
Proteomics Profiling of the Urine of Patients with Hyperthyroidism after Anti-Thyroid Treatment


Abstract
1. Introduction
2. Results
2.1. Anthropometric and Biochemical Data
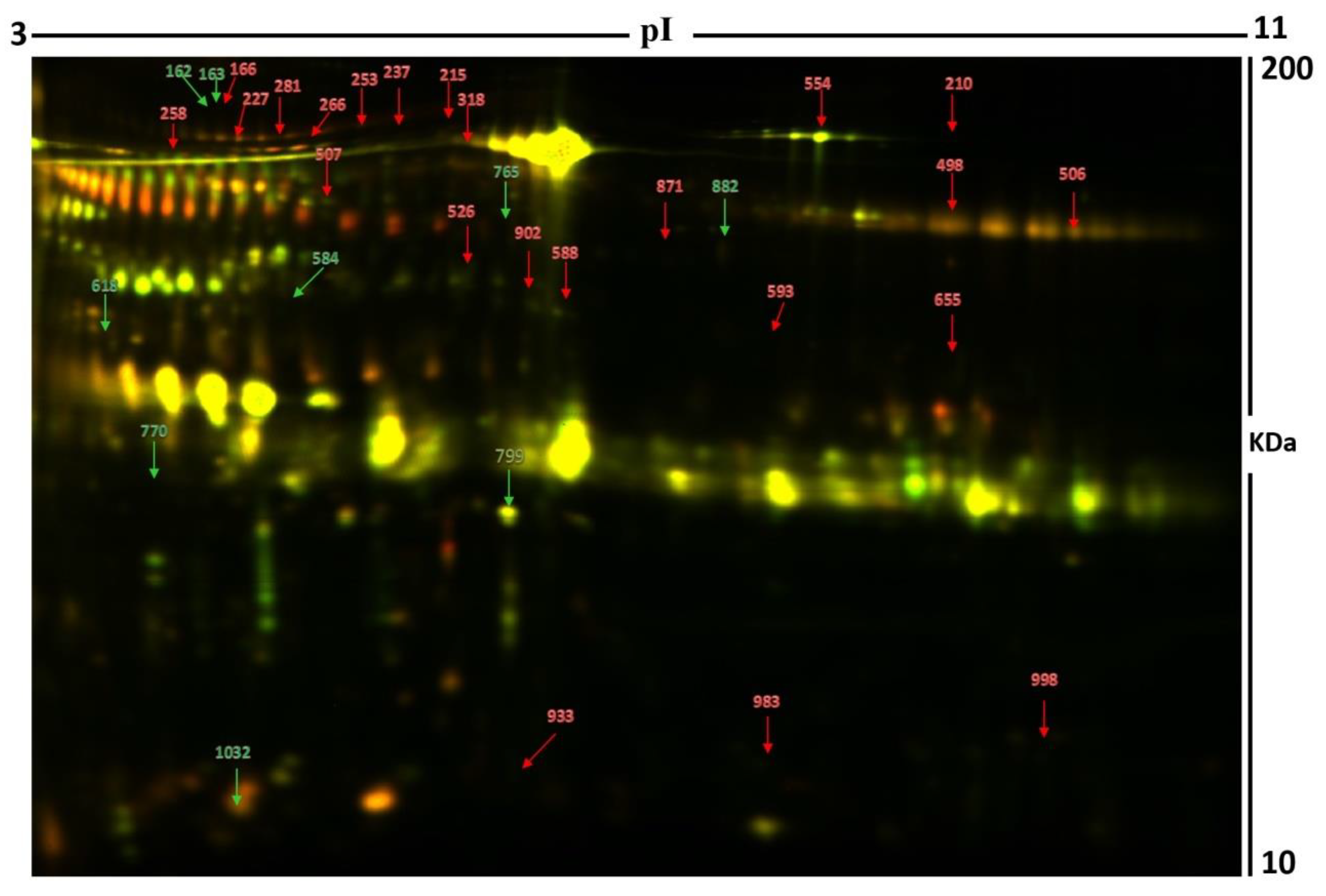
2.2. 2D-DIGE Analysis and Identification of Differentially Expressed Proteins
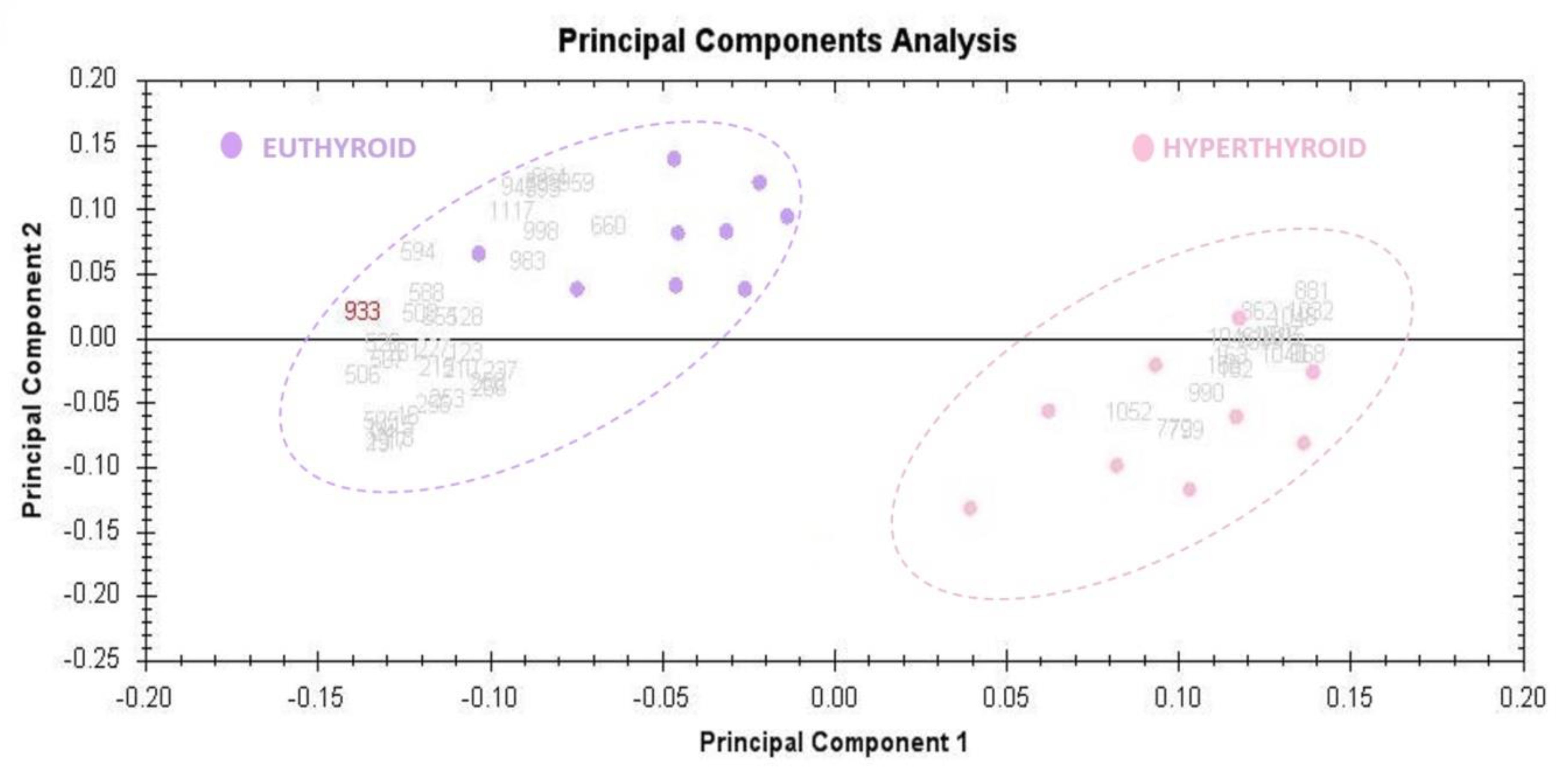
2.3. Principal Component Analysis
2.4. Network Pathway Analysis
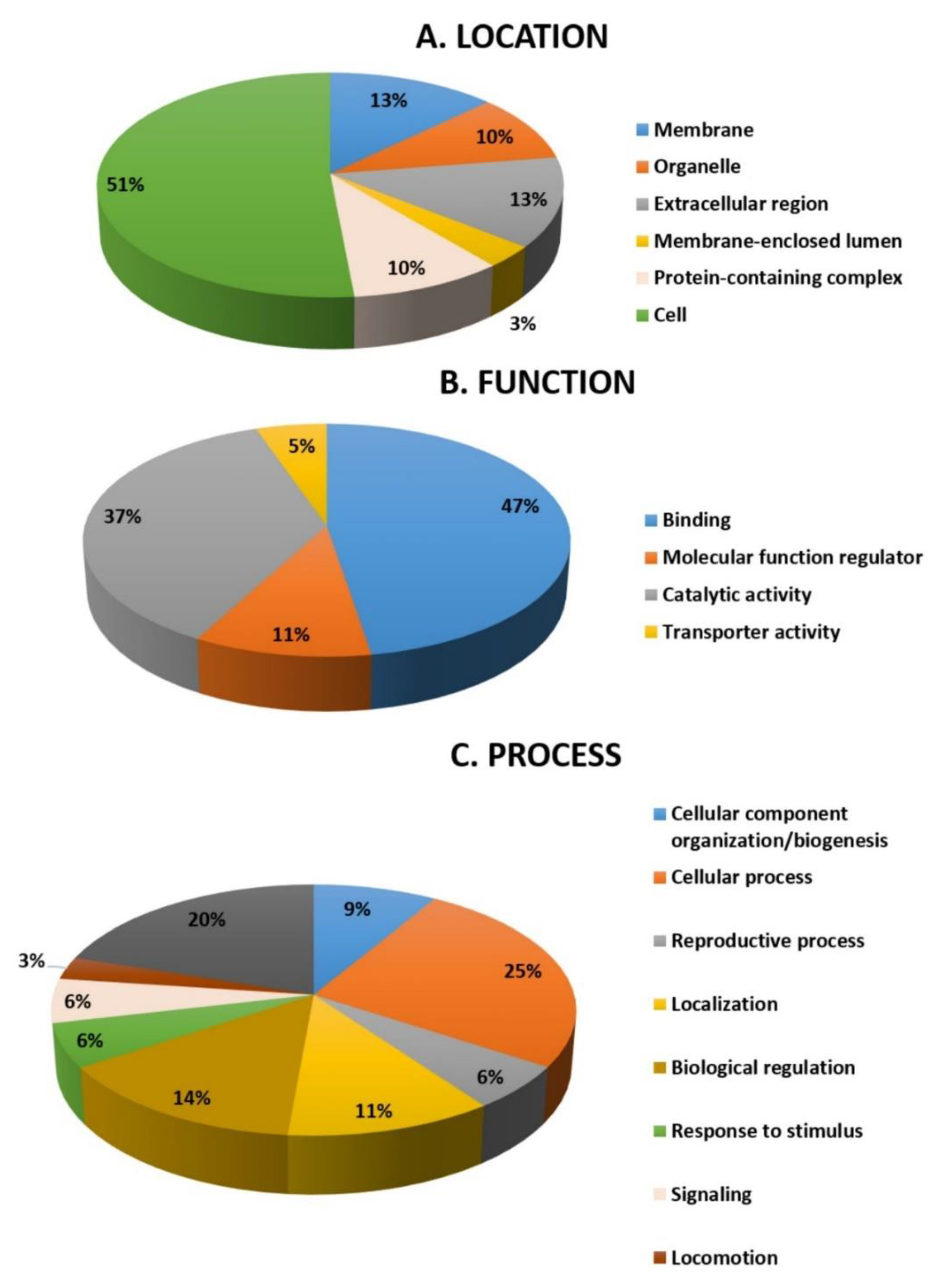
2.5. Function Analysis
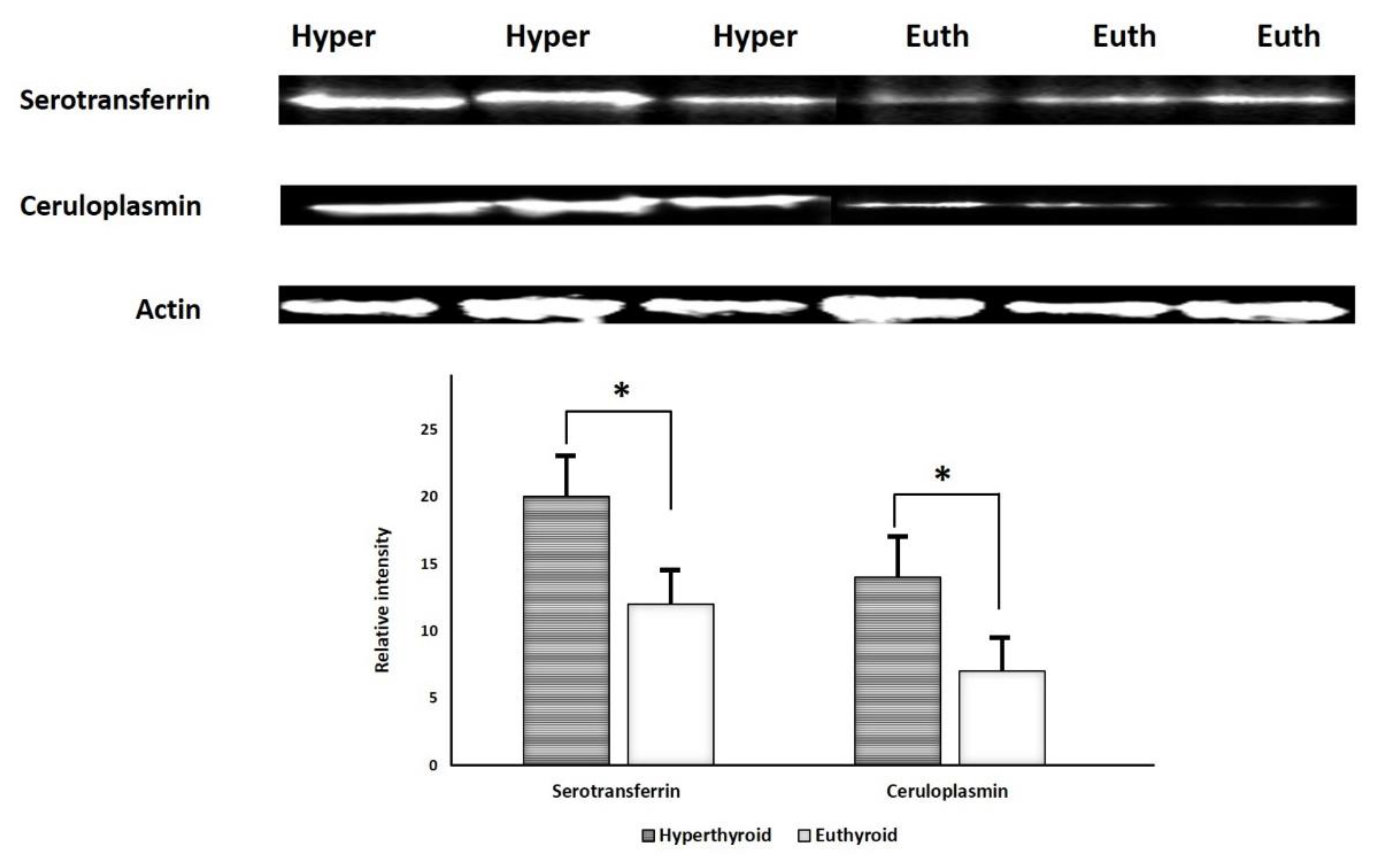
2.6. Immunoblotting Confirmation of Changes in Selected Proteins
3. Discussion and Conclusions
3.1. Proteins Increased in the Urine of Patients with Hyperthyroidism
3.2. Proteins Decreased in the Patients with Hyperthyroidism
4. Materials and Methods
4.1. Ethical Considerations and Informed Consent
4.2. Study Design
4.3. Protein Extraction from Urine Samples
4.4. Fluorescence Labeling, 2D-DIGE, and MALDI-TOF-MS Analysis
4.5. Bioinformatic Analysis: Functional Classification of Proteins and Pathway Analysis
4.6. Statistical Analysis
4.7. Immunoblotting
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Iglesias, P.; Díez, J. Thyroid dysfunction and kidney disease. Eur. J. Endocrinol. 2009, 160, 503–515. [Google Scholar] [CrossRef]
- Thongboonkerd, V.; Malasit, P. Renal and urinary proteomics: Current applications and challenges. Proteomics 2005, 5, 1033–1042. [Google Scholar] [CrossRef]
- Katz, A.I.; Emmanouel, D.S.; Lindheimer, M.D. Thyroid hormone and the kidney. Nephron 1975, 15, 223–249. [Google Scholar] [CrossRef]
- Kobori, H.; Ichihara, A.; Miyashita, Y.; Hayashi, M.; Saruta, T. Mechanism of hyperthyroidism-induced renal hypertrophy in rats. J. Endocrinol. 1998, 159, 9. [Google Scholar] [CrossRef][Green Version]
- Marre, M. Recent advance in the treatment of diabetic nephropathy: Angiotensin I converting enzyme (ACE) inhibitors. J. Diabet. Complicat. 1991, 5, 91. [Google Scholar] [CrossRef]
- Masood, A.; Benabdelkamel, H.; Ekhzaimy, A.A.; Alfadda, A.A. Plasma-Based Proteomics Profiling of Patients with Hyperthyroidism after Antithyroid Treatment. Molecules 2020, 25, 2831. [Google Scholar] [CrossRef]
- Alfadda, A.A.; Benabdelkamel, H.; Masood, A.; Jammah, A.A.; Ekhzaimy, A.A. Differences in the plasma proteome of patients with hypothyroidism before and after thyroid hormone replacement: A proteomic analysis. Int. J. Mol. Sci. 2018, 19, 88. [Google Scholar] [CrossRef] [PubMed]
- Masood, A.; Benabdelkamel, H.; Jammah, A.A.; Ekhzaimy, A.A.; Alfadda, A.A. Identification of Protein Changes in the Urine of Hypothyroid Patients Treated with Thyroxine Using Proteomics Approach. ACS Omega 2021, 6, 2367–2378. [Google Scholar] [CrossRef]
- Kravets, I. Hyperthyroidism: Diagnosis and treatment. Am. Fam. Physician 2016, 93, 363–370. [Google Scholar] [PubMed]
- Carroll, M.F.; Temte, J.L. Proteinuria in adults: A diagnostic approach. Am. Fam. Physician 2000, 62, 1333–1340. [Google Scholar] [PubMed]
- Zhao, M.; Li, M.; Yang, Y.; Guo, Z.; Sun, Y.; Shao, C.; Li, M.; Sun, W.; Gao, Y. A comprehensive analysis and annotation of human normal urinary proteome. Sci. Rep. 2017, 7, 1–13. [Google Scholar] [CrossRef]
- Adachi, J.; Kumar, C.; Zhang, Y.; Olsen, J.V.; Mann, M. The human urinary proteome contains more than 1500 proteins, including a large proportion of membrane proteins. Genome Biol. 2006, 7, 1–16. [Google Scholar] [CrossRef]
- KELER-BACOKA, M.; Hahn, A. Serum Copper and Ceruloplasmin (PPD Oxidase) in Hyperthyroidism. Radovi Medicinskog faculteta u Zagrebu 1960, 8, 244–249. [Google Scholar]
- Stříž, I.; Trebichavský, I. Calprotectin—A pleiotropic molecule in acute and chronic inflammation. Physiol. Res. 2004, 53, 245–253. [Google Scholar]
- Turnier, J.L.; Fall, N.; Thornton, S.; Witte, D.; Bennett, M.R.; Appenzeller, S.; Klein-Gitelman, M.S.; Grom, A.A.; Brunner, H.I. Urine S100 proteins as potential biomarkers of lupus nephritis activity. Arthritis Res. Ther. 2017, 19, 1–11. [Google Scholar] [CrossRef]
- Ito, Y.; Yoshida, H.; Tomoda, C.; Uruno, T.; Miya, A.; Kobayashi, K.; Matsuzuka, F.; Kakudo, K.; Kuma, K.; Miyauchi, A. S100A4 expression is an early event of papillary carcinoma of the thyroid. Oncology 2004, 67, 397–402. [Google Scholar] [CrossRef] [PubMed]
- Nipp, M.; Elsner, M.; Balluff, B.; Meding, S.; Sarioglu, H.; Ueffing, M.; Rauser, S.; Unger, K.; Höfler, H.; Walch, A. S100-A10, thioredoxin, and S100-A6 as biomarkers of papillary thyroid carcinoma with lymph node metastasis identified by MALDI imaging. J. Mol. Med. 2012, 90, 163–174. [Google Scholar] [CrossRef] [PubMed]
- Nishimura, R.; Yokose, T.; Mukai, K. S-100 protein is a differentiation marker in thyroid carcinoma of follicular cell origin: An immunohistochemical study. Pathol. Int. 1997, 47, 673–679. [Google Scholar] [CrossRef] [PubMed]
- Stehle, J.R., Jr.; Weeks, M.E.; Lin, K.; Willingham, M.C.; Hicks, A.M.; Timms, J.F.; Cui, Z. Mass spectrometry identification of circulating alpha-1-B glycoprotein, increased in aged female C57BL/6 mice. Biochim. Biophys. Acta (BBA) Gen. Subj. 2007, 1770, 79–86. [Google Scholar] [CrossRef]
- Ishioka, N.; Takahashi, N.; Putnam, F.W. Amino acid sequence of human plasma alpha 1B-glycoprotein: Homology to the immunoglobulin supergene family. Proc. Natl. Acad. Sci. USA 1986, 83, 2363–2367. [Google Scholar] [CrossRef] [PubMed]
- Piyaphanee, N.; Ma, Q.; Kremen, O.; Czech, K.; Greis, K.; Mitsnefes, M.; Devarajan, P.; Bennett, M.R. Discovery and initial validation of α 1-B glycoprotein fragmentation as a differential urinary biomarker in pediatric steroid-resistant nephrotic syndrome. Proteom. Clin. Appl. 2011, 5, 334–342. [Google Scholar] [CrossRef] [PubMed]
- Kreunin, P.; Zhao, J.; Rosser, C.; Urquidi, V.; Lubman, D.M.; Goodison, S. Bladder cancer associated glycoprotein signatures revealed by urinary proteomic profiling. J. Proteome Res. 2007, 6, 2631–2639. [Google Scholar] [CrossRef] [PubMed]
- Yoon, S.Y.; Kim, J.-M.; Oh, J.-H.; Jeon, Y.-J.; Lee, D.-S.; Kim, J.H.; Choi, J.Y.; Ahn, B.M.; Kim, S.; Yoo, H.-S. Gene expression profiling of human HBV-and/or HCV-associated hepatocellular carcinoma cells using expressed sequence tags. Int. J. Oncol. 2006, 29, 315–327. [Google Scholar] [CrossRef] [PubMed]
- Tian, M.; Cui, Y.-Z.; Song, G.-H.; Zong, M.-J.; Zhou, X.-Y.; Chen, Y.; Han, J.-X. Proteomic analysis identifies MMP-9, DJ-1 and A1BG as overexpressed proteins in pancreatic juice from pancreatic ductal adenocarcinoma patients. BMC Cancer 2008, 8, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Lunardi, F.; Villano, G.; Perissinotto, E.; Agostini, C.; Rea, F.; Gnoato, M.; Bradaschia, A.; Valente, M.; Pontisso, P.; Calabrese, F. Overexpression of SERPIN B3 promotes epithelial proliferation and lung fibrosis in mice. Lab. Investig. 2011, 91, 945–954. [Google Scholar] [CrossRef]
- Kang, B.S.; Cooper, D.R.; Jelen, F.; Devedjiev, Y.; Derewenda, U.; Dauter, Z.; Otlewski, J.; Derewenda, Z.S. PDZ tandem of human syntenin: Crystal structure and functional properties. Structure 2003, 11, 459–468. [Google Scholar] [CrossRef]
- Sarkar, D.; Boukerche, H.; Su, Z.-z.; Fisher, P.B. mda-9/Syntenin: More than just a simple adapter protein when it comes to cancer metastasis. Cancer Res. 2008, 68, 3087–3093. [Google Scholar] [CrossRef]
- Koo, T.H.; Lee, J.-J.; Kim, E.-M.; Kim, K.-W.; Do Kim, H.; Lee, J.-H. Syntenin is overexpressed and promotes cell migration in metastatic human breast and gastric cancer cell lines. Oncogene 2002, 21, 4080–4088. [Google Scholar] [CrossRef] [PubMed]
- Latysheva, N.; Muratov, G.; Rajesh, S.; Padgett, M.; Hotchin, N.A.; Overduin, M.; Berditchevski, F. Syntenin-1 is a new component of tetraspanin-enriched microdomains: Mechanisms and consequences of the interaction of syntenin-1 with CD63. Mol. Cell. Biol. 2006, 26, 7707–7718. [Google Scholar] [CrossRef] [PubMed]
- Levy, S.; Shoham, T. The tetraspanin web modulates immune-signalling complexes. Nat. Rev. Immunol. 2005, 5, 136–148. [Google Scholar] [CrossRef] [PubMed]
- Hemler, M.E. Tetraspanin proteins mediate cellular penetration, invasion, and fusion events and define a novel type of membrane microdomain. Annu. Rev. Cell Dev. Biol. 2003, 19, 397–422. [Google Scholar] [CrossRef]
- Yu, H.; Bowden, D.W.; Spray, B.J.; Rich, S.S.; Freedman, B.I. Identification of human plasma kallikrein gene polymorphisms and evaluation of their role in end-stage renal disease. Hypertension 1998, 31, 906–911. [Google Scholar] [CrossRef][Green Version]
- Avigdor, S.; Alhenc-Gelas, F.; Bouhnik, J. Effects of thyroid hormones on urinary and renal kallikreins. Am. J. Physiol. Endocrinol. Metab. 1992, 263, E430–E434. [Google Scholar] [CrossRef] [PubMed]
- Alexander-Curtis, M.; Pauls, R.; Chao, J.; Volpi, J.J.; Bath, P.M.; Verdoorn, T.A. Human tissue kallikrein in the treatment of acute ischemic stroke. Ther. Adv. Neurol. Disord. 2019, 12, 1756286418821918. [Google Scholar] [CrossRef]
- Erem, C.; Ersoz, H.; Karti, S.; Ukinç, K.; Hacihasanoglu, A.; Değer, O.; Telatar, M. Blood coagulation and fibrinolysis in patients with hyperthyroidism. J. Endocrinol. Investig. 2002, 25, 345–350. [Google Scholar] [CrossRef] [PubMed]
- Greenberg, C.S.; Birckbichler, P.J.; Rice, R.H. Transglutaminases: Multifunctional cross-linking enzymes that stabilize tissues. FASEB J. 1991, 5, 3071–3077. [Google Scholar] [CrossRef]
- Parry-Billings, M.; Dimitriadis, G.; Leighton, B.; Bond, J.; Bevan, S.; Opara, E.; Newsholme, E. Effects of hyperthyroidism and hypothyroidism on glutamine metabolism by skeletal muscle of the rat. Biochem. J. 1990, 272, 319–322. [Google Scholar] [CrossRef] [PubMed]
- Ugi, S.; Imamura, T.; Ricketts, W.; Olefsky, J.M. Protein phosphatase 2A forms a molecular complex with Shc and regulates Shc tyrosine phosphorylation and downstream mitogenic signaling. Mol. Cell. Biol. 2002, 22, 2375–2387. [Google Scholar] [CrossRef]
- Östman, A.; Böhmer, F.-D. Regulation of receptor tyrosine kinase signaling by protein tyrosine phosphatases. Trends Cell Biol. 2001, 11, 258–266. [Google Scholar] [CrossRef]
- Shorning, B.Y.; Dass, M.S.; Smalley, M.J.; Pearson, H.B. The PI3K-AKT-mTOR pathway and prostate cancer: At the crossroads of AR, MAPK, and WNT signaling. Int. J. Mol. Sci. 2020, 21, 4507. [Google Scholar] [CrossRef]
- Wessel, D.M.; Flügge, U.I. A method for the quantitative recovery of protein in dilute solution in the presence of detergents and lipids. Anal. Biochem. 1984, 138, 141–143. [Google Scholar] [CrossRef]
- Alfadda, A.A.; Benabdelkamel, H.; Masood, A.; Moustafa, A.; Sallam, R.; Bassas, A.; Duncan, M. Proteomic analysis of mature adipo cytes from obese patients in relation to aging. Exp. Gerontol. 2013, 48, 1196–1203. [Google Scholar] [CrossRef] [PubMed]
- Benabdelkamel, H.; Masood, A.; Almidani, G.M.; Alsadhan, A.A.; Bassas, A.F.; Duncan, M.W.; Alfadda, A.A. Mature adipocyte proteome reveals differentially altered protein abundances between lean, overweight and morbidly obese human subjects. Mol. Cell. Endocrinol. 2015, 401, 142–154. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Normal Range | Hyperthyroid | Euthyroid | p-Value | |
|---|---|---|---|---|
| N | 9 | 9 | ||
| Age (years) | 39.6 ± 10.6 | 39.6 ± 10.6 | ||
| Glucose (mmol/L) | 4.07–5.83 | 5.3 ± 0.8 | 5.2 ± 0.5 | 0.19 |
| Urea (mmol/L) | 2.5–6.4 | 3.9 ± 0.9 | 4.6 ± 0.8 | 0.18 |
| Creatinine (µmol/L) | 53–115 | 61.1 ± 12.7 | 66.3 ± 12.0 | 0.39 |
| Sodium (mmol/L) | 136–145 | 138.6 ± 2.1 | 138 ± 0.9 | 0.16 |
| Potassium (mmol/L) | 3.5–5.1 | 4.2 ± 0.2 | 4.3 ± 0.4 | 0.40 |
| Aspartate transaminase (IU/L) | 15–37 | 34.8 ± 9.1 | 31.8 ± 4.5 | 0.38 |
| Alanine transaminase (IU/L) | 20–65 | 17.5 ± 5 | 15.8 ± 2.0 | 0.40 |
| Alkaline phosphatase (IU/L) | 40–150 | 114.6 ± 53.7 | 120.2 ± 28.9 | 0.30 |
| FT4 (pmol/L) | 11.5–22.7 | 35.4 ± 9.9 | 17.0 ± 2.8 | 0.001 |
| TSH (mIU/L) | 0.25–0.5 | 0.014 ± 0.01 | 0.8 ± 0.4 | 0.00068 |
| Total cholesterol (mmol/L) | 3.2–5.2 | 4.6 ± 1.0 | 4.9 ± 0.8 | 0.10 |
| HDL cholesterol (mmol/L) | 0.96–2.15 | 2.8 ± 0.9 | 3.1 ± 0.7 | 0.02 |
| LDL cholesterol (mmol/L) | 1.84–4.25 | 2.8 ± 0.9 | 3.1 ± 0.7 | 0.09 |
| Triglycerides (mmol/L) | 0.4–1.48 | 1.1 ± 0.6 | 0.9 ± 0.2 | 0.14 |
| Sl No | Spot No. | Accession No. a | Protein Name | MASCOT ID | p-Value b (ANOVA) | Ratio c Hyper\Euth | EXP d |
|---|---|---|---|---|---|---|---|
| 1 | 554 | P02787 | Serotransferrin | TRFE_HUMAN | 0.048 | 1.7 | UP |
| 2 | 871 | P06702 | Protein S100-A9 | S10A9_HUMAN | 0.049 | 3.4 | UP |
| 3 | 933 | P04217 | Alpha-1B-glycoprotein | A1BG_HUMAN | 9.410−4 | 2 | UP |
| 4 | 227 | P02768 | Serum albumin | ALBU_HUMAN | 0.006 | 2.1 | UP |
| 5 | 210 | Q6ZMW3 | Echinoderm microtubule-associated protein-like 6 | EMAL6_HUMAN | 0.001 | 1.6 | UP |
| 6 | 983 | P02766 | Transthyretin | TTHY_HUMAN | 0.004 | 2.5 | UP |
| 7 | 253 | P02768 | Serum albumin | ALBU_HUMAN | 0.017 | 1.6 | UP |
| 8 | 237 | P01833 | Polymeric immunoglobulin receptor | PIGR_HUMAN | 0.017 | 1.7 | UP |
| 9 | 258 | P04217 | Alpha-1B-glycoprotein | A1BG_HUMAN | 0.024 | 1.6 | UP |
| 10 | 166 | P00450 | Ceruloplasmin | CERU_HUMAN | 0.029 | 1.8 | UP |
| 11 | 588 | Q9UQ35 | Serine/arginine repetitive matrix protein 2 | SRRM2_HUMAN | 0.031 | 1.7 | UP |
| 12 | 593 | O00443 | Phosphatidylinositol 4-phosphate 3-kinase C2 domain-containing subunit alpha | P3C2A_HUMAN | 0.034 | 3.2 | UP |
| 13 | 318 | P02768 | Serum albumin | ALBU_HUMAN | 0.036 | 1.6 | UP |
| 14 | 526 | Q16769 | Glutaminyl-peptide cyclotransferase | QPCT_HUMAN | 0.01 | 1.8 | UP |
| 15 | 215 | Q6FIF0 | AN1-type zinc finger protein 6 | ZFAN6_HUMAN | 0.02 | 1.6 | UP |
| 16 | 266 | Q92878 | DNA repair protein RAD50 | RAD50_HUMAN | 0.047 | 1.5 | UP |
| 17 | 584 | Q9BUB4 | tRNA-specific adenosine deaminase 1 | ADAT1_HUMAN | 0.051 | −1.5 | DOWN |
| 18 | 162 | P03952 | Plasma kallikrein | KLKB1_HUMAN | 0.023 | −2.5 | DOWN |
| 19 | 618 | Q15257 | Serine/threonine-protein phosphatase 2A activator | PTPA_HUMAN | 0.03 | −2.0 | DOWN |
| 20 | 163 | Q96PF1 | Protein glutamine gamma-glutamyl transferase Z | TGM7_HUMAN | 0.031 | −1.6 | DOWN |
| 21 | 799 | Q6U7Q0 | Zinc finger protein 322 | ZN322_HUMAN | 0.029 | −2.5 | DOWN |
| 22 | 882 | P29508 | SERPINB3 | SPB3_HUMAN | 0.009 | 3 | UP |
| 23 | 765 | Q9UKF7 | Cytoplasmic phosphatidylinositol transfer protein 1 | PITC1_HUMAN | 0.031 | 1.7 | UP |
| 24 | 902 | Q16769 | Glutaminyl-peptide cyclotransferase | QPCT_HUMAN | 0.048 | 1.5 | UP |
| 25 | 281 | P01833 | Polymeric immunoglobulin receptor | PIGR_HUMAN | 0.011 | 1.7 | UP |
| 26 | 998 | Q8N3U1 | Putative uncharacterized protein LOC400692 | YS014_HUMAN | 0.011 | 1.5 | UP |
| 27 | 506 | Q4V348 | Zinc finger protein 658B | Z658B_HUMAN | 0.016 | 1.5 | UP |
| 28 | 655 | O00560 | Syntenin-1 | SDCB1_HUMAN | 0.016 | 1.8 | UP |
| 29 | 1032 | Q9H299 | SH3 domain-binding glutamic acid-rich-like protein 3 | SH3L3_HUMAN | 0.02 | −3.2 | DOWN |
| 30 | 959 | P13645 | Keratin, type I cytoskeletal 10 | K1C10_HUMAN | 0.028 | 2.6 | UP |
| 31 | 770 | P01833 | Polymeric immunoglobulin receptor | PIGR_HUMAN | 0.054 | −1.9 | DOWN |
| 32 | 498 | P04746 | Pancreatic alpha-amylase | AMYP_HUMAN | 0.052 | 1.5 | UP |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Benabdelkamel, H.; Masood, A.; Ekhzaimy, A.A.; Alfadda, A.A. Proteomics Profiling of the Urine of Patients with Hyperthyroidism after Anti-Thyroid Treatment. Molecules 2021, 26, 1991. https://doi.org/10.3390/molecules26071991
Benabdelkamel H, Masood A, Ekhzaimy AA, Alfadda AA. Proteomics Profiling of the Urine of Patients with Hyperthyroidism after Anti-Thyroid Treatment. Molecules. 2021; 26(7):1991. https://doi.org/10.3390/molecules26071991
Chicago/Turabian StyleBenabdelkamel, Hicham, Afshan Masood, Aishah A. Ekhzaimy, and Assim A. Alfadda. 2021. "Proteomics Profiling of the Urine of Patients with Hyperthyroidism after Anti-Thyroid Treatment" Molecules 26, no. 7: 1991. https://doi.org/10.3390/molecules26071991
APA StyleBenabdelkamel, H., Masood, A., Ekhzaimy, A. A., & Alfadda, A. A. (2021). Proteomics Profiling of the Urine of Patients with Hyperthyroidism after Anti-Thyroid Treatment. Molecules, 26(7), 1991. https://doi.org/10.3390/molecules26071991