
Synergistic Roles of Curcumin in Sensitising the Cisplatin Effect on a Cancer Stem Cell-Like Population Derived from Non-Small Cell Lung Cancer Cell Lines




Abstract
1. Introduction
2. Results
2.1. Curcumin Sensitises the Cisplatin Effect Leading to Decreased Cells in
2.1.1. Proliferation Activity
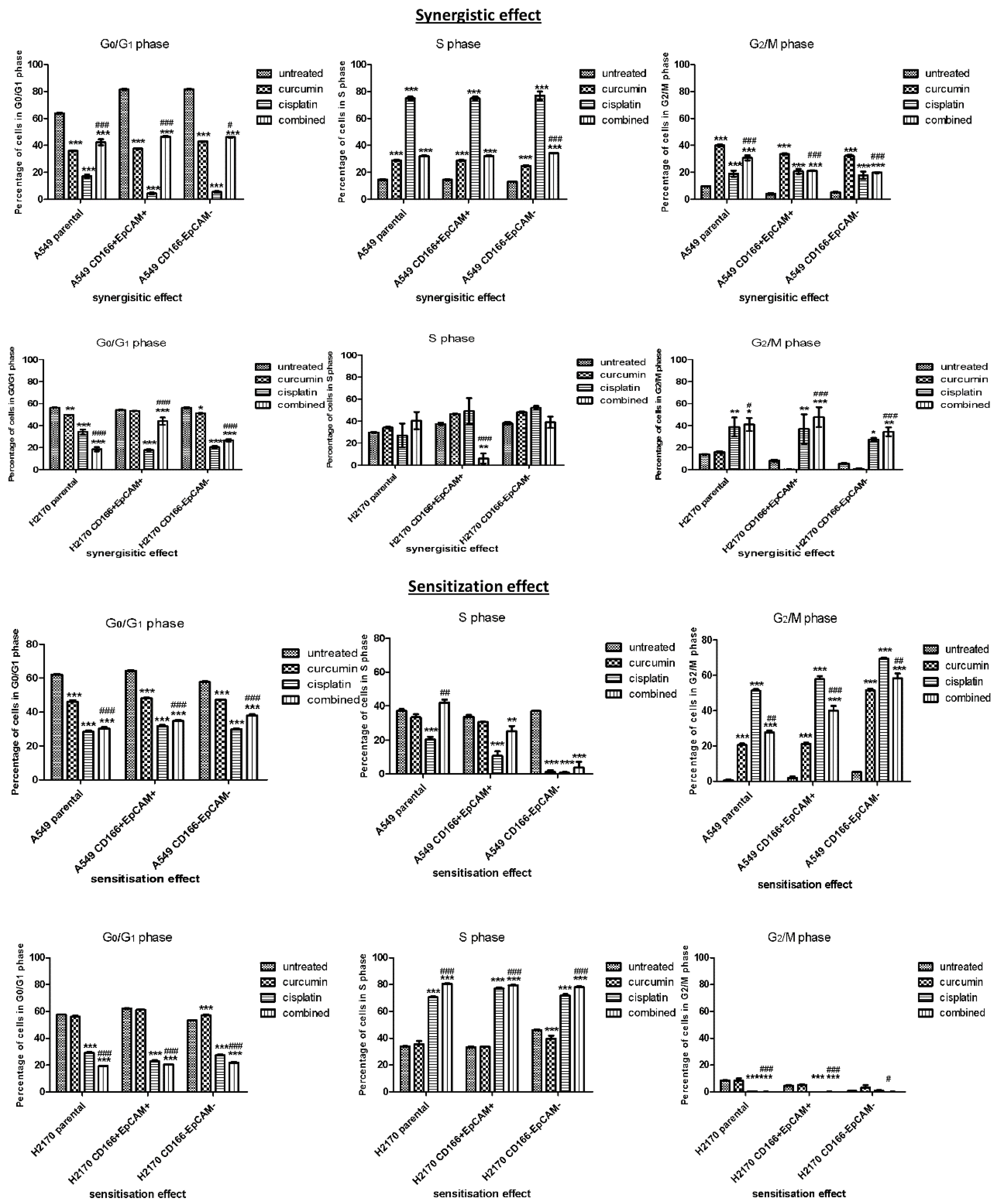
2.1.2. Cell Cycle Regulation in CSCs of NSCLC
Curcumin Arrests the Cell Cycle in CSCs Subpopulation by Synergistic and Sensitisation Treatment Groups
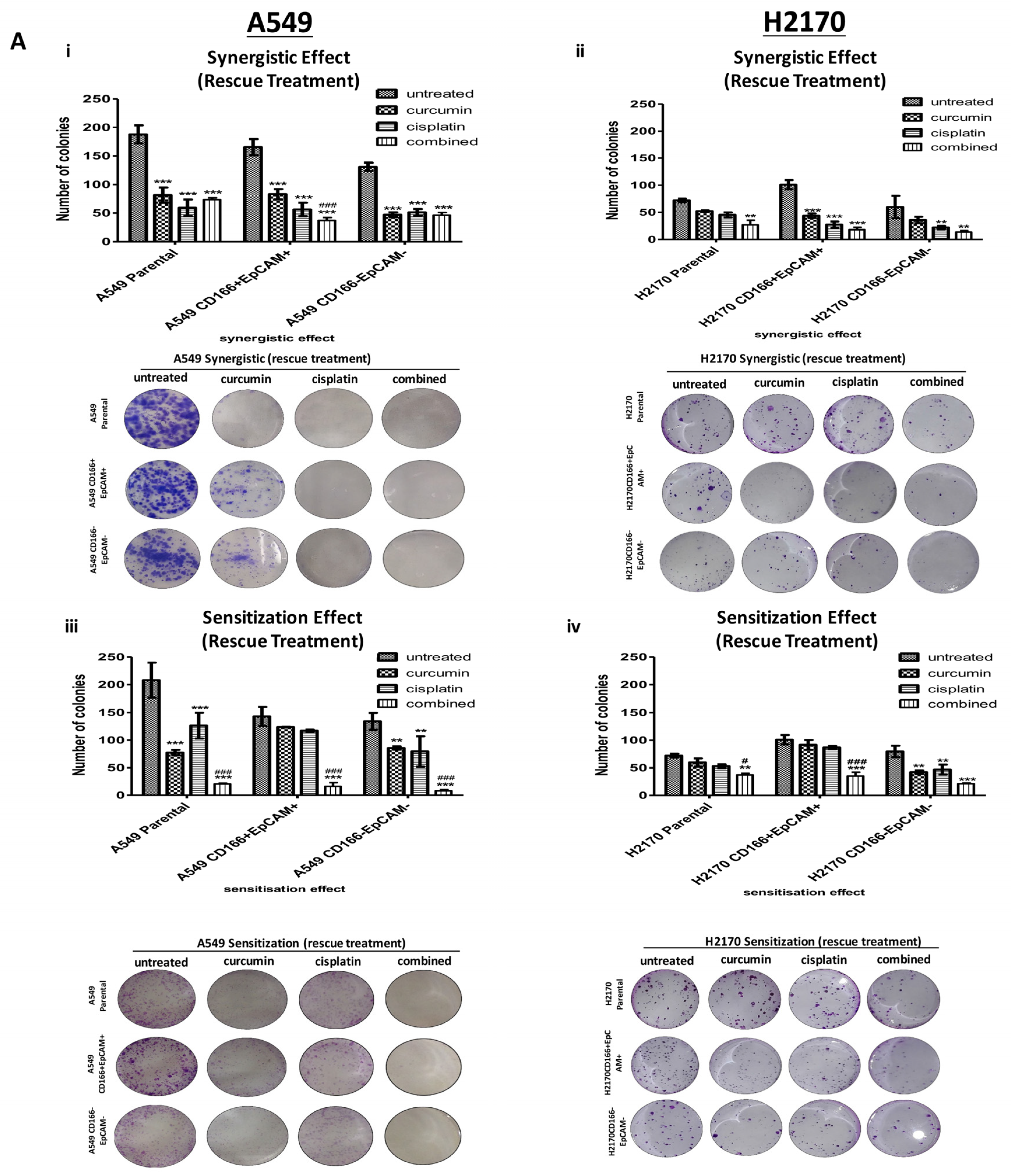
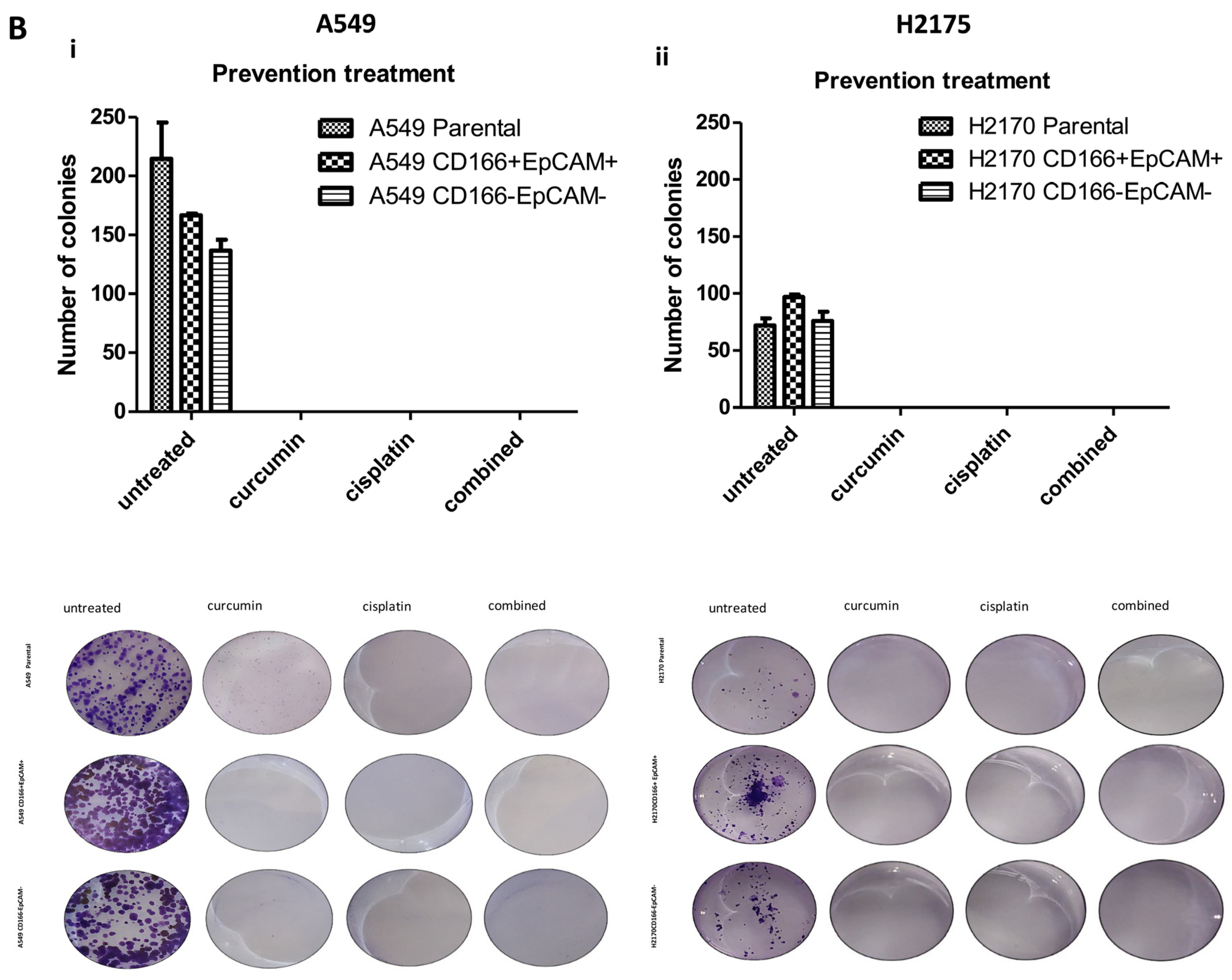
2.2. Curcumin Suppresses and Prevents the Self-Renewal Characteristics of CSCs Subpopulation
2.2.1. Curcumin Suppresses and Prevents the Colonies Formation
2.2.2. Curcumin Suppresses and Prevents Spheroid Development
2.3. Curcumin Regulated Stemness-Related Genes in CSC and Non-CSC Subpopulations
2.4. Secreted Proteins Present in the CSC and Non-CSC Subpopulations
3. Discussion
4. Materials and Methods
4.1. Cell Culture
4.2. Preparation of Curcumin and Cisplatin
4.3. Cell Proliferation Assay
4.4. Cell Cycle Analysis by Flow Cytometry
4.5. Clonogenic Assay
4.6. Spheroid Assay
4.7. Quantitative Real-Time-Polymerase Chain Reaction (qRT-PCR)
4.8. Preparation of Condition Medium
4.9. Trichloroacetic Acid (TCA) Precipitation
4.10. Micro BCA Assay
4.11. In-Solution Protein Digestion
4.12. Liquid Chromatography-Mass Spectrometry (LC-MS/MS) Analysis
4.13. Protein Identification and Analysis
4.14. Functional Group Analysis
4.15. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Templeton, A.K.; Miyamoto, S.; Babu, A.; Munshi, A.; Ramesh, R. Cancer stem cells: Progress and challenges in lung cancer. Stem Cell Investig. 2014, 1, 9. [Google Scholar] [PubMed]
- Zakaria, N.; Abdul Satar, N.; Abu Halim, N.H.; Ngalim, S.H.; Mohd Yusof, N.; Lin, J.; Yahaya, B.H. Targeting lung cancer stem cells: Research and clinical impacts. Front. Oncol. 2017, 7, 80. [Google Scholar] [CrossRef] [PubMed]
- Yagui-Beltrán, A.; Jablons, D.M. A translational approach to lung cancer research: From EGFRs to Wnt and cancer stem cells. Ann. Thorac. Cardiovasc. Surg. 2009, 15, 213–220. [Google Scholar] [PubMed]
- Al-Hajj, M.; Wicha, M.S.; Benito-Hernandez, A.; Morrison, S.J.; Clarke, M.F. Prospective identification of tumorigenic breast cancer cells. Proc. Natl. Acad. Sci. USA 2003, 100, 3983–3988. [Google Scholar] [CrossRef] [PubMed]
- Talkowski, K.; Kiełbasiński, K.; Peszek, W.; Grabarek, B.O.; Boroń, D.; Oplawski, M. Salinomycin Modulates the Expression of mRNAs and miRNAs Related to Stemness in Endometrial Cancer. Curr. Pharm. Biotechnol. 2020, 21, 1–10. [Google Scholar] [CrossRef]
- Phi, L.T.H.; Sari, I.T.; Yang, Y.G.; Lee, S.H.; Jun, N.; Kim, K.S.; Lee, Y.K.; Kwon, H.Y. Cancer Stem Cells (CSCs) in Drug Resistance and their Therapeutic Implications in Cancer Treatment. Stem Cells Int. 2018, 2018, 5416923. [Google Scholar] [CrossRef]
- Nunes, T.; Hamdan, D.; Leboeuf, C.; Bouchtaoui, M.E.; Gapihan, G.; Nguyen, T.T.; Meles, S.; Angeli, E.; Ratajczak, P.; Lu, H.; et al. Targeting cancer stem cells to overcome chemoresistance. Int. J. Mol. Sci. 2018, 19, 4036. [Google Scholar] [CrossRef]
- De Angelis, M.L.; Francescangeli, F.; La Torre, F.; Zeuner, A. Stem cell plasticity and dormancy in the development of cancer therapy resistance. Front. Oncol. 2019, 9, 626. [Google Scholar] [CrossRef]
- Bao, B.; Ahmad, A.; Azmi, A.S.; Ali, S.; Sarkar, F.H. Overview of cancer stem cells (CSCS) and mechanisms of their regulation: Implications for cancer therapy. Curr. Protoc. Pharmacol. 2013, 61, 14–25. [Google Scholar] [CrossRef]
- Calabrese, C.; Poppleton, H.; Kocak, M.; Hogg, T.L.; Fuller, C.; Hamner, B.; Oh, E.Y.; Gaber, M.W.; Finklestein, D.; Allen, M.; et al. A Perivascular Niche for Brain Tumor Stem Cells. Cancer Cell 2007, 11, 69–82. [Google Scholar] [CrossRef]
- Hill, R.P.; Marie-Egyptienne, D.T.; Hedley, D.W. Cancer Stem Cells, Hypoxia and Metastasis. Semin. Radiat. Oncol. 2009, 19, 106–111. [Google Scholar] [CrossRef]
- Rahmani, A.H.; Al Zohairy, M.A.; Aly, S.M.; Khan, M.A. Curcumin: A potential candidate in prevention of cancer via modulation of molecular pathways. Biomed Res. Int. 2014, 2014, 761608. [Google Scholar] [CrossRef] [PubMed]
- Tomeh, M.A.; Hadianamrei, R.; Zhao, X. A review of curcumin and its derivatives as anticancer agents. Int. J. Mol. Sci. 2019, 20, 1033. [Google Scholar] [CrossRef] [PubMed]
- Zhou, H.S.; Beevers, C.; Huang, S. The Targets of Curcumin. Curr. Drug Targets 2012, 12, 332–347. [Google Scholar] [CrossRef] [PubMed]
- Baharuddin, P.; Abdul Satar, N.; Fakiruddin, K.S.; Zakaria, N.; Lim, M.N.; Mohd Yusoff, N.; Zakaria, Z.; Yahaya, B.H. Curcumin improves the efficacy of cisplatin by targeting cancer stem-like cells through p21 and cyclin D1-mediated tumour cell inhibition in non-small cell lung cancer cell lines. Oncol. Rep. 2016, 35, 13–25. [Google Scholar] [CrossRef] [PubMed]
- Zakaria, N.; Mohd Yusoff, N.; Zakaria, Z.; Lim, M.N.; Baharuddin, P.; Fakiruddin, K.S.; Yahaya, B.H. Human non-small cell lung cancer expresses putative cancer stem cell markers and exhibits the transcriptomic profile of multipotent cells. BMC Cancer 2015, 15, 84. [Google Scholar] [CrossRef]
- Halim, N.H.A.; Zakaria, N.; Satar, N.A.; Yahaya, B.H. Isolation and characterization of cancer stem cells of the non-small-cell lung cancer (A549) cell line. In Methods in Molecular Biology; Humana Press Inc.: New York, NY, USA, 2016; Volume 1516, pp. 371–388. [Google Scholar]
- Zeuner, A. The secret life of quiescent cancer stem cells. Mol. Cell. Oncol. 2015, 2, 1–2. [Google Scholar] [CrossRef][Green Version]
- Fuertes, M.A.; Alonso, C.; Perez, J.M. Biochemical Modulation of Cisplatin Mechanisms of Action: Enhancement of Antitumor Activity and Circumvention of Drug Resistance. Chem. Rev. 2003, 103, 3, 645–662. [Google Scholar] [CrossRef]
- Kelland, L. The resurgence of platinum-based cancer chemotherapy. Nat. Rev. Cancer 2007, 7, 573–584. [Google Scholar] [CrossRef]
- Zhu, J.Y.; Yang, X.; Chen, Y.; Jiang, Y.; Wang, S.J.; Li, Y.; Wang, X.Q.; Meng, Y.; Zhu, M.M.; Ma, X.; et al. Curcumin Suppresses Lung Cancer Stem Cells via Inhibiting Wnt/β-catenin and Sonic Hedgehog Pathways. Phyther. Res. 2017, 31, 680–688. [Google Scholar] [CrossRef]
- Chung, S.S.; Vadgama, J.V. Curcumin and epigallocatechin gallate inhibit the cancer stem cell phenotype via down-regulation of STAT3-NFκB signaling. Anticancer Res. 2015, 35, 39–46. [Google Scholar] [PubMed]
- Takaishi, S.; Okumura, T.; Tu, S.; Wang, S.S.W.; Shibata, W.; Vigneshwaran, R.; Gordon, S.A.K.; Shimada, Y.; Wang, T.C. Identification of gastric cancer stem cells using the cell surface marker CD44. Stem Cells 2009, 27, 1006–1020. [Google Scholar] [CrossRef] [PubMed]
- Izumiya, M.; Kabashima, A.; Higuchi, H.; Igarashi, T.; Sakai, G.; Iizuka, H.; Nakamura, S.; Adachi, M.; Hamamoto, Y.; Funakoshi, S.; et al. Chemoresistance is associated with cancer stem cell-like properties and epithelial-to-mesenchymal transition in pancreatic cancer cells. Anticancer Res. 2012, 32, 3847–3853. [Google Scholar] [PubMed]
- Aggarwal, B.B.; Shishodia, S. Molecular targets of dietary agents for prevention and therapy of cancer. Biochem. Pharmacol. 2006, 71, 1397–1421. [Google Scholar] [CrossRef] [PubMed]
- Siddappa, G.; Kulsum, S.; Ravindra, D.R.; Kumar, V.V.; Raju, N.; Raghavan, N.; Sudheendra, H.V.; Sharma, A.; Sunny, S.P.; Jacob, T.; et al. Curcumin and metformin-mediated chemoprevention of oral cancer is associated with inhibition of cancer stem cells. Mol. Carcinog. 2017, 56, 2446–2460. [Google Scholar] [CrossRef] [PubMed]
- Zhang, S.; Xiong, X.; Sun, Y.; Self-renewal, S.C. Functional characterization of SOX2 as an anticancer target. Signal Transduct. Target. Ther. 2020, 5, 1–7. [Google Scholar] [CrossRef]
- Lu, Y.; Zhu, H.; Shan, H.; Lu, J.; Chang, X.; Li, X.; Lu, J.; Fan, X.; Zhu, S.; Wang, Y.; et al. Knockdown of Oct4 and Nanog expression inhibits the stemness of pancreatic cancer cells. Cancer Lett. 2013, 340, 113–123. [Google Scholar] [CrossRef]
- Shan, J.; Shen, J.; Liu, L.; Xia, F.; Xu, C.; Duan, G.; Xu, Y.; Ma, Q.; Yang, Z.; Zhang, Q.; et al. Nanog regulates self-renewal of cancer stem cells through the insulin-like growth factor pathway in human hepatocellular carcinoma. Hepatology 2012, 56, 1004–1014. [Google Scholar] [CrossRef]
- Müller, M.; Hermann, P.C.; Liebau, S.; Weidgang, C.; Seufferlein, T.; Kleger, A.; Perkhofer, L. The role of pluripotency factors to drive stemness in gastrointestinal cancer. Stem Cell Res. 2016, 16, 349–357. [Google Scholar] [CrossRef]
- Lacina, L.; Plzak, J.; Kodet, O.; Szabo, P.; Chovanec, M.; Dvorankova, B.; Smetana Jr, K. Cancer microenvironment: What can we learn from the stem cell niche. Int. J. Mol. Sci. 2015, 16, 24094–24110. [Google Scholar] [CrossRef]
- Plaks, V.; Kong, N.; Werb, Z. The cancer stem cell niche: How essential is the niche in regulating stemness of tumor cells? Cell Stem Cell 2015, 16, 225–238. [Google Scholar] [CrossRef] [PubMed]
- Huang, C.P.; Tsai, M.F.; Chang, T.H.; Tang, W.C.; Chen, S.Y.; Lai, H.H.; Lin, T.Y.; Yang, J.C.H.; Yang, P.C.Y.; Shih, J.Y.; et al. ALDH-positive lung cancer stem cells confer resistance to epidermal growth factor receptor tyrosine kinase inhibitors. Cancer Lett. 2013, 328, 144–151. [Google Scholar] [CrossRef] [PubMed]
- Xu, X.; Chai, S.; Wang, P.; Zhang, C.; Yang, Y.; Yang, Y.; Wang, K. Aldehyde dehydrogenases and cancer stem cells. Cancer Lett. 2015, 369, 50–57. [Google Scholar] [CrossRef] [PubMed]
- Hsu, N.Y.; Ho, H.C.; Chow, K.C.; Lin, T.Y.; Shih, C.S.; Wang, L.S.; Tsai, C.M. Overexpression of dihydrodiol dehydrogenase as a prognostic marker of non-small cell lung cancer. Cancer Res. 2001, 61, 2727–2731. [Google Scholar] [PubMed]
- Deng, H.B.; Adikari, M.; Parekh, H.K.; Simpkins, H. Ubiquitous induction of resistance to platinum drugs in human ovarian, cervical, germ-cell and lung carcinoma tumor cells overexpressing isoforms 1 and 2 of dihydrodiol dehydrogenase. Cancer Chemother. Pharmacol. 2004, 54, 301–307. [Google Scholar] [CrossRef] [PubMed]
- Seo, D.C.; Sung, J.M.; Cho, H.J.; Yi, H.; Seo, K.H.; Choi, I.N.; Kim, D.K.; Kim, J.S.; Abd El-Aty, A.M.; Shin, H.C. Gene expression profiling of cancer stem cell in human lung adenocarcinoma A549 cells. Mol. Cancer 2007, 6, 2–9. [Google Scholar] [CrossRef] [PubMed]
- Martin, H.J.; Breyer-Pfaff, U.; Wsol, V.; Venz, S.; Block, S.; Maser, E. Purification and characterization of AKR1B10 from human liver: Role in carbonyl reduction of xenobiotics. Drug Metab. Dispos. 2006, 34, 464–470. [Google Scholar] [CrossRef]
- Balendiran, G.K.; Martin, H.J.; El-Hawari, Y.; Maser, E. Cancer biomarker AKR1B10 and carbonyl metabolism. Chem. Biol. Interact. 2009, 178, 134–137. [Google Scholar] [CrossRef]
- Zhao, Y.; Dong, Q.; Li, J.; Zhang, K.; Qin, J.; Zhao, J.; Sun, Q.; Wang, Z.; Wartmann, T.; Jauch, K.W.; et al. Seminars in Cancer Biology Targeting cancer stem cells and their niche: Perspectives for future therapeutic targets and strategies. Semin. Cancer Biol. 2018, 53, 139–155. [Google Scholar] [CrossRef]
- Sleeman, J.P.; Cremers, N. New concepts in breast cancer metastasis: Tumor initiating cells and the microenvironment. Clin. Exp. Metastasis. 2007, 24, 707–715. [Google Scholar] [CrossRef]
- Ribatti, D.; Mangialardi, G.; Vacca, A. Stephen Paget and the ‘seed and soil’ theory of metastatic dissemination. Clin. Exp. Med. 2006, 6, 145–149. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.F.; Xiao, Z.Q.; Li, M.X.; Li, M.Y.; Zhang, P.F.; Li, C.; Li, F.; Chen, Y.H.; Yi, H.; Yao, H.X.; et al. Quantitative proteome analysis reveals annexin A3 as a novel biomarker in lung adenocarcinoma. J. Pathol. 2009, 217, 54–64. [Google Scholar] [CrossRef] [PubMed]
- Maschler, S.; Gebeshuber, C.A.; Wiedemann, E.M.; Alacakaptan, M.; Schreiber, M.; Custir, I.; Beug, H. Annexin A1 attenuates EMT and metastatic potential in breast cancer. EMBO Mol. Med. 2010, 2, 401–414. [Google Scholar] [CrossRef] [PubMed]
- Geary, L.A.; Nash, K.A.; Adisetiyo, H.; Liang, M.; Liao, C.P.; Jeong, J.H.; Zandi, E.; Roy-burman, P. CAF-secreted annexin A1 induces prostate cancer cells to gain stem cell-like features. Mol. Cancer Res. 2014, 12, 607–621. [Google Scholar] [CrossRef]
- Chen, S.; Huang, E.H. The Colon Cancer Stem Cell Microenvironment Holds Keys to Future Cancer Therapy. J. Gastrointest. Surg. 2014, 18, 1040–1048. [Google Scholar] [CrossRef]
- Harvey, K.F.; Zhang, X.; Thomas, D.M. The Hippo pathway and human cancer. Nat. Rev. Cancer 2013, 13, 246–257. [Google Scholar] [CrossRef]
- Lee, S.M.; Na, Y.K.; Hong, H.S.; Jang, E.J.; Yoon, G.S.; Park, J.Y.; Kim, D.S. Hypomethylation of the thymosin ß10 gene is not associated with its overexpression in non-small cell lung cancer. Mol. Cells 2011, 32, 343–348. [Google Scholar] [CrossRef]
- Zhang, X.J.; Su, Y.R.; Liu, D.; Xu, D.B.; Zheng, M.Z.; Chen, W.K. Thymosin beta 10 correlates with lymph node metastases of papillary thyroid carcinoma. J. Surg. Res. 2014, 192, 487–493. [Google Scholar] [CrossRef]
- Ding, Z.; Joy, M.; Bhargava, R.; Gunsaulus, M.; Lakshman, N.; Miron-Mendoza, M.; Petroll, M.; Condeelis, J.; Wells, A.; Roy, P. Profilin-1 downregulation has contrasting effects on early vs late steps of breast cancer metastasis. Oncogene 2014, 33, 2065–2074. [Google Scholar] [CrossRef]
- Zou, L.; Jaramillo, M.; Whaley, D.; Wells, A.; Panchapakesa, V.; Das, T.; Roy, P. Profilin-1 is a negative regulator of mammary carcinoma aggressiveness. Br. J. Cancer 2007, 97, 1361–1371. [Google Scholar] [CrossRef]
- Du, S.; Guan, Z.; Hao, L.; Song, Y.; Wang, L.; Gong, L.; Liu, L.; Qi, X.; Hou, Z.; Shao, S. Fructose-bisphosphate aldolase a is a potential metastasis-associated marker of lung squamous cell carcinoma and promotes lung cell tumorigenesis and migration. PLoS ONE 2014, 9, e85804. [Google Scholar] [CrossRef] [PubMed]
- Morisaki, T.; Yashiro, M.; Kakehashi, A.; Inagaki, A.; Kinoshita, H.; Fukuoka, T.; Kasashima, H.; Masuda, G.; Sakurai, K.; Kubo, N.; et al. Comparative proteomics analysis of gastric cancer stem cells. PLoS ONE 2014, 9, e110736. [Google Scholar] [CrossRef]
- Chang, Y.S.; Wu, W.; Walsh, G.; Hong, W.K.; Mao, L. Enolase-α Is Frequently Down-Regulated in Non-Small Cell Lung Cancer and Predicts Aggressive Biological Behavior. Clin. Cancer Res. 2003, 9, 3641–3644. [Google Scholar] [PubMed]
- Zhou, X.; Zhang, Y.; Han, N.; Guo, S.; Xiao, T.; Cheng, S.; Gao, Y.; Zhang, K. α-enolase (ENO1) inhibits epithelial-mesenchymal transition in the A549 cell line by suppressing ERK1/2 phosphorylation. Chin. J. Lung Cancer 2013, 16, 221–226. [Google Scholar]
Sample Availability: Samples of the compounds are not available from the authors. |








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Group | KEGG Pathway | Genes | % | p-Value | Benjamin |
|---|---|---|---|---|---|
| A549 CD166-EPCAM-CSCs SUBPOPULATION | Steroid hormone biosynthesis | 4 | 15.4 | 4.2 × 10−4 | 2.3 × 10−2 |
| Glycolysis | 4 | 15.4 | 6.4 × 10−4 | 1.8 × 10−2 | |
| Biosynthesis of amino acids | 4 | 15.4 | 8.6 × 10−4 | 1.6 × 10−2 | |
| Carbon metabolism | 4 | 15.4 | 2.9 × 10−3 | 4.0 × 10−2 | |
| Fructose and mannose metabolism | 3 | 11.5 | 3.0 × 10−3 | 3.3 × 10−2 | |
| * Hippo signalling pathway | 4 | 15.4 | 6.6 × 10−3 | 6.0 × 10−2 | |
| * Metabolism of xenobiotics by cytochrome P450 | 3 | 11.5 | 1.5 × 10−2 | 1.2 × 10−1 | |
| Biosynthesis of antibiotics | 4 | 15.4 | 1.7 × 10−2 | 1.1 × 10−1 | |
| Metabolic pathways | 8 | 30.8 | 2.8 × 10−2 | 1.6 × 10−1 | |
| Oocyte meiosis | 3 | 11.5 | 3.2 × 10−2 | 1.7 × 10−1 | |
| * Cell cycle | 3 | 11.5 | 4.1 × 10−2 | 1.9 × 10−1 | |
| Hepatitis B | 3 | 11.5 | 5.4 × 10−2 | 2.3 × 10−2 | |
| * PI3-AKT signalling pathway | 4 | 15.4 | 5.8 × 10−2 | 2.3 × 10−1 | |
| Influenza A | 3 | 11.5 | 7.4 × 10−2 | 2.7 × 10−1 | |
| Epstein-Barr virus infection | 3 | 11.5 | 8.6 × 10−2 | 2.9 × 10−1 | |
| Viral carcinogenesis | 3 | 11.5 | 9.8 × 10−2 | 3.0 × 10−1 | |
| A549 CD166-EPCAM-NON-CSCs SUBPOPULATION | * Hippo signalling pathway | 8 | 24.2 | 7.2 × 10−7 | 4.5 × 10−4 |
| * Cell cycle | 7 | 21.2 | 4.06 × 10−6 | 1.2 × 10−4 | |
| Oocyte meiosis | 6 | 18.2 | 3.7 × 10−5 | 7.7 × 10−4 | |
| Epstein-Barr virus infection | 7 | 21.2 | 4.6 × 10−5 | 7.2 × 10−4 | |
| Viral carcinogenesis | 7 | 21.2 | 7.0 × 10−5 | 8.8 × 10−4 | |
| * PI3-AKT signalling pathway | 7 | 21.2 | 1.2 × 10−3 | 1.2 × 10−2 | |
| Hypertrophic cardiomyopathy (HCM) | 4 | 12.1 | 2.7 × 10−3 | 2.4 × 10−2 | |
| Dilated cardiomyopathy | 4 | 12.1 | 3.3 × 10−3 | 2.6 × 10−2 | |
| Regulation of actin cytoskeleton | 5 | 15.2 | 6.5 × 10−3 | 4.4 × 10−2 | |
| Shigellosis | 3 | 9.1 | 2.2 × 10−2 | 1.3 × 10−1 | |
| Bacterial invasion of epithelial cells | 3 | 9.1 | 3.2 × 10−2 | 1.7 × 10−1 | |
| Salmonella infection | 3 | 9.1 | 3.6 × 10−2 | 1.7 × 10−1 | |
| Hepatitis B | 3 | 9.1 | 9.6 × 10−2 | 3.9 × 10−1 |
| Accession Number | Gene Symbol | Amplicon Length |
|---|---|---|
| Hs01053049_s1 | SOX2 | 91 |
| Hs00999632_g1 | POU51F | 77 |
| Hs04399610_g1 | NANOG | 101 |
| Hs00358836_m1 | KLF4 | 110 |
| Hs02758991_g1 | GAPDH | 93 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Abdul Satar, N.; Ismail, M.N.; Yahaya, B.H. Synergistic Roles of Curcumin in Sensitising the Cisplatin Effect on a Cancer Stem Cell-Like Population Derived from Non-Small Cell Lung Cancer Cell Lines. Molecules 2021, 26, 1056. https://doi.org/10.3390/molecules26041056
Abdul Satar N, Ismail MN, Yahaya BH. Synergistic Roles of Curcumin in Sensitising the Cisplatin Effect on a Cancer Stem Cell-Like Population Derived from Non-Small Cell Lung Cancer Cell Lines. Molecules. 2021; 26(4):1056. https://doi.org/10.3390/molecules26041056
Chicago/Turabian StyleAbdul Satar, Nazilah, Mohd Nazri Ismail, and Badrul Hisham Yahaya. 2021. "Synergistic Roles of Curcumin in Sensitising the Cisplatin Effect on a Cancer Stem Cell-Like Population Derived from Non-Small Cell Lung Cancer Cell Lines" Molecules 26, no. 4: 1056. https://doi.org/10.3390/molecules26041056
APA StyleAbdul Satar, N., Ismail, M. N., & Yahaya, B. H. (2021). Synergistic Roles of Curcumin in Sensitising the Cisplatin Effect on a Cancer Stem Cell-Like Population Derived from Non-Small Cell Lung Cancer Cell Lines. Molecules, 26(4), 1056. https://doi.org/10.3390/molecules26041056