
Chemical Diversity and Antimicrobial Potential of Cultivable Fungi from Deep-Sea Sediments of the Gulf of Mexico

 ,
,  ,
,  , , , , , and
, , , , , and
Abstract
:
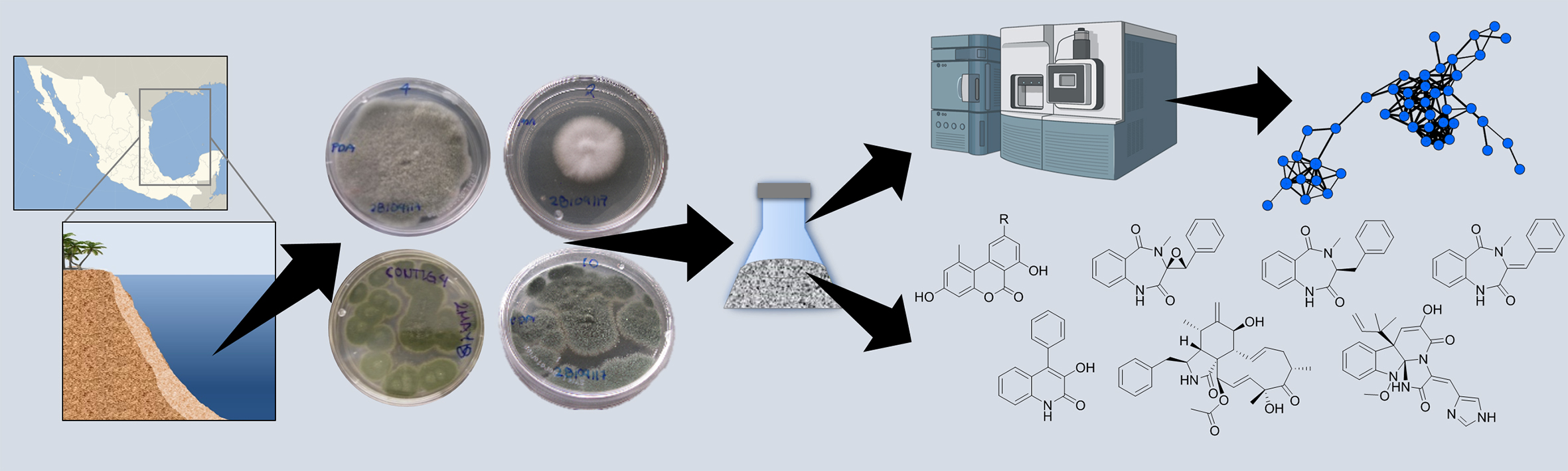
1. Introduction
2. Results and Discussion
2.1. Antimicrobial Screening of the Fungal Collection and Effects of Culture Conditions
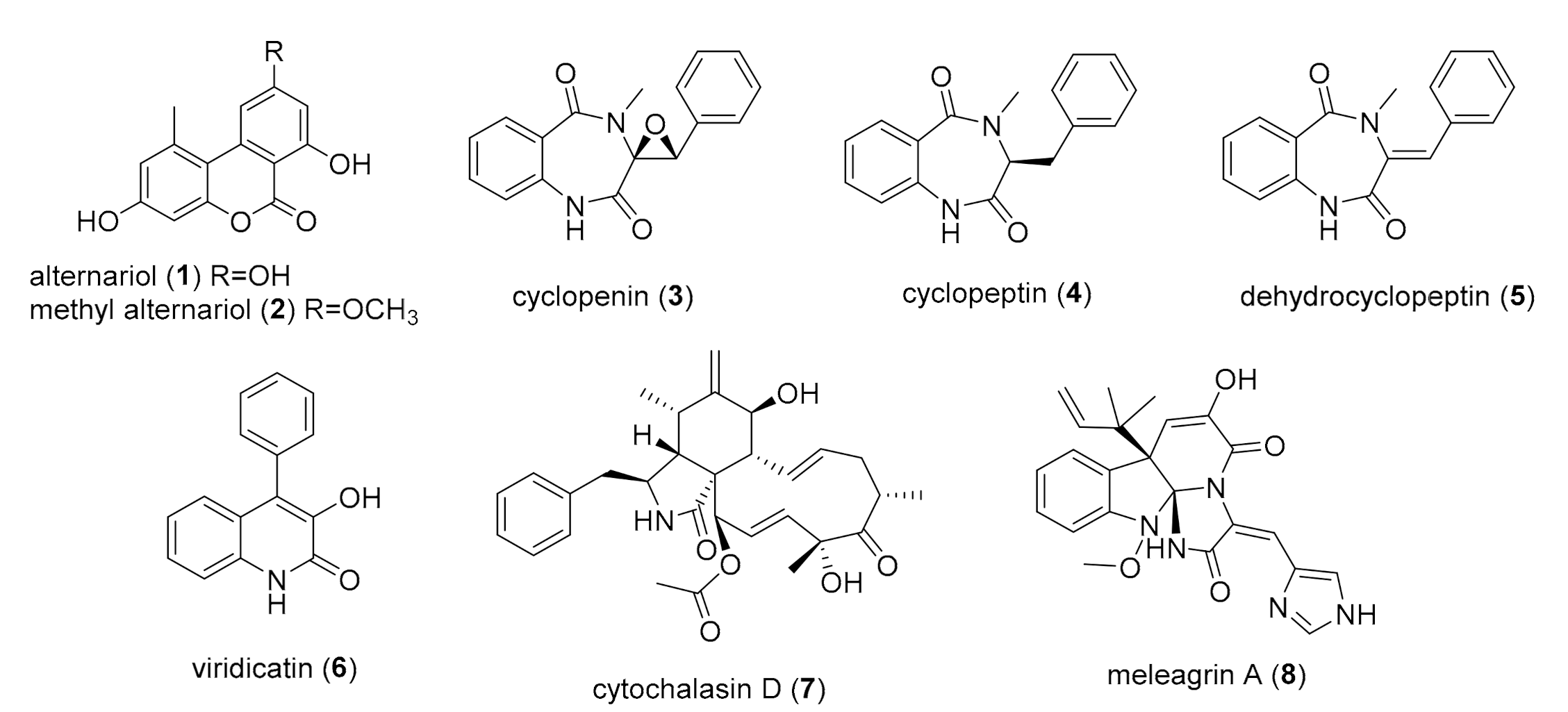
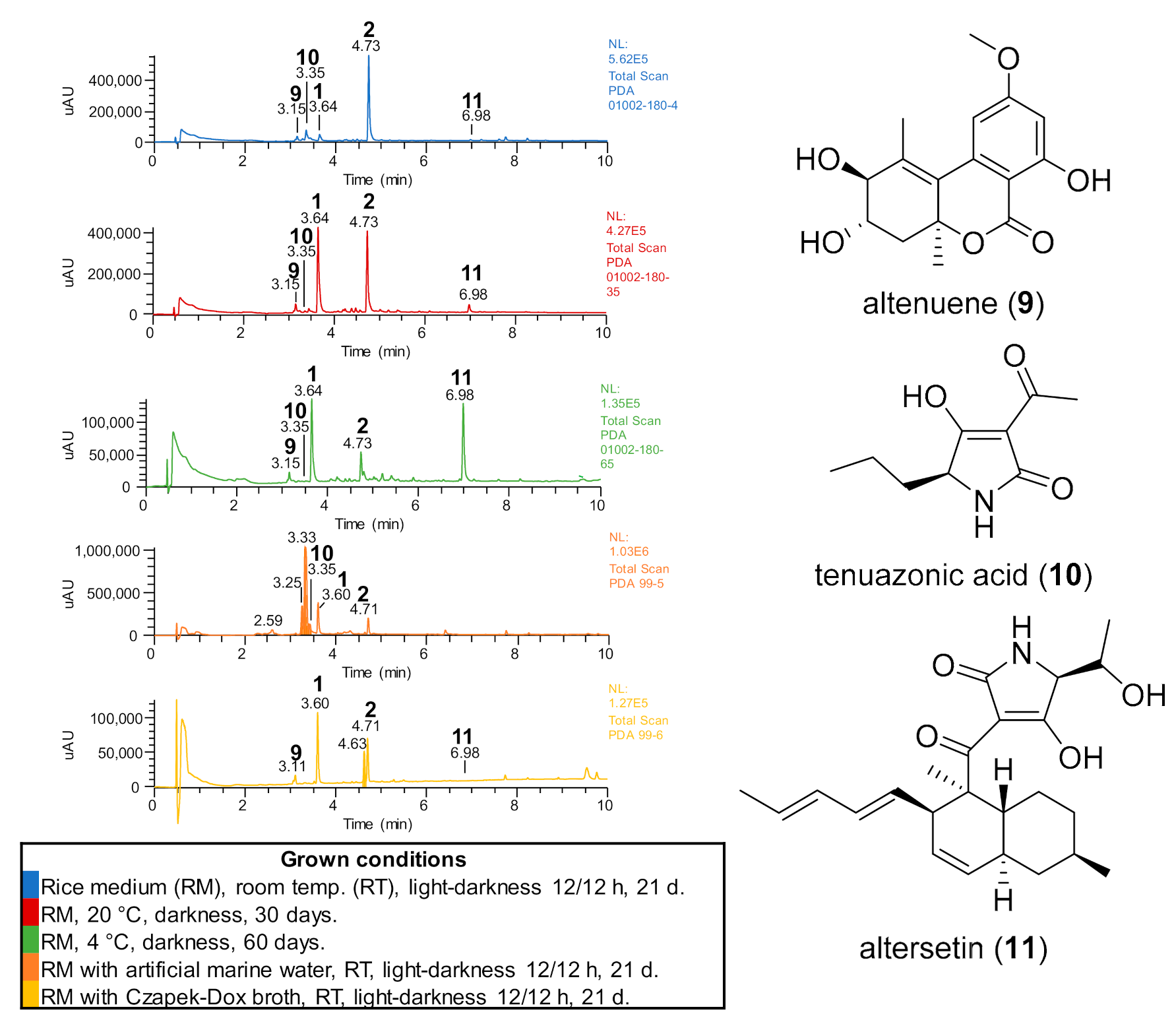
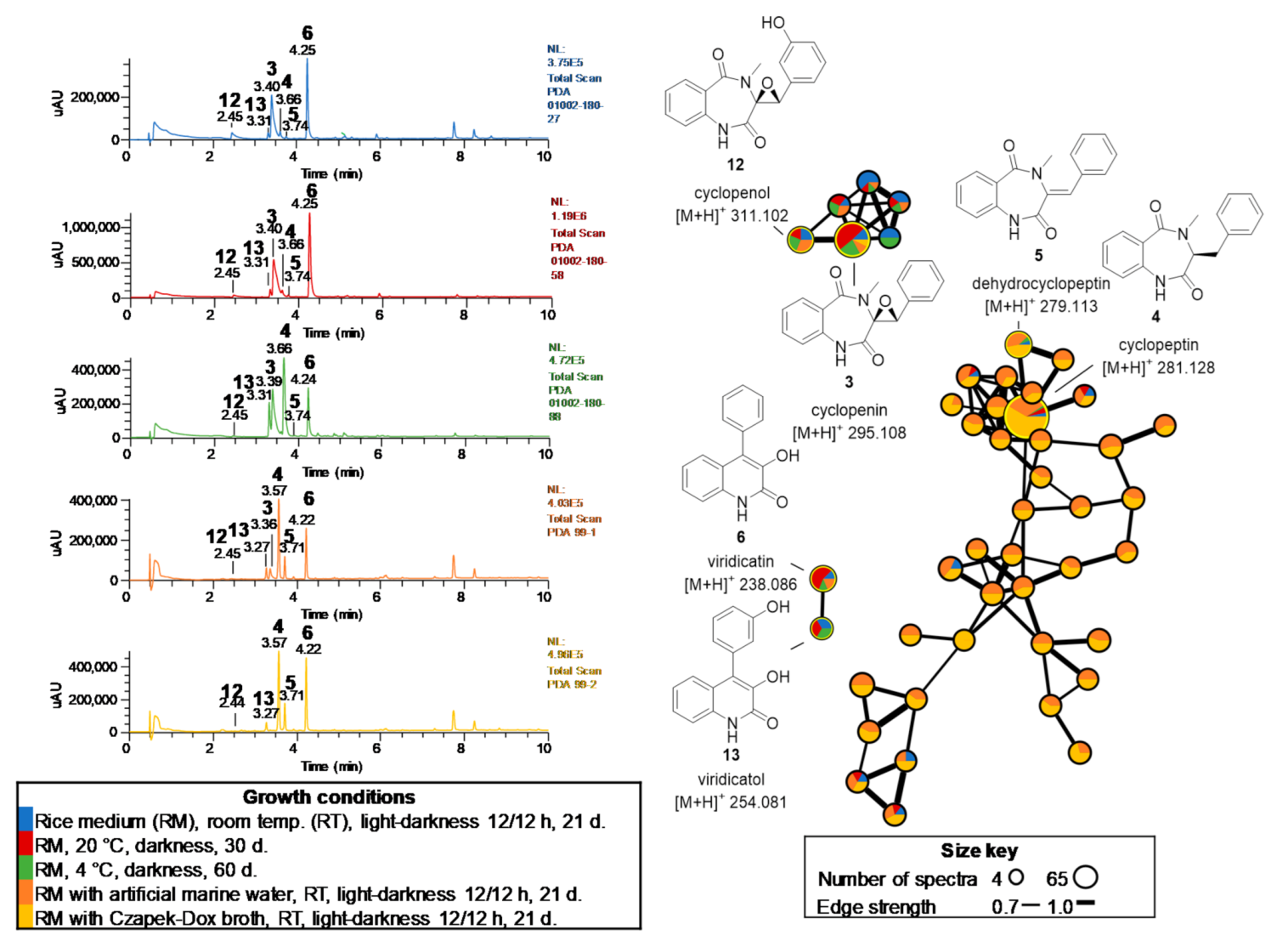
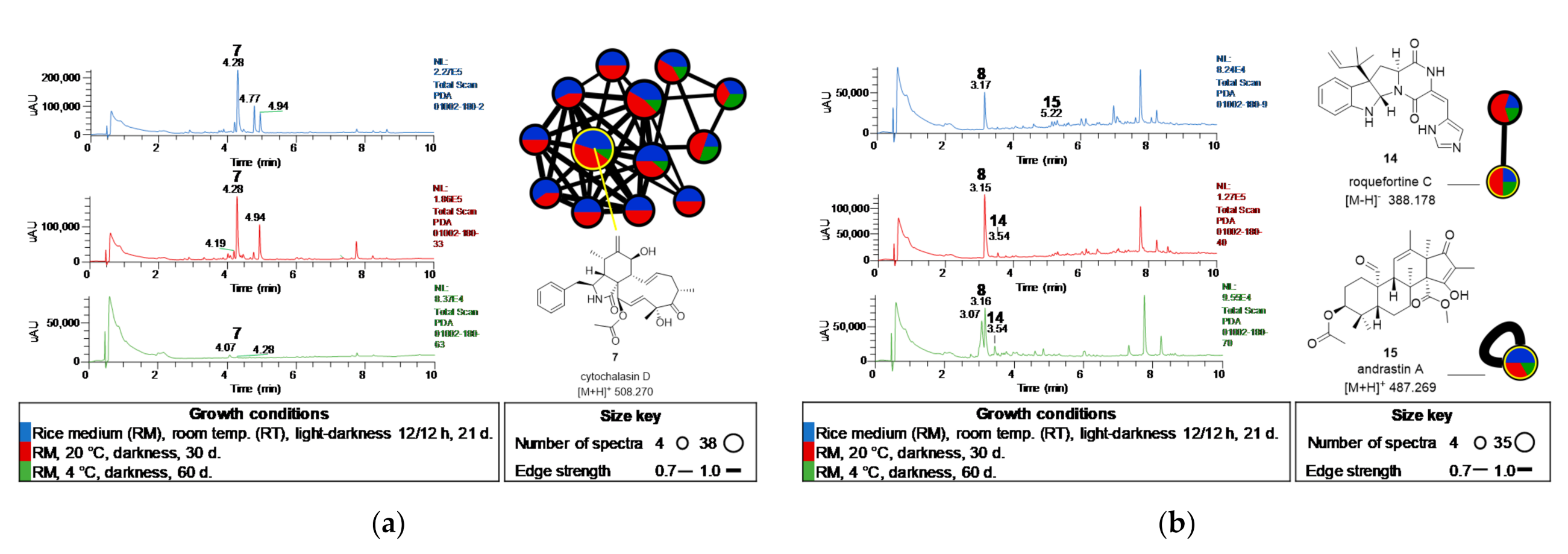
2.2. Chemical Study of Selected Fungal Strains
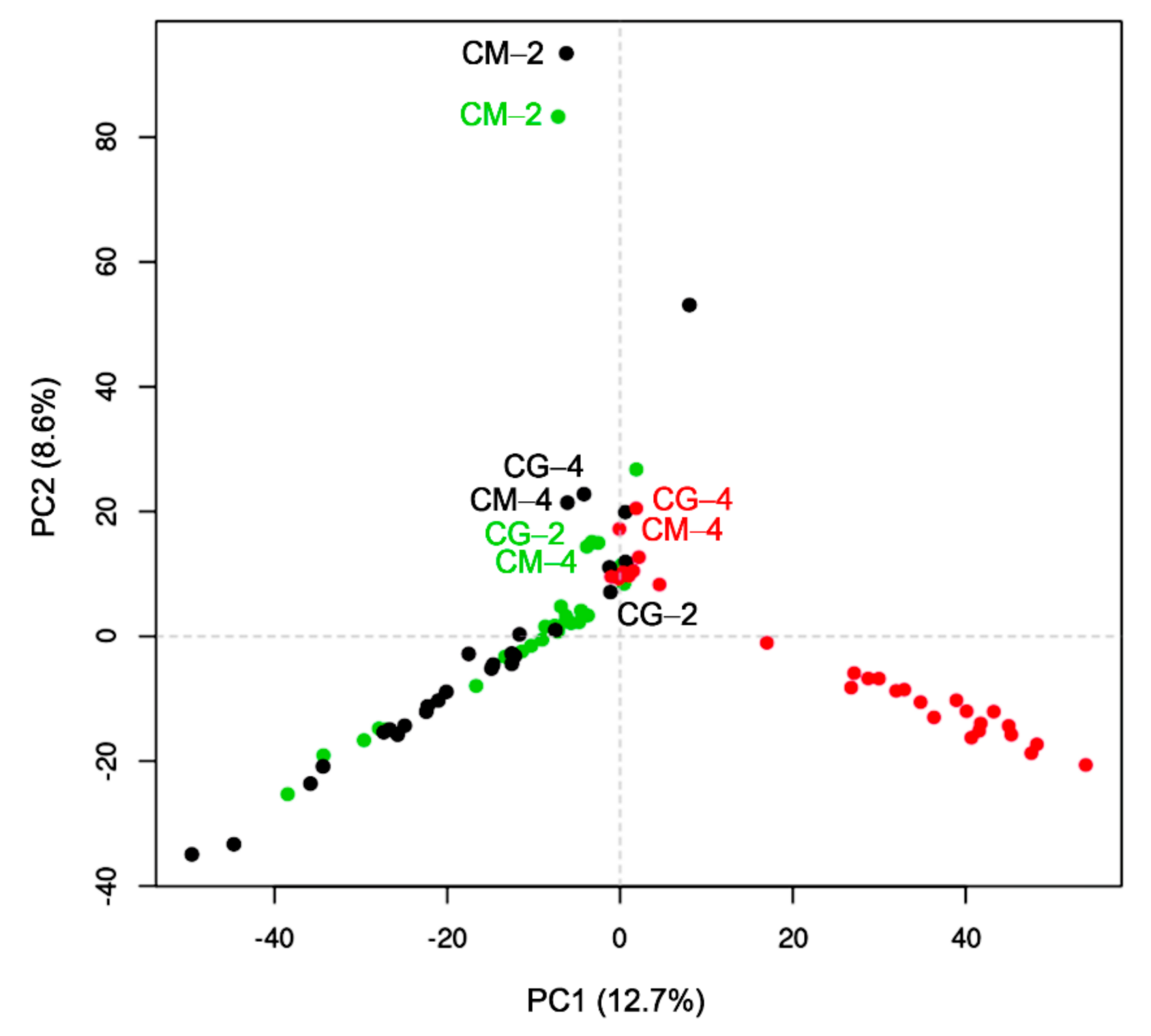
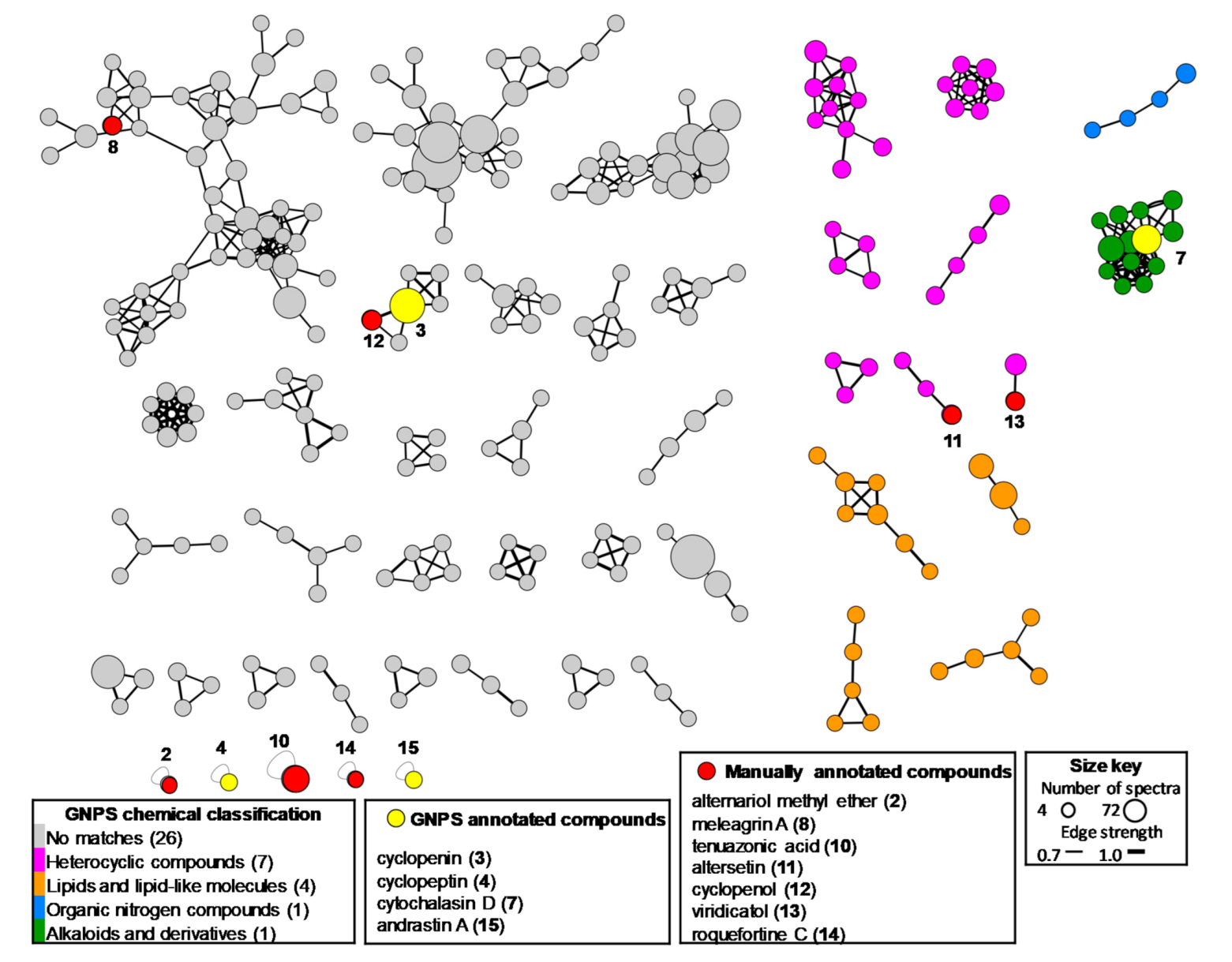
2.3. Metabolomic Study of Selected GoM Fungal Strains
2.4. Biological Activity of 1–8
3. Materials and Methods
3.1. Strains, Cultures, and Extract Preparations
3.2. In Vitro Biological Testing
3.3. Chemical Study of Selected Fungal Strains
3.4. LC-MS/MS, Untargeted Metabolomic and Molecular Network Analysis
3.5. Data Availability
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Sample Availability
References
- Dash, H.R.; Das, S. Chapter 4. Molecular methods for studying microorganisms from atypical environments. In Methods in Microbiology; Gurtler, V., Trevors, J.T., Eds.; Academic Press: Cambridge, MA, USA, 2018; Volume 45, pp. 89–122. [Google Scholar]
- Salazar, G.; Sunagawa, S. Marine microbial diversity. Curr. Biol. 2017, 27, R489–R494. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Newman, D.J.; Cragg, G.M. Natural products as sources of new drugs over the nearly four decades from 01/1981 to 09/2019. J. Nat. Prod. 2020, 83, 770–803. [Google Scholar] [CrossRef] [PubMed]
- Gladfelter, A.S.; James, T.Y.; Amend, A.S. Marine fungi. Curr. Biol. 2019, 29, R191–R195. [Google Scholar] [CrossRef] [PubMed]
- Rédou, V.; Vallet, M.; Meslet-Cladière, L.; Kumar, A.; Pang, K.-L.; Pouchus, Y.-F.; Barbier, G.; Grovel, O.; Bertrand, S.; Prado, S.; et al. Marine Fungi. In The Marine Microbiome: An Untapped Source of Biodiversity and Biotechnological Potential; Stal, L.J., Cretoiu, M.S., Eds.; Springer International Publishing: Cham, Switzerland, 2016; pp. 99–153. [Google Scholar]
- Poli, A.; Finore, I.; Romano, I.; Gioiello, A.; Lama, L.; Nicolaus, B. Microbial diversity in extreme marine habitats and their biomolecules. Microorganisms 2017, 5, 25. [Google Scholar] [CrossRef] [Green Version]
- Amend, A.; Burgaud, G.; Cunliffe, M.; Edgcomb, V.P.; Ettinger, C.L.; Gutiérrez, M.H.; Heitman, J.; Hom, E.F.Y.; Ianiri, G.; Jones, A.C.; et al. Fungi in the marine environment: Open questions and unsolved problems. mBio 2019, 10, e01189-18. [Google Scholar] [CrossRef] [Green Version]
- Vargas-Gastélum, L.; Chong-Robles, J.; Lago-Lestón, A.; Darcy, J.L.; Amend, A.S.; Riquelme, M. Targeted ITS1 sequencing unravels the mycodiversity of deep-sea sediments from the Gulf of Mexico. Environ. Microbiol. 2019, 21, 4046–4061. [Google Scholar] [CrossRef]
- Kimes, N.; Callaghan, A.; Aktas, D.; Smith, W.; Sunner, J.; Golding, B.; Drozdowska, M.; Hazen, T.; Suflita, J.; Morris, P. Metagenomic analysis and metabolite profiling of deep–sea sediments from the Gulf of Mexico following the deepwater horizon oil spill. Front. Microbiol. 2013, 4, 50. [Google Scholar] [CrossRef] [Green Version]
- King, G.M.; Kostka, J.E.; Hazen, T.C.; Sobecky, P.A. Microbial responses to the Deepwater Horizon oil spill: From coastal wetlands to the deep sea. Ann. Rev. Mar. Sci. 2015, 7, 377–401. [Google Scholar] [CrossRef] [Green Version]
- Dombrowski, N.; Donaho, J.A.; Gutierrez, T.; Seitz, K.W.; Teske, A.P.; Baker, B.J. Reconstructing metabolic pathways of hydrocarbon-degrading bacteria from the Deepwater Horizon oil spill. Nat. Microbiol. 2016, 1, 16057. [Google Scholar] [CrossRef]
- Bacosa, H.P.; Erdner, D.L.; Rosenheim, B.E.; Shetty, P.; Seitz, K.W.; Baker, B.J.; Liu, Z. Hydrocarbon degradation and response of seafloor sediment bacterial community in the northern Gulf of Mexico to light Louisiana sweet crude oil. ISME J. 2018, 12, 2532–2543. [Google Scholar] [CrossRef]
- Noirungsee, N.; Hackbusch, S.; Viamonte, J.; Bubenheim, P.; Liese, A.; Müller, R. Influence of oil, dispersant, and pressure on microbial communities from the Gulf of Mexico. Sci. Rep. 2020, 10, 7079. [Google Scholar] [CrossRef]
- Ramírez, D.; Vega-Alvarado, L.; Taboada, B.; Estradas-Romero, A.; Soto, L.; Juárez, K. Bacterial diversity in surface sediments from the continental shelf and slope of the North West Gulf of Mexico and the presence of hydrocarbon degrading bacteria. Mar. Pollut. Bull. 2020, 150, 110590. [Google Scholar] [CrossRef]
- Cerqueda-García, D.; García-Maldonado, J.Q.; Aguirre-Macedo, L.; García-Cruz, U. A succession of marine bacterial communities in batch reactor experiments during the degradation of five different petroleum types. Mar. Pollut. Bull. 2020, 150, 110775. [Google Scholar] [CrossRef]
- Escobedo-Hinojosa, W.; Pardo-López, L. Analysis of bacterial metagenomes from the Southwestern Gulf of Mexico for pathogens detection. Pathog. Dis. 2017, 75, ftx058. [Google Scholar] [CrossRef]
- Velez, P.; Gasca-Pineda, J.; Riquelme, M. Cultivable fungi from deep-sea oil reserves in the Gulf of Mexico: Genetic signatures in response to hydrocarbons. Mar. Environ. Res. 2020, 153, 104816. [Google Scholar] [CrossRef]
- Romero-Hernández, L.; Velez, P.; Betanzo-Gutiérrez, I.; Camacho-López, M.D.; Vázquez-Duhalt, R.; Riquelme, M. Extra-heavy crude oil degradation by Alternaria sp. isolated from deep-sea sediments of the Gulf of Mexico. Appl. Sci. 2021, 11, 6090. [Google Scholar] [CrossRef]
- Godoy-Lozano, E.E.; Escobar-Zepeda, A.; Raggi, L.; Merino, E.; Gutierrez-Rios, R.M.; Juarez, K.; Segovia, L.; Licea-Navarro, A.F.; Gracia, A.; Sanchez-Flores, A.; et al. Bacterial diversity and the geochemical landscape in the southwestern Gulf of Mexico. Front. Microbiol. 2018, 9, 2528. [Google Scholar] [CrossRef]
- Bode, H.B.; Bethe, B.; Höfs, R.; Zeeck, A. Big effects from small changes: Possible ways to explore nature’s chemical diversity. Chembiochem 2002, 3, 619–627. [Google Scholar] [CrossRef]
- Wang, M.; Carver, J.J.; Phelan, V.V.; Sanchez, L.M.; Garg, N.; Peng, Y.; Nguyen, D.D.; Watrous, J.; Kapono, C.A.; Luzzatto-Knaan, T.; et al. Sharing and community curation of mass spectrometry data with Global Natural Products Social Molecular Networking. Nat. Biotechnol. 2016, 34, 828–837. [Google Scholar] [CrossRef] [Green Version]
- Aly, A.H.; Edrada-Ebel, R.; Indriani, I.D.; Wray, V.; Müller, W.E.; Totzke, F.; Zirrgiebel, U.; Schächtele, C.; Kubbutat, M.H.; Lin, W. Cytotoxic metabolites from the fungal endophyte Alternaria sp. and their subsequent detection in its host plant Polygonum senegalense. J. Nat. Prod. 2008, 71, 972–980. [Google Scholar] [CrossRef]
- Tan, N.; Tao, Y.; Pan, J.; Wang, S.; Xu, F.; She, Z.; Lin, Y.; Gareth Jones, E.B. Isolation, structure elucidation, and mutagenicity of four alternariol derivatives produced by the mangrove endophytic fungus No. 2240. Chem. Nat. Compd. 2008, 44, 296–300. [Google Scholar] [CrossRef]
- Kusano, M.; Koshino, H.; Uzawa, J.; Fujioka, S.; Kawano, T.; Kimura, Y. Nematicidal alkaloids and related compounds produced by the fungus Penicillium cf. simplicissimum. Biosci. Biotechnol. Biochem. 2000, 64, 2559–2568. [Google Scholar] [CrossRef]
- Abdel-Hadi, A.; Awad, M.; Abo-Dahab, N.; El-Shanawany, A.; Abdelkader, M.; Abo-Kadoum, M.V. Viridicatin and dehydrocyclopeptine, two bioactive alkaloids isolated from marine derived fungus Penicillium aurantiogriseum AUMC 9759. IBSPR 2015, 3, 115–122. [Google Scholar] [CrossRef]
- Kishimoto, S.; Hara, K.; Hashimoto, H.; Hirayama, Y.; Champagne, P.A.; Houk, K.N.; Tang, Y.; Watanabe, K. Enzymatic one-step ring contraction for quinolone biosynthesis. Nat. Commun. 2018, 9, 2826. [Google Scholar] [CrossRef]
- Jikai, L.; Jianwen, T.; Zejun, D.; Zhihui, D.; Xianghua, W.; Peigui, L. Neoengleromycin, a novel compound from Engleromyces goetzii. Helv. Chim. Acta 2002, 85, 1439–1442. [Google Scholar] [CrossRef]
- Kawai, K.; Nozawa, K.; Nakajima, S.; Iitaka, Y. Studies on fungal products. VII. The structures of meleagrin and 9-Op-bromobenzoylmeleagrin. Chem. Pharm. Bull. 1984, 32, 94–98. [Google Scholar] [CrossRef] [Green Version]
- Sumner, L.W.; Amberg, A.; Barrett, D.; Beale, M.H.; Beger, R.; Daykin, C.A.; Fan, T.W.-M.; Fiehn, O.; Goodacre, R.; Griffin, J.L.; et al. Proposed minimum reporting standards for chemical analysis: Chemical Analysis Working Group (CAWG) Metabolomics Standards Initiative (MSI). Metabolomics 2007, 3, 211–221. [Google Scholar] [CrossRef] [Green Version]
- El-Elimat, T.; Figueroa, M.; Ehrmann, B.M.; Cech, N.B.; Pearce, C.J.; Oberlies, N.H. High-resolution MS, MS/MS, and UV database of fungal secondary metabolites as a dereplication protocol for bioactive natural products. J. Nat. Prod. 2013, 76, 1709–1716. [Google Scholar] [CrossRef] [Green Version]
- Romano, S.; Jackson, S.A.; Patry, S.; Dobson, A.D.W. Extending the “One Strain Many Compounds” (OSMAC) principle to marine microorganisms. Mar. Drugs 2018, 16, 244. [Google Scholar] [CrossRef] [Green Version]
- Balouiri, M.; Sadiki, M.; Ibnsouda, S.K. Methods for in vitro evaluating antimicrobial activity: A review. J. Pharm. Anal. 2016, 6, 71–79. [Google Scholar] [CrossRef] [Green Version]
- Cho, S.; Lee, H.S.; Franzblau, S. Microplate Alamar Blue Assay (MABA) and Low Oxygen Recovery Assay (LORA) for Mycobacterium tuberculosis. In Mycobacteria Protocols; Parish, T., Roberts, D.M., Eds.; Springer: New York, NY, USA, 2015; pp. 281–292. [Google Scholar]
- Gao, W.; Kim, J.-Y.; Anderson, J.R.; Akopian, T.; Hong, S.; Jin, Y.-Y.; Kandror, O.; Kim, J.-W.; Lee, I.-A.; Lee, S.-Y.; et al. The cyclic peptide ecumicin targeting ClpC1 is active against Mycobacterium tuberculosis in vivo. Antimicrob. Agents Chemother. 2015, 59, 880–889. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhou, B.; Shetye, G.; Yu, Y.; Santarsiero, B.D.; Klein, L.L.; Abad-Zapatero, C.; Wolf, N.M.; Cheng, J.; Jin, Y.; Lee, H.; et al. Antimycobacterial rufomycin analogues from Streptomyces atratus strain MJM3502. J. Nat. Prod. 2020, 83, 657–667. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, Y.; Yang, M.-H.; Wang, X.-B.; Li, T.-X.; Kong, L.-Y. Bioactive metabolites from the endophytic fungus Alternaria alternata. Fitoterapia 2014, 99, 153–158. [Google Scholar] [CrossRef] [PubMed]
- Zhang, C.-C.; Ding, S.-S.; Shi, W.-S.; Cao, F.; Zhu, H.-J.; Wen, M.-L. A new quinolinone from freshwater lake-derived fungus Myrothecium verrucaria. Nat. Prod. Res. 2017, 31, 99–103. [Google Scholar] [CrossRef]
- Cunningham, K.G.; Freeman, G.G. The isolation and some chemical properties of viridicatin, a metabolic product of Penicillium viridicatum Westling. Biochem. J. 1953, 53, 328–332. [Google Scholar] [CrossRef] [Green Version]
- Hamed, A.; Abdel-Razek, A.S.; Araby, M.; Abu-Elghait, M.; El-Hosari, D.G.; Frese, M.; Soliman, H.S.; Stammler, H.G.; Sewald, N.; Shaaban, M. Meleagrin from marine fungus Emericella dentata Nq45: Crystal structure and diverse biological activity studies. Nat. Prod. Res. 2021, 35, 3830–3838. [Google Scholar] [CrossRef]
- Zheng, C.J.; Sohn, M.-J.; Lee, S.; Kim, W.-G. Meleagrin, a new FabI inhibitor from Penicillium chryosogenum with at least one additional mode of action. PLoS ONE 2013, 8, e78922. [Google Scholar] [CrossRef] [Green Version]
- Aparicio-Cuevas, M.A.; Rivero-Cruz, I.; Sánchez-Castellanos, M.; Menéndez, D.; Raja, H.A.; Joseph-Nathan, P.; González, M.d.C.; Figueroa, M. Dioxomorpholines and derivatives from a marine-facultative Aspergillus species. J. Nat. Prod. 2017, 80, 2311–2318. [Google Scholar] [CrossRef] [Green Version]
- VanderMolen, K.M.; Raja, H.A.; El-Elimat, T.; Oberlies, N.H. Evaluation of culture media for the production of secondary metabolites in a natural products screening program. AMB Express 2013, 3, 71. [Google Scholar] [CrossRef] [Green Version]
- Lê, S.; Josse, J.; Husson, F. FactoMineR: An R package for multivariate analysis. J. Stat. Softw. 2015, 25, 1–18. [Google Scholar] [CrossRef] [Green Version]
- Ernst, M.; Kang, K.B.; Caraballo-Rodríguez, A.M.; Nothias, L.-F.; Wandy, J.; Chen, C.; Wang, M.; Rogers, S.; Medema, M.H.; Dorrestein, P.C.; et al. MolNetEnhancer: Enhanced molecular networks by integrating metabolome mining and annotation tools. Metabolites 2019, 9, 144. [Google Scholar] [CrossRef] [Green Version]
- Shannon, P.; Markiel, A.; Ozier, O.; Baliga, N.S.; Wang, J.T.; Ramage, D.; Amin, N.; Schwikowski, B.; Ideker, T. Cytoscape: A software environment for integrated models of biomolecular interaction networks. Genome Res. 2003, 13, 2498–2504. [Google Scholar] [CrossRef]
- Dictionary of Natural Products 30.1. Available online: https://dnp.chemnetbase.com/faces/chemical/ChemicalSearch.xhtml (accessed on 26 November 2021).
- SciFinder—CAS. Available online: https://scifinder.cas.org (accessed on 26 November 2021).








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
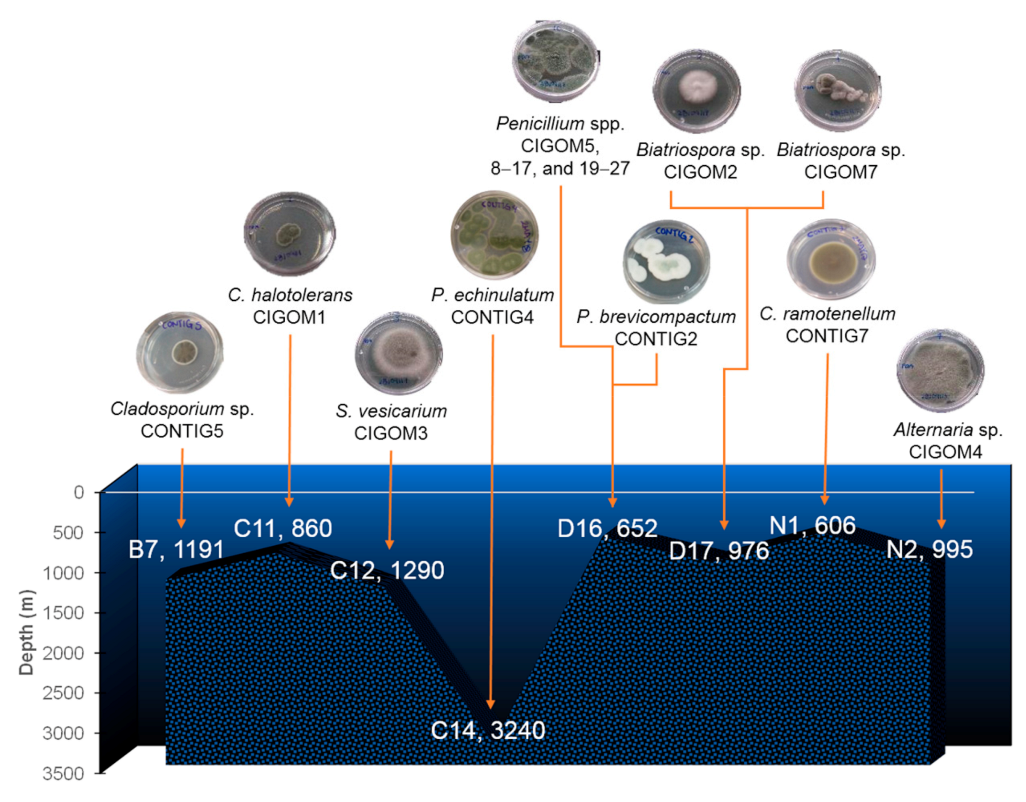
| Station | Depth (m) | Strain |
|---|---|---|
| B7 (Perdido) | 1191 | Cladosporium sp. CONTIG5 |
| C11 (Coatzacoalcos) | 860 | Cladosporium halotolerans CIGOM1 |
| C12 (Coatzacoalcos) | 1290 | Stemphylium vesicarium CIGOM3 |
| C14 (Coatzacoalcos) | 3240 | Penicillium echinulatum CONTIG4 |
| D16 (Coatzacoalcos) | 652 | Penicillium spp. CIGOM5, 8–17, and 19–27 P. brevicompactum CONTIG2 |
| D17 (Coatzacoalcos) | 976 | Biatriospora sp. CIGOM2 y CIGOM7 |
| N1 (Perdido) | 606 | Cladosporium ramotenellum CONTIG7 |
| N2 (Perdido) | 995 | Alternaria sp. CIGOM4 |
| Strain | Extract Condition | E. coli ATCC 10536 | S. typhi ATCC 9992v | MSSA | B. subtilis ATCC 6633 |
|---|---|---|---|---|---|
| P.echinulatum CONTIG4 | B | ||||
| C. ramotenellum CONTIG7 | A B | ||||
| S. vesicarium CIGOM3 | C | ||||
| Alternaria sp. CIGOM4 | B | ||||
| C | |||||
| Biatriospora sp. CIGOM7 | C | ||||
| Penicillium sp. CIGOM16 | C | ||||
| Penicillium sp. CIGOM17 | C | ||||
| Penicillium sp. CIGOM20 | C | ||||
| Penicillium sp. CIGOM21 | C | ||||
| Penicillium sp. CIGOM22 | C | ||||
| Penicillium sp. CIGOM24 | A | ||||
| Penicillium sp. CIGOM26 | A | ||||
| C | |||||
| MIC ampicillin (μg/mL) | 1.6 | 0.4 | 0.1 | 16.3 | |
| Inhibition level | |||||
| Total inhibition at 200 μg/mL and 20 μg/mL. | |||||
| Total inhibition at 200 μg/mL. | |||||
| Partial inhibition at 200 μg/mL and 20 μg/mL. | |||||
| Partial inhibition at 200 μg/mL. | |||||
| No inhibition. | |||||
| Compound | Adduct | Observed Ion a | Molecular Formula | Exact Mass d | Mass Accuracy (ppm) |
|---|---|---|---|---|---|
| Alternariol (1) | [M − H]− | 257.045 | C14H10O5 | 257.0452 | −1.3 |
| Alternariol methyl ether (2) | [M + H]+ | 273.076 | C15H12O5 | 273.0755 | −0.9 |
| Cyclopenin (3) | [M + H]+ | 295.108 | C17H14N2O3 | 295.1074 | −1.10 |
| Cyclopeptin (4) | [M + H]+ | 281.128 | C17H16N2O2 | 281.1282 | −0.9 |
| Dehydrocyclopeptin (5) | [M + H]+ | 279.113 | C17H14N2O2 | 279.1126 | −0.7 |
| Viridicatin (6) | [M + H]+ | 238.086 | C15H11NO2 | 238.0860 | −1.1 |
| Cytochalasin D (7) | [M + H]+ | 508.270 | C30H37NO6 | 508.2689 | −0.7 |
| Meleagrin A (8) | [M + H]+ | 434.182 | C23H23N5O4 | 434.1820 | −0.6 |
| Altenuene (9) b | [M + H]+ | 293.102 | C15H16O6 | 293.1017 | −0.9 |
| Tenuazonic acid (10) b | [M + H]+ | 198.113 | C10H15NO3 | 198.1123 | −0.9 |
| Altersetin (11) b | [M + H]+ | 400.248 | C24H33NO4 | 400.2479 | −0.8 |
| Cyclopenol (12) b | [M + H]+ | 311.102 | C17H14N2O4 | 311.1023 | −1.1 |
| Viridicatol (13) b | [M + H]+ | 254.081 | C15H11NO3 | 254.0811 | −0.3 |
| Roquefortine C (14) b | [M − H]− | 388.178 | C22H23N5O2 | 388.1781 | +0.5 |
| Andrastin A (15) c | [M + H]+ | 487.269 | C28H38O7 | 487.2689 | −3.0 |
| Compound | MSSA | MRSA | M. tuberculosis H37Rv | M avium ATCC 15769 | Vero Cell ATCC CCL-81 6 | |
|---|---|---|---|---|---|---|
| MABA 4 | LORA 5 | MABA 4 | ||||
| Alternariol (1) | 87 (ND) | >50 | ND | ND | ||
| Viridicatin (6) | 100 (43.8) | >50 | >50 | >50 | ||
| Meleagrin A (8) | 101 (48.0) | >50 | 12.3 | >50 | ||
| MIC positive control (μM) | 572.41 1 | 0.86 2 | 100 (0.03) 3 | 0.08 3 | 0.05 3 | >100 3 |
| Inhibition level | ||||||
| Total inhibition at 100 μg/mL and partial at 10 μg/mL. | ||||||
| Total inhibition at 100 μg/mL. | ||||||
| Partial inhibition at 100 μg/mL. | ||||||
| No inhibition. | ||||||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Villanueva-Silva, R.; Velez, P.; Riquelme, M.; Fajardo-Hernández, C.A.; Martínez-Cárdenas, A.; Arista-Romero, A.; Wan, B.; Ma, R.; Qader, M.; Franzblau, S.G.; et al. Chemical Diversity and Antimicrobial Potential of Cultivable Fungi from Deep-Sea Sediments of the Gulf of Mexico. Molecules 2021, 26, 7328. https://doi.org/10.3390/molecules26237328
Villanueva-Silva R, Velez P, Riquelme M, Fajardo-Hernández CA, Martínez-Cárdenas A, Arista-Romero A, Wan B, Ma R, Qader M, Franzblau SG, et al. Chemical Diversity and Antimicrobial Potential of Cultivable Fungi from Deep-Sea Sediments of the Gulf of Mexico. Molecules. 2021; 26(23):7328. https://doi.org/10.3390/molecules26237328
Chicago/Turabian StyleVillanueva-Silva, Rodrigo, Patricia Velez, Meritxell Riquelme, Carlos A. Fajardo-Hernández, Anahí Martínez-Cárdenas, Alejandra Arista-Romero, Baojie Wan, Rui Ma, Mallique Qader, Scott G. Franzblau, and et al. 2021. "Chemical Diversity and Antimicrobial Potential of Cultivable Fungi from Deep-Sea Sediments of the Gulf of Mexico" Molecules 26, no. 23: 7328. https://doi.org/10.3390/molecules26237328
APA StyleVillanueva-Silva, R., Velez, P., Riquelme, M., Fajardo-Hernández, C. A., Martínez-Cárdenas, A., Arista-Romero, A., Wan, B., Ma, R., Qader, M., Franzblau, S. G., & Figueroa, M. (2021). Chemical Diversity and Antimicrobial Potential of Cultivable Fungi from Deep-Sea Sediments of the Gulf of Mexico. Molecules, 26(23), 7328. https://doi.org/10.3390/molecules26237328