
Anticancer Potential of Damnacanthal and Nordamnacanthal from Morinda elliptica Roots on T-lymphoblastic Leukemia Cells

Abstract
1. Introduction
2. Results
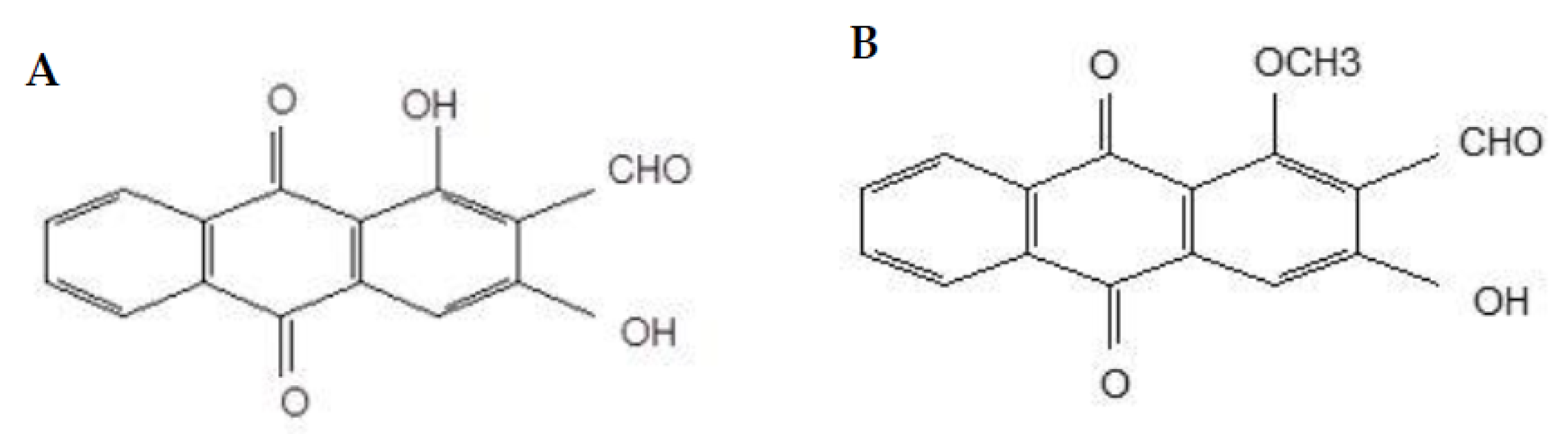
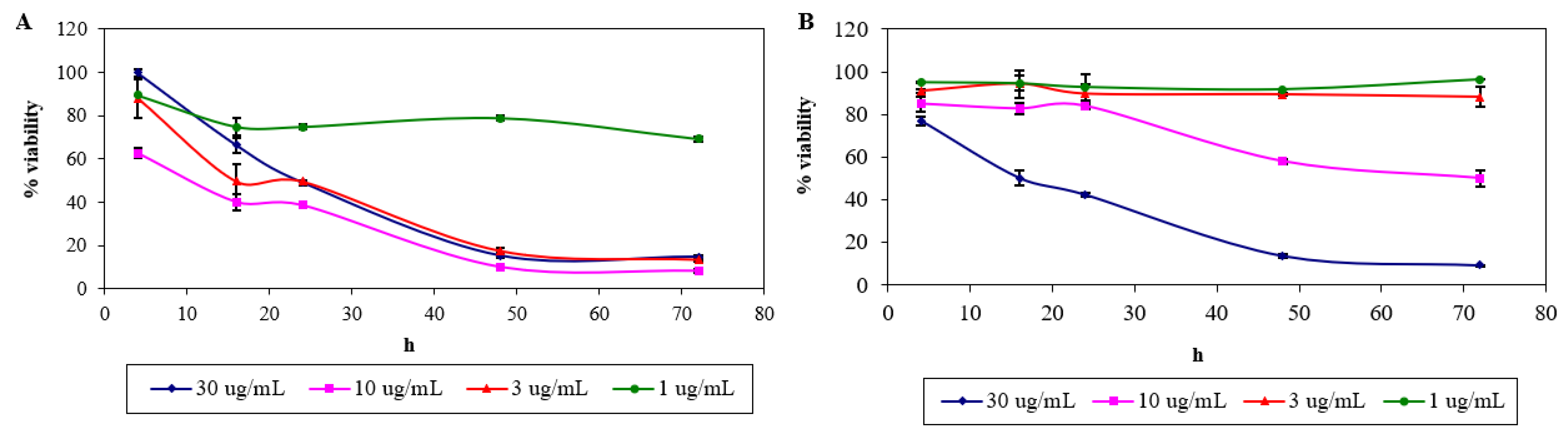
2.1. Cytotoxicity of Nordamnacanthal and Damnacanthal
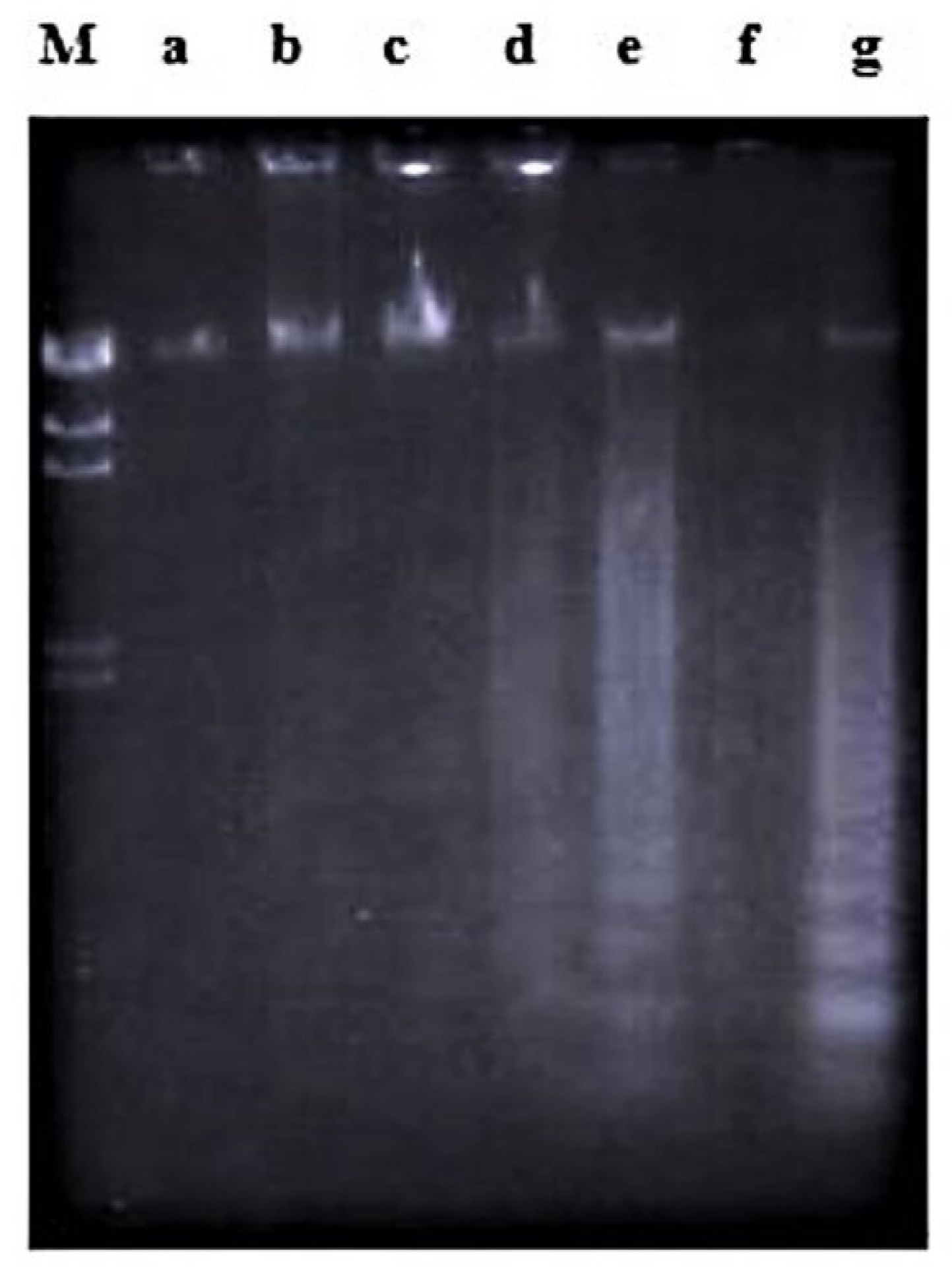
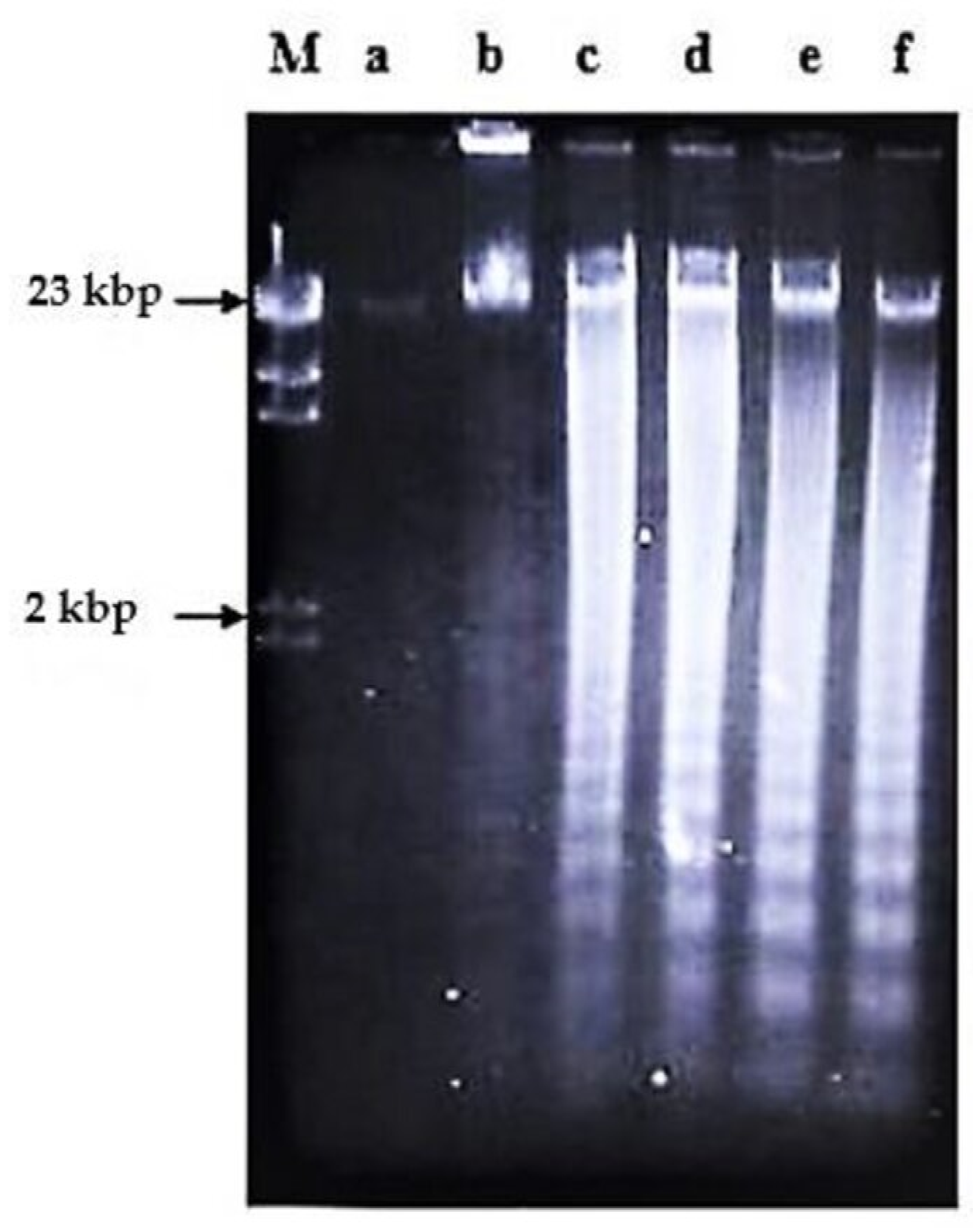
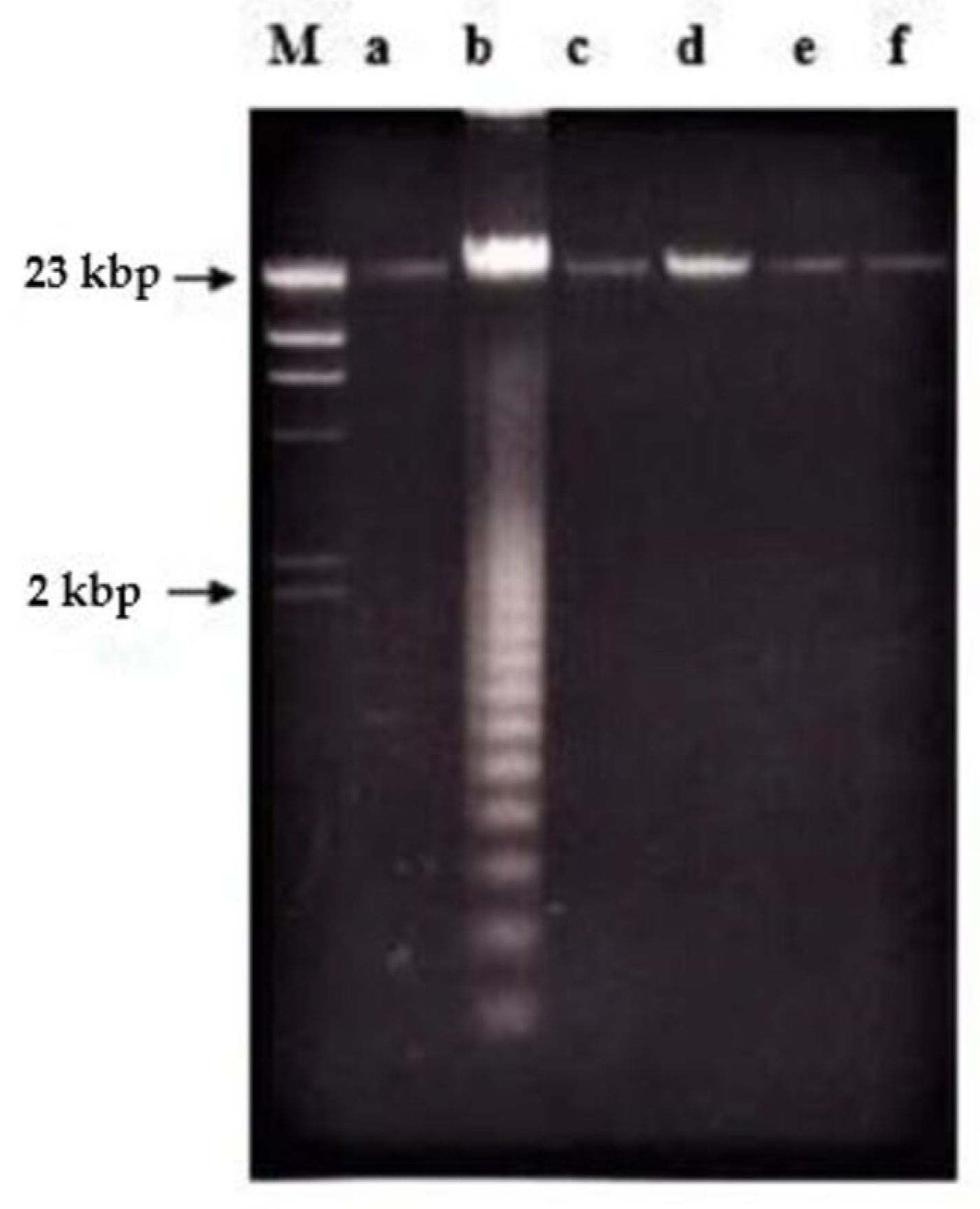
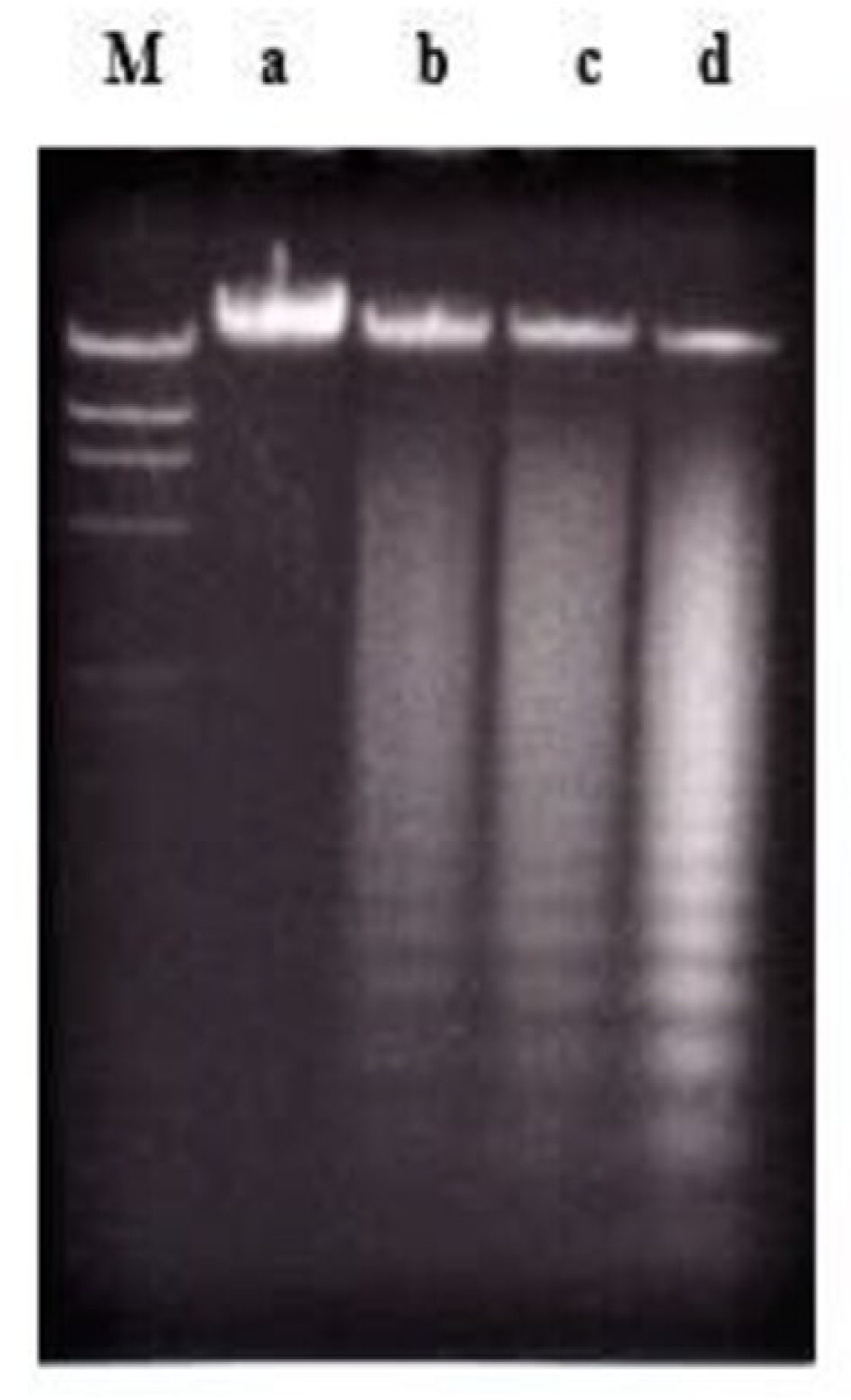
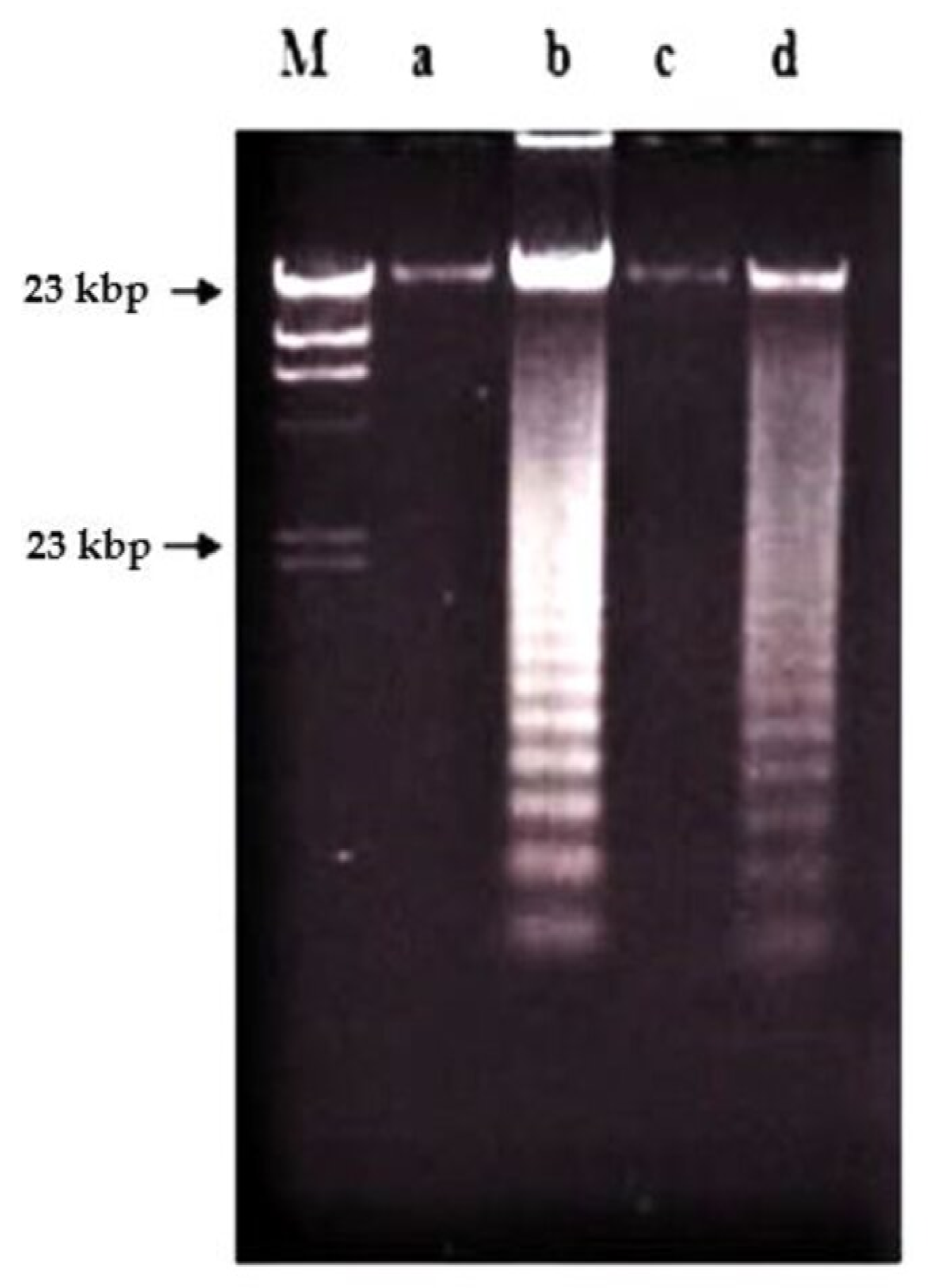
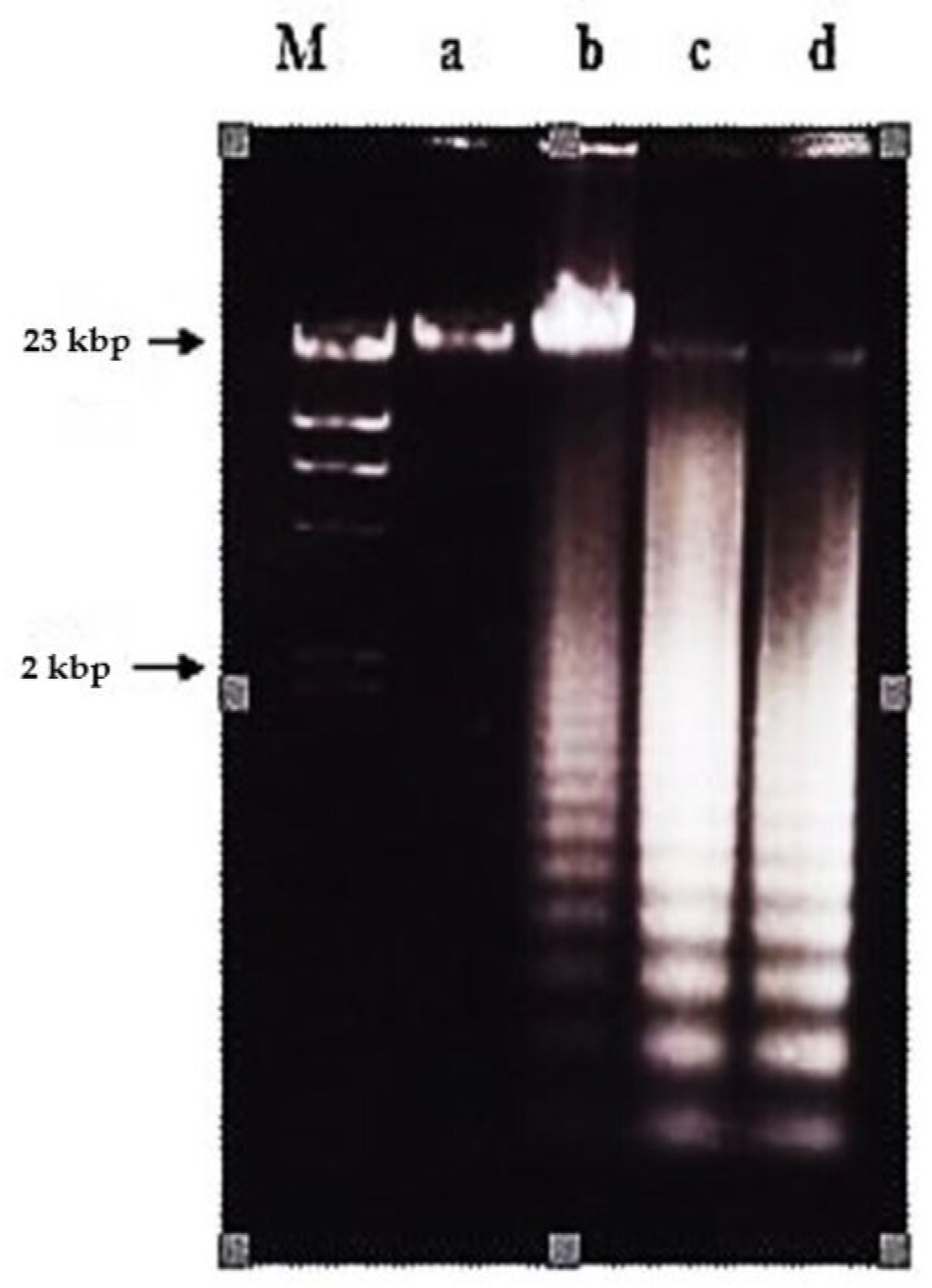
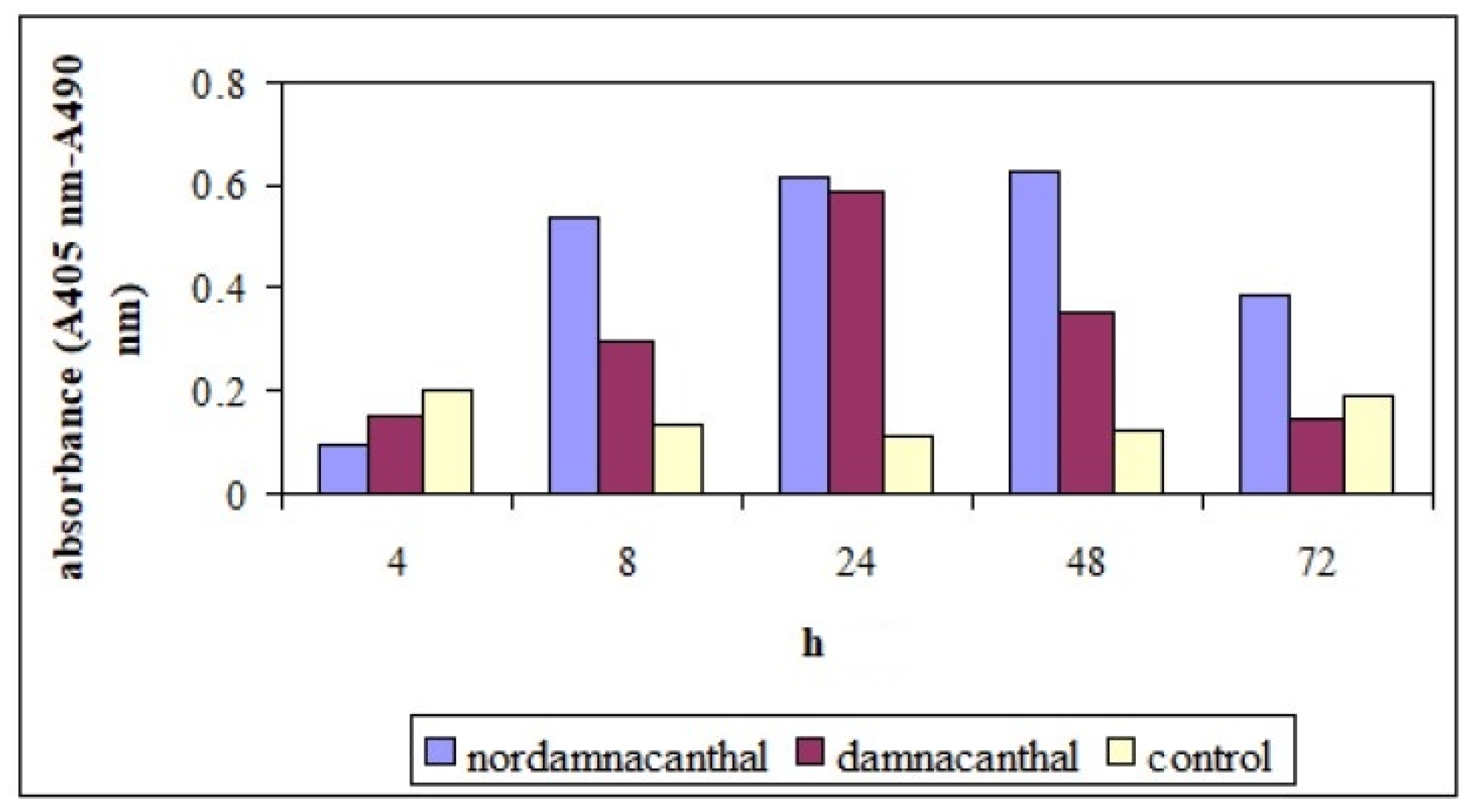
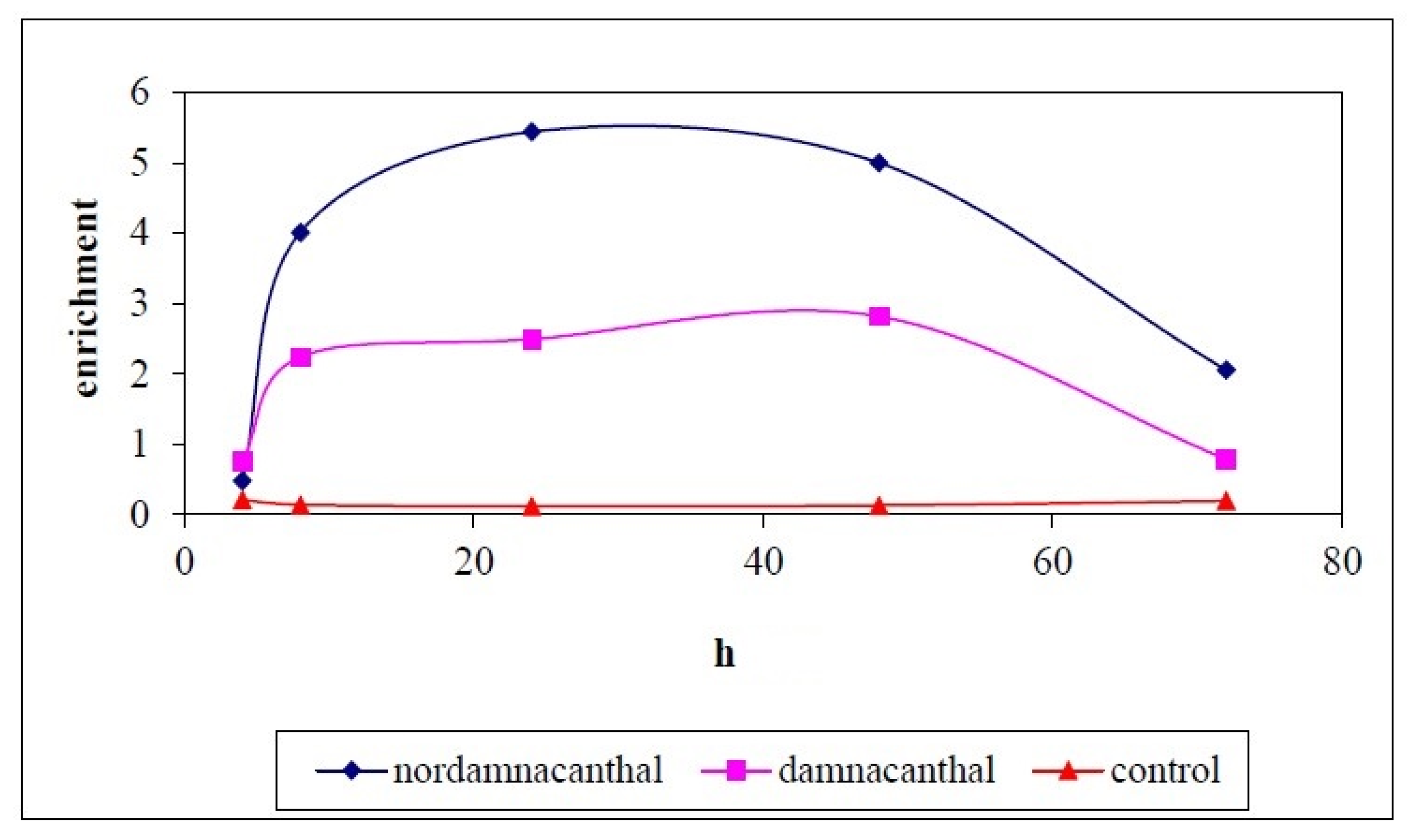
2.2. Effects of Nordamnacanthal and Damnacanthal on DNA Fragmentation in CEM-SS Cells
2.2.1. ELISSA
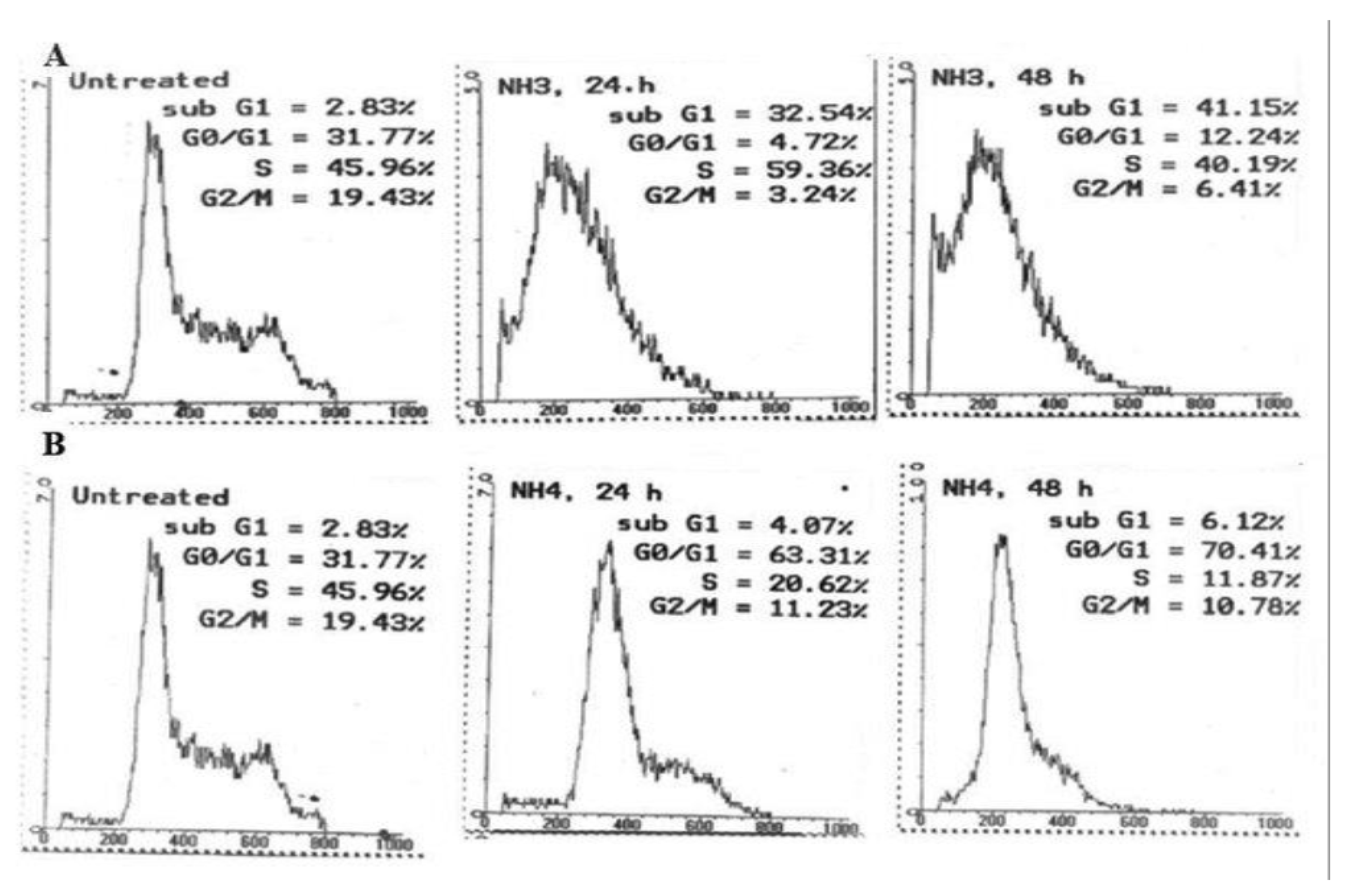
2.2.2. Cell Cycle Analysis
3. Discussion
4. Materials and Methods
4.1. Materials
4.1.1. Cells and Compounds
4.1.2. Compounds
4.2. Methods
4.2.1. Cell Lines
4.2.2. Cytotoxicity Assay
4.2.3. DNA Fragmentation Assay
4.2.4. Working Procedure for the ELISA
4.2.5. Flow Cytometric Analyses
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
Sample Availability
References
- GLOBOCAN. World. 2018. Available online: https://gco.iarc.fr/today/data/factsheets/populations/900-world-fact-sheets.pdf (accessed on 1 December 2020).
- Bray, F.; Ferlay, J.; Soerjomataram, I.; Siegel, R.L.; Torre, L.A.; Jemal, A. Global cancer statistics 2018: GLOBOCAN estimates of incidence and mortality worldwide for 36 cancers in 185 countries. CA Cancer J. Clin. 2018, 68, 394–424. [Google Scholar] [CrossRef]
- Greaves, M.A. Causal mechanism for childhood acute lymphoblastic leukaemia. Nat. Rev. Cancer 2018, 18, 471–484. [Google Scholar] [CrossRef]
- Samra, B.; Jabbour, E.; Ravandi, F.; Kantarjian, H.; Short, N.J. Evolving therapy of adult acute lymphoblastic leukemia: State-of-the-art treatment and future directions. J. Hematol. Oncol. 2020, 13, 70. [Google Scholar] [CrossRef] [PubMed]
- Alias, H.; Doris Lau, S.C.; Loh, C.K.; Harrison, C.J.; Eswaran, J. Improved Treatment of Childhood ALL in Malaysia. Blood 2019, 134, 5828. [Google Scholar] [CrossRef]
- Burkill, I.H. A Dictionary of the Economic Products of the Malay Peninsula; Ministry of Agriculture and Co-Operatives: Kuala Lumpur, Malaysia, 1966; Volume 1–2. [Google Scholar]
- Jasril, L.N.H.; Lajis, N.H.; Abdullah, M.A.; Sukari, M.A.; Ali, A.M. Antitumor promoting and antioxidant activities of anthraquinones isolated from cell suspension culture of Morinda elliptica. Asia Pac. J. Mol. Biol. Biotechnol. 2003, 11, 3–7. [Google Scholar]
- Ali, A.M.; Ismail, N.H.; Mackeen, M.M.; Yazan, L.S.; Mohamed, S.M.; Ho, A.S.H.; Lajis, N.H. Antiviral, cytotoxic and antimicrobial activites of anthraquinones isolated from the roots of Morinda elliptica. Pharm. Biol. 2000, 38, 298–301. [Google Scholar] [CrossRef]
- Akhtar, M.N.; Zareen, S.; Yeap, S.K.; Ho, W.Y.; Lo, K.M.; Hasan, A.; Alitheen, N.B. Total Synthesis, Cytotoxic Effects of Damnacanthal, Nordamnacanthal and Related Anthraquinone Analogues. Molecules 2013, 18, 10042–10055. [Google Scholar] [CrossRef]
- Subramani, T.; Yeap, S.K.; Ho, W.Y.; Ho, C.L.; Osman, C.P.; Ismail, N.H.; Rahman, N.M.; Alitheen, N.B. Nordamnacanthal potentiates the cytotoxic effects of tamoxifen in human breast cancer cells. Oncol. Lett. 2015, 9, 335–340. [Google Scholar] [CrossRef]
- Abu, N.; Zamberi, N.R.; Yeap, S.K.; Nordin, N.; Mohamad, N.E.; Romli, M.F.; Rasol, N.E.; Subramani, T.; Ismail, N.H.; Alitheen, N.B. Subchronic toxicity, immunoregulation and anti-breast tumor effect of Nordamnacantal, an anthraquinone extracted from the stems of Morinda citrifolia L. BMC Complement. Altern. Med. 2018, 18, 31. [Google Scholar] [CrossRef]
- Lv, L.; Chen, H.; Ho, C.T.; Sang, S. Chemical components of the roots of Noni (Morinda citrifolia) and their cytotoxic effects. Fitoterapia 2011, 82, 704–708. [Google Scholar] [CrossRef]
- Nualsanit, T.; Rojanapanthu, P.; Gritsanapan, W.; Lee, S.H.; Lawson, D.; Baek, S.J. Damnacanthal, a noni component, exhibits antitumorigenic activity in human colorectal cancer cells. J. Nutr. Biochem. 2012, 23, 915–923. [Google Scholar] [CrossRef]
- Shaghayegh, G.; Alabsi, A.M.; Ali-Saeed, R.; Ali, A.M.; Vincent-Chong, V.K.; Ismail, N.H.; Choon, Y.F.; Zain, R.B. Effects of Damnacanthal and Nordamnacanthal on Proliferation, Apoptosis, and Migration of Oral Squamous Cell Carcinoma Cells. Asian Pac. J. Cancer Prev. 2017, 18, 3333–3341. [Google Scholar] [PubMed]
- García-Vilas, J.A.; Quesadal, A.R.; Medina, M.A. Damnacanthal, a noni anthraquinone, inhibits c-Met and is a potent antitumor compound against Hep G2 human hepatocellular carcinoma cells. Sci. Rep. 2015, 5, 8021. [Google Scholar] [CrossRef]
- Ismail, N.; Mohamad, H.; Mohidin, A.; Lajis, N.H. Antioxidant activity of anthraquinones from Morinda elliptica. Nat. Prod. Sci. 2002, 8, 48–51. [Google Scholar]
- Shami, A.M.M. Antibacterial and antioxidant properties of anthraquinones fractions from Morinda citrifolia fruit. J. Rep. Pharm. Sci. 2018, 7, 379–388. [Google Scholar]
- Kamei, H.; Koide, T.; Kojima, T.; Hashimoto, Y.; Hasegawa, M. Inhibition of cell growth in culture by quinones. Cancer Biother. Radiopharm. 1998, 13, 185–188. [Google Scholar] [CrossRef] [PubMed]
- Kamil, M.; Haque, E.; Mir, S.S.; Irfan, S.; Hasan, A.; Sheikh, S.; Alam, A.; Ansari, K.M.; Nazir, A. Hydroxyl Group Difference between Anthraquinone Derivatives Regulate Different Cell Death Pathways via Nucleo-Cytoplasmic Shuttling of p53. Anticancer Agents Med. Chem. 2019, 19, 184–193. [Google Scholar] [CrossRef]
- Li, Y.; Jiang, J.G. Health functions and structure-activity relationships of natural anthraquinones from plants. Food Funct. 2018, 9, 6063–6080. [Google Scholar] [CrossRef]
- Sachet, M.; Liang, Y.Y.; Oehler, R. The immune response to secondary necrotic cells. Apoptosis 2017, 22, 1189–1204. [Google Scholar] [CrossRef] [PubMed]
- Saadat, R.Y.; Saeidi, N.; Zununi Vahed, S.; Barzegari, A.; Barar, J. An update to DNA ladder assay for apoptosis detection. Bioimpacts 2015, 5, 25–28. [Google Scholar]
- Shaghayegh, G.; Alabsi, A.M.; Ali-Saeed, R.; Ali, A.M.; Vincent-Chong, V.K.; Zain, R.B. Cell cycle arrest and mechanism of apoptosis induction in H400 oral cancer cells in response to Damnacanthal and Nordamnacanthal isolated from Morinda citrifolia. Cytotechnology 2016, 68, 1999–2013. [Google Scholar] [CrossRef] [PubMed]
- Walker, P.R.; Kokileva, L.; LeBlanc, J.; Sikorska, M. Detection of the intial stages of DNA fragmentation in apoptosis. Biotechniques 1993, 15, 1032–1040. [Google Scholar]
- Hao, L.; Zhao, Y.; Li, Z.; He, H.; Liang, Q.; Zhang, Z.; Shi, Z.; Zhang, P.; Han, C. Tumor necrosis factor-related apoptosis-inducing ligand inhibits proliferation and induces apoptosis of prostate and bladder cancer cells. Oncol. Lett. 2017, 13, 3638–3640. [Google Scholar] [CrossRef][Green Version]
- Xiao, Q.; Lu, Y.; Chen, X. Oleanolic Acid Induces Apoptosis and Necrosis in LO2 Cells. Indian J. Pharm. Sci. 2020, 82, 361–368. [Google Scholar] [CrossRef]
- Duvall, E.; Wyllie, A.H. Death and the cell. Immunol. Today 1986, 7, 115–119. [Google Scholar] [CrossRef]
- Kumar, V.; Chichili, V.P.; Tang, X.; Sivaraman, J. A novel trans conformation of ligand-free calmodulin. PLoS ONE 2013, 8, e54834. [Google Scholar] [CrossRef]
- Waring, P.; Kos, F.J.; Mullbacher, A. Apoptosis or programmed cell death. Med. Res. Rev. 1991, 11, 219–236. [Google Scholar] [CrossRef] [PubMed]
- Marini, M.; Musiani, D. Micromolar Zinc Affects Endonucleolytic Activity in Hydrogen Peroxide-Mediated Apoptosis. Exp. Cell Res. 1998, 239, 393–398. [Google Scholar] [CrossRef] [PubMed]
- Gammoh, N.Z.; Rink, L. Zinc in infection and inflammation. Nutrients 2017, 9, 624. [Google Scholar] [CrossRef] [PubMed]
- Harmon, B.V.; Forster, T.H.; Collins, R.J. Hyperthermia-Induced apoptosis. In Programmed Cell Death: The Cellular and Molecular Biology of Apoptosis; Lavin, M., Watters, D., Eds.; Harwood Academic Publishers: Victoria, BC, Canada, 1993; pp. 297–314. [Google Scholar]
- Bicknell, G.R.; Snowden, R.T.; Cohen, G.M. Formation of high molecular mass DNA fragments is a marker of apoptosis in the human leukaemic cell line, U397. J. Cell Sci. 1994, 107, 2483–2489. [Google Scholar]
- Eron, S.J.; MacPherson, D.J.; Dagbay, K.B.; Hardy, J.A. Multiple Mechanisms of Zinc-Mediated Inhibition for the Apoptotic Caspases-3, -6, -7, and -8. ACS Chem. Biol. 2018, 13, 1279–1290. [Google Scholar] [CrossRef]
- Radford, R.J.; Chyan, W.; Lippard, S.J. Peptide-Based Targeting of Fluorescent Zinc Sensors to the Plasma Membrane of Live Cells. Chem. Sci. 2013, 4, 3080–3084. [Google Scholar] [CrossRef]
- Haneji, T.; Hirashima, K.; Teramachi, J.; Morimoto, H. Okadaic acid activates the PKR pathway and induces apoptosis through PKR stimulation in MG63 osteoblast-like cells. Int. J. Oncol. 2013, 42, 1904–1910. [Google Scholar] [CrossRef] [PubMed]
- Takai, A.; Eto, M.; Hirano, K.; Takeya, K.; Wakimoto, T.; Watanabe, M. Protein phosphatases 1 and 2A and their naturally occurring inhibitors: Current topics in smooth muscle physiology and chemical biology. J. Physiol. Sci. 2018, 68, 1–17. [Google Scholar] [CrossRef] [PubMed]
- Dedinszki, D.; Kiss, A.; Márkász, L.; Márton, A.; Tóth, E.; Székely, L.; Erdődi, F. Inhibition of protein phosphatase-1 and -2A decreases the chemosensitivity of leukemic cells to chemotherapeutic drugs. Cell. Signal. 2015, 27, 363–372. [Google Scholar] [CrossRef]
- Janeczko, M.; Masłyk, M.; Kubiński, K.; Golczyk, H. Emodin, a natural inhibitor of protein kinase CK2, suppresses growth, hyphal development, and biofilm formation of Candida albicans. Yeast 2017, 34, 253–265. [Google Scholar] [CrossRef] [PubMed]
- Nishizuka, Y. The molecular heterogenity of protein kinase C and its implications for cellular regulation. Nature 1988, 334, 661–665. [Google Scholar] [CrossRef]
- Song, Q.; Baxter, G.D.; Kovacs, E.M.; Findik, D.; Lavin, M.F. Inhibition of apoptosis in human tumour cells by okadaic acid. J. Cell. Physiol. 1992, 153, 550–556. [Google Scholar] [CrossRef]
- Goodall, K.J.; Finch-Edmondson, M.L.; van Vuuren, J.; Yeoh, G.C.; Gentle, I.E.; Vince, J.E.; Ekert, P.G.; Vaux, D.L.; Callus, B.A. Cycloheximide Can Induce Bax/Bak Dependent Myeloid Cell Death Independently of Multiple BH3-Only Proteins. PLoS ONE 2016, 11, e0164003. [Google Scholar] [CrossRef]
- Sánchez-Alcázar, J.A.; Khodjakov, A.; Schneider, E. Anticancer Drugs Induce Increased Mitochondrial Cytochrome C Expression That Precedes Cell Death. Cancer Res. 2001, 61, 1038–1044. [Google Scholar]
- Hoshino, M.; Qi, M.L.; Yoshimura, N.; Miyashita, T.; Tagawa, K.; Wada, Y.; Enokido, Y.; Marubuchi, S.; Harjes, P.; Arai, N.; et al. Transcriptional repression induces a slowly progressive atypical neuronal death associated with changes of YAP isoforms and p73. J. Cell Biol. 2006, 172, 589–604. [Google Scholar] [CrossRef]
- Chow, S.C.; Peters, I.; Orrenius, S. Reevaluation of the role of de novo protein synthesis in rat thymocyte apoptosis. Exp. Cell Res. 1995, 216, 149–159. [Google Scholar] [CrossRef]
- Bensaude, O. Inhibiting eukaryotic transcription. Which compound to choose? How to evaluate its activity? Transcription 2011, 2, 103–108. [Google Scholar] [CrossRef]
- Choi, J.W.; Jung, S.E. Lovastatin-Induced proliferation inhibition and apoptosis in C6 glial cells. J. Pharmacol. Exp. Ther. 1999, 289, 572–579. [Google Scholar]
- Cohen, J.J.; Duke, R.C.; Fadok, V.A.; Sellins, K.S. Apoptosis and programmed cell death in immunity. Annu. Rev. Immunol. 1992, 10, 267–293. [Google Scholar] [CrossRef]
- Rizzuto, R.; Pinton, P.; Ferrari, D.; Chami, M.; Szabadkai, G.; Magalhães, P.J.; Virgilio, F.D.; Pozzan, T. Calcium and apoptosis: Facts and hypotheses. Oncogene 2003, 22, 8619–8627. [Google Scholar] [CrossRef] [PubMed]
- Aoki, K.; Parent, A.; Zhang, J. Mechanism of damnacanthal-induced [Ca2+]i elevation in human dermal fibroblasts. Eur. J. Pharmacol. 2000, 387, 119–124. [Google Scholar] [CrossRef]
- La Rovere, R.M.L.; Roest, G.; Bultynck, G.; Parys, J.B. Intracellular Ca2+ signaling and Ca2+ microdomains in the control of cell survival, apoptosis and autophagy. Cell Calcium 2016, 60, 74–87. [Google Scholar] [CrossRef] [PubMed]
- Gerschenson, L.E.; Rotello, R.J. Apoptosis: A different type of cell death. FASEB J. 1992, 6, 2450–2455. [Google Scholar] [CrossRef] [PubMed]
- Dibwe, D.F.; Awale, S.; Kadota, S.; Tezuka, Y. Damnacanthal from the Congolese medicinal plant Garcinia huillensis has a potent preferential cytotoxicity against human pancreatic cancer PANC-1 cells. Phytother. Res. 2012, 26, 1920–1926. [Google Scholar] [CrossRef] [PubMed]
- Ohashi, K.; Sampei, K.; Nakagawa, M.; Uchiumi, N.; Amanuma, T.; Aiba, S.; Oikawa, M.; Mizuno, K. Damnacanthal, an effective inhibitor of LIM-kinase, inhibits cell migration and invasion. Mol. Biol. Cell 2014, 25, 828–840. [Google Scholar] [CrossRef]
- García-Vilas, J.A.; Pino-Ángeles, A.; Martínez-Poveda, B.; Quesada, A.R.; Medina, M.Á. The noni anthraquinone damnacanthal is a multi-kinase inhibitor with potent anti-angiogenic effects. Cancer Lett. 2017, 385, 1–11. [Google Scholar] [CrossRef]
- Jim, H.S.; Jacobsen, P.B.; Phillips, K.M.; Wenham, R.M.; Roberts, W.; Small, B.J. Lagged relationships among sleep disturbance, fatigue, and depressed mood during chemotherapy. Health Psychol. 2013, 32, 768–774. [Google Scholar] [CrossRef]
- Ansari, L.; Shiehzadeh, F.; Taherzadeh, Z.; Nikoofal-Sahlabadi, S.; Momtazi-borojeni, A.A.; Sahebkar, A.; Eslami, S. The most prevalent side effects of pegylated liposomal doxorubicin monotherapy in women with metastatic breast cancer: A systematic review of clinical trials. Cancer Gene Ther. 2017, 24, 189–193. [Google Scholar] [CrossRef] [PubMed]
- Dar, W.; Hussain, M.; Aziz, S.A.; Mohammad, G.; Wani, B.; Latief, M. Uncommon Adverse Effects of Commonly Used Chemotherapeutic Agents in Medical Oncology Practice: A Series of Two Cases of Hand-Foot Syndrome. Indian J. Med. Paediatr. Oncol. 2017, 38, 380–382. [Google Scholar] [CrossRef] [PubMed]
- Jameel, P.Z.; Lohiya, S.; Dongre, A.; Damke, S.; Lakhkar, B.B. Concurrent diabetic ketoacidosis and pancreatitis in Paediatric acute lymphoblastic leukemia receiving L-asparaginase. BMC Pediatr. 2020, 20, 228. [Google Scholar] [CrossRef]
- Qi, L.; Luo, Q.; Zhang, Y.; Jia, F.; Zhao, Y.; Wang, F. Advances in Toxicological Research of the Anticancer Drug Cisplatin. Chem. Res. Toxicol. 2019, 32, 1469–1486. [Google Scholar] [CrossRef] [PubMed]
- Rios, A.; Cen, P.; Dinh, B.; Mays, S.R.; Patel, A.B. Dramatic response of nivolumab-associated psoriasiform dermatitis to etoposide. Eur. J. Cancer 2019, 107, 97–99. [Google Scholar] [CrossRef] [PubMed]
- Ismail, N.H.; Ali, A.M.; Aimi, N.; Kitajima, M.; Takayama, H.; Lajis, N.H. Anthraquinones from Morinda elliptica. Phytochemistry 1997, 45, 1723–1725. [Google Scholar] [CrossRef]
- Hirose, Y. Syntheses of Damnacanthal, Damnacanthol, Norjuzunal and Norjuzunol, the Coloring Matters of Damnacanthus spp. Chem. Pharm. Bull. 1960, 8, 417–426. [Google Scholar] [CrossRef]
- Adnan, N.E. Isolation and photophysical properties of Di- and Tri-substituted natural anthraquinones from Malaysian Morinda citrifolia. Sains Malays. 2018, 47, 903–906. [Google Scholar] [CrossRef]
- Mossman, T. Rapid colorimetric assay for cellular growth and survival: Application to proliferation and cytotoxicity assays. J. Immunol. Methods 1983, 65, 55–63. [Google Scholar] [CrossRef]

















{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Time after Treatment | |||
|---|---|---|---|
| Control (0 h) | 24 h | 48 h | |
| Nordamnacanthal | |||
| Sub-G1 (%) | 2.83 | 32.54 | 41.15 |
| G0/G1 (%) | 31.77 | 4.72 | 12.24 |
| S (%) | 45.96 | 59.36 | 40.19 |
| G2/M (%) | 19.43 | 3.24 | 6.41 |
| Damnacanthal | |||
| Sub-G1 (%) | 2.83 | 4.07 | 6.12 |
| G0/G1 (%) | 31.77 | 63.31 | 70.41 |
| S (%) | 45.96 | 20.62 | 11.87 |
| G2/M (%) | 19.43 | 11.23 | 10.78 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Latifah, S.Y.; Gopalsamy, B.; Abdul Rahim, R.; Manaf Ali, A.; Haji Lajis, N. Anticancer Potential of Damnacanthal and Nordamnacanthal from Morinda elliptica Roots on T-lymphoblastic Leukemia Cells. Molecules 2021, 26, 1554. https://doi.org/10.3390/molecules26061554
Latifah SY, Gopalsamy B, Abdul Rahim R, Manaf Ali A, Haji Lajis N. Anticancer Potential of Damnacanthal and Nordamnacanthal from Morinda elliptica Roots on T-lymphoblastic Leukemia Cells. Molecules. 2021; 26(6):1554. https://doi.org/10.3390/molecules26061554
Chicago/Turabian StyleLatifah, Saiful Yazan, Banulata Gopalsamy, Raha Abdul Rahim, Abdul Manaf Ali, and Nordin Haji Lajis. 2021. "Anticancer Potential of Damnacanthal and Nordamnacanthal from Morinda elliptica Roots on T-lymphoblastic Leukemia Cells" Molecules 26, no. 6: 1554. https://doi.org/10.3390/molecules26061554
APA StyleLatifah, S. Y., Gopalsamy, B., Abdul Rahim, R., Manaf Ali, A., & Haji Lajis, N. (2021). Anticancer Potential of Damnacanthal and Nordamnacanthal from Morinda elliptica Roots on T-lymphoblastic Leukemia Cells. Molecules, 26(6), 1554. https://doi.org/10.3390/molecules26061554