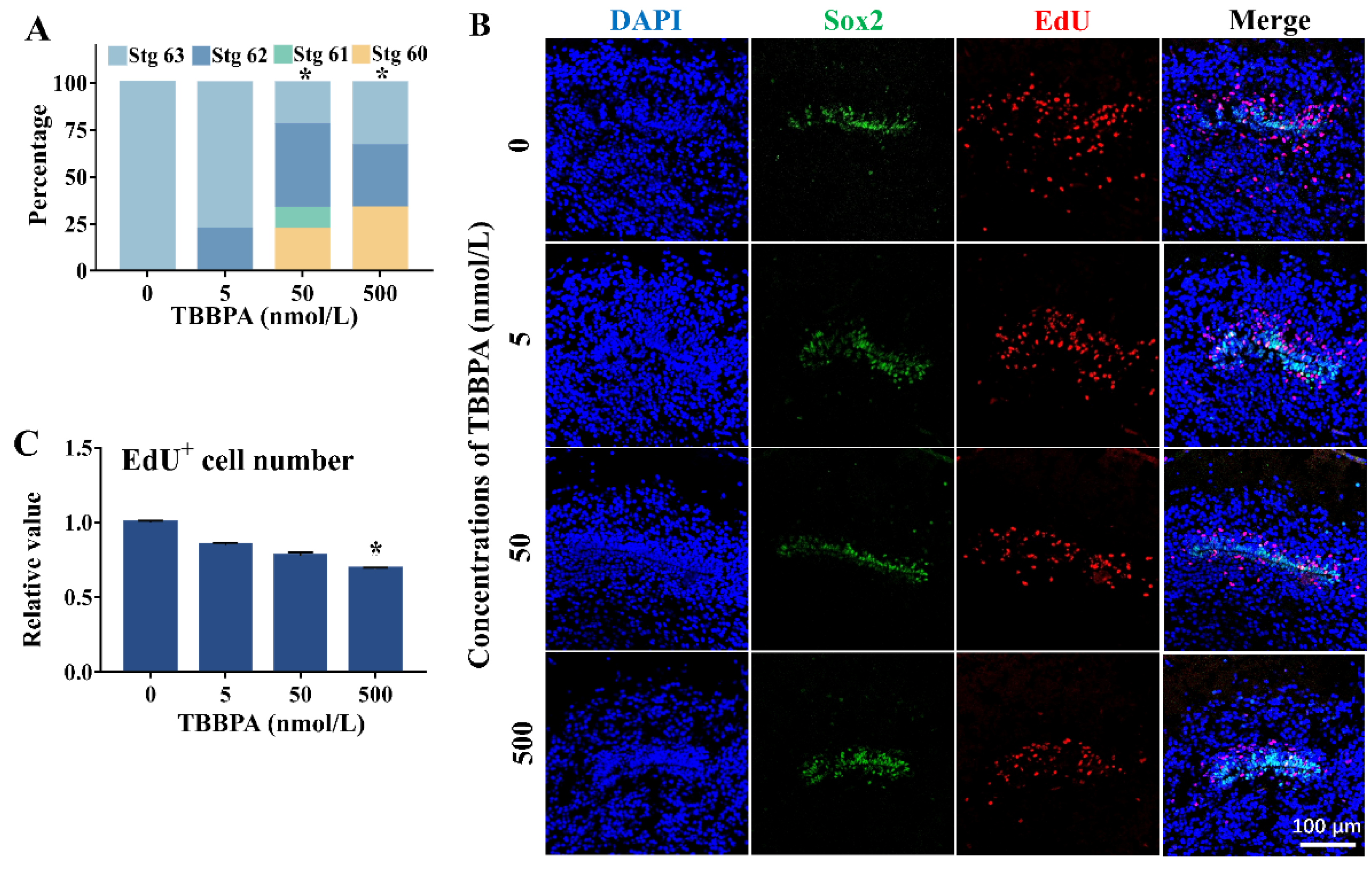
2.3. TBBPA Affects Cell Proliferation in the Telencephalon in the Presence of T3
2.4. TBBPA Affects Neurodifferentiation in the Telencephalon in the Presence of T3
2.5. TBBPA Disturbs Brain Development in Spontaneous Metamorphosis Assay
3. Discussion
4. Materials and Methods
4.1. Chemicals
4.2. Animal Housing
4.3. T3-Induced Xenopus Metamorphosis Assay (TIXMA)
4.4. Spontaneous Metamorphosis Assay
4.5. RNA Extraction and RT-qPCR Analysis
4.6. Analysis for Gross Morphology
4.7. IF Staining
4.8. EdU Proliferation Assay
4.9. Data Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Malkoske, T.; Tang, Y.L.; Xu, W.Y.; Yu, S.L.; Wang, H.T. A review of the environmental distribution, fate, and control of tetrabromobisphenol A released from sources. Sci. Total Environ. 2016, 569–570, 1608–1617. [Google Scholar] [CrossRef] [PubMed]
- Yang, S.W.; Wang, S.R.; Liu, H.L.; Yan, Z.G. Tetrabromobisphenol A: Tissue distribution in fish, and seasonal variation in water and sediment of Lake Chaohu, China. Environ. Sci. Pollut. Res. Int. 2012, 19, 4090–4096. [Google Scholar] [CrossRef] [PubMed]
- Cariou, R.; Antignac, J.P.; Zalko, D.; Berrebi, A.; Cravedi, J.P.; Maume, D.; Marchand, P.; Monteau, F.; Riu, A.; Andre, F.; et al. Exposure assessment of French women and their newborns to tetrabromobisphenol-A: Occurrence measurements in maternal adipose tissue, serum, breast milk and cord serum. Chemosphere 2008, 73, 1036–1041. [Google Scholar] [CrossRef] [PubMed]
- Li, A.J.; Zhuang, T.F.; Shi, W.; Liang, Y.; Liao, C.Y.; Song, M.Y.; Jiang, G.B. Serum concentration of bisphenol analogues in pregnant women in China. Sci. Total Environ. 2020, 707, 136100. [Google Scholar] [CrossRef] [PubMed]
- Shi, Z.X.; Jiao, Y.; Hu, Y.; Sun, Z.W.; Zhou, X.Q.; Feng, J.F.; Li, J.G.; Wu, Y.N. Levels of tetrabromobisphenol A, hexabromocyclododecanes and polybrominated diphenyl ethers in human milk from the general population in Beijing, China. Sci. Total Environ. 2013, 452–453, 10–18. [Google Scholar] [CrossRef] [PubMed]
- European Chemicals Bureau. European Union Risk Assessment Report-2,2′,6,6′-Tetrabromo-4,4′-Isopropylidenediphenol (Tetrabromobisphenol-A or TBBP-A) (CAS: 79-94-7) Part-Human Health, II-Human Health; European Chemicals Bureau: Luxembourg, 2006. [Google Scholar]
- EFSA. Scientific Opinion on Tetrabromobisphenol A (TBBPA) and its derivatives in food. EFSA J. 2011, 9. [Google Scholar]
- Yu, Y.J.; Yu, Z.L.; Chen, H.B.; Han, Y.J.; Xiang, M.D.; Chen, X.C.; Ma, R.X.; Wang, Z.D. Tetrabromobisphenol A: Disposition, kinetics and toxicity in animals and humans. Environ. Pollut. 2019, 253, 909–917. [Google Scholar] [CrossRef] [PubMed]
- Lu, L.; Zhan, T.; Ma, M.; Xu, C.; Wang, J.; Zhang, C.; Liu, W.; Zhuang, S. Correction to “Thyroid Disruption by Bisphenol S Analogues via Thyroid Hormone Receptor β: In Vitro, In Vivo, and Molecular Dynamics Simulation Study”. Environ. Sci. Technol. 2018, 52, 6617–6625. [Google Scholar] [CrossRef]
- Mengeling, B.J.; Wei, Y.; Dobrawa, L.N.; Streekstra, M.; Louisse, J.; Singh, V.; Singh, L.; Lein, P.J.; Wulff, H.; Murk, A.J.; et al. A multi-tiered, in vivo, quantitative assay suite for environmental disruptors of thyroid hormone signaling. Aquat. Toxicol. 2017, 190, 1–10. [Google Scholar] [CrossRef]
- Fini, J.B.; Le Mével, S.; Palmier, K.; Darras, V.M.; Punzon, I.; Richardson, S.J.; Clerget-Froidevaux, M.S.; Demeneix, B.A. Thyroid Hormone Signaling in the Xenopus laevis Embryo Is Functional and Susceptible to Endocrine Disruption. Endocrinol. 2012, 153, 5068–5081. [Google Scholar] [CrossRef]
- Kitamura, S.; Kato, T.; Iida, M.; Jinno, N.; Suzuki, T.; Ohta, S.; Fujimoto, N.; Hanada, H.; Kashiwagi, K.; Kashiwagi, A. Anti-thyroid hormonal activity of tetrabromobisphenol A, a flame retardant, and related compounds: Affinity to the mammalian thyroid hormone receptor, and effect on tadpole metamorphosis. Life Sci. 2005, 76, 1589–1601. [Google Scholar] [CrossRef]
- Zhang, Y.F.; Xu, W.; Lou, Q.Q.; Li, Y.Y.; Zhao, Y.X.; Wei, W.J.; Qin, Z.F.; Wang, H.L.; Li, J.Z. Tetrabromobisphenol A disrupts vertebrate development via thyroid hormone signaling pathway in a developmental stage-dependent manner. Environ. Sci. Technol. 2014, 48, 8227–8234. [Google Scholar] [CrossRef]
- Kacew, S.; Hayes, A.W. Absence of neurotoxicity and lack of neurobehavioral consequences due to exposure to tetrabromobisphenol A (TBBPA) exposure in humans, animals and zebrafish. Arch. Toxicol. 2020, 94, 59–66. [Google Scholar] [CrossRef]
- Chen, J.F.; Tanguay, R.L.; Simonich, M.; Nie, S.F.; Zhao, Y.X.; Li, L.L.; Bai, C.L.; Dong, Q.X.; Huang, C.J.; Lin, K.F. TBBPA chronic exposure produces sex-specific neurobehavioral and social interaction changes in adult zebrafish. Neurotoxicol. Teratol. 2016, 56, 9–15. [Google Scholar] [CrossRef]
- Crump, D.; Werry, K.; Veldhoen, N.; Van Aggelen, G.; Helbing, C.C. Exposure to the herbicide acetochlor alters thyroid hormone-dependent gene expression and metamorphosis in Xenopus laevis. Environ. Health Perspect. 2002, 110, 1199–1205. [Google Scholar] [CrossRef]
- Morvan-Dubois, G.; Demeneix, B.A.; Sachs, L.M. Xenopus laevis as a model for studying thyroid hormone signalling: From development to metamorphosis. Mol. Cell Endocrinol. 2008, 293, 71–79. [Google Scholar] [CrossRef][Green Version]
- Tata, J.R. Amphibian metamorphosis as a model for the developmental actions of thyroid hormone. Mol. Cell. Endocrinol 2006, 246, 10–20. [Google Scholar] [CrossRef]
- Yao, X.F.; Chen, X.Y.; Zhang, Y.F.; Li, Y.Y.; Wang, Y.; Zheng, Z.M.; Qin, Z.F.; Zhang, Q.D. Optimization of the T3-induced Xenopus metamorphosis assay for detecting thyroid hormone signaling disruption of chemicals. J. Environ. Sci. 2017, 52, 314–324. [Google Scholar] [CrossRef]
- Zhu, M.; Chen, X.Y.; Li, Y.Y.; Yin, N.Y.; Faiola, F.; Qin, Z.F.; Wei, W.J. Bisphenol F Disrupts Thyroid Hormone Signaling and Postembryonic Development in Xenopus laevis. Environ. Sci. Technol. 2018, 52, 1602–1611. [Google Scholar] [CrossRef] [PubMed]
- Niu, Y.; Zhu, M.; Dong, M.Q.; Li, J.B.; Li, Y.Y.; Xiong, Y.M.; Liu, P.Y.; Qin, Z.F. Bisphenols disrupt thyroid hormone (TH) signaling in the brain and affect TH-dependent brain development in Xenopus laevis. Aquat. Toxicol. 2021, 237, 105902. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Li, Y.Y.; Qin, Z.F.; Wei, W.J. Re-evaluation of thyroid hormone signaling antagonism of tetrabromobisphenol A for validating the T3-induced Xenopus metamorphosis assay. J. Environ. Sci. 2017, 52, 325–332. [Google Scholar] [CrossRef] [PubMed]
- D’Amico, L.A.; Boujard, D.; Coumailleau, P. Proliferation, migration and differentiation in juvenile and adult Xenopus laevis brains. Brain Res. 2011, 1405, 31–48. [Google Scholar] [CrossRef]
- Sun, H.; Shen, O.X.; Wang, X.R.; Zhou, L.; Zhen, S.Q.; Chen, X.D. Anti-thyroid hormone activity of bisphenol A, tetrabromobisphenol A and tetrachlorobisphenol A in an improved reporter gene assay. Toxicol. Vitro 2009, 23, 950–954. [Google Scholar] [CrossRef]
- Wong, J.; Shi, Y.-B. Coordinated regulation of and transcriptional activation by Xenopus thyroid hormone and retinoid X receptors. J. Biol. Chem. 1995, 270, 18479–18483. [Google Scholar] [CrossRef]
- Shi, Y.-B.; Wong, J.; Puzianowska-Kuznicka, M.; Stolow, M.A. Tadpole competence and tissue-specific temporal regulation of amphibian metamorphosis: Roles of thyroid hormone and its receptors. Bioessays 1996, 18, 391–399. [Google Scholar] [CrossRef] [PubMed]
- Zhang, R.; Engler, A.; Taylor, V. Notch: An interactive player in neurogenesis and disease. Cell Tissue Res. 2018, 371, 73–89. [Google Scholar] [CrossRef] [PubMed]
- Yin, N.Y.; Liang, S.J.; Liang, S.X.; Yang, R.J.; Hu, B.W.; Qin, Z.F.; Liu, A.F.; Faiola, F. TBBPA and Its Alternatives Disturb the Early Stages of Neural Development by Interfering with the NOTCH and WNT Pathways. Environ. Sci. Technol. 2018, 52, 5459–5468. [Google Scholar] [CrossRef] [PubMed]
- Liang, S.J.; Liang, S.X.; Zhou, H.; Yin, N.Y.; Faiola, F. Typical halogenated flame retardants affect human neural stem cell gene expression during proliferation and differentiation via glycogen synthase kinase 3 beta and T3 signaling. Ecotoxicol. Environ. Saf. 2019, 183, 109498. [Google Scholar] [CrossRef]
- Horn, S.; Heuer, H. Thyroid hormone action during brain development: More questions than answers. Mol. Cell Endocrinol. 2010, 315, 19–26. [Google Scholar] [CrossRef] [PubMed]
- Rovet, J.F. The role of thyroid hormones for brain development and cognitive function. Endocr. Dev. 2014, 26, 26–43. [Google Scholar] [CrossRef] [PubMed]
- Fini, J.B.; Anne, R.; Laurent, D.; Anne, H.; Le, M.S.; Sylvie, C. Parallel biotransformation of tetrabromobisphenol A in Xenopus laevis and mammals: Xenopus as a model for endocrine perturbation studies. Toxicol. Sci. 2012, 125, 359–367. [Google Scholar] [CrossRef]
- Liu, H.L.; Ma, Z.Y.; Tao, Z.; Yu, N.Y.; Su, G.Y.; Giesy, J.P.L.; Yu, H.X. Pharmacokinetics and effects of tetrabromobisphenol a (TBBPA) to early life stages of zebrafish (Danio rerio). Chemosphere 2018, 190, 243–252. [Google Scholar] [CrossRef]
- Hakk, H.; Letcher, R.J. Metabolism in the toxicokinetics and fate of brominated flame retardants—A review. Environ. Int. 2003, 29, 801–828. [Google Scholar] [CrossRef]
- Dong, M.Q.; Li, Y.Y.; Zhu, M.; Qin, Z.F. Tetrabromobisphenol A: A neurotoxicant or not? Environ. Sci. Pollut. Res. Int. 2021, 28, 54466–54476. [Google Scholar] [CrossRef] [PubMed]
- Cai, M.; Li, Y.Y.; Zhu, M.; Li, J.B.; Qin, Z.F. Evaluation of the effects of low concentrations of bisphenol AF on gonadal development using the Xenopus laevis model: A finding of testicular differentiation inhibition coupled with feminization. Environ. Pollut. 2020, 260, 113980. [Google Scholar] [CrossRef] [PubMed]
- Nieuwkoop, P.; Faber, J. Normal Table of Xenopus Laevis (Daudin); Garland Publishing: New York, NY, USA, 1994; p. 252. [Google Scholar]
- Shi, Y.B.; Liang, C.T. Cloning and characterization of the ribosomal protein L8 gene from Xenopus laevis. Biochim. Biophys. Acta 1994, 1217, 227–228. [Google Scholar] [CrossRef]
- Gunderson, M.P.; Veldhoen, N.; Skirrow, R.C.; Macnab, M.K.; Ding, W.; van Aggelen, G.; Helbing, C.C. Effect of low dose exposure to the herbicide atrazine and its metabolite on cytochrome P450 aromatase and steroidogenic factor-1 mRNA levels in the brain of premetamorphic bullfrog tadpoles (Rana catesbeiana). Aquat. Toxicol. 2011, 102, 31–38. [Google Scholar] [CrossRef]
- Hogan, N.S.; Crump, K.L.; Duarte, P.; Lean, D.R.; Trudeau, V.L. Hormone cross-regulation in the tadpole brain: Developmental expression profiles and effect of T3 exposure on thyroid hormone- and estrogen-responsive genes in Rana pipiens. Gen. Comp. Endocrinol. 2007, 154, 5–15. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)) Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Dong, M.; Li, Y.; Zhu, M.; Li, J.; Qin, Z. Tetrabromobisphenol A Disturbs Brain Development in Both Thyroid Hormone-Dependent and -Independent Manners in Xenopus laevis. Molecules 2022, 27, 249. https://doi.org/10.3390/molecules27010249
Dong M, Li Y, Zhu M, Li J, Qin Z. Tetrabromobisphenol A Disturbs Brain Development in Both Thyroid Hormone-Dependent and -Independent Manners in Xenopus laevis. Molecules. 2022; 27(1):249. https://doi.org/10.3390/molecules27010249
Chicago/Turabian StyleDong, Mengqi, Yuanyuan Li, Min Zhu, Jinbo Li, and Zhanfen Qin. 2022. "Tetrabromobisphenol A Disturbs Brain Development in Both Thyroid Hormone-Dependent and -Independent Manners in Xenopus laevis" Molecules 27, no. 1: 249. https://doi.org/10.3390/molecules27010249
APA StyleDong, M., Li, Y., Zhu, M., Li, J., & Qin, Z. (2022). Tetrabromobisphenol A Disturbs Brain Development in Both Thyroid Hormone-Dependent and -Independent Manners in Xenopus laevis. Molecules, 27(1), 249. https://doi.org/10.3390/molecules27010249