
Probing the Role of the Conserved Arg174 in Formate Dehydrogenase by Chemical Modification and Site-Directed Mutagenesis

 ,
, 
Abstract
1. Introduction
2. Results and Discussion
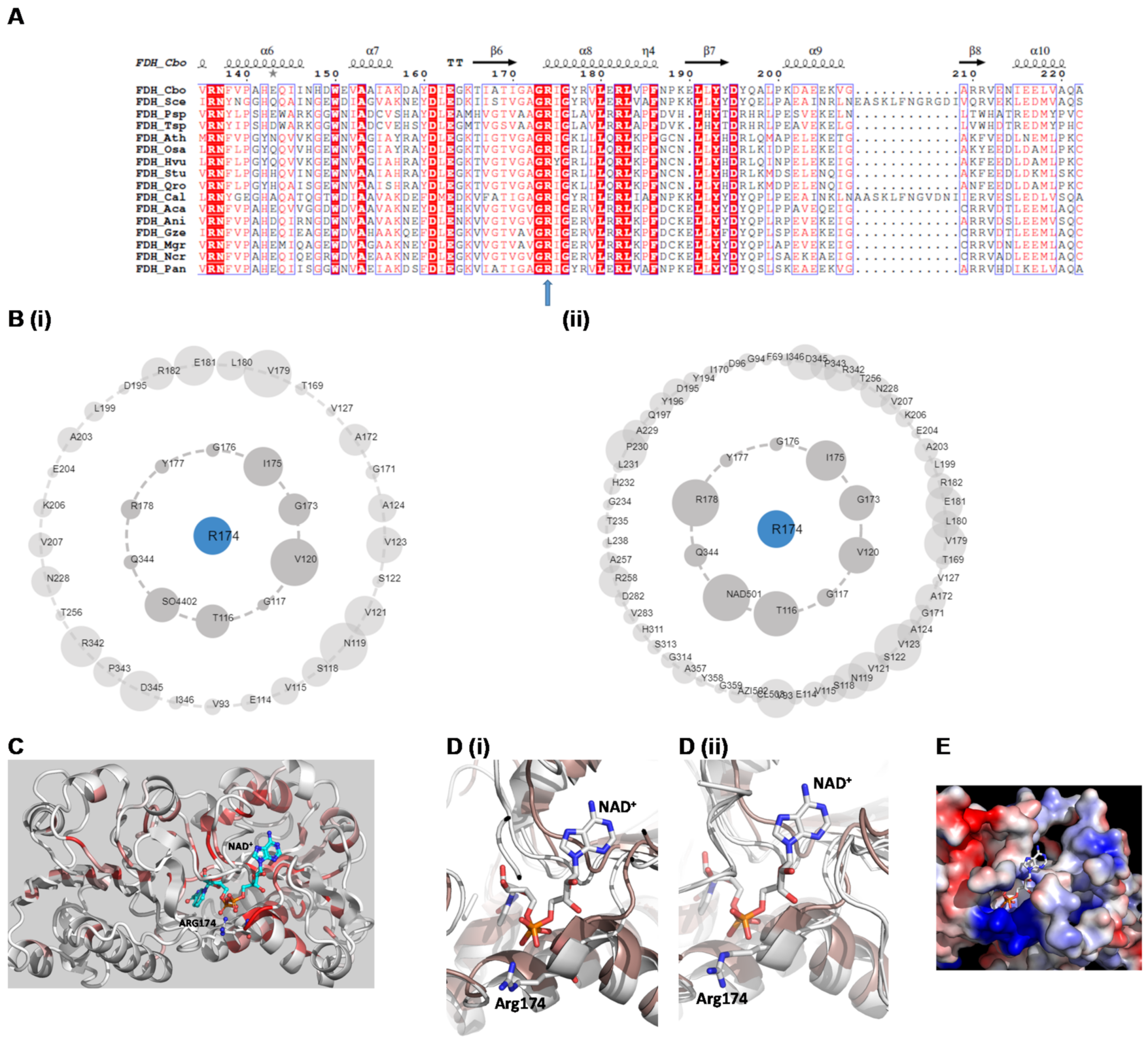
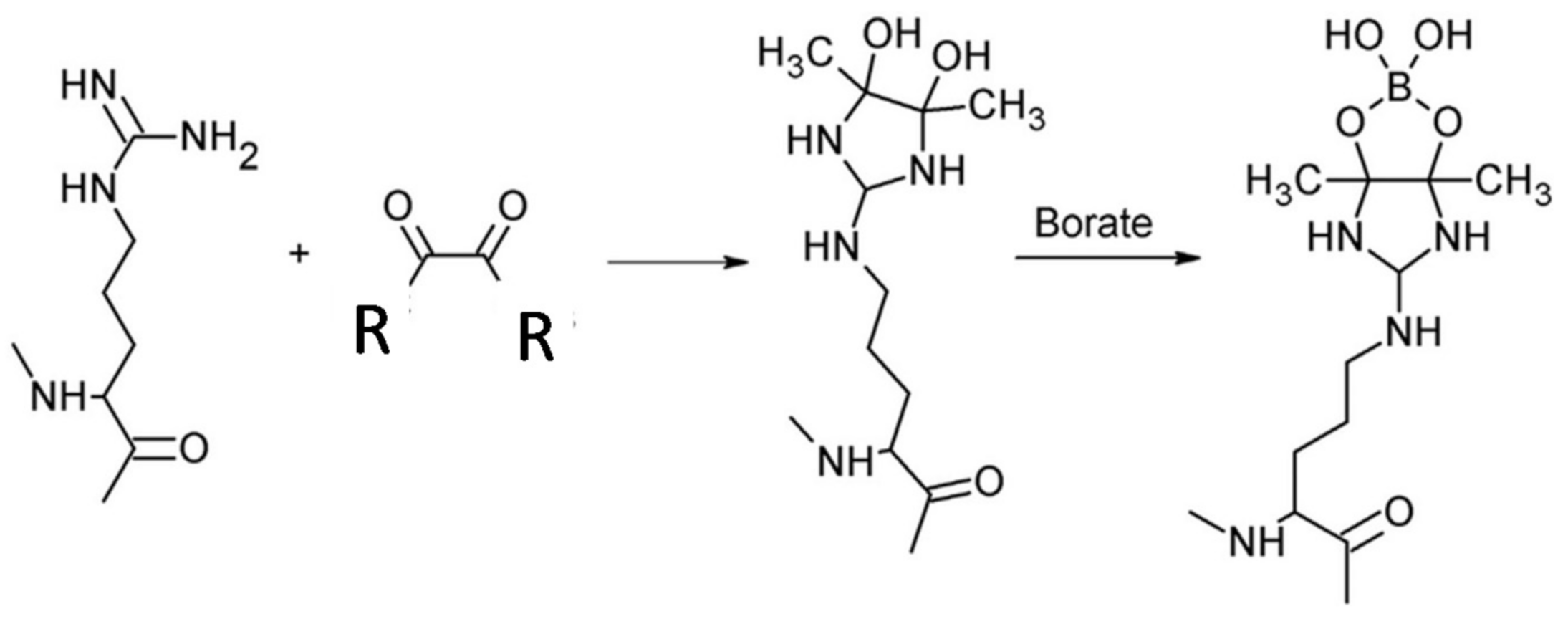
2.1. Structural Context
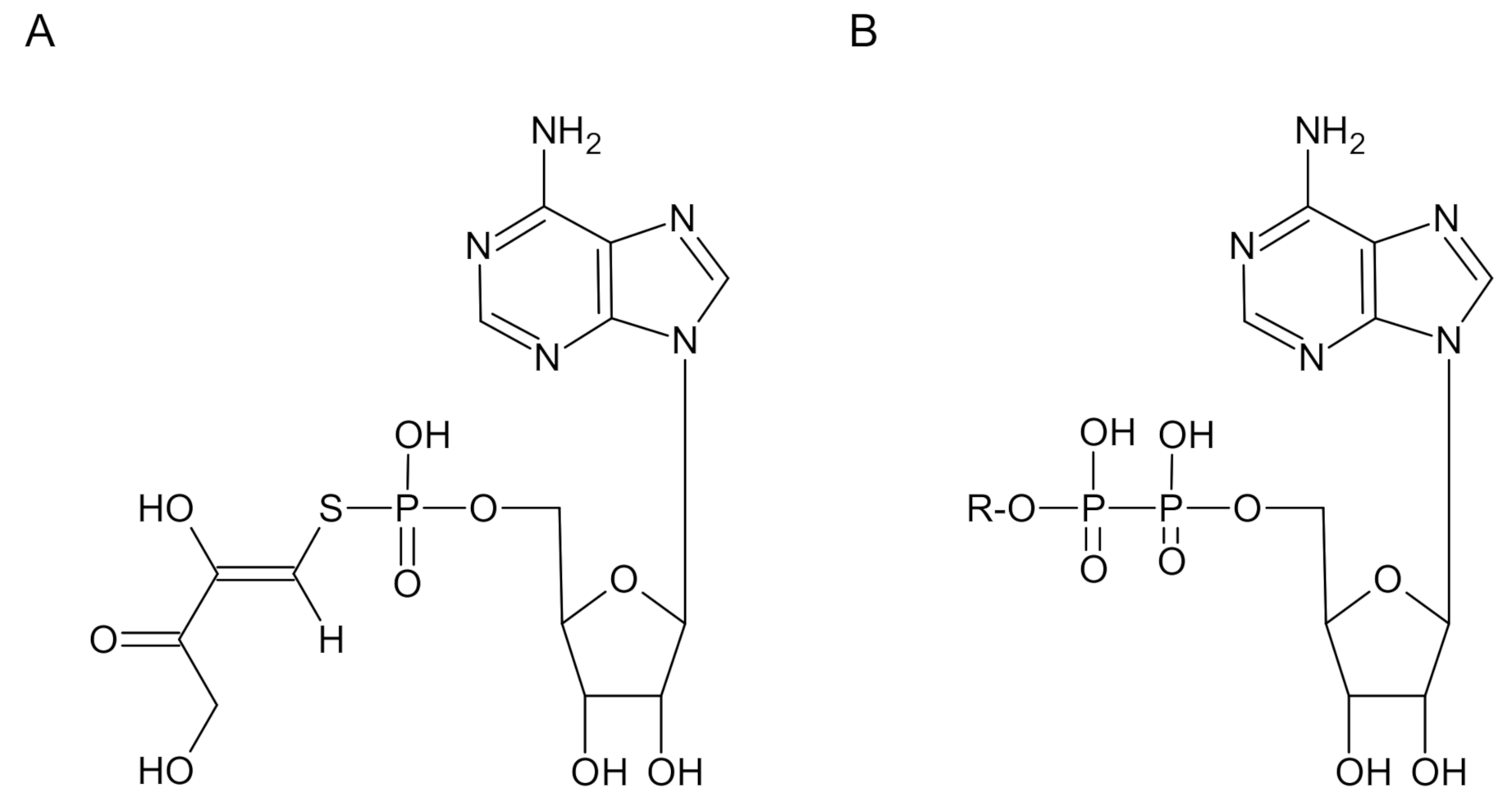
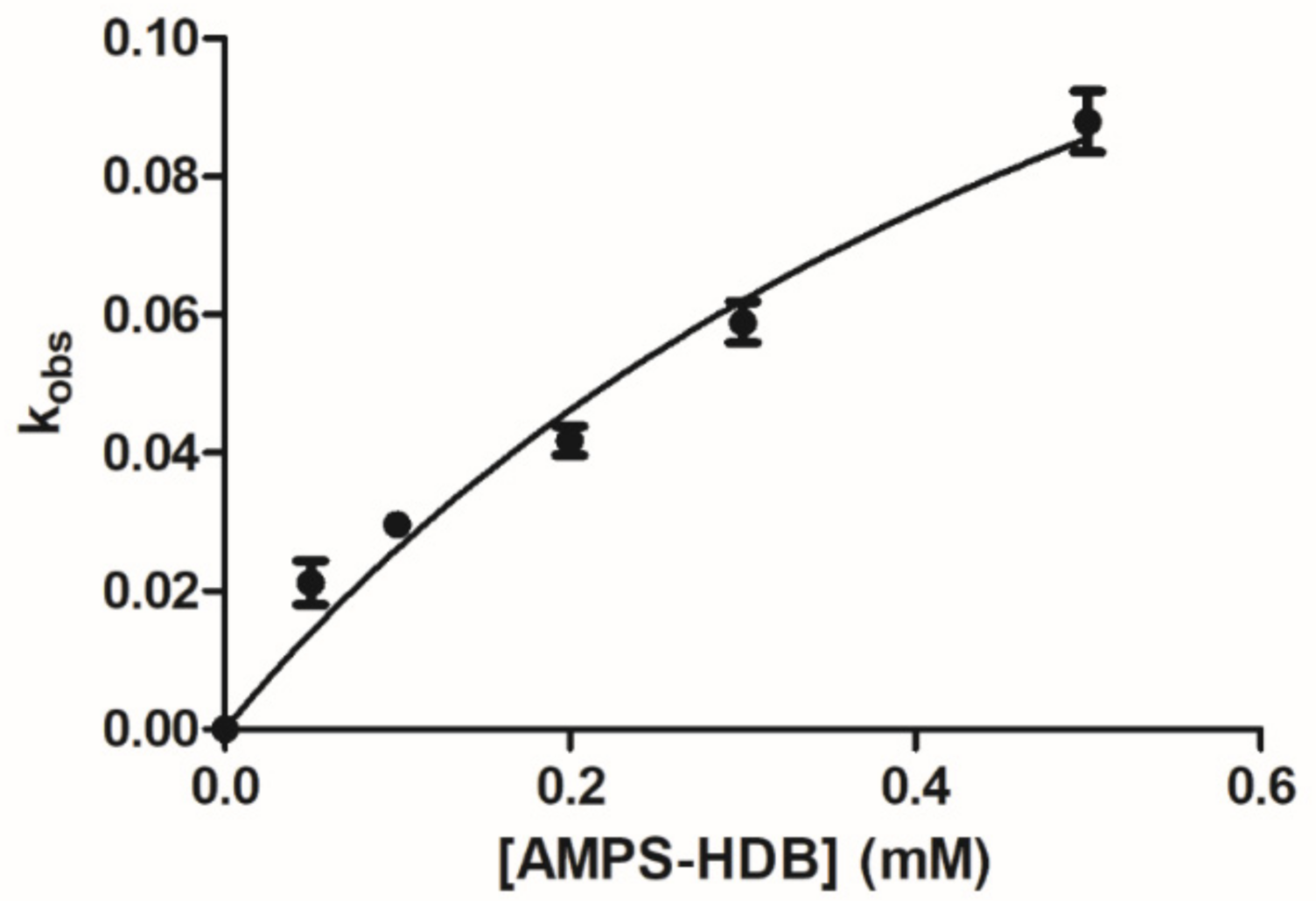
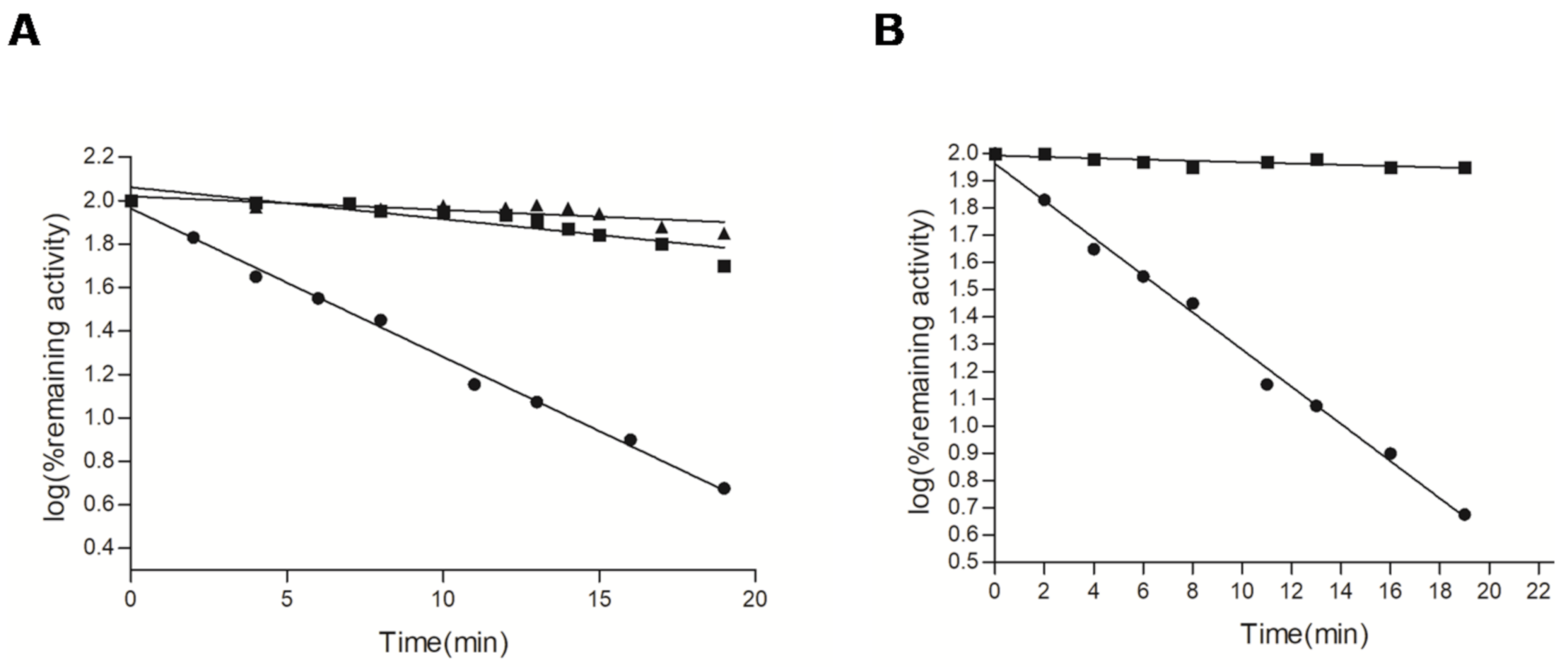
2.2. Kinetics of Reaction of AMPS-HDB with CbFDH
2.3. Effect of Substrates on the Reaction of AMPS-HDB with CbFDH
2.4. Site-Directed Mutagenesis of Arg174 Mutant Enzyme
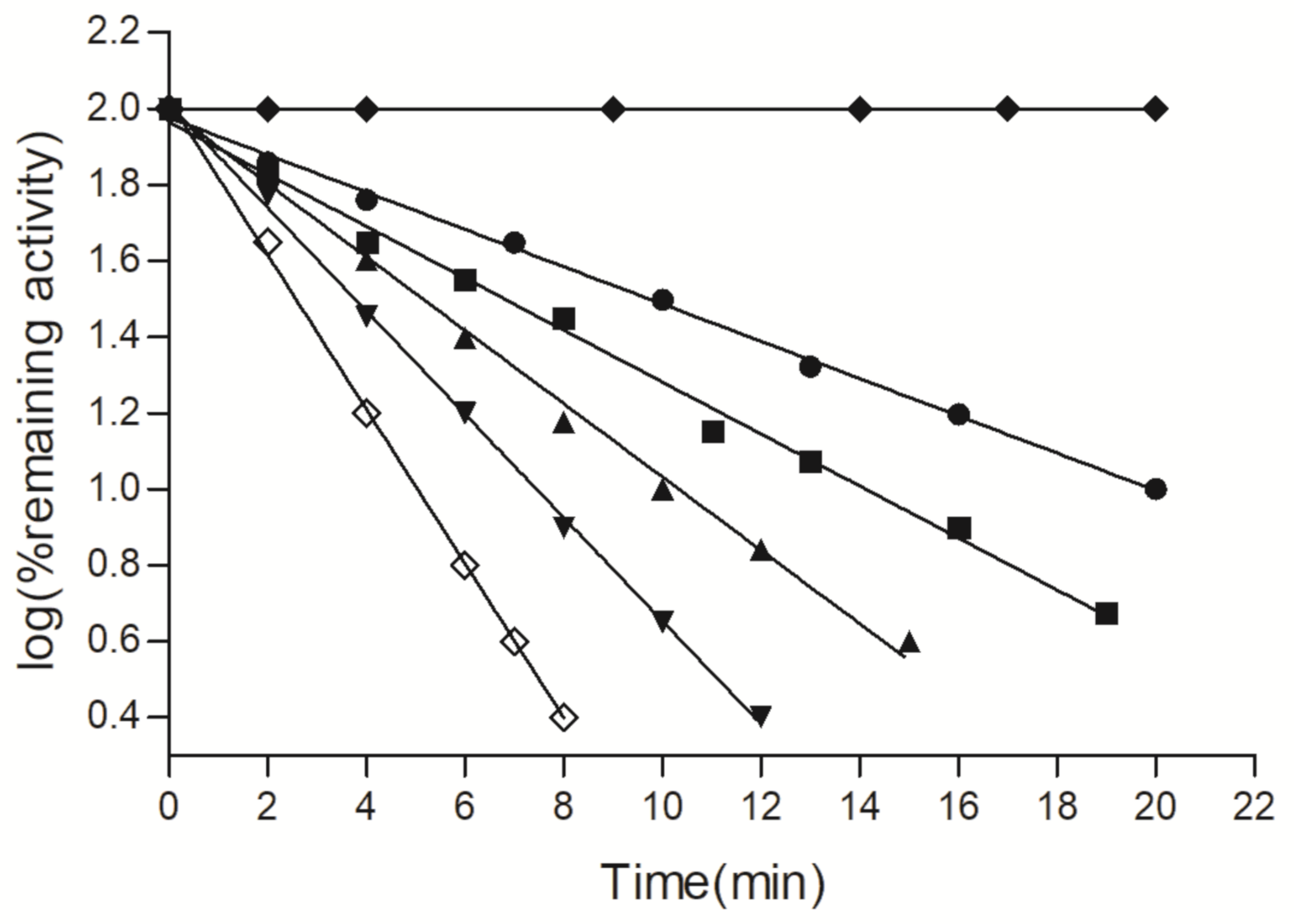
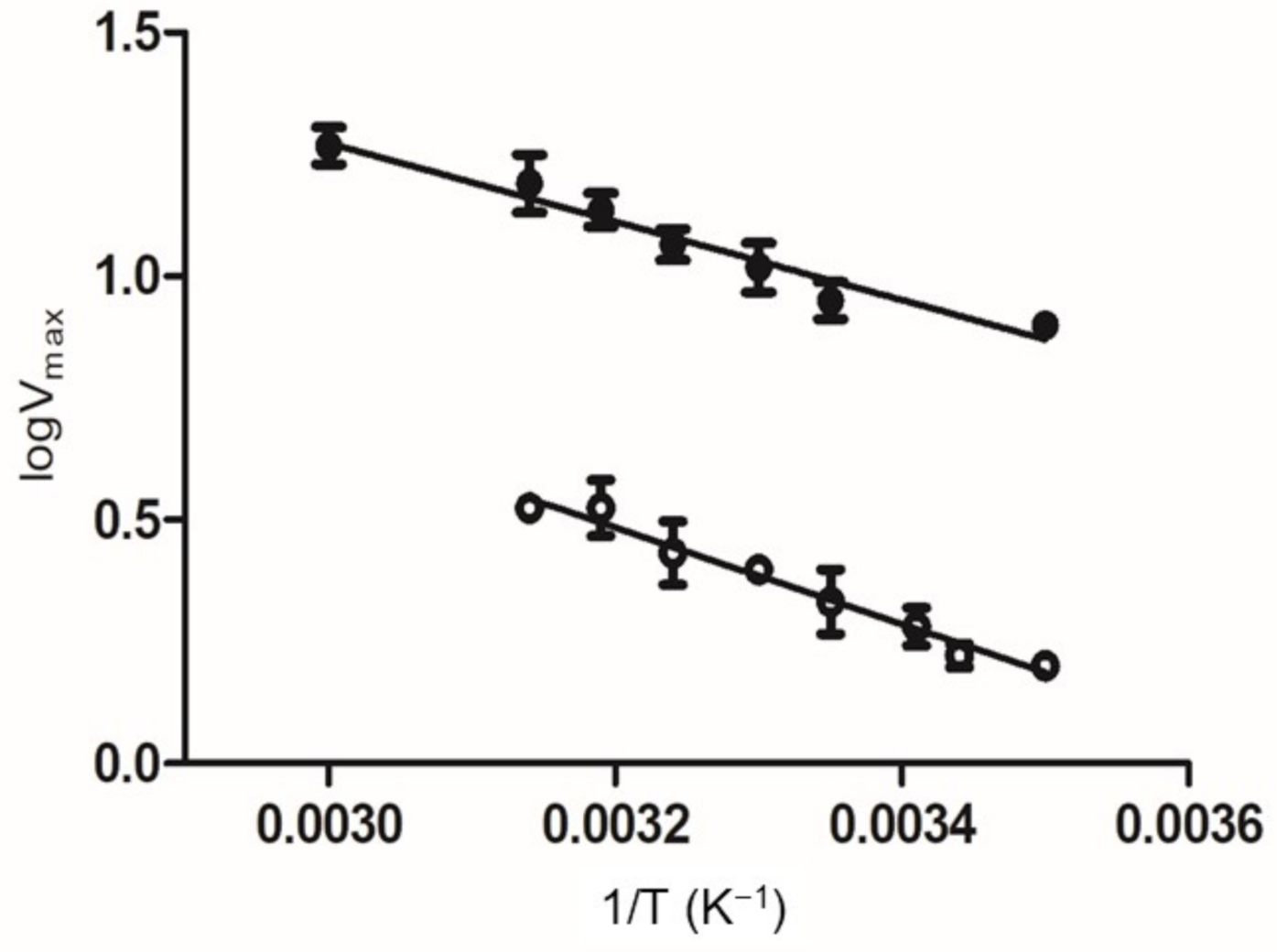
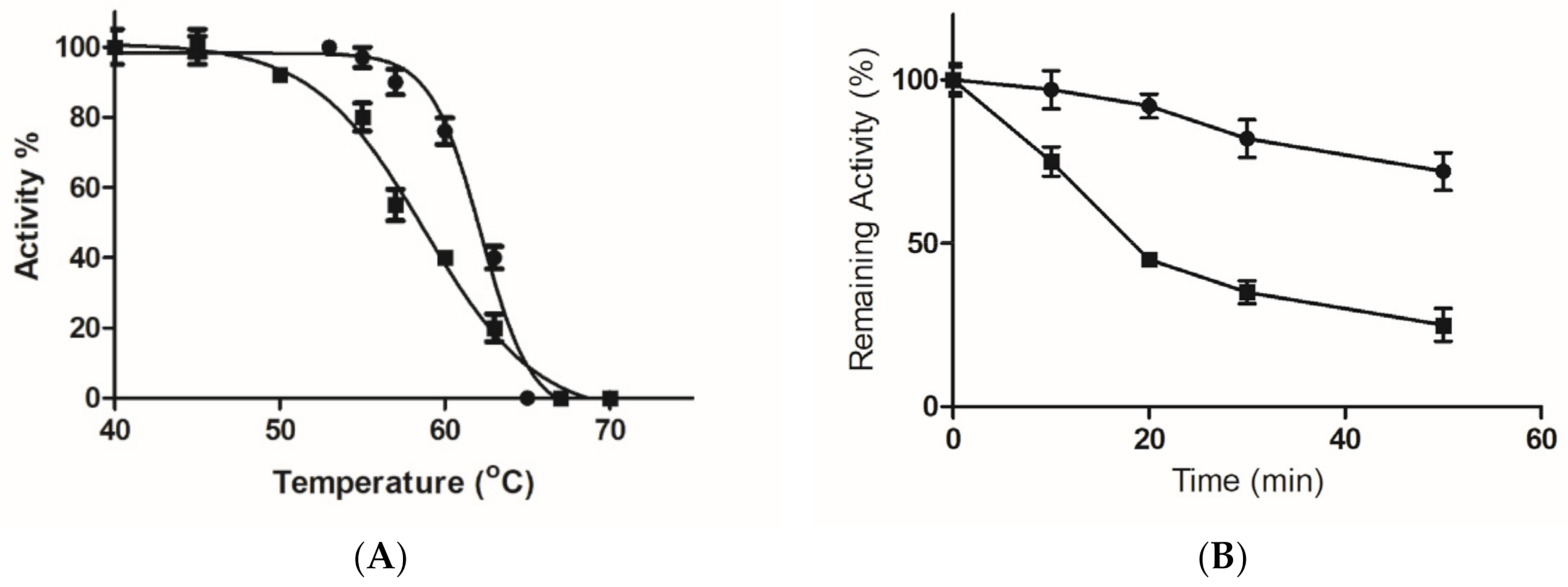
2.5. Kinetics and Stability Analysis of the Wild-Type and Mutant Arg174Asn Enzymes
3. Materials and Methods
3.1. Materials
3.2. Methods
3.2.1. CbFDH Assays
3.2.2. Determination of Protein Concentration
3.2.3. Expression and Purification of CbFDH
3.2.4. Enzyme Inactivation and Inhibition Studies
3.2.5. Site-Directed Mutagenesis
3.2.6. Kinetic Analysis of Wild-Type and Mutant Enzymes
3.2.7. Effect of Temperature on Enzyme Activity
3.2.8. Thermal Stability of the Wild-Type and Mutant Enzyme
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Sample Availability: Samples of the compounds are not available from the authors. |
References
- Popov, V.O.; Lamzin, V.S. NAD+-dependent formate dehydrogenase. Biochem. J. 1994, 301, 625–643. [Google Scholar] [CrossRef]
- Bulut, H.; Valjakka, J.; Yuksel, B.; Yilmazer, B.; Turunen, O.; Binay, B. Effect of Metal Ions on the Activity of Ten NAD-Dependent Formate Dehydrogenases. Protein J. 2020, 39, 519–530. [Google Scholar] [CrossRef]
- Nielsen, C.F.; Lange, L.; Meyer, A.S. Classification and enzyme kinetics of formate dehydrogenases for biomanufacturing via CO2 utilization. Biotechnol. Adv. 2019, 37, 107408. [Google Scholar] [CrossRef]
- Jormakka, M.; Byrne, B.; Iwata, S. Formate dehydrogenase—A versatile enzyme in changing environments. Curr. Opin. Struct. Biol. 2003, 13, 418–423. [Google Scholar] [CrossRef]
- Andreadeli, A.; Platis, D.; Tishkov, V.; Popov, V.; Labrou, N.E. Structure-guided alteration of coenzyme specificity of formate dehydrogenase by saturation mutagenesis to enable efficient utilization of NADP+. FEBS J. 2008, 275, 3859–3869. [Google Scholar] [CrossRef]
- Jiang, H.-W.; Chen, Q.; Pan, J.; Zheng, G.-W.; Xu, J.-H. Rational Engineering of Formate Dehydrogenase Substrate/Cofactor Affinity for Better Performance in NADPH Regeneration. Appl. Biochem. Biotechnol. 2020, 192, 530–543. [Google Scholar] [CrossRef] [PubMed]
- Guo, X.; Wang, X.; Liu, Y.; Li, Q.; Wang, J.; Liu, W.; Zhao, Z.K. Structure-Guided Design of Formate Dehydrogenase for Regeneration of a Non-Natural Redox Cofactor. Chem.-A Eur. J. 2020, 26, 16611–16615. [Google Scholar] [CrossRef] [PubMed]
- Luo, W.; Zhu, J.; Zhao, Y.; Zhang, H.; Yang, X.; Liu, Y.; Rao, Z.; Yu, X. Cloning and Expression of a Novel Leucine Dehydrogenase: Characterization and L-tert-Leucine Production. Front. Bioeng. Biotechnol. 2020, 8, 186–190. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.; Lindner, S.N.; Aslan, S.; Yishai, O.; Wenk, S.; Schann, K.; Bar-Even, A. Growth of E. coli on formate and methanol via the reductive glycine pathway. Nat. Chem. Biol. 2020, 16, 538–545. [Google Scholar] [CrossRef]
- Guo, X.; Liu, Y.; Wang, Q.; Wang, X.; Li, Q.; Liu, W.; Zhao, Z.K. Non-natural Cofactor and Formate-Driven Reductive Carboxylation of Pyruvate. Angew. Chem. Int. Ed. 2019, 59, 3143–3146. [Google Scholar] [CrossRef] [PubMed]
- Agomuo, E.N.; Amadi, P.U. Biochemical Implications of Biotransformation of Some Toxic Floras Using Natural Local Enzyme Sources. Recent Patents Biotechnol. 2020, 14, 16–32. [Google Scholar] [CrossRef]
- Miyaji, A.; Amao, Y. How does methylviologen cation radical supply two electrons to the formate dehydrogenase in the catalytic reduction process of CO2 to formate? Phys. Chem. Chem. Phys. 2020, 22, 18595–18605. [Google Scholar] [CrossRef]
- Tian, Y.; Zhou, Y.; Zong, Y.; Li, J.; Yang, N.; Zhang, M.; Guo, Z.; Song, H. Construction of Functionally Compartmental Inorganic Photocatalyst–Enzyme System via Imitating Chloroplast for Efficient Photoreduction of CO2 to Formic Acid. ACS Appl. Mater. Interfaces 2020, 12, 34795–34805. [Google Scholar] [CrossRef]
- Calzadiaz-Ramirez, L.; Calvó-Tusell, C.; Stoffel, G.M.M.; Lindner, S.N.; Osuna, S.; Erb, T.J.; Garcia-Borràs, M.; Bar-Even, A.; Acevedo-Rocha, C.G. In Vivo Selection for Formate Dehydrogenases with High Efficiency and Specificity toward NADP+. ACS Catal. 2020, 10, 7512–7525. [Google Scholar] [CrossRef]
- Ren, S.; Wang, Z.; Bilal, M.; Feng, Y.; Jiang, Y.; Jia, S.; Cui, J. Co-immobilization multienzyme nanoreactor with co-factor regeneration for conversion of CO2. Int. J. Biol. Macromol. 2020, 155, 110–118. [Google Scholar] [CrossRef]
- Çakar, M.M.; Ruupunen, J.; Mangas-Sanchez, J.; Birmingham, W.R.; Yildirim, D.; Turunen, O.; Turner, N.J.; Valjakka, J.; Binay, B. Engineered formate dehydrogenase from Chaetomium thermophilum, a promising enzymatic solution for biotechnical CO2 fixation. Biotechnol. Lett. 2020, 42, 2251–2262. [Google Scholar] [CrossRef]
- Satanowski, A.; Bar-Even, A. A one-carbon path for fixing CO2: C1 compounds, produced by chemical catalysis and upgraded via microbial fermentation, could become key intermediates in the valorization of CO2 into commodity chemicals. EMBO Rep. 2020, 21, e50273. [Google Scholar] [CrossRef] [PubMed]
- Yadav, R.K.; Baeg, J.-O.; Oh, G.H.; Park, N.-J.; Kong, K.-J.; Kim, J.; Hwang, D.W.; Biswas, S.K. A Photocatalyst–Enzyme Coupled Artificial Photosynthesis System for Solar Energy in Production of Formic Acid from CO2. J. Am. Chem. Soc. 2012, 134, 11455–11461. [Google Scholar] [CrossRef]
- Artiukhov, A.V.; Pometun, A.A.; Zubanova, S.A.; Tishkov, V.I.; Bunik, V.I. Advantages of formate dehydrogenase reaction for efficient NAD+ quantification in biological samples. Anal. Biochem. 2020, 603, 113797. [Google Scholar] [CrossRef]
- Hovda, K.E.; Gadeholt, G.; Evtodienko, V.; Jacobsen, D. A novel bedside diagnostic test for methanol poisoning using dry chemistry for formate. Scand. J. Clin. Lab. Investig. 2015, 75, 610–614. [Google Scholar] [CrossRef]
- Hovda, K.E.; Urdal, P.; Jacobsen, D. Increased Serum Formate in the Diagnosis of Methanol Poisoning. J. Anal. Toxicol. 2005, 29, 586–588. [Google Scholar] [CrossRef] [PubMed]
- Pohanka, M. Antidotes Against Methanol Poisoning: A Review. Mini-Rev. Med. Chem. 2019, 19, 1126–1133. [Google Scholar] [CrossRef]
- Shin, W.-H.; Kihara, D. 55 Years of the Rossmann Fold. Methods Mol. Biol. 2019, 1958, 1–13. [Google Scholar]
- Hanukoglu, I. Proteopedia: Rossmann fold: A beta-alpha-beta fold at dinucleotide binding sites. Biochem. Mol. Biol. Educ. 2015, 43, 206–209. [Google Scholar] [CrossRef] [PubMed]
- Munsamy, G.; Soliman, M.E.S. Unveiling a New Era in Malaria Therapeutics: A Tailored Molecular Approach Towards the Design of Plasmepsin IX Inhibitors. Protein J. 2019, 38, 616–627. [Google Scholar] [CrossRef]
- Messaoudi, A.; Zoghlami, M.; Basharat, Z.; Sadfi-Zouaoui, N. Identification of a Potential Inhibitor Targeting MurC Ligase of the Drug Resistant Pseudomonas aeruginosa Strain through Structure-Based Virtual Screening Approach and In Vitro Assay. Curr. Pharm. Biotechnol. 2019, 20, 1203–1212. [Google Scholar] [CrossRef] [PubMed]
- Prathiviraj, R.; Chellapandi, P. Deciphering Molecular Virulence Mechanism of Mycobacterium tuberculosis Dop isopeptidase Based on Its Sequence–Structure–Function Linkage. Protein J. 2020, 39, 33–45. [Google Scholar] [CrossRef] [PubMed]
- Neetu, N.; Sharma, M.; Mahto, J.K.; Kumar, P. Biophysical and In-Silico Studies of Phytochemicals Targeting Chorismate Synthase from Drug-Resistant Moraxella Catarrhalis. Protein J. 2020, 39, 449–460. [Google Scholar] [CrossRef]
- Kataria, R.; Khatkar, A. Molecular Docking of Natural Phenolic Compounds for the Screening of Urease Inhibitors. Curr. Pharm. Biotechnol. 2019, 20, 410–421. [Google Scholar] [CrossRef]
- Mozafari, M.; Tariverdian, T.; Beynaghi, A. Trends in Biotechnology at the Turn of the Millennium. Recent Patents Biotechnol. 2020, 14, 78–82. [Google Scholar] [CrossRef]
- Dalmizrak, O.; Teralı, K.; Asuquo, E.B.; Ogus, I.H.; Ozer, N. The Relevance of Glutathione Reductase Inhibition by Fluoxetine to Human Health and Disease: Insights Derived from a Combined Kinetic and Docking Study. Protein J. 2019, 38, 515–524. [Google Scholar] [CrossRef] [PubMed]
- Bazaes, S.E. Affinity labels as probes for the study of the nucleotide binding site of enzymes. Biol. Res. 1996, 29, 21–30. [Google Scholar]
- Labrou, N.E.; Muharram, M.M.; Abdelkader, M.S. Delineation of the structural and functional role of Arg111 in GSTU4-4 from Glycine max by chemical modification and site-directed mutagenesis. Biochim. Biophys. Acta (BBA)-Proteins Proteom. 2016, 1864, 1315–1321. [Google Scholar] [CrossRef]
- Colman, R.F.; Huang, Y.-C.; King, M.M.; Erb, M. 6-[(4-bromo-2,3-dioxobutyl)thio]-6-deaminoadenosine 5′-monophosphate and 5′-diphosphate: New affinity labels for purine nucleotide sites in proteins. Biochemistry 1984, 23, 3281–3286. [Google Scholar] [CrossRef] [PubMed]
- Chabot, N.; Vinatier, V.; Gefflaut, T.; Baudoin, C.; Rodriguez, F.; Blonski, C.; Hoffmann, P. Irreversible inhibition of aldolase by a phosphorylated α-dicarbonyl compound. J. Enzym. Inhib. Med. Chem. 2008, 23, 21–27. [Google Scholar] [CrossRef]
- Patthy, L.; Thész, J. Origin of the selectivity of alpha-dicarbonyl reagents for arginyl residues of anion-binding sites. JBIC J. Biol. Inorg. Chem. 1980, 105, 387–393. [Google Scholar] [CrossRef] [PubMed]
- Egorov, A.M.; Tishkov, V.I.; Popov, V.O.; Berezin, I.V. Study of the role of arginine residues in bacterial formate dehydrogenase. Biochim. Biophys. Acta. 1981, 659, 141–149. [Google Scholar] [CrossRef]
- Kayikci, M.; Venkatakrishnan, A.J.; Scott-Brown, J.; Ravarani, C.N.J.; Flock, T.; Babu, M.M. Visualization and analysis of non-covalent contacts using the Protein Contacts Atlas. Nat. Struct. Mol. Biol. 2018, 25, 185–194. [Google Scholar] [CrossRef]
- Vollmer, S.H.; Walner, M.B.; Tarbell, K.V.; Colman, R.F. Guanosine 5′-0-[S-(4-Bromo-2,3-dioxobutyl)]thiophosphate and Adenosine 5′-0-[S-(4-Bromo-2,3- dioxobutyl)]thiophosphate: New Nucleotide Affinity Labels which React with Rabbit Muscle Pyruvate Kinase. J. Biol. Chem. 1994, 269, 8082–8090. [Google Scholar] [CrossRef]
- Kitz, R.; Wilson, I.B. Esters of Methanesulfonic Acid as Irreversible Inhibitors of Acetylcholinesterase. J. Biol. Chem. 1962, 237, 3245–3249. [Google Scholar] [CrossRef]
- Kotzia, G.A.; Labrou, N.E. S-(2,3-dichlorotriazinyl)glutathione. A new affinity label for probing the structure and function of glutathione transferases. Eur. J. Biochem. 2004, 271, 3503–3511. [Google Scholar] [CrossRef] [PubMed]
- AlQarni, M.H.; Muharram, M.M.; Labrou, N.E.; AlQarni, M.H. Ligand-induced glutathione transferase degradation as a therapeutic modality: Investigation of a new metal-mediated affinity cleavage strategy for human GSTP1-1. Int. J. Biol. Macromol. 2018, 116, 84–90. [Google Scholar] [CrossRef] [PubMed]
- Platis, M.; Vlachakis, D.; Foudah, A.I.; Muharram, M.M.; AlQarni, M.H.; Papageorgiou, A.C.; Labrou, N.E. The interaction of Schistosoma japonicum glutathione transferase with Cibacron blue 3GA and its fragments. Med. Chem. 2020, 16, 1–28. [Google Scholar] [CrossRef] [PubMed]
- Sokalingam, S.; Raghunathan, G.; Soundrarajan, N.; Lee, S.-G. A Study on the Effect of Surface Lysine to Arginine Mutagenesis on Protein Stability and Structure Using Green Fluorescent Protein. PLoS ONE 2012, 7, e40410. [Google Scholar] [CrossRef]
- Kumar, S.; Tsai, C.-J.; Nussinov, R. Factors enhancing protein thermostability. Protein Eng. Des. Sel. 2000, 13, 179–191. [Google Scholar] [CrossRef]
- Marinou, M.; Platis, D.; Ataya, F.S.; Chronopoulou, E.; Vlachakis, D.; Labrou, N.E. Structure-based design and application of a nucleotide coenzyme mimetic ligand: Application to the affinity purification of nucleotide dependent enzymes. J. Chromatogr. A 2018, 1535, 88–100. [Google Scholar] [CrossRef]
- Patel, N.; Shahane, S.; Majumdar, R.; Mishra, U. Mode of Action, Properties, Production, and Application of Laccase: A Review. Recent Patents Biotechnol. 2019, 13, 19–32. [Google Scholar] [CrossRef]
- Fan, Y.-X.; McPHIE, P.; Miles, E.W. Regulation of Tryptophan Synthase by Temperature, Monovalent Cations, and an Allosteric Ligand. Evidence from Arrhenius Plots, Absorption Spectra, and Primary Kinetic Isotope Effects. Biochemistry 2000, 39, 4692–4703. [Google Scholar] [CrossRef]
- Peng, G.; Fritzsch, G.; Zickermann, V.; Schagger, H.; Mentele, R.; Lottspeich, F.; Bostina, M.; Radermacher, M.; Huber, R.; Stetter, K.O.; et al. Isolation, Characterization and Electron Microscopic Single Particle Analysis of the NADH:Ubiquinone Oxidoreductase (Complex I) from the Hyperthermophilic Eubacterium Aquifex aeolicus. Biochemistry 2003, 42, 3032–3039. [Google Scholar] [CrossRef]
- Zeeshan, F.; Tabbassum, M.; Kesharwani, P. Investigation on Secondary Structure Alterations of Protein Drugs as an Indicator of Their Biological Activity Upon Thermal Exposure. Protein J. 2019, 38, 551–564. [Google Scholar] [CrossRef]
- Strzelecka, D.; Chmielinski, S.; Bednarek, S.; Jemielity, J.; Kowalska, J. Analysis of mononucleotides by tandem mass spectrometry: Investigation of fragmentation pathways for phosphate- and ribose-modified nucleotide analogues. Sci. Rep. 2017, 7, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Labrou, N.E.; Rigden, D.J. Active-site characterization of Candida boidinii formate dehydrogenase. Biochem. J. 2001, 354, 455–463. [Google Scholar] [CrossRef] [PubMed]
- Bradford, M.A. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Kato, N.; Sahm, H.; Wagner, F. Steady-state kinetics of formaldehyde dehydrogenase and formate dehydrogenase from a methanol-utilizing yeast, Candida boidinii. Biochim. Biophys. Acta (BBA)-Enzym. 1979, 566, 12–20. [Google Scholar] [CrossRef]
- Deng, W.P.; Nickoloff, J.A. Site-directed mutagenesis of virtually any plasmid by eliminating a unique site. Anal. Biochem. 1992, 200, 81–90. [Google Scholar] [CrossRef]
- Labrou, N.E.; Kotzia, G.A.; Clonis, Y.D. Engineering the xenobiotic substrate specificity of maize glutathione S-transferase I. Protein Eng. Des. Sel. 2004, 17, 741–748. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Enzyme | Km, (μM) (NAD+) | Km, (mM) (Formate) | kcat (% of the Wild-Type) | Ki (mM) (NADH) | Ki (mM) (AMP) |
|---|---|---|---|---|---|
| Wild-type | 0.04 ± 0.003 | 2.4 ± 0.1 | 100 | 0.03 ± 0.005 | 1.6 ± 0.1 |
| Arg174Asn | 1.27 ± 0.1 | 3.1 ± 0.2 | 17.5 | 1.1 ± 0.05 | 9.2 ± 0.3 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Alqarni, M.H.; Foudah, A.I.; Muharram, M.M.; Budurian, H.; Labrou, N.E. Probing the Role of the Conserved Arg174 in Formate Dehydrogenase by Chemical Modification and Site-Directed Mutagenesis. Molecules 2021, 26, 1222. https://doi.org/10.3390/molecules26051222
Alqarni MH, Foudah AI, Muharram MM, Budurian H, Labrou NE. Probing the Role of the Conserved Arg174 in Formate Dehydrogenase by Chemical Modification and Site-Directed Mutagenesis. Molecules. 2021; 26(5):1222. https://doi.org/10.3390/molecules26051222
Chicago/Turabian StyleAlqarni, Mohammed Hamed, Ahmed Ibrahim Foudah, Magdy Mohamed Muharram, Haritium Budurian, and Nikolaos E. Labrou. 2021. "Probing the Role of the Conserved Arg174 in Formate Dehydrogenase by Chemical Modification and Site-Directed Mutagenesis" Molecules 26, no. 5: 1222. https://doi.org/10.3390/molecules26051222
APA StyleAlqarni, M. H., Foudah, A. I., Muharram, M. M., Budurian, H., & Labrou, N. E. (2021). Probing the Role of the Conserved Arg174 in Formate Dehydrogenase by Chemical Modification and Site-Directed Mutagenesis. Molecules, 26(5), 1222. https://doi.org/10.3390/molecules26051222