
Neuroprotective Effects of Thymoquinone by the Modulation of ER Stress and Apoptotic Pathway in In Vitro Model of Excitotoxicity




Abstract

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
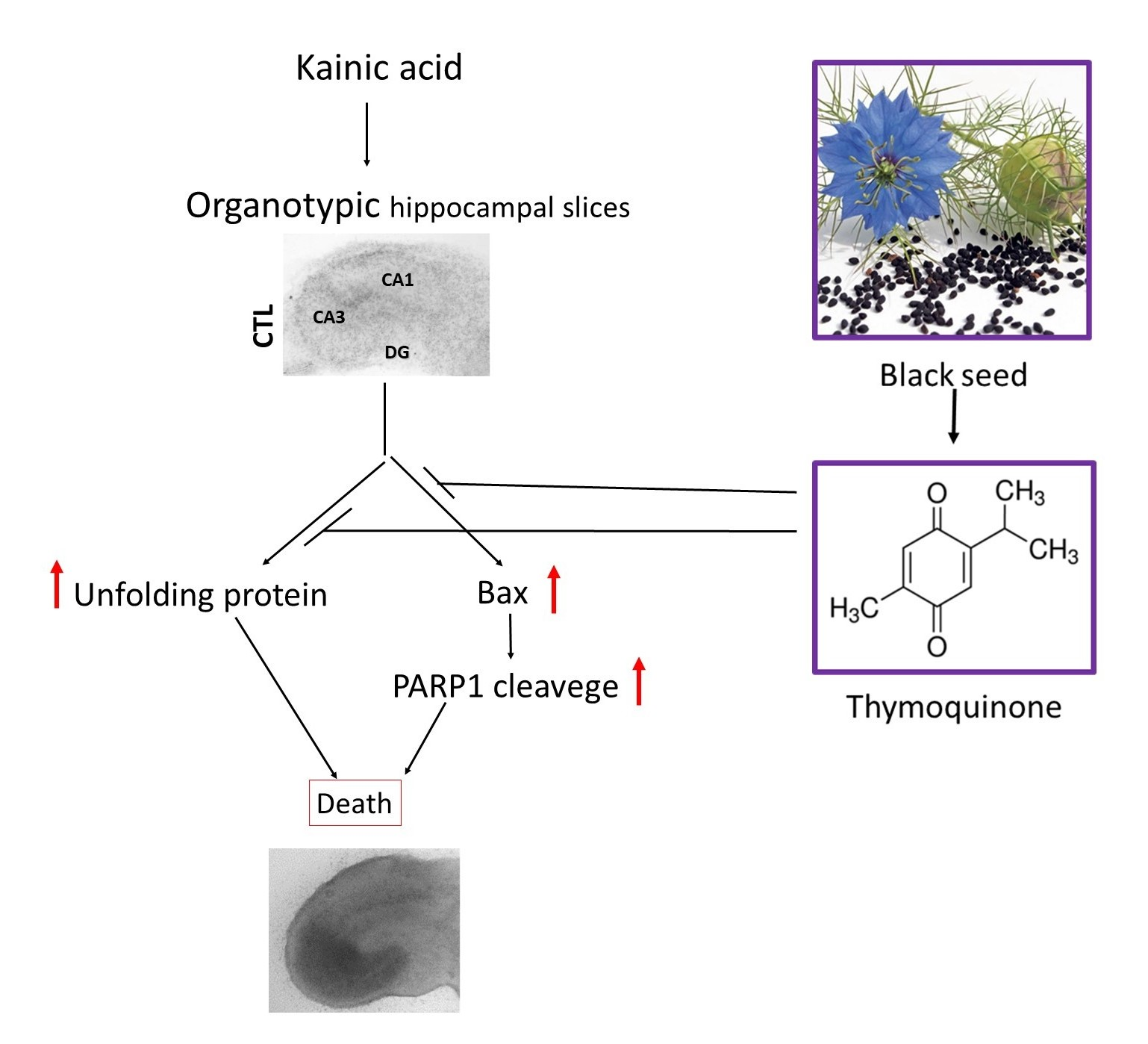
1. Introduction
2. Results
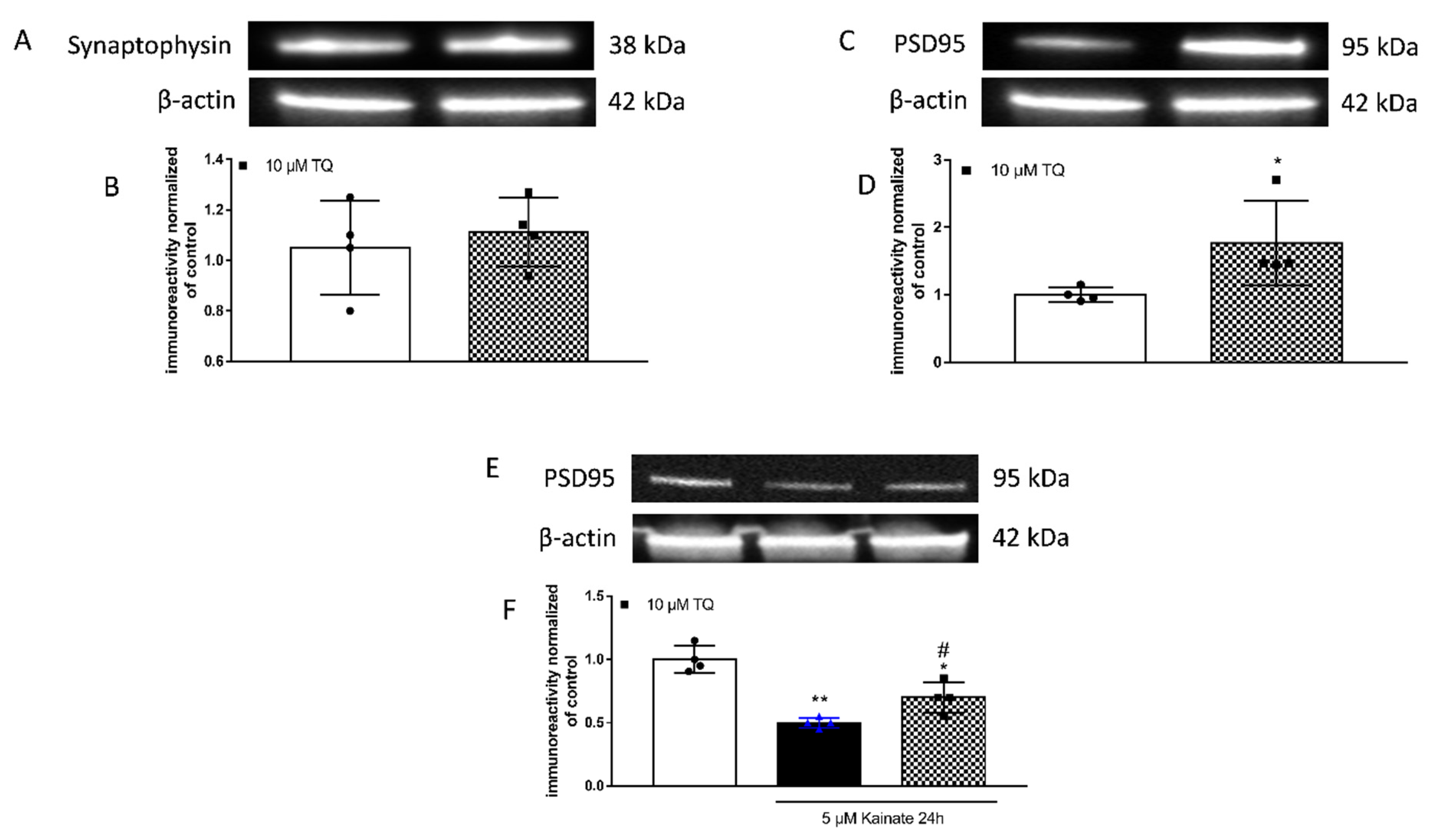
2.1. Effect of TQ on PSD95 Levels
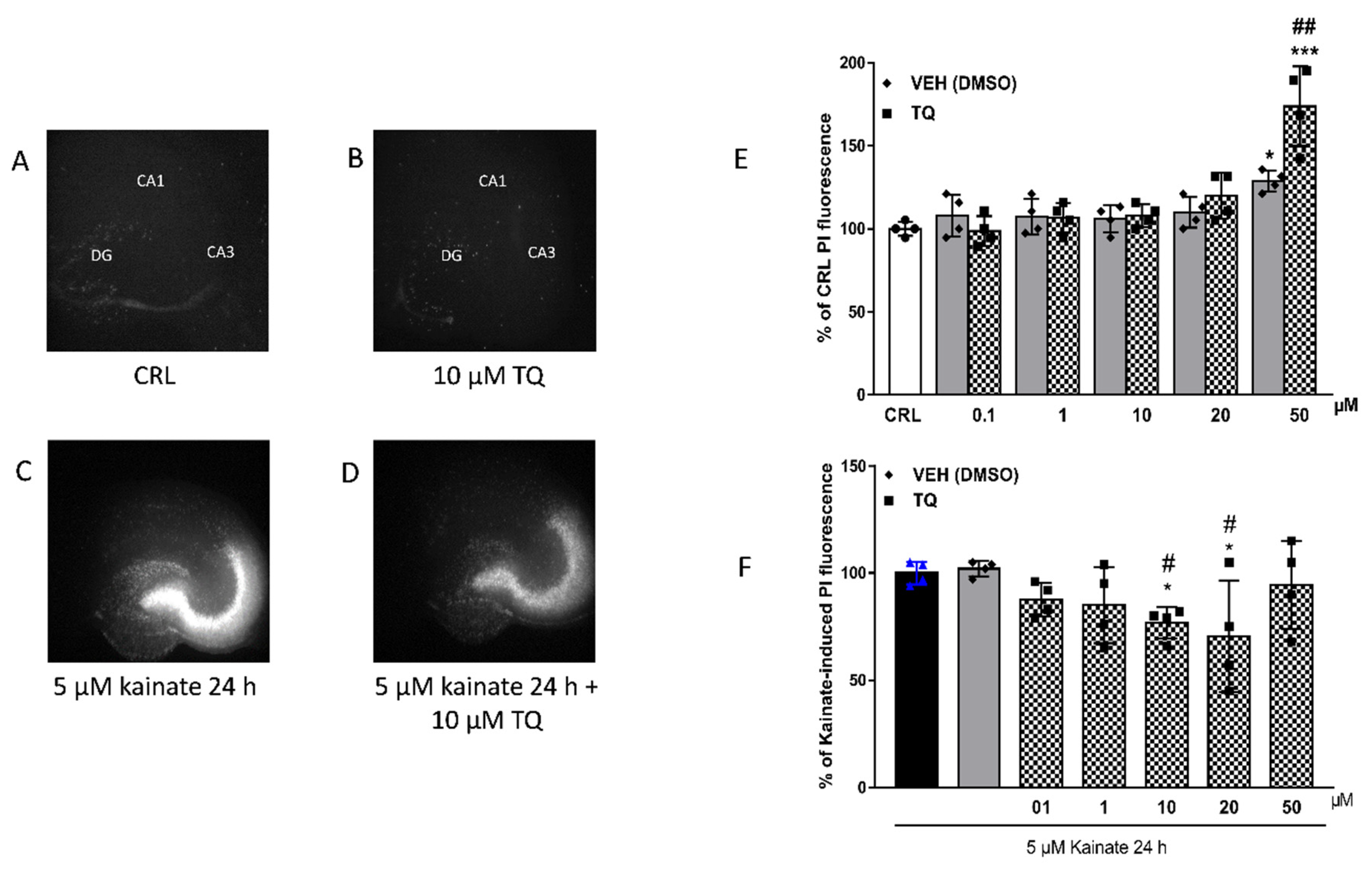
2.2. Effect of TQ on KA-Induced Neuronal Cell Death
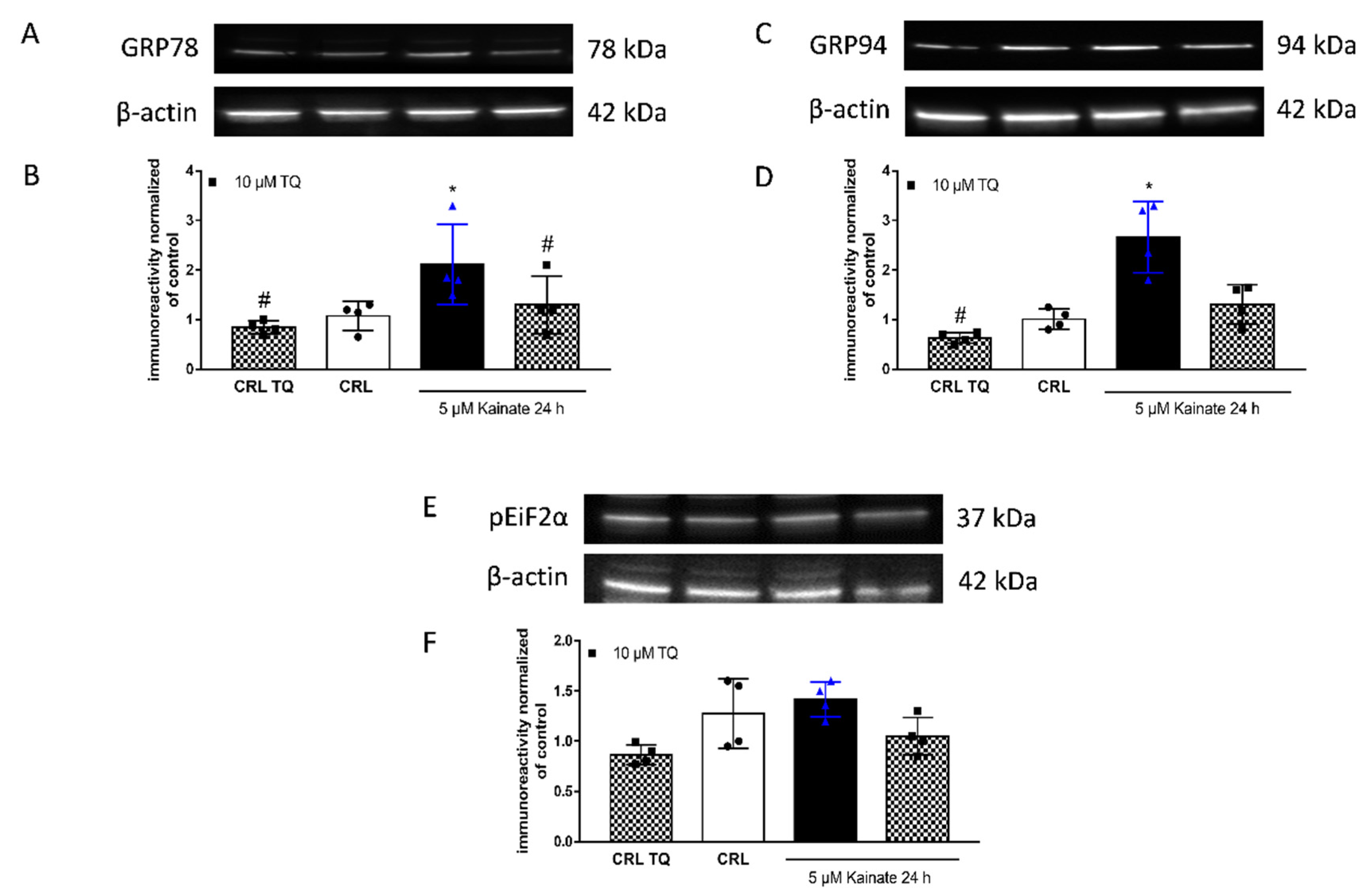
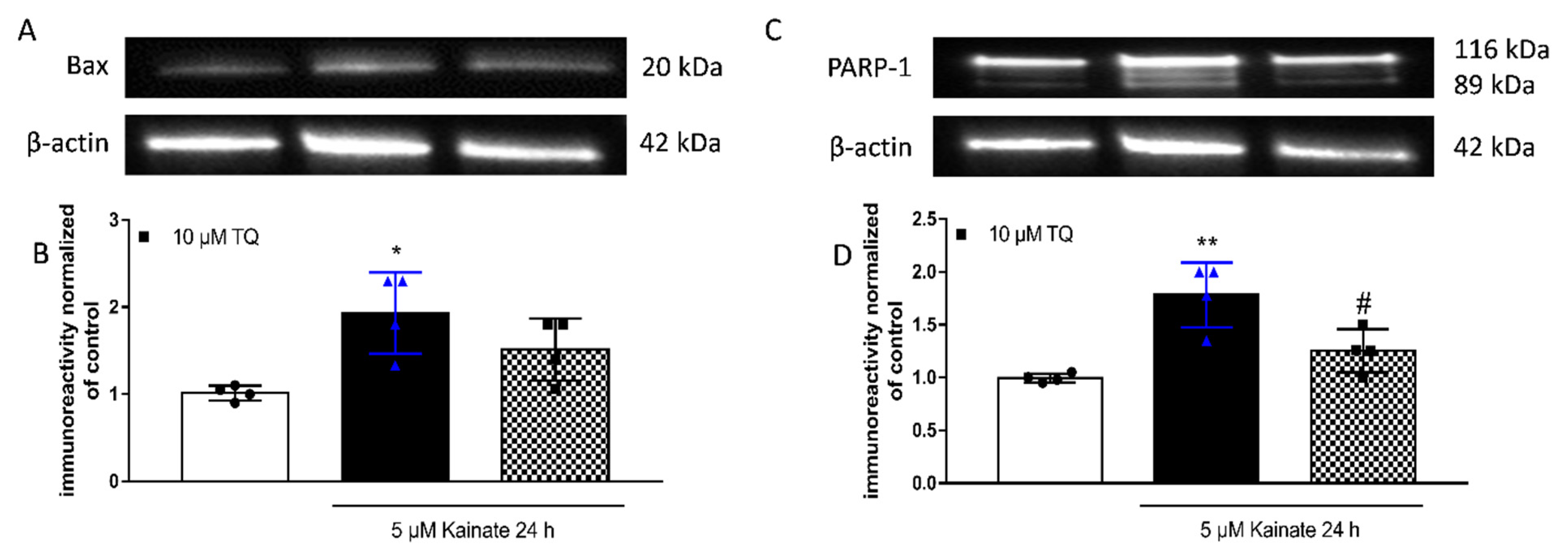
2.3. Effect of TQ on KA-Induced Apoptosis
3. Discussion
4. Materials and Methods
4.1. Animals
4.2. Materials
4.3. Preparation of Rat Organotypic Hippocampal Slice Cultures
4.4. Western Blot Analysis
4.5. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Sample Availability
Abbreviations
| TQ, thymoquinone; |
| SCE, Schisandra chinensis extract; |
| AA, ascorbic acid; |
| PSD95, postsynaptic density protein 95; |
| BDNF, brain-derived neurotrophic factor; |
| ER stress, Endoplasmic Reticulum stress; |
| TRIB3, tribblespseudokinase 3; |
| AKT, protein kinase B; |
| KA, kainic acid; |
| I/R, ischemia-reperfusion; |
| TLE, temporal lobe epilepsy; |
| CA3, Cornu Ammonis 3; |
| WB, Western blotting; |
| GRP78, Glucose Regulated Protein 78; |
| GRP94, Glucose Regulated Protein 94; |
| eIF2a, eukaryotic translation initiation factor 2 subunit alpha; |
| PTZ, pentylenetetrazole; |
| Bcl-2, B-cell lymphoma 2; |
| BAX, Bcl-2 Associated X-protein; |
| PARP, Poly [ADP-ribose] polymerase; |
| PARP-1, Poly [ADP-ribose]polymerase 1; |
| Bim, Bcl-2-like protein 11; |
| Bad, BCL2 associated agonist of cell death; |
| NSAIDs, nonsteroidal anti-inflammatory drugs; |
| COX-2, cyclooxygenase-2; |
| COXIBs, cyclooxygenase-2 selective inhibitors; |
| UPR, unfolded protein response; |
| PI, Propidium iodide; |
| SDS, Sodium Dodecyl Sulphate; |
| BCA, bicinchoninic acid; |
| TBS-T, Tris Buffered Saline-Tween; |
| HRP, Horseradish Peroxidase; |
| ECL-plus, Enhanced ChemiLuminescence; |
| AU, arbitrary units; |
| SEM, standard error of the mean. |
References
- Mansour, M.A.; Nagi, M.N.; El-Khatib, A.S.; Al-Bekairi, A.M. Effects of thymoquinone on antioxidant enzyme activities, lipid peroxidation and dt-diaphorase in different tissues of mice: A possible mechanism of action. Cell Biochem. Funct. 2002, 20, 143–151. [Google Scholar] [CrossRef] [PubMed]
- Badary, O.A.; Taha, R.A.; El-Din, A.M.G.; Abdel-Wahab, M.H. Thymoquinone is a potent superoxide anion scavenger. Drug Chem. Toxicol. 2003, 26, 87–98. [Google Scholar] [CrossRef]
- Chaieb, K.; Kouidhi, B.; Jrah, H.; Mahdouani, K.; Bakhrouf, A. Antibacterial activity of Thymoquinone, an active principle of Nigella sativa and its potency to prevent bacterial biofilm formation. BMC Complement. Altern. Med. 2011, 11, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Kouidhi, B.; Zmantar, T.; Jrah, H.; Souiden, Y.; Chaieb, K.; Mahdouani, K.; Bakhrouf, A. Antibacterial and resistance-modifying activities of thymoquinone against oral pathogens. Ann. Clin. Microbiol. Antimicrob. 2011, 10, 1–7. [Google Scholar] [CrossRef]
- Salem, M.L. Immunomodulatory and therapeutic properties of the Nigella sativa L. seed. Int. Immunopharmacol. 2005, 5, 1749–1770. [Google Scholar] [CrossRef]
- Mahgoub, A.A. Thymoquinone protects against experimental colitis in rats. Toxicol. Lett. 2003, 143, 133–143. [Google Scholar] [CrossRef]
- Houghton, P.J.; Zarka, R.; De Las Heras, B.; Hoult, J.R.S. Fixed oil of Nigella sativa and derived thymoquinone inhibit eicosanoid generation in leukocytes and membrane lipid peroxidation. Planta Med. 1995, 61, 33–36. [Google Scholar] [CrossRef] [PubMed]
- El Gazzar, M.; El Mezayen, R.; Marecki, J.C.; Nicolls, M.R.; Canastar, A.; Dreskin, S.C. Anti-inflammatory effect of thymoquinone in a mouse model of allergic lung inflammation. Int. Immunopharmacol. 2006, 6, 1135–1142. [Google Scholar] [CrossRef] [PubMed]
- El-Mahmoudy, A.; Shimizu, Y.; Shiina, T.; Matsuyama, H.; El-Sayed, M.; Takewaki, T. Successful abrogation by thymoquinone against induction of diabetes mellitus with streptozotocin via nitric oxide inhibitory mechanism. Int. Immunopharmacol. 2005, 5, 195–207. [Google Scholar] [CrossRef] [PubMed]
- Nagi, M.N.; Almakki, H.A. Thymoquinone supplementation induces quinone reductase and glutathione transferase in mice liver: Possible role in protection against chemical carcinogenesis and toxicity. Phyther. Res. 2009, 23, 1295–1298. [Google Scholar] [CrossRef] [PubMed]
- Alenzi, F.Q.; El-Sayed El-Bolkiny, Y.; Salem, M.L. Protective effects of Nigella sativa oil and thymoquinone against toxicity induced by the anticancer drug cyclophosphamide. Br. J. Biomed. Sci. 2010, 67, 20–28. [Google Scholar] [CrossRef] [PubMed]
- Khattab, M.M.; Nagi, M.N. Thymoquinone supplementation attenuates hypertension and renal damage in nitric oxide deficient hypertensive rats. Phyther. Res. 2007, 21, 410–414. [Google Scholar] [CrossRef] [PubMed]
- Pari, L.; Sankaranarayanan, C. Beneficial effects of thymoquinone on hepatic key enzymes in streptozotocin-nicotinamide induced diabetic rats. Life Sci. 2009, 85, 830–834. [Google Scholar] [CrossRef] [PubMed]
- Hawsawi, Z.A.; Ali, B.A.; Bamosa, A.O. Effect of Nigella sativa (black seed) and thymoquinone on blood glucose in albino rats. Ann. Saudi Med. 2001, 21, 242–244. [Google Scholar] [CrossRef]
- Banerjee, S.; Padhye, S.; Azmi, A.; Wang, Z.; Philip, P.A.; Kucuk, O.; Sarkar, F.H.; Mohammad, R.M. Review on molecular and therapeutic potential of thymoquinone in cancer. Nutr. Cancer 2010, 62, 938–946. [Google Scholar] [CrossRef]
- Shoieb, A.M.; Elgayyar, M.; Dudrick, P.S.; Bell, J.L.; Tithof, P.K. In vitro inhibition of growth and induction of apoptosis in cancer cell lines by thymoquinone. Int. J. Oncol. 2003, 22, 107–113. [Google Scholar] [CrossRef]
- Woo, C.C.; Loo, S.Y.; Gee, V.; Yap, C.W.; Sethi, G.; Kumar, A.P.; Tan, K.H.B. Anticancer activity of thymoquinone in breast cancer cells: Possible involvement of PPAR-γ pathway. Biochem. Pharmacol. 2011, 82, 464–475. [Google Scholar] [CrossRef] [PubMed]
- Samarghandian, S.; Farkhondeh, T.; Samini, F. A Review on Possible Therapeutic Effect of Nigella sativa and Thymoquinone in Neurodegenerative Diseases. CNS Neurol. Disord. Drug Targets 2018, 17, 412–420. [Google Scholar] [CrossRef] [PubMed]
- Ragheb, A.; Attia, A.; Eldin, W.S.; Elbarbry, F.; Gazarin, S.; Shoker, A. The protective effect of thymoquinone, an antioxidant and anti-inflammatory agent, against renal injury: A review. Saudi J. Kidney Dis. Transpl. 2009, 20, 741–752. [Google Scholar]
- Isaev, N.K.; Chetverikov, N.S.; Stelmashook, E.V.; Genrikhs, E.E.; Khaspekov, L.G.; Illarioshkin, S.N. Thymoquinone as a Potential Neuroprotector in Acute and Chronic Forms of Cerebral Pathology. Biochemistry 2020, 85, 167–176. [Google Scholar] [CrossRef] [PubMed]
- Shao, Y.Y.; Li, B.; Huang, Y.M.; Luo, Q.; Xie, Y.M.; Chen, Y.H. Thymoquinone attenuates brain injury via an antioxidative pathway in a status epilepticus rat model. Transl. Neurosci. 2017, 8, 9–14. [Google Scholar] [CrossRef]
- Jang, Y.; Lee, J.H.; Lee, M.J.; Kim, S.J.; Ju, X.; Cui, J.; Zhu, J.; Lee, Y.L.; Namgung, E.; Sung, H.W.J.; et al. Schisandra extract and ascorbic acid synergistically enhance cognition in mice through modulation of mitochondrial respiration. Nutrients 2020, 12, 897. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Wu, J.; Liu, F.; Tong, L.; Chen, Z.; Chen, J.; He, H.; Xu, R.; Ma, Y.; Huang, C. Neuroprotective effects of anthocyanins and its major component cyanidin-3-O-glucoside (C3G) in the central nervous system: An outlined review. Eur. J. Pharmacol. 2019, 858, 172500. [Google Scholar] [CrossRef]
- Bouhlel, A.; Mosbah, B.; Abdallah, H.; Ribault, C.; Viel, R.; Mannaï, S.; Corlu, A.; Abdennebi, B. Thymoquinone prevents endoplasmic reticulum stress and mitochondria-induced apoptosis in a rat model of partial hepatic warm ischemia reperfusion. Biomed. Pharmacother. 2017, 94, 964–973. [Google Scholar] [CrossRef] [PubMed]
- Landucci, E.; Llorente, I.L.; Anuncibay-Soto, B.; Pellegrini-Giampietro, D.E.; Fernández-López, A. Bicuculline Reverts the Neuroprotective Effects of Meloxicam in an Oxygen and Glucose Deprivation (OGD) Model of Organotypic Hippocampal Slice Cultures. Neuroscience 2018, 386, 68–78. [Google Scholar] [CrossRef] [PubMed]
- Ullah, I.; Badshah, H.; Naseer, M.I.; Lee, H.Y.; Kim, M.O. Thymoquinone and Vitamin C Attenuates Pentylenetetrazole-Induced Seizures Via Activation of GABAB1 Receptor in Adult Rats Cortex and Hippocampus. Neuromol. Med. 2015, 17, 35–46. [Google Scholar] [CrossRef]
- Landucci, E.; Gencarelli, M.; Mazzantini, C.; Laurino, A.; Pellegrini-Giampietro, D.E.; Raimondi, L. N-(3-Ethoxy-phenyl)-4-pyrrolidin-1-yl-3-trifluoromethyl-benzamide (EPPTB) prevents 3-iodothyronamine (T1AM)-induced neuroprotection against kainic acid toxicity. Neurochem. Int. 2019, 129, 104460. [Google Scholar] [CrossRef]
- Landucci, E.; Pellegrini-Giampietro, D.E.; Bilia, A.R.; Bergonzi, M.C. Enhanced neuroprotective effects of panax ginseng g115® and ginkgo biloba gk501® combinations in vitro models of excitotoxicity. Int. J. Mol. Sci. 2019, 20, 5872. [Google Scholar] [CrossRef]
- Korhonen, L.; Belluardo, N.; Lindholm, D. Regulation of X-chromosome-linked inhibitor of apoptosis protein in kainic acid-induced death in the rat hippocampus. Mol. Cell. Neurosci. 2001, 17, 364–372. [Google Scholar] [CrossRef]
- Zhao, Y.; Spigolon, G.; Bonny, C.; Culman, J.; Vercelli, A.; Herdegen, T. The JNK inhibitor D-JNKI-1 blocks apoptotic JNK signaling in brain mitochondria. Mol. Cell. Neurosci. 2012, 49, 300–310. [Google Scholar] [CrossRef]
- Radu, B.M.; Epureanu, F.B.; Radu, M.; Fabene, P.F.; Bertini, G. Nonsteroidal anti-inflammatory drugs in clinical and experimental epilepsy. Epilepsy Res. 2017, 131, 15–27. [Google Scholar] [CrossRef]
- Dhir, A. Curcumin in epilepsy disorders. Phyther. Res. 2018, 32, 1865–1875. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.R. Control of Granule Cell Dispersion by Natural Materials Such as Eugenol and Naringin: A Potential Therapeutic Strategy Against Temporal Lobe Epilepsy. J. Med. Food 2016, 19, 730–736. [Google Scholar] [CrossRef]
- Shanmugam, M.K.; Arfuso, F.; Kumar, A.P.; Wang, L.; Goh, B.C.; Ahn, K.S.; Bishayee, A.; Sethi, G. Modulation of diverse oncogenic transcription factors by thymoquinone, an essential oil compound isolated from the seeds of Nigella sativa Linn. Pharmacol. Res. 2018, 129, 357–364. [Google Scholar] [CrossRef]
- Shanmugam, M.K.; Ahn, K.S.; Hsu, A.; Woo, C.C.; Yuan, Y.; Tan, K.H.B.; Chinnathambi, A.; Alahmadi, T.A.; Alharbi, S.A.; Koh, A.P.F.; et al. Thymoquinone Inhibits Bone Metastasis of Breast Cancer Cells Through Abrogation of the CXCR4 Signaling Axis. Front. Pharmacol. 2018, 9, 1294. [Google Scholar] [CrossRef]
- Firdaus, F.; Zafeer, M.F.; Anis, E.; Ahmad, F.; Hossain, M.M.; Ali, A.; Afzal, M. Evaluation of phytomedicinal efficacy of thymoquinone against Arsenic induced mitochondrial dysfunction and cytotoxicity in SH-SY5Y cells. Phytomedicine 2019, 54, 224–230. [Google Scholar] [CrossRef]
- Tabeshpour, J.; Mehri, S.; Abnous, K.; Hosseinzadeh, H. Neuroprotective Effects of Thymoquinone in Acrylamide-Induced Peripheral Nervous System Toxicity Through MAPKinase and Apoptosis Pathways in Rat. Neurochem. Res. 2019, 44, 1101–1112. [Google Scholar] [CrossRef] [PubMed]
- Tabeshpour, J.; Mehri, S.; Abnous, K.; Hosseinzadeh, H. Role of Oxidative Stress, MAPKinase and Apoptosis Pathways in the Protective Effects of Thymoquinone Against Acrylamide-Induced Central Nervous System Toxicity in Rat. Neurochem. Res. 2020, 45, 254–267. [Google Scholar] [CrossRef] [PubMed]
- Zhang, P.; Lisman, J.E. Activity-dependent regulation of synaptic strength by PSD-95 in CA1 neurons. J. Neurophysiol. 2012, 107, 1058–1066. [Google Scholar] [CrossRef]
- De Bartolomeis, A.; Latte, G.; Tomasetti, C.; Iasevoli, F. Glutamatergic postsynaptic density protein dysfunctions in synaptic plasticity and dendritic spines morphology: Relevance to schizophrenia and other behavioral disorders pathophysiology, and implications for novel therapeutic approaches. Mol. Neurobiol. 2014, 49, 484–511. [Google Scholar] [CrossRef]
- Jiang, Q.; Wang, J.; Wu, X.; Jiang, Y. Alterations of NR2B and PSD-95 expression after early-life epileptiform discharges in developing neurons. Int. J. Dev. Neurosci. 2007, 25, 165–170. [Google Scholar] [CrossRef] [PubMed]
- Morimoto, K.; Fahnestock, M.; Racine, R.J. Kindling and status epilepticus models of epilepsy: Rewiring the brain. Prog. Neurobiol. 2004, 73, 1–60. [Google Scholar] [CrossRef] [PubMed]
- Frasca, A.; Aalbers, M.; Frigerio, F.; Fiordaliso, F.; Salio, M.; Gobbi, M.; Cagnotto, A.; Gardoni, F.; Battaglia, G.S.; Hoogland, G.; et al. Misplaced NMDA receptors in epileptogenesis contribute to excitotoxicity. Neurobiol. Dis. 2011, 43, 507–515. [Google Scholar] [CrossRef] [PubMed]
- Sun, Q.J.; Duan, R.S.; Wang, A.H.; Shang, W.; Zhang, T.; Zhang, X.Q.; Chi, Z.F. Alterations of NR2B and PSD-95 expression in hippocampus of kainic acid-exposed rats with behavioural deficits. Behav. Brain Res. 2009, 201, 292–299. [Google Scholar] [CrossRef] [PubMed]
- Sokka, A.L.; Putkonen, N.; Mudo, G.; Pryazhnikov, E.; Reijonen, S.; Khiroug, L.; Belluardo, N.; Lindholm, D.; Korhonen, L. Endoplasmic reticulum stress inhibition protects against excitotoxic neuronal injury in the rat brain. J. Neurosci. 2007, 27, 901–908. [Google Scholar] [CrossRef] [PubMed]
- Laurino, A.; Landucci, E.; Resta, F.; De Siena, G.; Pellegrini-Giampietro, D.E.; Masi, A.; Mannaioni, G.; Raimondi, L. Anticonvulsant and Neuroprotective Effects of the Thyroid Hormone Metabolite 3-Iodothyroacetic Acid. Thyroid 2018, 28, 1387–1397. [Google Scholar] [CrossRef]
- Wang, M.; Kaufman, R.J. Protein misfolding in the endoplasmic reticulum as a conduit to human disease. Nature 2016, 529, 326–335. [Google Scholar] [CrossRef]
- Ron, D.; Walter, P. Signal integration in the endoplasmic reticulum unfolded protein response. Nat. Rev. Mol. Cell Biol. 2007, 8, 519–529. [Google Scholar] [CrossRef]
- Hetz, C.; Zhang, K.; Kaufman, R.J. Mechanisms, regulation and functions of the unfolded protein response. Nat. Rev. Mol. Cell Biol. 2020, 21, 421–438. [Google Scholar] [CrossRef] [PubMed]
- Gerace, E.; Landucci, E.; Scartabelli, T.; Moroni, F.; Pellegrini-Giampietro, D.E. Rat hippocampal slice culture models for the evaluation of neuroprotective agents. Methods Mol. Biol. 2012, 846, 343–354. [Google Scholar] [PubMed]
- Landucci, E.; Filippi, L.; Gerace, E.; Catarzi, S.; Guerrini, R.; Pellegrini-Giampietro, D.E. Neuroprotective effects of topiramate and memantine in combination with hypothermia in hypoxic-ischemic brain injury in vitro and in vivo. Neurosci. Lett. 2018, 668, 103–107. [Google Scholar] [CrossRef] [PubMed]




Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Landucci, E.; Mazzantini, C.; Buonvicino, D.; Pellegrini-Giampietro, D.E.; Bergonzi, M.C. Neuroprotective Effects of Thymoquinone by the Modulation of ER Stress and Apoptotic Pathway in In Vitro Model of Excitotoxicity. Molecules 2021, 26, 1592. https://doi.org/10.3390/molecules26061592
Landucci E, Mazzantini C, Buonvicino D, Pellegrini-Giampietro DE, Bergonzi MC. Neuroprotective Effects of Thymoquinone by the Modulation of ER Stress and Apoptotic Pathway in In Vitro Model of Excitotoxicity. Molecules. 2021; 26(6):1592. https://doi.org/10.3390/molecules26061592
Chicago/Turabian StyleLanducci, Elisa, Costanza Mazzantini, Daniela Buonvicino, Domenico E. Pellegrini-Giampietro, and Maria Camilla Bergonzi. 2021. "Neuroprotective Effects of Thymoquinone by the Modulation of ER Stress and Apoptotic Pathway in In Vitro Model of Excitotoxicity" Molecules 26, no. 6: 1592. https://doi.org/10.3390/molecules26061592
APA StyleLanducci, E., Mazzantini, C., Buonvicino, D., Pellegrini-Giampietro, D. E., & Bergonzi, M. C. (2021). Neuroprotective Effects of Thymoquinone by the Modulation of ER Stress and Apoptotic Pathway in In Vitro Model of Excitotoxicity. Molecules, 26(6), 1592. https://doi.org/10.3390/molecules26061592