3.2. Extraction, Fractionation, and Bioassays
The shade and air-dried plant Onosma chitralicum (2 kg) were taken and ground to get powder, soaked in methanol (4 L) for 14 days, and were extracted three times during the 14 days at room temperature in the same solvent, and then filtered. The filtrates were evaporated under reduced pressure by vacuum rotary evaporator at 35 °C. The extract was dried and weighed. The crude extract weighed to 80 g, was further suspended in water, and partitioned successively with n-hexane, chloroform, ethyl acetate, and n-butanol to obtain their soluble fractions. The weight of each solvent soluble fraction was n-hexane (15 g), chloroform (25 g), ethyl acetate soluble (18 g), n-butanol (12 g), and aqueous fraction (10 g).
Antibacterial and antifungal activities of the crude extract, other solvent soluble fractions, and the compounds
1–
7 isolated from
Onosma chitralicum were investigated. Bacterial strains, such as
Escherichia coli, Pseudomonas aeruginosa,
Staphylococcus aureus, Salmonella typhi, and
Bacillus subtilis were used in antibacterial essay while fungal strains
Aspergillus flavus, Fusarium solani, Aspergillus fumigates, Aspergillus niger, and
Candida glabrata were used in antifungal assay. The bacterial and fungal strains used in this study were clinical isolates that were isolated and identified previously at Department of Microbiology of this university. Disc diffusion method was used for antifungal activity, while for antibacterial activities, the agar well diffusion method was used. In the disc diffusion technique, the bacterial culture was streaked on the surface of the solidified agar media in the sterile petri plate. Next, on the sterile disc, 10 μL of the extract or fraction and isolated compounds from their stock solutions were allowed to absorb in independent experiments, and zones of inhibition were measured after 24 h of incubation [
31].
In the agar-well diffusion method for bacterial strains, wells of 6 mm were dug in the sterile agar media by using a sterile plastic borer. Next, each well was given a specific number, and bacterial culture was inoculated on the surface of the solidified media. Stock solutions of crude extracts and fractions in one experiment and the isolated compounds in another experiment were added into respective wells. The zones of inhibition were measured after 24 h of incubation at 37 °C in an incubator [
32]. Doxycycline and Miconazole were used in the crude assays, whereas, levofloxacin and clotrimazole were used in the isolated compounds screening as standard positive controls, while DMSO was used as a negative control. The zones of inhibition of crude extract, fractions, and the isolated compounds were compared with the zones of inhibition of standard drugs.
3.3. Isolation of Chemical Constituents
The most potent activity was observed in ethyl acetate fraction and was further subjected to separation protocol using column chromatography. The elution through column chromatography was carried out based on the gradient of polarity (
n-C
6H
14 →
n-C
6H
14-EtOAc → pure-EtOAc). As a result, four fractions A, fractions B, fractions C and fractions D were obtained by eluting
n-C
6H
14-EtOAc of different fractions through column chromatography. Fraction A was obtained by using
n-C
6H
14-EtOAc (8:2), consisted of two pure compounds, compound
1 (4′,8-dimethoxy-7-hydroxyisoflavone) [
33] and compound
3 (2,3-dihydro-3-hydroxy-7-methoxyflavone) [
34] with a ratio of (8.50:1.50 & 7.50:2.50), respectively. Fraction B was also obtained as discussed above with
n-C
6H
14-EtOAc (6.0:4.0), consisted of compound
2 (2,3-dihydro-3,5-dihydroxy-7-methoxyflavone) [
35] and compound
4 (7,4′-dihydroxy-3′-methoxyisoflavone) [
36], with the ratio of (6.50:3.50 and 6.50:4.0), respectively. Fraction C was eluted by using
n-C
6H
14-EtOAc at the ratio of (9:1) was further subjected to column chromatography. Fraction C afforded only one pure compound, compound
5 (7,4′-dimethoxy-isoflavone) [
37]. Fraction D was eluted with
n- C
6H
14-EtOAc by using the ratio of (5:5) and 2 compounds, compound
6 (5,3′,3-trihydroxy-7,4′-dimethoxyflavanone) [
38] and compound
7 (5′,7,8-trihydroxy-6,3′,4′-trimethoxyflavanone) [
39] with the ratio of (2.50:7.50 & 4.50:5.50) were eluted, respectively.
For isolation of compounds, Eyela (EF-10 model) flash chromatography a type of flash column chromatography technique was used by using silica gel (E. Merck Si 60,230–400 mesh).
3.5. Physical and Spectral Data of Isolated Compounds 1 to 7
4′,8-dimethoxy-7-hydroxyisoflavone (
1): colorless solid. UV λ
max (CH
3Cl) 254,350 nm. IR(KBr) cm
−1: 1600, 1685, 3450, 1315,
1H-NMR (CDCl
3, 400 MHz) δ: 8.33 (2H s,
J = 0 Hz), 7.62 (5H d,
J = 8.3), 6.89 (6H d,
J = 9.4), 7.47 (2H′ d,
J = 8.81), 6.94 (3H′ d,
J = 8.81), 6.94 (5H′ d,
J = 8.81), 7.47 (6H′ d,
J = 8.81), 3.81 (8OCH
3 s, ), 3.79 (4-OCH
3′s,);
13C-NMR (CDCl
3) δ: 154.2 (C-2), 123.2 (C-3), 181.1(C-4), 123.9 (C-5), 122.5 (C-6), 163.1 (C-7), 145.5 (C-8), 159.6 (C-9), 106.1 (C-10), 123.6 (C-1′), 121 (C-2′), 122 (C-3′), 146.5 (C-4′), 122 (C-5′), 121 (C-6′), 58.4 (8-OCH
3), 61.3 (4-OCH
3′); EI-MS
m/
z 298. The above physical and spectral data are in complete agreement with the data reported in literature for compound
1 [33].
2,3-dihydro-3,5-dihydroxy-7-methoxyflavone (
2): colorless solid. UV λ
max (CH
3Cl) 250,323 nm. IR(KBr) cm
−1: 1605, 1710, 3250, 1310;
1H-NMR (CDCl
3,400 MHz) δ: 5.14 (2H d,
J = 11.0 Hz), 4.54 (3H d,
J = 11.0 Hz), 11.04 (5H s), 6.05 (6H d,
J = 2.3), 6.18 (8H d,
J = 2.3 Hz), 7.40 (2H′ m).5, 7.42 (3H′ m), 7.43 (4H′ m) 7.47 (5H′ m), 7.51 (7OCH
3 s);
13C-NMR (CDCl
3) δ: 77.2 (C-2), 149.7 (C-3), 181.9(C-4), 158.3 (C-5), 121.5 (C-6), 146.3 (C-7), 122.5 (C-8), 156.3 (C-9), 106.9 (C-10), 122.8 (C-1′), 121.9 (C-2′), 123.2 (C-3′), 123.7 (C-4′), 122.2 (C-5′), 121. (C-6′), 58.3 (8OCH
3); (calcd for C
16H
12O
5284) EI-MS
m/
z 284. The above physical and spectral data coincided with the data reported in literature for compound
2 [34].
2,3-dihydro-3-hydroxy-7-methoxyflavone (
3): colorless solid. UV λ
max (CH
3Cl) 254,350 nm. IR(KBr) cm
−1: 1600,1685,3450,1315;
1H-NMR (CDCl
3,400 MHz) δ: 8.33 (2H s,
J= 0 Hz), 7.62 (5H d,
J = 8.3), 6.89 (6H d,
J = 9.4), 7.47 (2H′ d,
J = 8.81), 6.94 (3H′ d,
J = 8.81), 6.94 (5H′ d,
J = 8.81), 7.47 (6H′ d,
J = 8.81), 3.81 (8OCH
3 s), 3.79 (4OCH
3′ s,);
13C-NMR (CDCl
3) δ: 154.2 (C-2), 123.2 (C-3), 181.1(C-4), 123.9 (C-5), 122.5 (C-6), 163.1 (C-7), 145.5 (C-8), 159.6 (C-9), 106.1 (C-10), 123.6 (C-1′), 121 (C-2′), 122 (C-3′), 146.5 (C-4′), 122 (C-5′), 121 (C-6′), 58.4 (8OCH
3), 61.3 (4OCH
3′); EI-MS
m/
z 268. The above physical and spectral data are in complete agreement with the data reported in literature for compound
3 [35].
7,4′-dihydroxy-3′-methoxyisoflavone (
4): colorless solid. UV λ
max (CH
3Cl) 255,336 nm. IR(KBr) cm
−1: 1610, 1680, 3450, 1325;
1H-NMR (CDCl
3,400 MHz) δ: 5.14 (2H d,
J = 11.2 Hz), 7.80 (5H d,
J = 8.9), 6.90 (6H d,
J = 8.9), 6.49 (8H d,
J = 2.7 Hz), 7.19 (2H′ d,
J =1.5), 7.42 (3H′ m), 6.76 (5H′ d,
J = 8.0) 6.79 (6H′ d,
J = 8.0), 3.76 (3-OCH
3);
13C-NMR (CDCl
3) δ: 151.4 (C-2), 153.5 (C-3), 173.5(C-4), 126.5 (C-5), 121.8 (C-6), 165.9 (C-7), 102 (C-8), 156.9 (C-9), 115.8 (C-10), 123.8 (C-1′), 114.3 (C-2′), 148.1 (C-3′), 145.4 (C-4′), 114.1 (C-5′), 118.5 (C-6′); EI-MS
m/
z 284. The above physical and spectral data are similar with the data reported in literature for compound
4 [36].
7,4′-dimethoxy-isoflavone (
5): colorless solid. UV λ
max (CH
3Cl) 250,323 nm. IR(KBr) cm
−1: 1605, 1670, 3350, 1340;
1H-NMR (CDCl
3,400 MHz) δ: 7.96 (2H, s), 8.31 (5H d,
J = 9.3 Hz), 6.97 (6H dd,
J = 9.3,2.5), 6.87 (8H d,
J = 2.5), 7.40 (2H′ d,
J = 8.1), 7.42 (5H′ d,
J = 8.4), 7.40 (6H′ d,
J = 8.1), 3.85 (7OCH
3 s), 3.89 (4OCH
3′ s),
13C-NMR (CDCl
3) δ: 150.2 (C-2), 123.1 (C-3), 181.3 (C-4), 123.8 (C-5), 122.8 (C-6), 131.8 (C-7), 122.5 (C-8), 159.6 (C-9), 106.1 (C-10), 123.6 (C-1′), 121 (C-2′), 122 (C-3′), 131.1 (C-4′), 122 (C-5′), 121 (C-6′), 61.2 (4OCH
3′) 61.3 (8-OCH
3); EI-MS
m/
z 282. The above physical and spectral data are in complete agreement with the data reported in literature for compound
5 [
37].
5,3′,3-trihydroxy-7,4′-dimethoxyflavanone (
6): colorless solid. UV λ
max (CH
3Cl) 260,388 nm. IR(KBr) cm
−1: 1605, 1680, 3410, 1340;
1H-NMR (CDCl
3,400 MHz) δ: 4.59 (2H d,
J = 11.5 Hz), 5.04 (3H d,
J = 11.5 Hz), 11.24 (5H s) 6.19 (6H d,
J = 2.8), 6.18 (8H d,
J = 2.8), 6.96 (2H′ d,
J = 2.8), 6.91 (5H′ d,
J = 7.8), 7.09 (6H′ d,d
J = 7.8,2.8), 3.81 (7OCH
3 s,), 3.89 (4OCH
3′ s);
13C-NMR (CDCl
3) δ: 85.6 (C-2), 77.4 (C-3), 194.9(C-4), 166.6 (C-5), 99.1(C-6), 167.8 (C-7), 98.7 (C-8), 168.9 (C-9), 100.2 (C-10), 128.8 (C-1′), 109.1 (C-2′), 144.5 (C-3′), 145.6 (C-4′), 115.6 (C-5′), 122.1 (C-6′), 56.8 (4OCH
3 ’) 56.9 (8-OCH
3); EI-MS
m/
z 360. The above physical and spectral data coincided with the data reported in literature for compound
6 [38].
5′,7,8-trihydroxy-6,3′,4′-trimethoxyflavanone (
7): colorless solid. UV λ
max (CH
3Cl) 250,330 nm. IR(KBr) cm
−1: 1603, 1695, 3310, 1350;
1H-NMR (CDCl
3,400 MHz) δ: 8.35(2H s, 6.71 (5H s), 3.89 (3OCH
3′ s), 3.78 (4OCH
3′ s), 7.47 (6H′ d,
J = 2.9), 3.47 (6OCH
3 s,);
13C-NMR (CDCl
3) δ: 156.3 (C-2), 123.8 (C-3), 185.6(C-4), 118.1 (C-5), 154.9 (C-6), 151.1 (C-7), 136.6 (C-8), 159.8 (C-9), 107.1 (C-10), 124.8 (C-1′), 124.4 (C-2′), 149.1 (C-3′), 149.9 (C-4′), 136.8 (C-5′), 136.4 (C-6′), 60.9(3OCH
3′), 59.1 (4-OCH
3′) 58.1 (6-OCH
3); EI-MS
m/
z 332. The above physical and spectral data are similar with the data reported in literature for compound
7 [39].
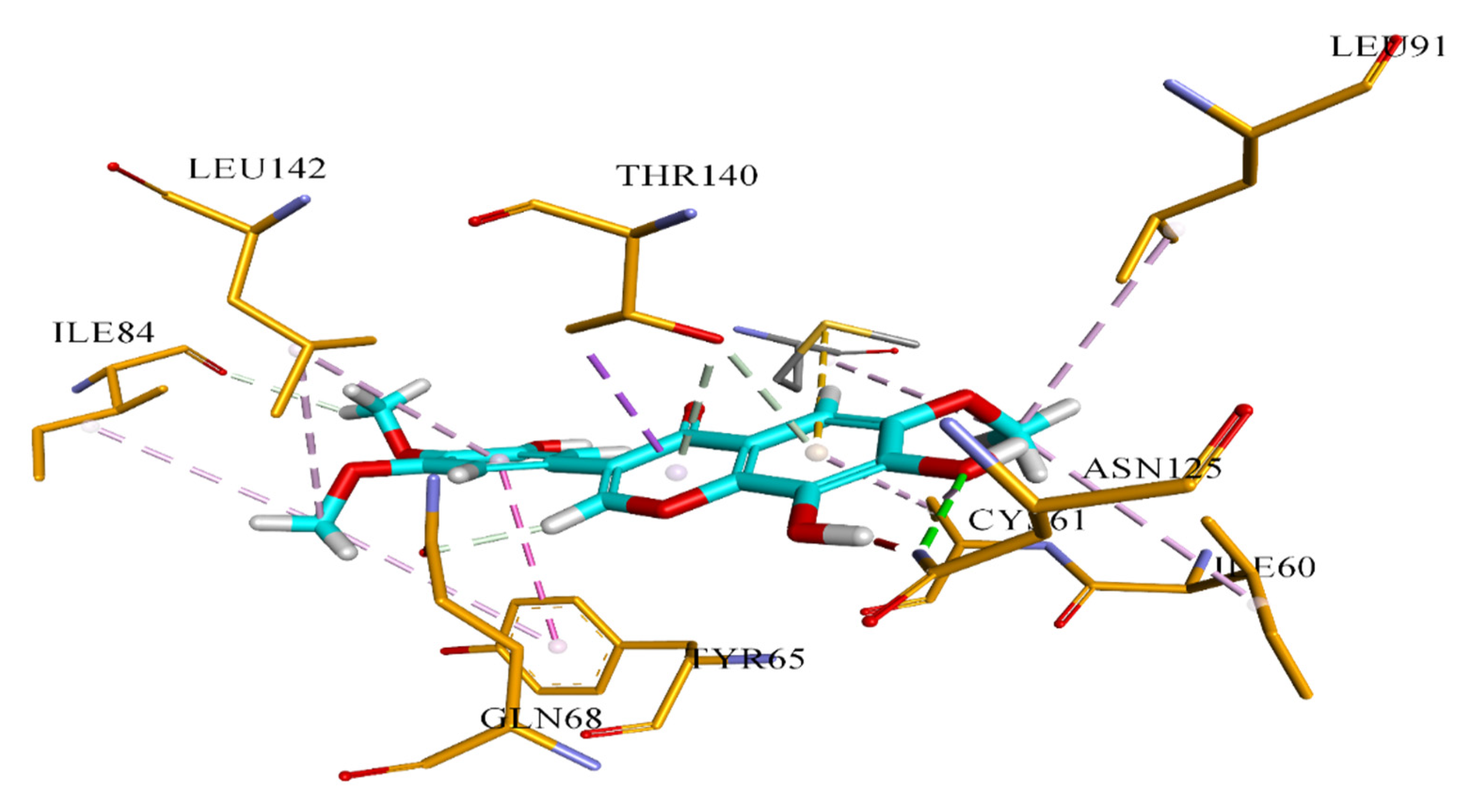
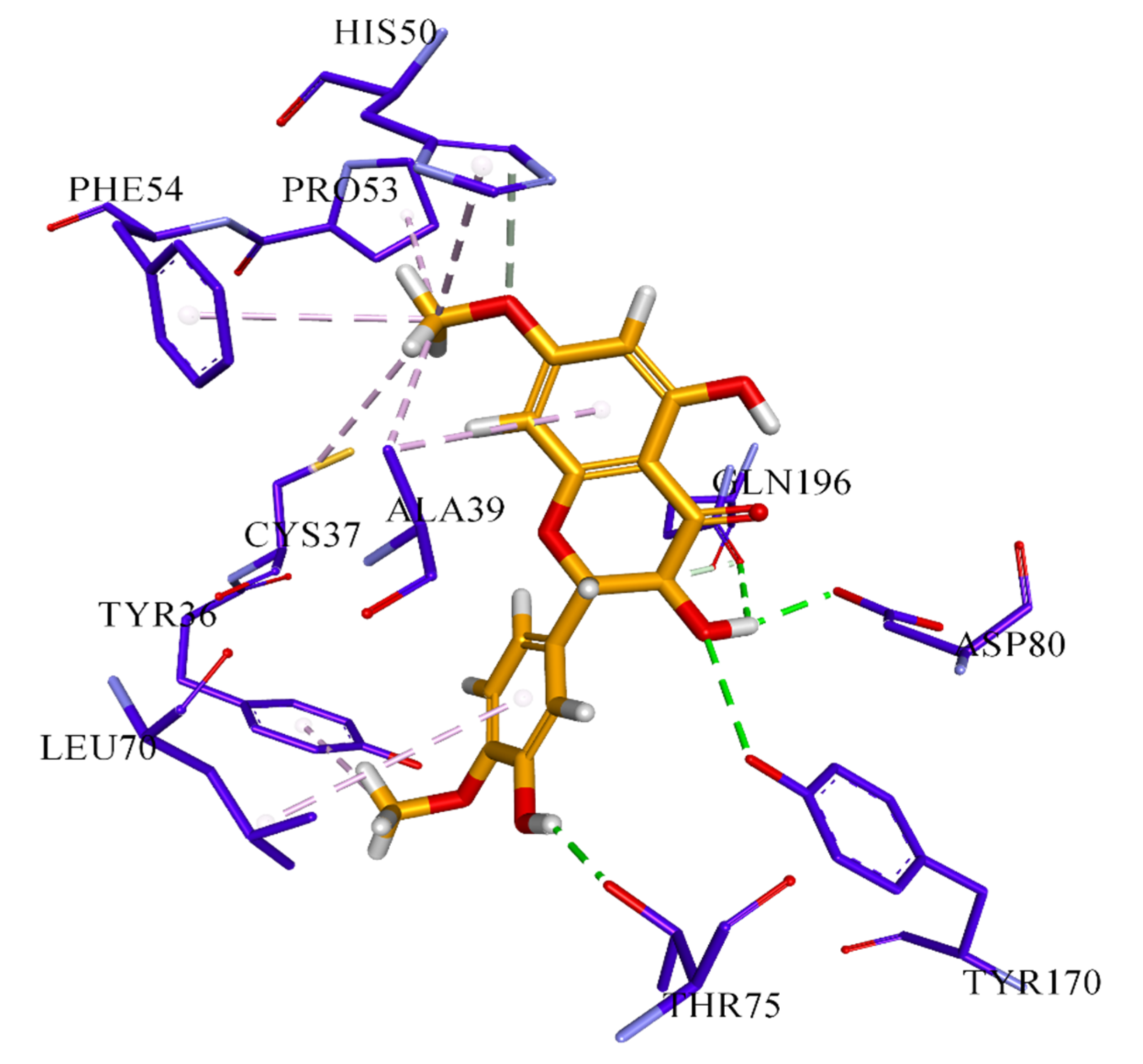
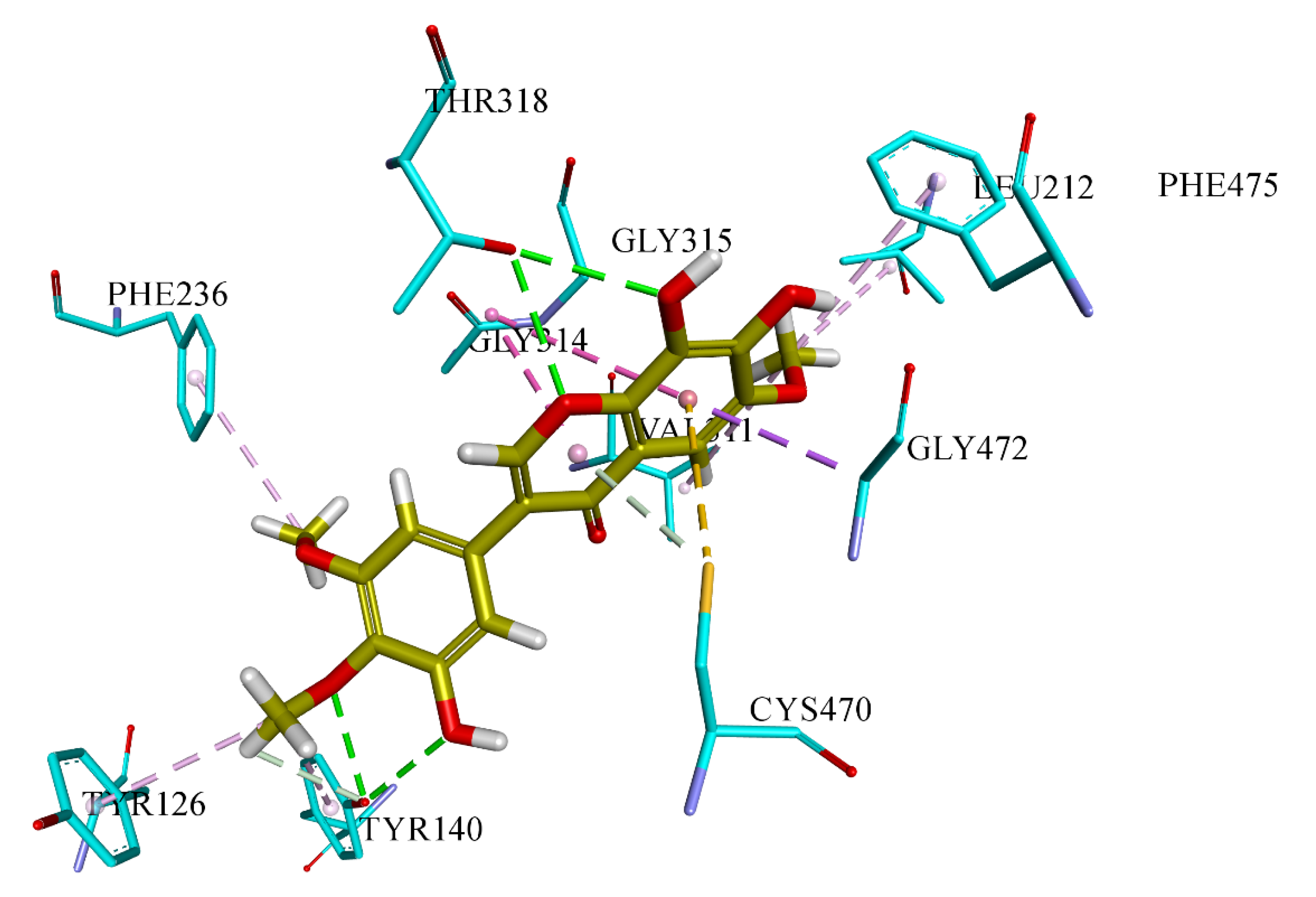
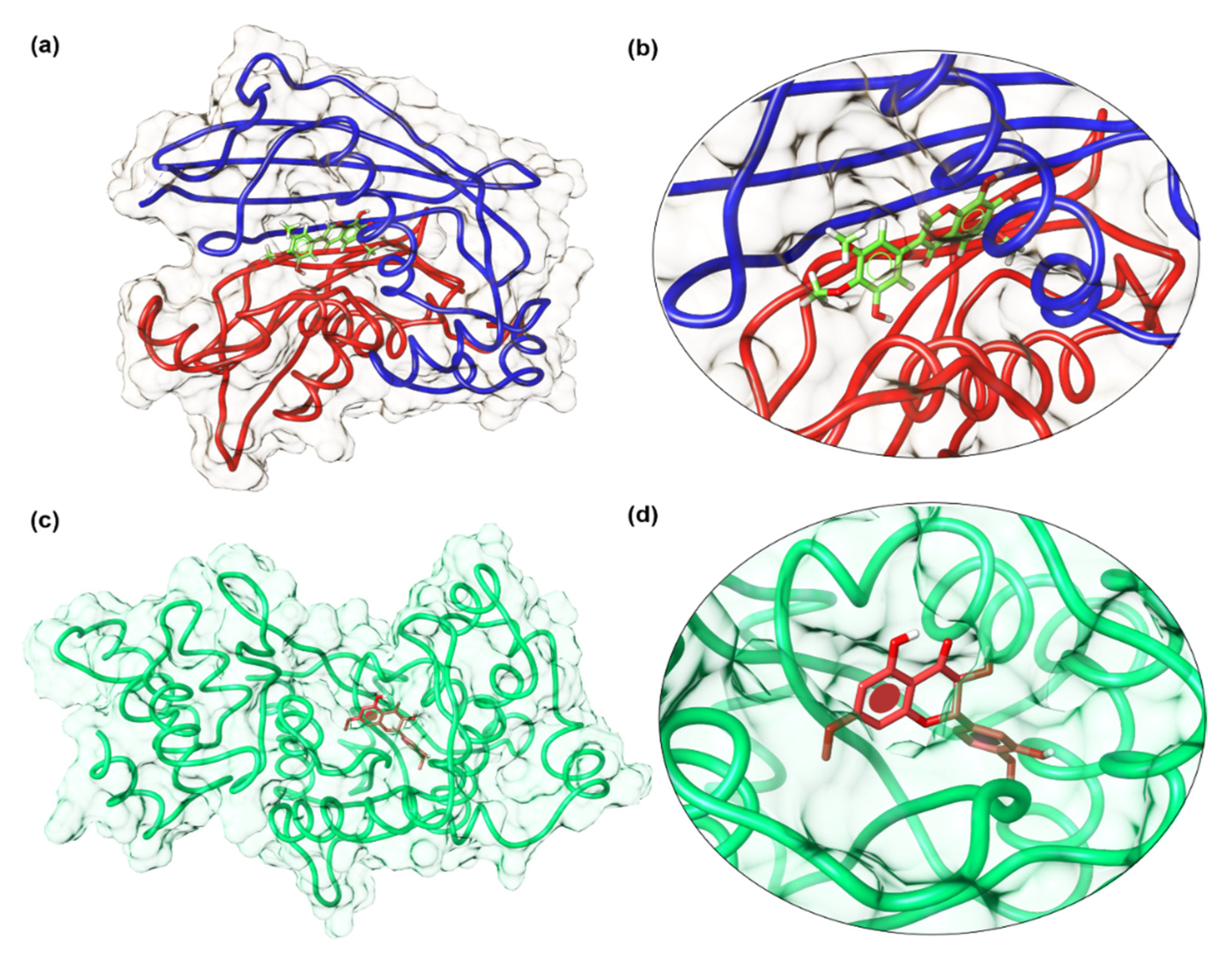
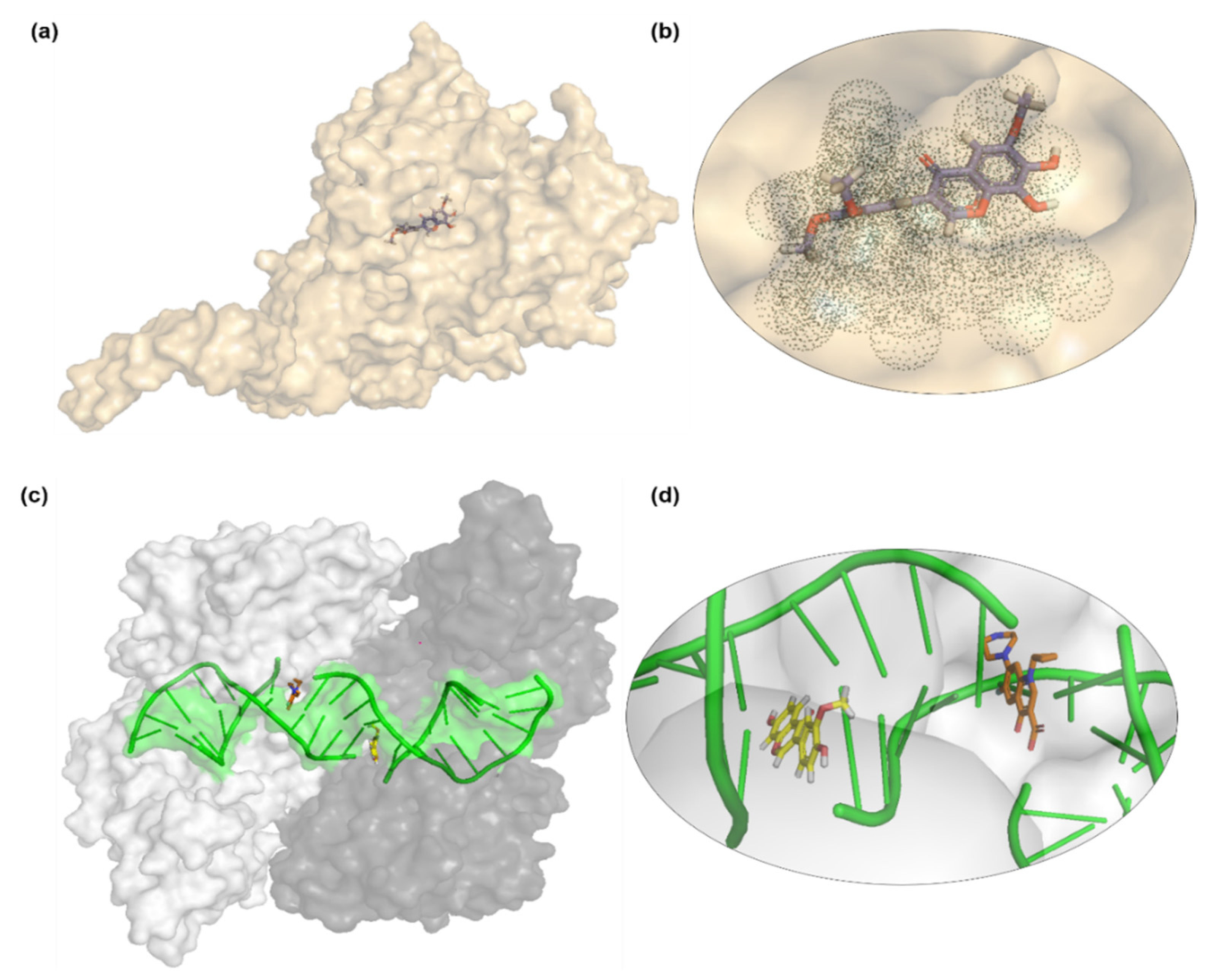
3.6. Docking Analysis Protocol
The molecular docking studies of all seven compounds (compounds
1–
7) were carried out to investigate the putative binding interaction within the target proteins. The starting three-dimensional (3D) structure of
S. aureus tyrosyl-tRNA synthetase (PDB id: 1JIJ) and topoisomerase II DNA gyrase (PDB id: 2XCT) and beta-hydroxyacyl-ACP dehydratase HadAB (PDB id: 4RLT) were obtained from the Protein Data Bank (PDB) [
40]. Ligand molecules were sketched using the Chem Draw Professional v17 (PerkinElmer, Akron, OH, USA). The molecules were converted into 3D using Chem3D (PerkinElmer, Akron, OH, USA) [
41]. Before performing the docking protocol, chemically correct models of the ligands were generated using the OMEGA module of OpenEye Scientific Software, and the receptor structures were prepared using the MAKERECEPTOR Wizard Module of Openeye Scientific Software (Santa Fe, NM, USA) [
42]. Molecular docking was carried out using the FRED ligand-docking module. FRED requires a set of input conformers for each compound. The conformers of each ligand were generated by OMEGA 3.0.0 (Santa Fe, NM, USA) [
43]. Default settings of OMEGA were used for the generation of multi-conformers. Receptor grids were generated using the PDB2Receptor grid generation module. Grids were generated for the prepared proteins. For the
S. aureus gyrase complex, the grid was generated around ciprofloxacin, while for the
S. aureus tyrosyl-tRNA synthetase complex, the grid was generated around co-crystal SB-239629 ligand. Moreover, in the case of beta-hydroxyacyl-ACP dehydratase HadAB grid box was selected around Fisetin. In the case of
S. cerevisiae co-crystallized, the grid was generated around co-crystal fluconazole ligand. The boundary box was set at default value, which was spacious enough to encompass the binding regions in target proteins. The docking protocol was optimized using the re-docking of the co-crystal ligand within the active site of the target protein. FRED generated ten poses for each ligand and the pose with the lowest Chemguass4 was selected for further analysis. Binding interactions of best-docked poses were visualized using Discovery Studio client v16.1.0 [
44].


 ,
, 


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}





