
Antiviral Properties of the NSAID Drug Naproxen Targeting the Nucleoprotein of SARS-CoV-2 Coronavirus

 , , , ,
, , , ,  ,
,  and
and 
Abstract
1. Introduction
2. Results
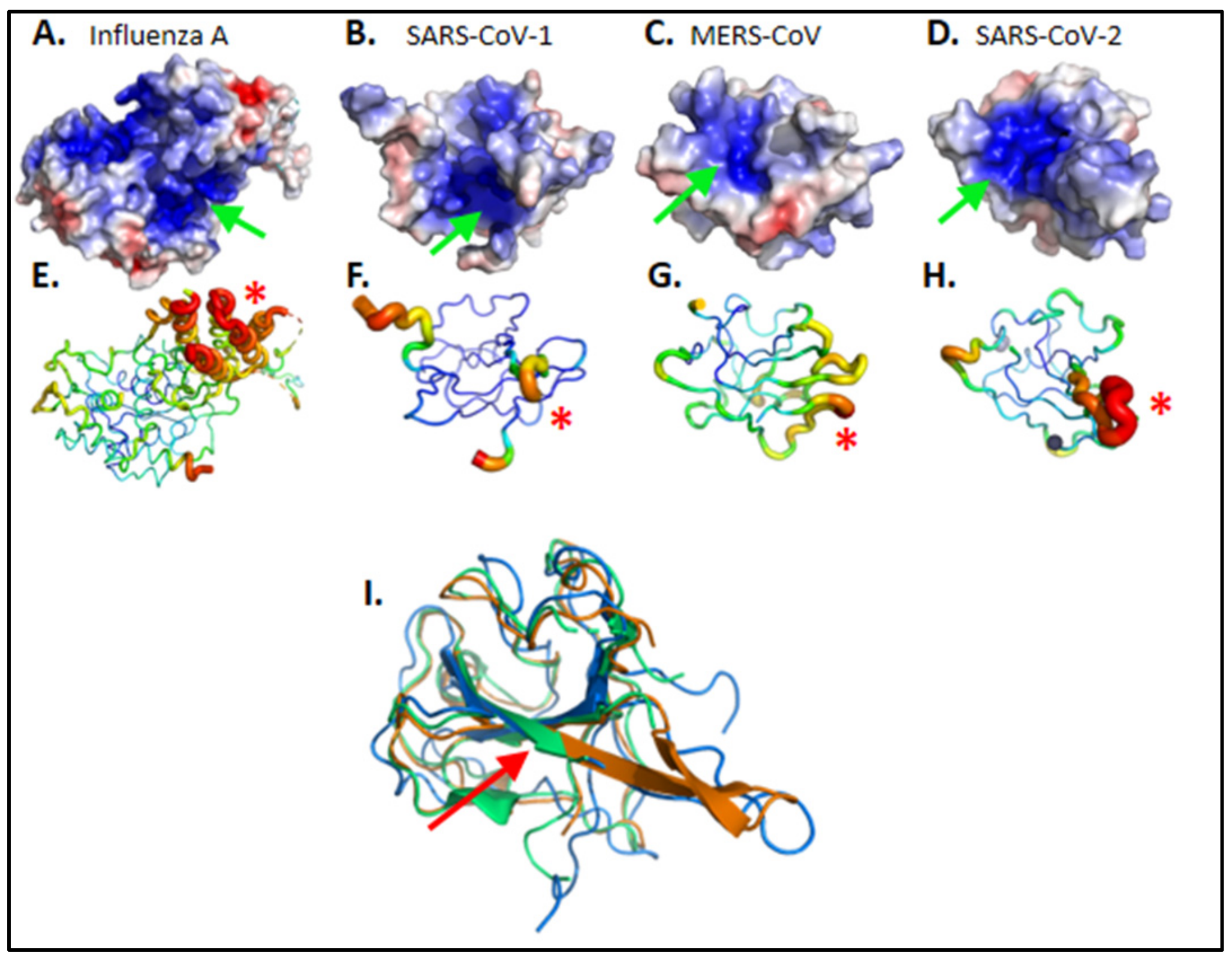
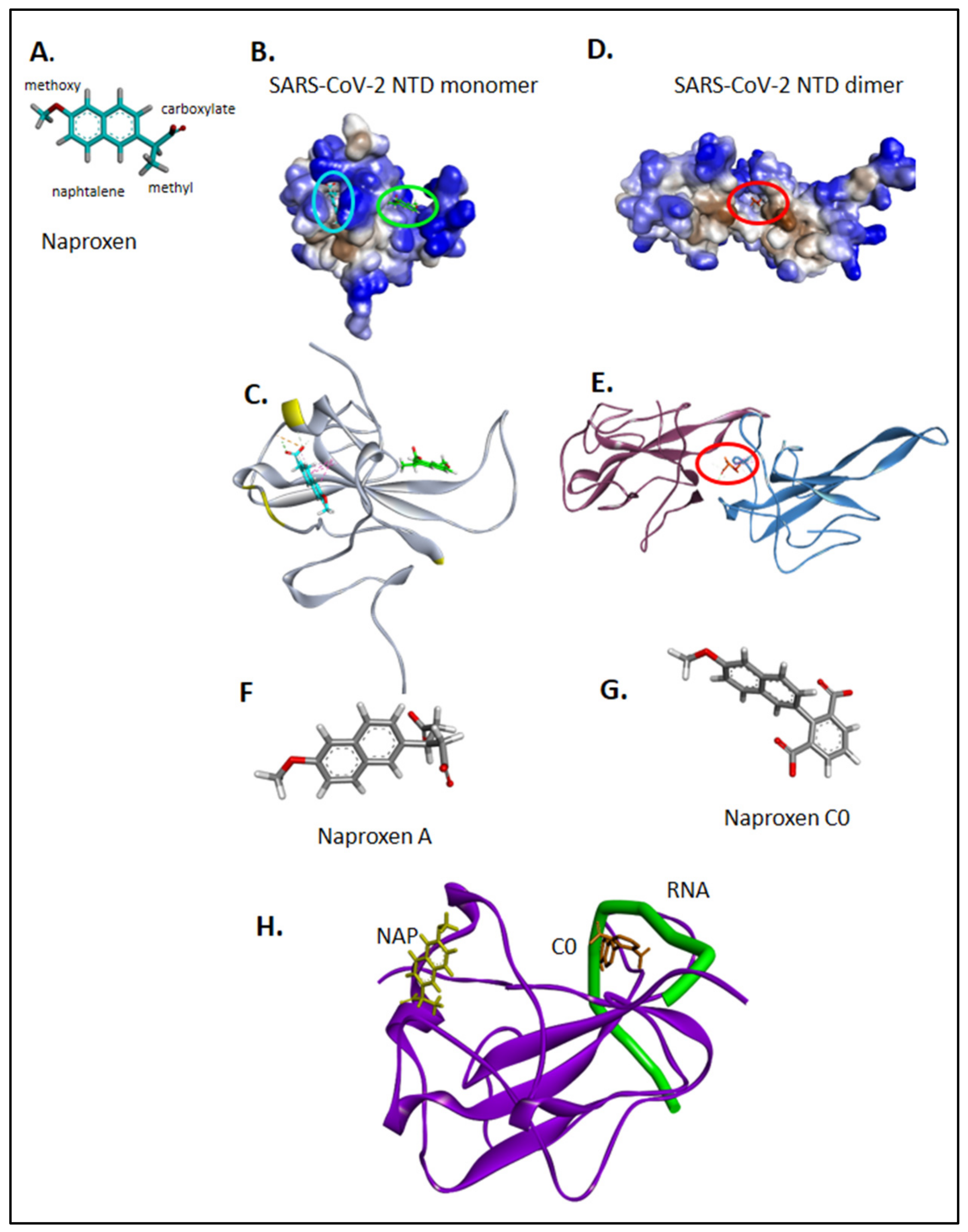
2.1. Structure-Based Modeling of Naproxen Binding to the Nucleoprotein of SARS-CoV-2
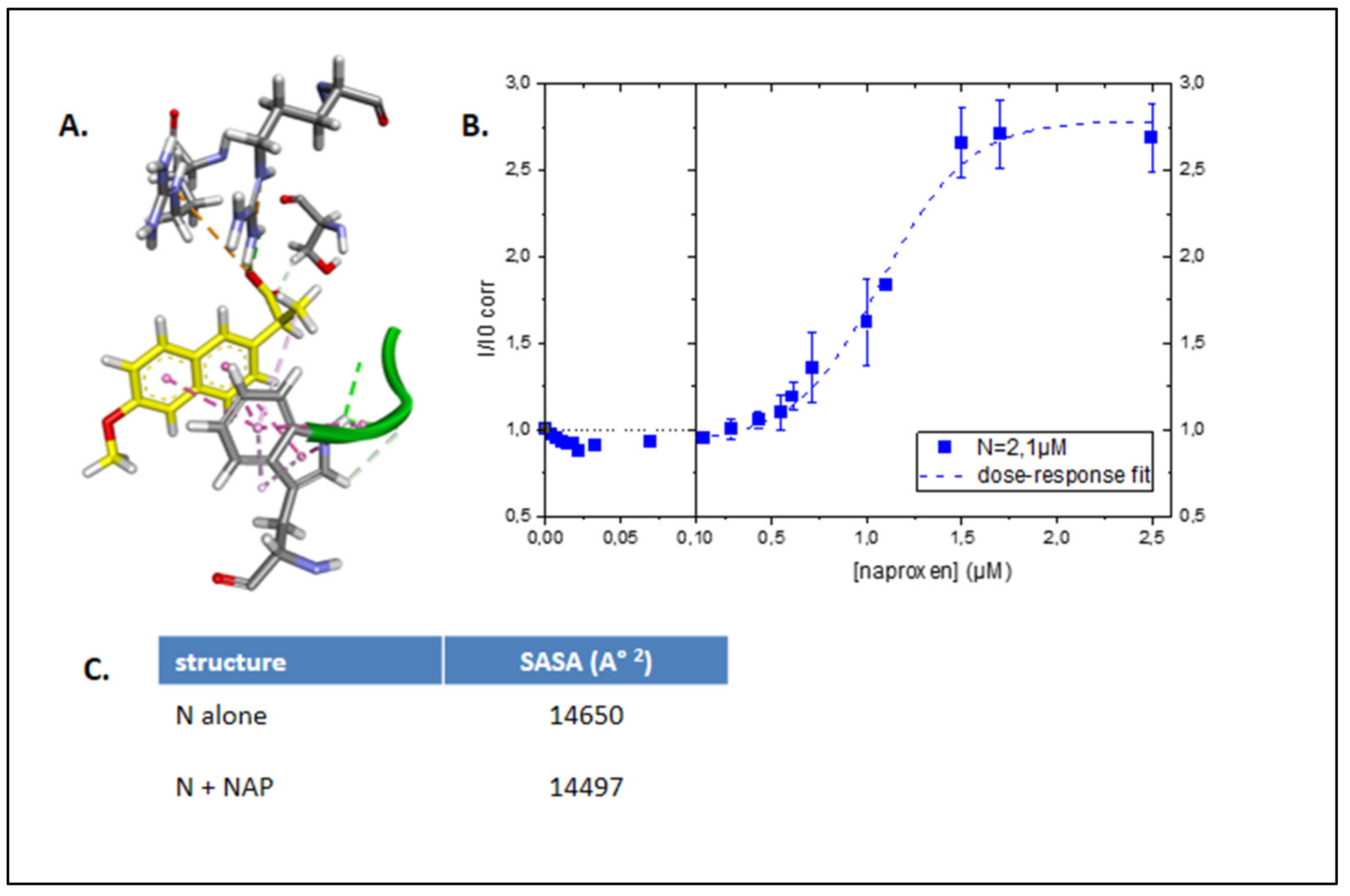
2.2. Evidence for In Vitro Binding of Naproxen to Recombinant N-Terminal Domain of SARS-CoV-2 N
2.2.1. Fluorescence Assay
2.2.2. Thermal Shift Assay Monitored by Dynamic Light Scattering (DLS)
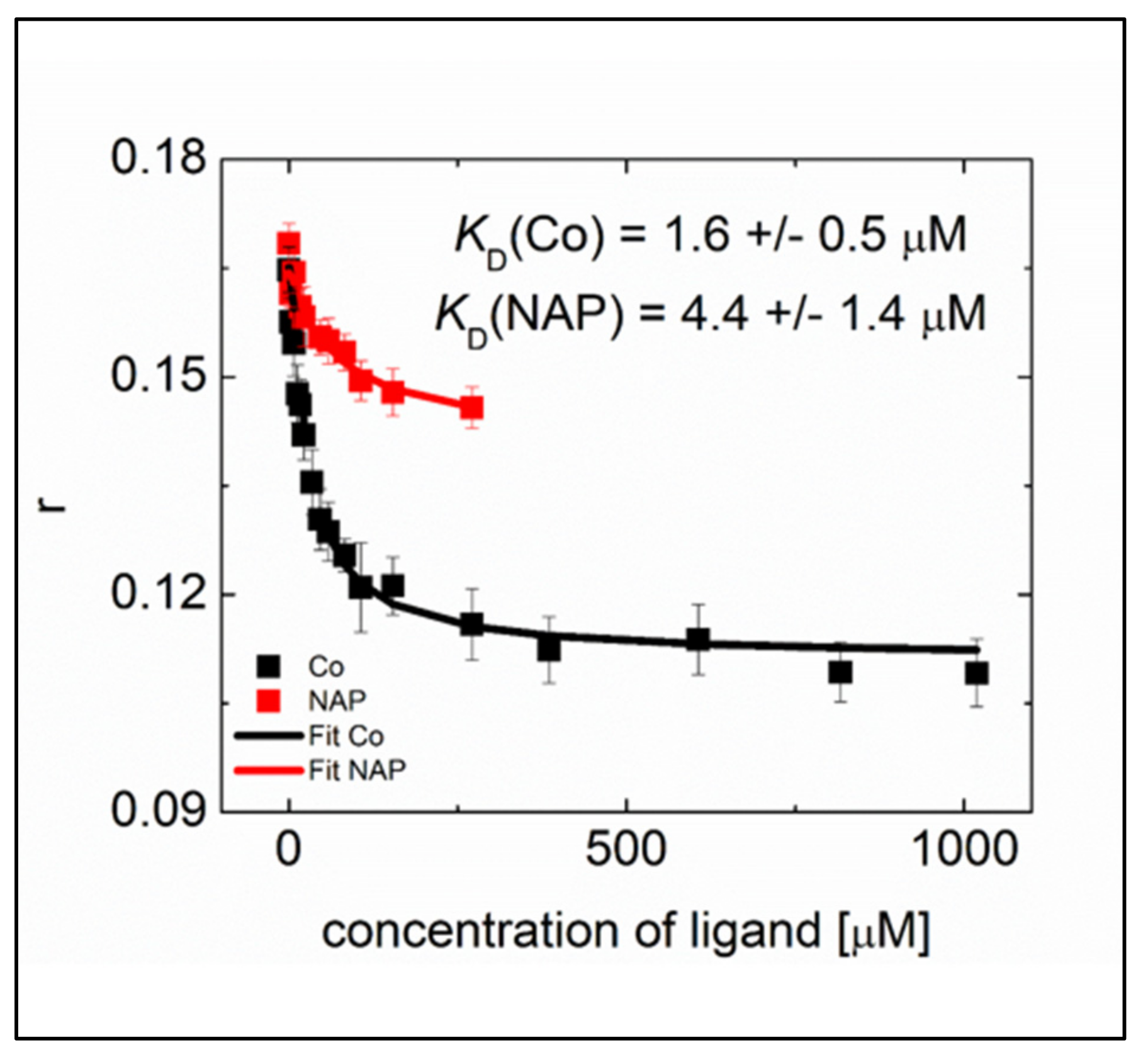
2.2.3. Competition on N NTD Binding of RNA and the Ligands Evidenced by Fluorescence Anisotropy
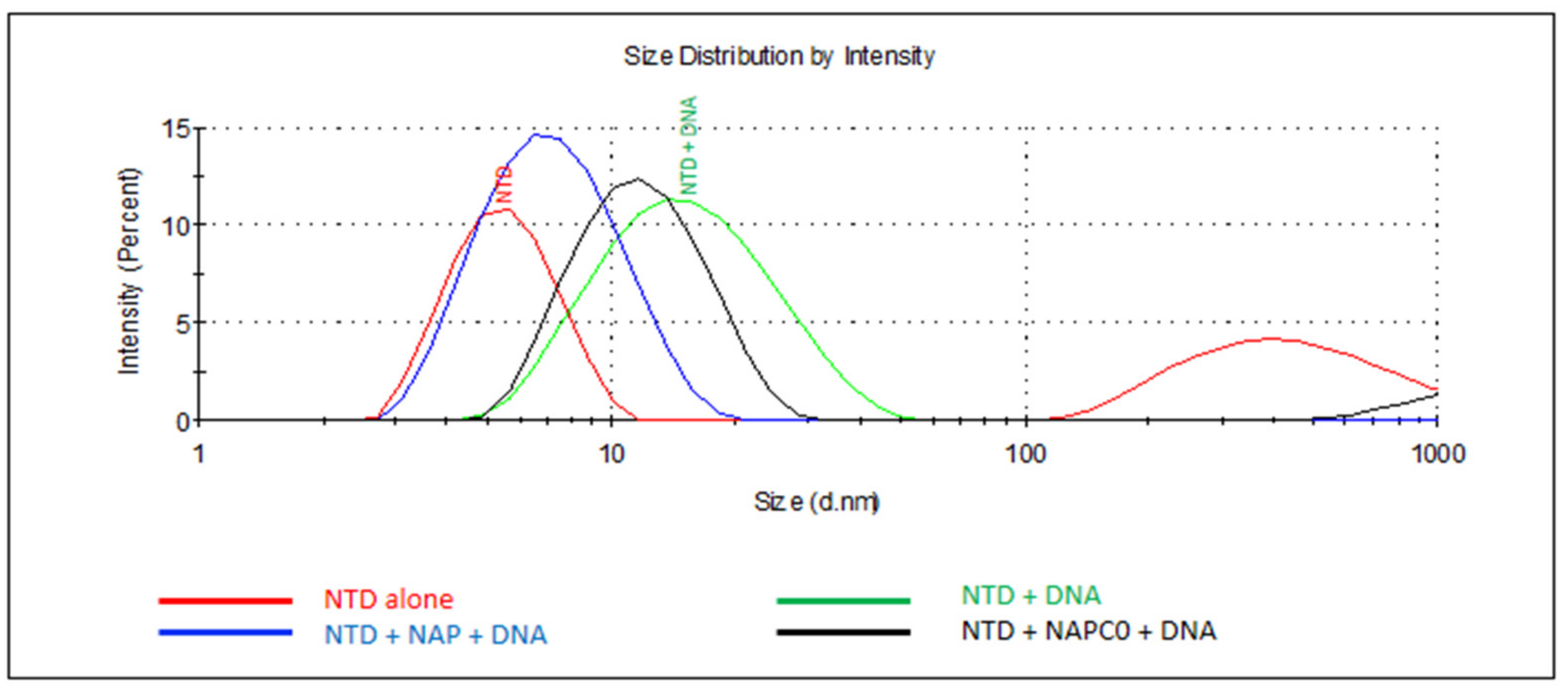
2.2.4. N Oligomerization Assay Monitored by DLS in the Presence of Nucleic Acids and the Effect of Various Ligands Thereof
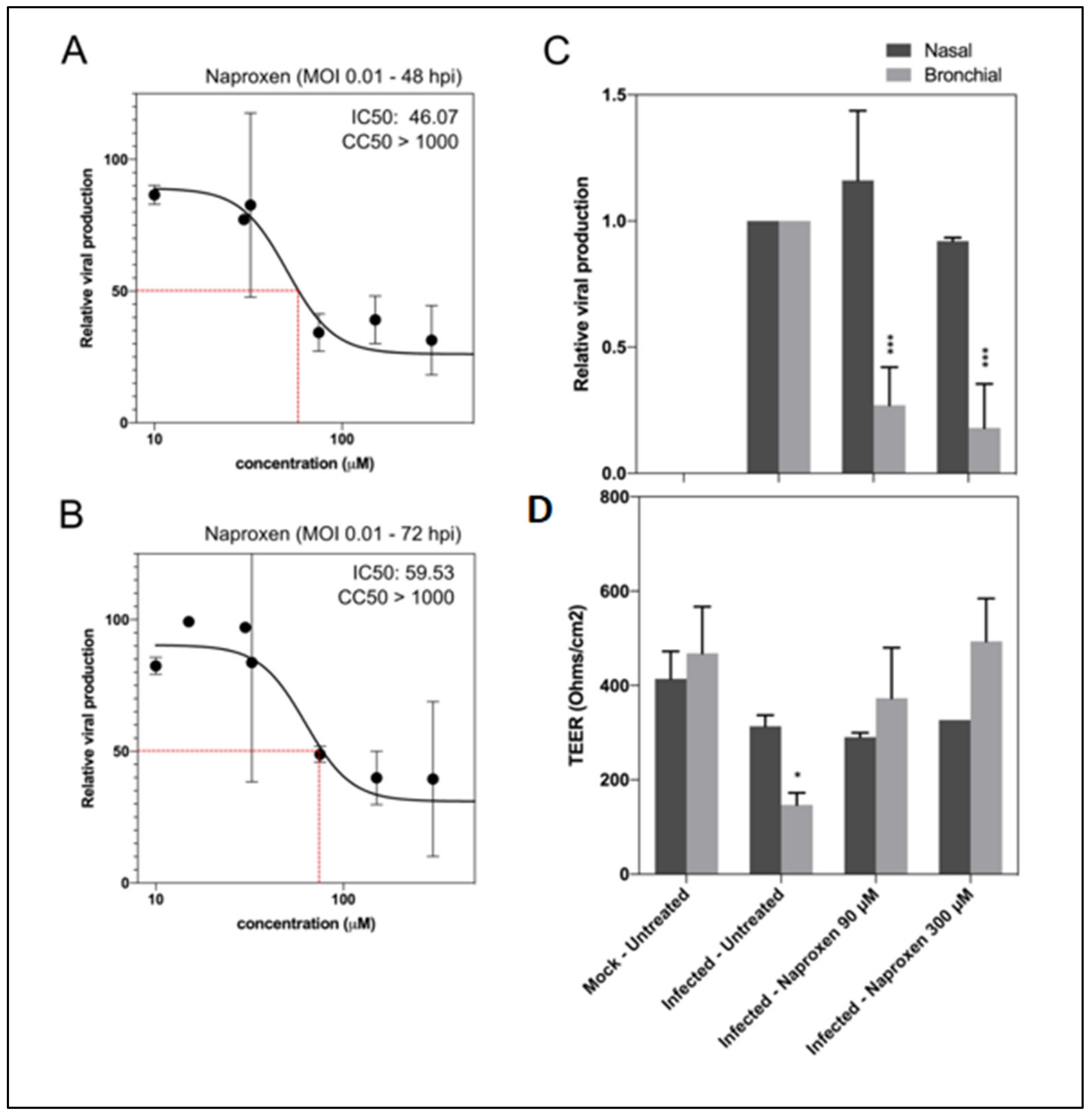
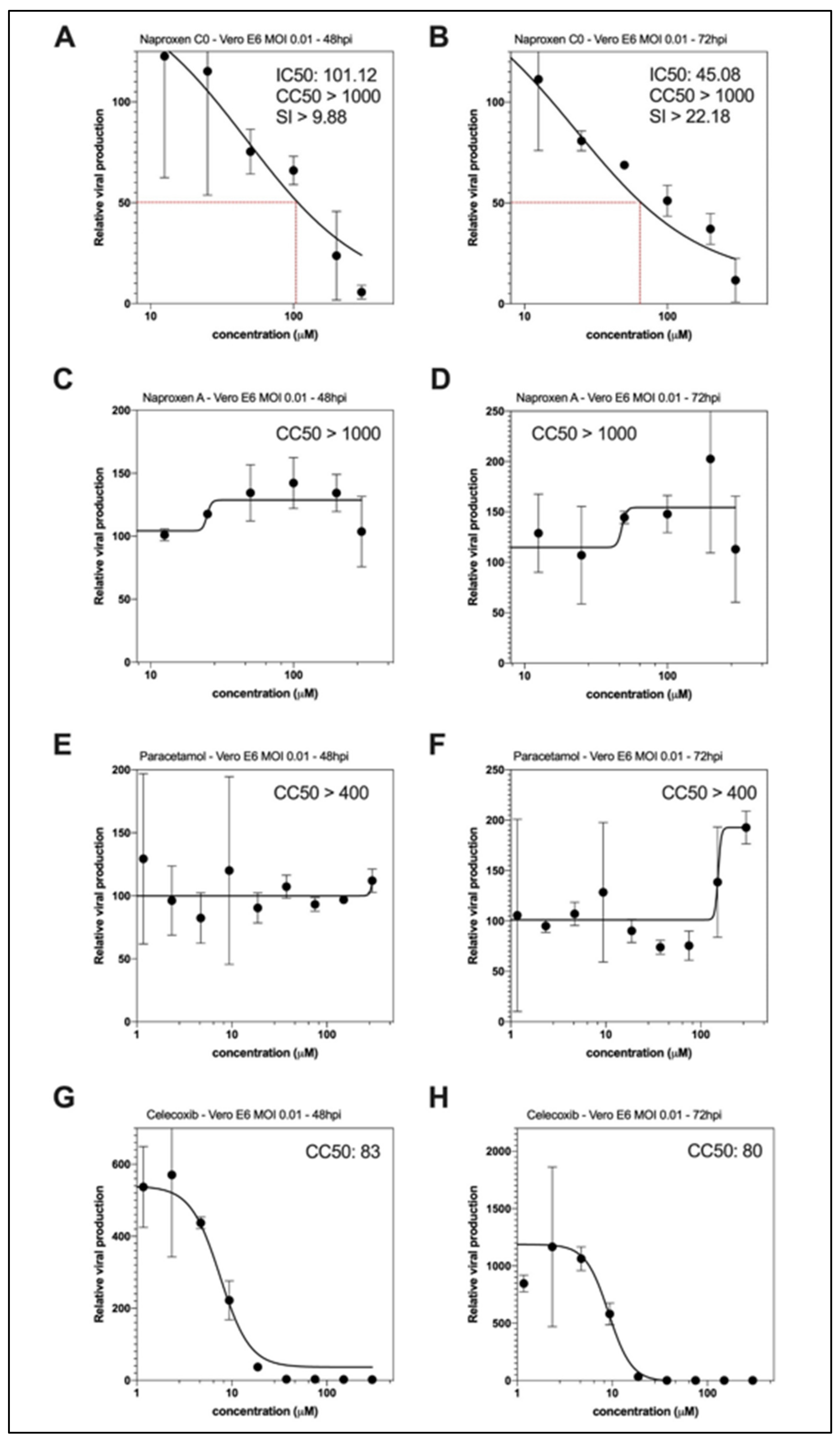
2.3. Naproxen Inhibits SARS-CoV-2 Infection in Vero E6 Cells and in Reconstituted Human Airway Epithelia (HAE)
3. Discussion
4. Methods
4.1. Modeling
4.2. Materials
4.3. Size-Exclusion Chromatography
4.4. Fluorescence Measurements
4.5. Fluorescence Anisotropy
4.6. Dynamic Light Scattering Measurements
4.7. Virus
4.8. Dose–Response Antiviral Evaluation in Vero E6 Cells
4.9. Evaluation of Antiviral Activity in Reconstituted Human Airway Epithelia (HAE)
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Sample Availability
References
- Huang, C.; Wang, Y.; Li, X.; Ren, L.; Zhao, J.; Hu, Y.; Zhang, L.; Fan, G.; Xu, J.; Gu, X.; et al. Clinical features of patients infected with 2019 novel coronavirus in Wuhan, China. Lancet 2020, 395, 497–506. [Google Scholar] [CrossRef]
- Zhou, F.; Yu, T.; Du, R.; Fan, G.; Liu, Y.; Liu, Z.; Xiang, J.; Wang, Y.; Song, B.; Gu, X.; et al. Clinical course and risk factors for mortality of adult inpatients with COVID-19 in Wuhan, China: A retrospective cohort study. Lancet 2020, 395, 1054–1062. [Google Scholar] [CrossRef]
- Fukuyama, S.; Kawaoka, Y. The pathogenesis of influenza virus infections: The contributions of virus and host factors. Curr. Opin. Immunol. 2011, 23, 481–486. [Google Scholar] [CrossRef]
- Horby, P.; Lim, W.S.; Emberson, J.R.; Mafham, M.; Bell, J.L.; Linsell, L.; Staplin, N.; Brightling, C.; Ustianowski, A.; Elmahi, E.; et al. Dexamethasone in Hospitalized Patients with Covid-19—Preliminary Report. N. Engl. J. Med. 2020. [Google Scholar] [CrossRef]
- Stebbing, J.; Phelan, A.; Griffin, I.; Tucker, C.; Oechsle, O.; Smith, D.; Richardson, P. COVID-19: Combining antiviral and anti-inflammatory treatments. Lancet Infect. Dis. 2020, 20, 400–402. [Google Scholar] [CrossRef]
- Verrall, G.M. Scientific Rationale for a Bottom-Up Approach to Target the Host Response in Order to Try and Reduce the Numbers Presenting With Adult Respiratory Distress Syndrome Associated With COVID-19. Is There a Role for Statins and COX-2 Inhibitors in the Prevention and Early Treatment of the Disease? Front. Immunol. 2020, 11, 2167. [Google Scholar] [CrossRef]
- Chen, J.S.; Alfajaro, M.M.; Chow, R.D.; Wei, J.; Filler, R.B.; Eisenbarth, S.C.; Wilen, C.B. Non-steroidal anti-inflammatory drugs dampen the cytokine and antibody response to SARS-CoV-2 infection. J. Virol. 2021. [Google Scholar] [CrossRef] [PubMed]
- Prasher, P.; Sharma, M.; Gunupuru, R. Targeting cyclooxygenase enzyme for the adjuvantCOVID-19 therapy. Drug Dev. Res. 2021. [Google Scholar] [CrossRef] [PubMed]
- Ong, S.W.X.; Tan, W.Y.T.; Chan, Y.; Fong, S.; Renia, L.; Ng, L.F.; Leo, Y.; Lye, D.C.; Young, B.E. Safety and potential efficacy of cyclooxygenase-2 inhibitors in coronavirus disease 2019. Clin. Transl. Immunol. 2020, 9, e1159. [Google Scholar] [CrossRef]
- Castro, V.M.; Ross, R.A.; McBride, S.M.; Perlis, R.H. Brief Report: Identifying common pharmacotherapies associated with reduced COVID-19 morbidity using electronic health records. MedRxiv 2020. [Google Scholar] [CrossRef]
- Lejal, N.; Tarus, B.; Bouguyon, E.; Chenavas, S.; Bertho, N.; Delmas, B.; Ruigrok, R.W.H.; Di Primo, C.; Slama-Schwok, A. Structure-Based Discovery of the Novel Antiviral Properties of Naproxen against the Nucleoprotein of Influenza A Virus. Antimicrob. Agents Chemother. 2013, 57, 2231–2242. [Google Scholar] [CrossRef]
- Tarus, B.; Bertrand, H.; Zedda, G.; Di Primo, C.; Quideau, S.; Slama-Schwok, A. Structure-based design of novel naproxen de-rivatives targeting monomeric nucleoprotein of Influenza A virus. J. Biomol. Struct. Dyn. 2015, 33, 1899–1912. [Google Scholar] [CrossRef] [PubMed]
- Dilly, S.; Fotso Fotso, A.; Lejal, N.; Zedda, G.; Chebbo, M.; Rahman, F. From Naproxen Repurposing to Naproxen Ana-logues and Their Antiviral Activity against Influenza A Virus. J. Med. Chem. 2018, 61, 7202–7217. [Google Scholar] [CrossRef] [PubMed]
- Hu, Y.; Li, W.; Gao, T.; Cui, Y.; Jin, Y.; Li, P.; Ma, Q.; Liu, X.; Cao, C. The Severe Acute Respiratory Syndrome Coronavirus Nucleocapsid Inhibits Type I Interferon Production by Interfering with TRIM25-Mediated RIG-I Ubiquitination. J. Virol. 2017, 91, e02143-16. [Google Scholar] [CrossRef] [PubMed]
- Mu, J.; Xu, J.; Zhang, L.; Shu, T.; Wu, D.; Huang, M. SARS-CoV-2-encoded nucleocapsid protein acts as a viral suppressor of RNA interference in cells. Sci. China Life Sci. 2020, 63, 1–4. [Google Scholar] [CrossRef]
- Gordon, D.E.; Jang, G.M.; Bouhaddou, M.; Xu, J.; Obernier, K.; White, K.M. A SARS-CoV-2 protein interaction map reveals targets for drug repurposing. Nature 2020, 583, 459–468. [Google Scholar] [CrossRef] [PubMed]
- Tung, H.Y.L.; Limtung, P. Mutations in the phosphorylation sites of SARS-CoV-2 encoded nucleocapsid protein and struc-ture model of sequestration by protein 14-3-3. Biophys. Res. Commun. 2020, 532, 134–138. [Google Scholar] [CrossRef]
- Mondal, A.; Potts, G.K.; Dawson, A.R.; Coon, J.J.; Mehle, A. Phosphorylation at the Homotypic Interface Regulates Nucleoprotein Oligomerization and Assembly of the Influenza Virus Replication Machinery. PLoS Pathog. 2015, 11, e1004826. [Google Scholar] [CrossRef]
- Yan, X.; Hao, Q.; Mu, Y.; Timani, K.A.; Ye, L.; Zhu, Y. Nucleocapsid protein of SARS-CoV activates the expression of cy-clooxygenase-2 by binding directly to regulatory elements for nuclear factor-kappa B and CCAAT/enhancer binding pro-tein. Int. J. Biochem. Cell Biol. 2006, 38, 1417–1428. [Google Scholar] [CrossRef]
- Chenavas, S.; Crepin, T.; Delmas, B.; Ruigrok, R.W.; Slama-Schwok, A. Influenza virus nucleoprotein: Structure, RNA binding, oligomerization and antiviral drug target. Future Microbiol. 2013, 8, 1537–1545. [Google Scholar] [CrossRef] [PubMed]
- Lin, S.M.; Lin, S.C.; Hsu, J.N.; Chang, C.K.; Chien, C.M.; Wang, Y.S. Structure-Based Stabilization of Non-native Pro-tein-Protein Interactions of Coronavirus Nucleocapsid Proteins in Antiviral Drug Design. J. Med. Chem. 2020, 63, 3131–3141. [Google Scholar] [CrossRef]
- Pizzorno, A.; Padey, B.; Terrier, O.; Rosa-Calatrava, M. Drug Repurposing Approaches for the Treatment of Influenza Viral Infection: Reviving Old Drugs to Fight Against a Long-Lived Enemy. Front. Immunol. 2019, 10, 531. [Google Scholar] [CrossRef]
- Lejal, N.; Truchet, S.; Bechor, E.; Bouguyon, E.; Khedkar, V.; Bertho, N.; Vidic, J.; Adenot, P.; Solier, S.; Pick, E.; et al. Turning off NADPH oxidase-2 by impeding p67 phox activation in infected mouse macrophages reduced viral entry and inflammation. Biochim. Biophys. Acta (BBA)-Gen. Subj. 2018, 1862, 1263–1275. [Google Scholar] [CrossRef] [PubMed]
- Tarus, B.; Bakowiez, O.; Chenavas, S.; Duchemin, L.; Estrozi, L.; Bourdieu, C.; Lejal, N.; Bernard, J.; Moudjou, M.; Chevalier, C.; et al. Oligomerization paths of the nucleoprotein of influenza A virus. Biochime 2012, 94, 776–785. [Google Scholar] [CrossRef] [PubMed]
- Chang, C.; Michalska, K.; Jedrzejczak, R.; Maltseva, N.; Endres, M.; Godzik, A.; Kim, Y.; Joachimiak, A. Crystal Structure of RNA Binding Domain of Nucleocapsid Phosphoprotein from SARS Coronavirus 2 to be Published. 2020. Available online: https://www.rcsb.org/structure/6VYO (accessed on 13 April 2021).
- Huang, Q.; Yu, L.; Petros, A.M.; Gunasekera, A.; Liu, Z.; Xu, N.; Hajduk, P.; Mack, J.; Fesik, S.W.; Olejniczak, E.T. Structure of the N-Terminal RNA-Binding Domain of the SARS CoV Nucleocapsid Protein. Biocheme 2004, 43, 6059–6063. [Google Scholar] [CrossRef]
- Kang, S.; Yang, M.; Hong, Z.; Zhang, L.; Huang, Z.; Chen, X. Crystal structure of SARS-CoV-2 nucleocapsid protein RNA binding domain reveals potential unique drug targeting sites. Acta Pharm. Sin. B 2020, 10, 1228–1238. [Google Scholar] [CrossRef] [PubMed]
- Dinesh, D.C.; Chalupska, D.; Silhan, J.; Koutna, E.; Nencka, R.; Veverka, V. Structural basis of RNA recognition by the SARS-CoV-2 nucleocapsid phosphoprotein. PLoS Pathog. 2020, 16, e1009100. [Google Scholar] [CrossRef] [PubMed]
- Tarus, B.; Chevalier, C.; Richard, C.A.; Delmas, B.; Di Primo, C.; Slama-Schwok, A. Molecular dynamics studies of the nucleo-protein of influenza A virus: Role of the protein flexibility in RNA binding. PLoS ONE 2012, 7, e30038. [Google Scholar] [CrossRef]
- Peng, Y.; Du, N.; Lei, Y.; Dorje, S.; Qi, J.; Luo, T.; Gao, G.F.; Song, H. Structures of the SARS-CoV-2 nucleocapsid and their perspectives for drug design. EMBO J. 2020, 39. [Google Scholar] [CrossRef]
- Zeng, W.; Liu, G.; Ma, H.; Zhao, D.; Yang, Y.; Liu, M. Biochemical characterization of SARS-CoV-2 nucleocapsid protein. Biochem. Biophys. Res. Commun. 2020, 527, 618–623. [Google Scholar] [CrossRef]
- Ye, Q.; West, A.M.V.; Silletti, S.; Corbett, K.D. Architecture and self-assembly of the SARS-CoV-2 nucleocapsid protein. Protein Sci. 2020, 29, 1890–1901. [Google Scholar] [CrossRef]
- Pizzorno, A.; Padey, B.; Julien, T.; Trouillet-Assant, S.; Traversier, A.; Errazuriz-Cerda, E.; Fouret, J.; Duboi, J.; Gaymard, A.; Dulière, V.; et al. Charac-terization and treatment of SARS-CoV-2 in nasal and bronchial human airway épithélium. Antivir. Res. 2020, 181, 104878. [Google Scholar] [CrossRef]
- Bar-On, Y.M.; Flamholz, A.; Phillips, R.; Milo, R. SARS-CoV-2 (COVID-19) by the numbers. Elife 2020, 9, e57309. [Google Scholar] [CrossRef]
- Rockx, B.; Kuiken, T.; Herfst, S.; Bestebroer, T.; Lamers, M.M.; Munnink, B.B.O.; De Meulder, D.; Van Amerongen, G.; Brand, J.V.D.; Okba, N.M.A.; et al. Comparative pathogenesis of COVID-19, MERS, and SARS in a nonhuman primate model. Science 2020, 368, 1012–1015. [Google Scholar] [CrossRef] [PubMed]
- Gordon, D.E.; Jang, G.M.; Bouhaddou, M.; Xu, J.; Obernier, K.; O’Meara, M.J. A SARS-CoV-2-Human Protein-Protein Inter-action Map Reveals Drug Targets and Potential Drug-Repurposing. bioRxiv 2020. [Google Scholar] [CrossRef]
- Rinott, E.; Kozer, E.; Shapira, Y.; Bar-Haim, A.; Youngster, I. Ibuprofen use and clinical outcomes in COVID-19 patients. Clin. Microbiol. Infect. 2020, 26, 1259.e5–1259.e7. [Google Scholar] [CrossRef]
- Robb, C.T.; Goepp, M.; Rossi, A.G.; Yao, C. Non-steroidal anti-inflammatory drugs, prostaglandins, and COVID-19. Br. J. Phar-macol. 2020, 177, 4899–4920. [Google Scholar] [CrossRef] [PubMed]
- Chiou, W.-C.; Hsu, M.-S.; Chen, Y.-T.; Yang, J.-M.; Tsay, Y.-G.; Huang, H.-C. Repurposing existing drugs: Identification of SARS-CoV-2 3C-like protease inhibitors. J. Enzyme Inhib. Med. Chem. 2021, 36, 147–153. [Google Scholar] [CrossRef] [PubMed]
- Zheng, W.; Fan, W.; Zhang, S.; Jiao, P.; Shang, Y.; Cui, L.; Mahesutihan, M.; Li, J.; Wang, D.; Gao, G.F.; et al. Naproxen Exhibits Broad Anti-influenza Virus Activity in Mice by Impeding Viral Nucleoprotein Nuclear Export. Cell Rep. 2019, 27, 1875–1885.e5. [Google Scholar] [CrossRef]
- Pan, T.; Peng, Z.; Tan, L.; Zou, F.; Zhou, N.; Liu, B.; Liang, L.; Chen, C.; Liu, J.; Wu, L.; et al. Nonsteroidal Anti-inflammatory Drugs Potently Inhibit the Replication of Zika Viruses by Inducing the Degradation of AXL. J. Virol. 2018, 92. [Google Scholar] [CrossRef]
- Hung, I.F.N.; To, K.K.W.; Chan, J.F.W.; Cheng, V.C.C.; Liu, K.S.H.; Tam, A. Efficacy of Clarithromycin-Naproxen-Oseltamivir Combination in the Treatment of Patients Hospitalized for Influenza A(H3N2) Infection: An Open-label Randomized, Controlled, Phase IIb/III Trial. Chest 2017, 151, 1069–1080. [Google Scholar] [CrossRef] [PubMed]
- Efficacy of Addition of Naproxen in the Treatment of critically ill Patients Hospitalized for COVID-19 Infection/Enacovid Study. Available online: https://www.clinicaltrialsregister.eu/ctr-search/search?query=eudract_number:2020-001301-23 (accessed on 13 April 2021).
- Ye, Q.; Krug, R.M.; Tao, Y.J. The mechanism by which influenza A virus nucleoprotein forms oligomers and binds RNA. Nat. Cell Biol. 2006, 444, 1078–1082. [Google Scholar] [CrossRef] [PubMed]
- Trott, O.; Olson, A.J. AutoDock Vina: Improving the speed and accuracy of docking with a new scoring function, efficient optimization, and multithreading. J. Comput. Chem. 2010, 31, 455–461. [Google Scholar] [CrossRef] [PubMed]
- Brooks, B.R.; Bruccoleri, R.E.; Olafson, B.D.; States, D.J.; Swaminathan, S.; Karplus, M. CHARMM: A program for macromolecular energy, minimization, and dynamics calculations. J. Comput. Chem. 1983, 4, 187–217. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Compound | N Dimer.PDB 6VYO ΔG (Kcal/mol)/RMSD (Å) | Binding Site | Score Dimer/% Population | N NTD Monomer PDB 7ACT Binding Site | Score Monomer/ % Population |
|---|---|---|---|---|---|
| Naproxen | −37 ± 1 /2.9 ±1.2 | A: W52, I157 D: R92, L104, S105, R107, Y112 | −7.9/40% | W52, I146, R149 | −6.4/33% |
| –32 ± 2 /8 ±4 | A: T148, N150 D: R92, I94 | −6.4/25% | A50, A90, R92, R93 | −5.4/13% | |
| Naproxen A1 | A: N77, W52, I146 D: T54, R92, R107, Y109 | −7.1/35% | A90, R92, R107, Y109 | −6.1/15% | |
| Naproxen C01 | A: W52, N75, I157 D: R107 | −7.9/25% | A90, R92, R107, Y109 | −6.3/50% | |
| Acetaminophen (paracetamol) | A: H145, I146 | −5.7/50% | W52, I146, R149 | −5.0/25% | |
| Celecoxib1 | A: W52, N75, I157 D: A55,R107 | −8.1/40% | W52, I146, R149 | −7.4 /50% |
| Compound | COX Inhibition | N Binding µM Fluo /Fluo Anisotropy | Tm (°C)/ΔTm (°C) 4 | Oligomer Inhibition/Enhancement | Oligomer Size (nm).5 | Mean IC50 µM | CC50 µM |
|---|---|---|---|---|---|---|---|
| None | 51 ± 1 | 15.7 ± 0.7 | |||||
| Naproxen | COX1 + COX2 | 1.1 ± 0.1 2/ 4.4 ± 1.4 3 | 56 ± 1 /+5 | +++ | 7.9 ± 0.3 | 52 ± 6 | >1000 |
| Naproxen A | None (ref 11) | Nd | 47 ± 2, 58 sh /−4 | - | 16.0 ± 0.8 | No inhibition | >1000 |
| Naproxen C0 | None (ref 11) | / 1.6 ± 0.5 3 | - | + | 12.7 ± 0.7 | 73 ± 25 | >1000 |
| Acetaminophen (paracetamol) | COX1 + COX2 ? 1 | Nd | 46 ± 2, 54sh /−5 | - | 14.0 ± 1.0 | No inhibition | >400 |
| Celecoxib | COX2 | Nd | −6 | /Enhancement | 7 Formation of large particles of 3 µm as compared to N + DMSO (12 nm) | proviral | 80 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Terrier, O.; Dilly, S.; Pizzorno, A.; Chalupska, D.; Humpolickova, J.; Bouřa, E.; Berenbaum, F.; Quideau, S.; Lina, B.; Fève, B.; et al. Antiviral Properties of the NSAID Drug Naproxen Targeting the Nucleoprotein of SARS-CoV-2 Coronavirus. Molecules 2021, 26, 2593. https://doi.org/10.3390/molecules26092593
Terrier O, Dilly S, Pizzorno A, Chalupska D, Humpolickova J, Bouřa E, Berenbaum F, Quideau S, Lina B, Fève B, et al. Antiviral Properties of the NSAID Drug Naproxen Targeting the Nucleoprotein of SARS-CoV-2 Coronavirus. Molecules. 2021; 26(9):2593. https://doi.org/10.3390/molecules26092593
Chicago/Turabian StyleTerrier, Olivier, Sébastien Dilly, Andrés Pizzorno, Dominika Chalupska, Jana Humpolickova, Evžen Bouřa, Francis Berenbaum, Stéphane Quideau, Bruno Lina, Bruno Fève, and et al. 2021. "Antiviral Properties of the NSAID Drug Naproxen Targeting the Nucleoprotein of SARS-CoV-2 Coronavirus" Molecules 26, no. 9: 2593. https://doi.org/10.3390/molecules26092593
APA StyleTerrier, O., Dilly, S., Pizzorno, A., Chalupska, D., Humpolickova, J., Bouřa, E., Berenbaum, F., Quideau, S., Lina, B., Fève, B., Adnet, F., Sabbah, M., Rosa-Calatrava, M., Maréchal, V., Henri, J., & Slama-Schwok, A. (2021). Antiviral Properties of the NSAID Drug Naproxen Targeting the Nucleoprotein of SARS-CoV-2 Coronavirus. Molecules, 26(9), 2593. https://doi.org/10.3390/molecules26092593