
Lentzeacins A-E, New Bacterial-Derived 2,5- and 2,6-Disubstituted Pyrazines from a BGC-Rich Soil Bacterium Lentzea sp. GA3-008

Abstract
1. Introduction
2. Results
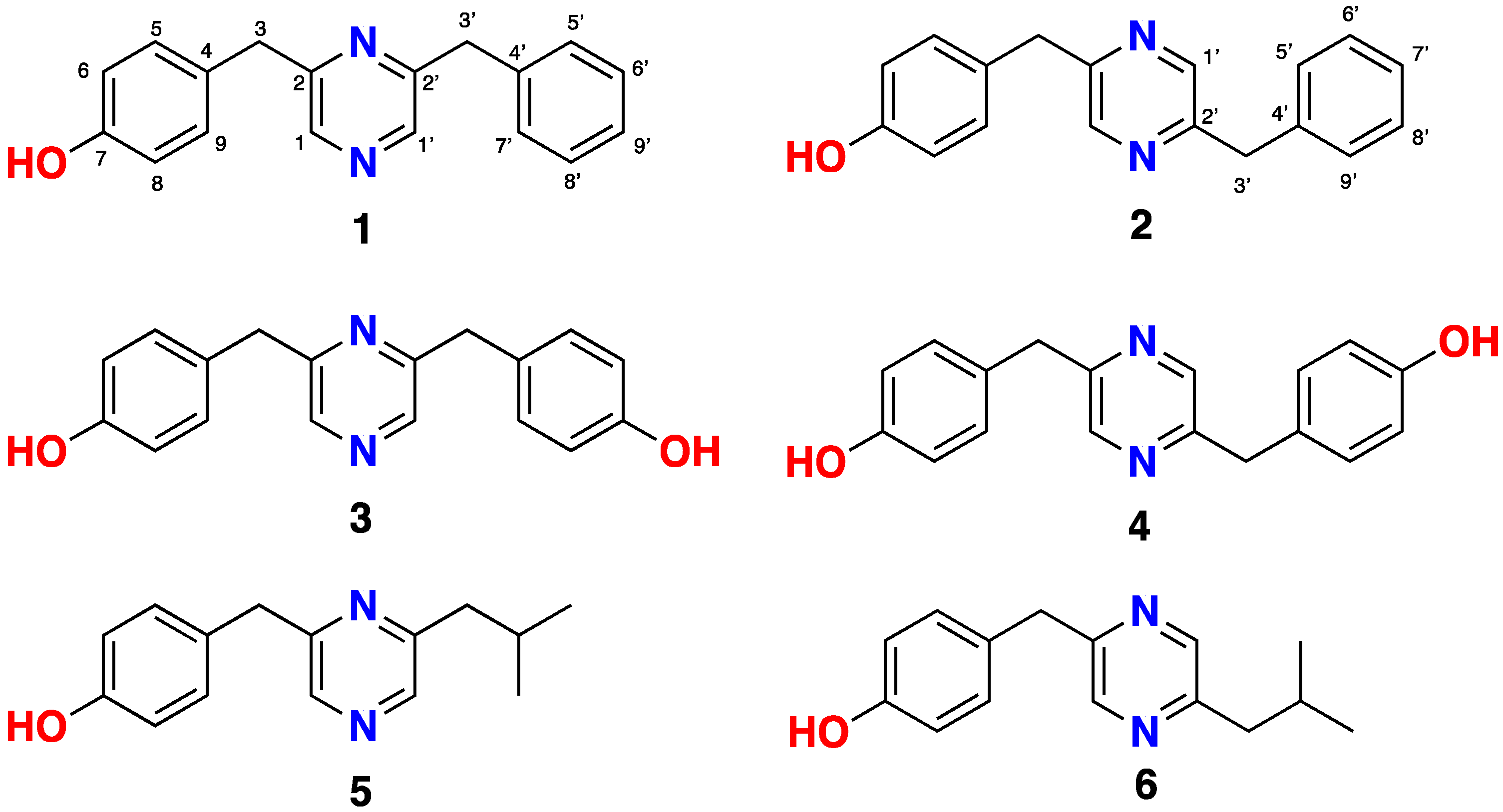
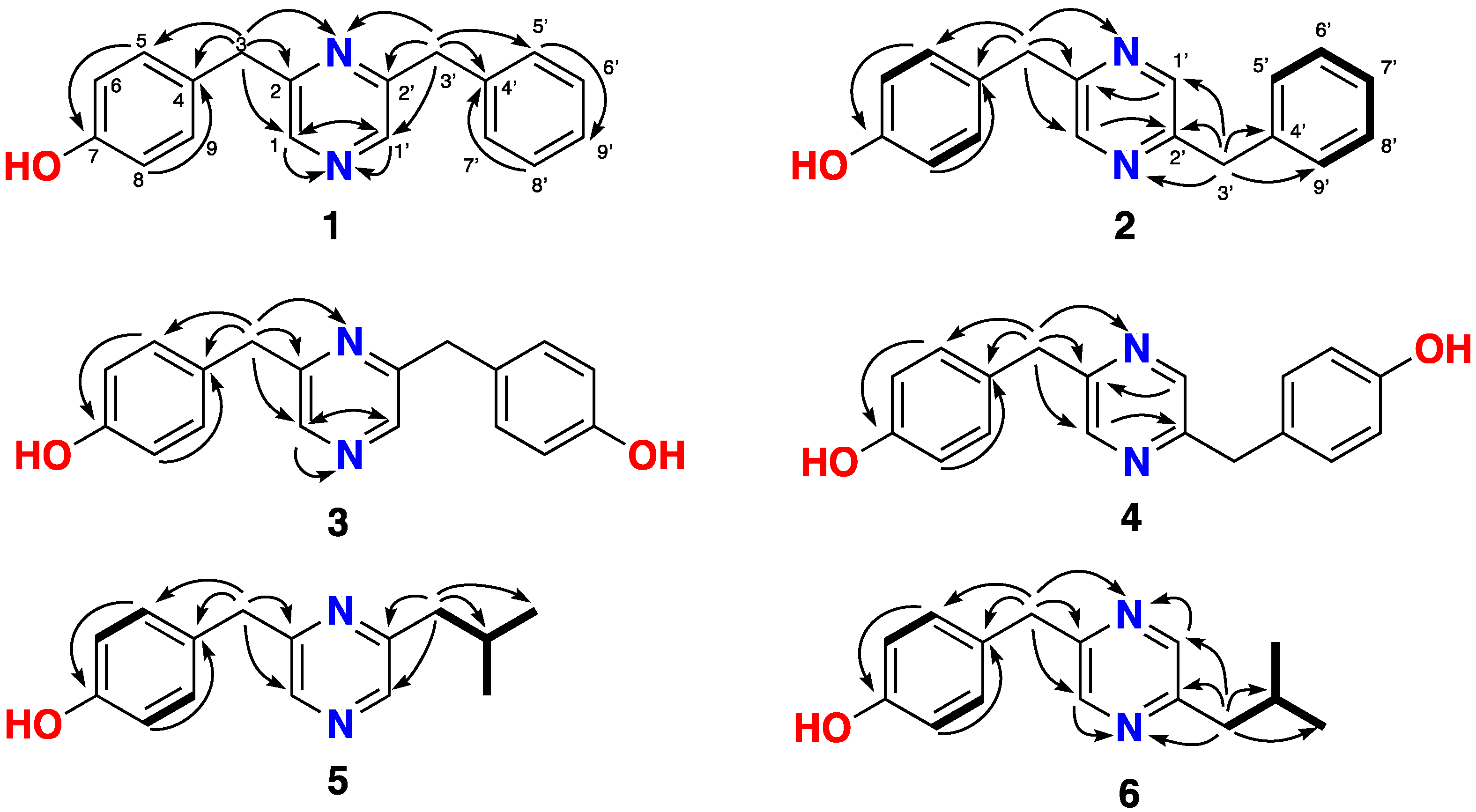
2.1. Isolation and Structure Determination of Lentzeacins and Zincphyrin
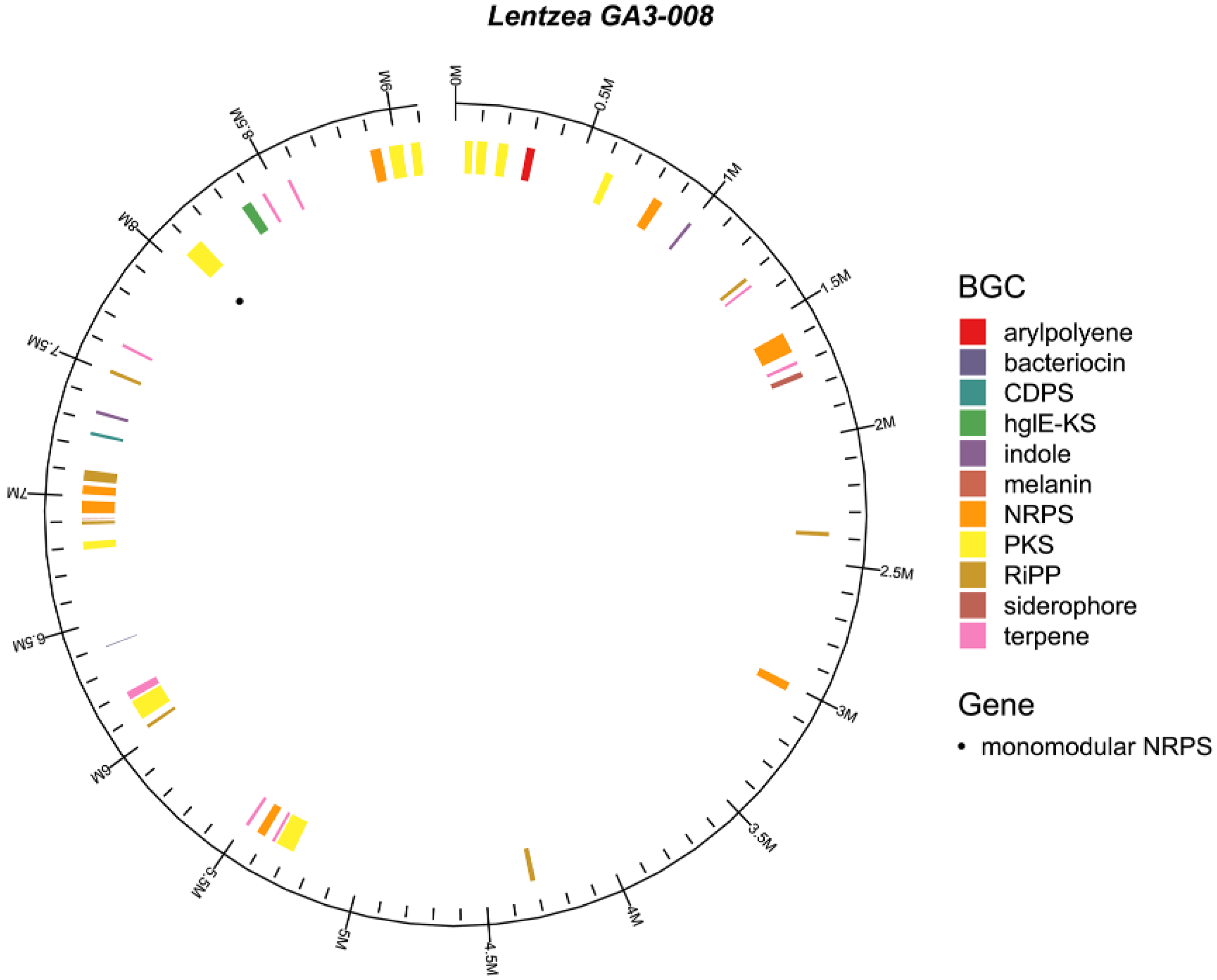
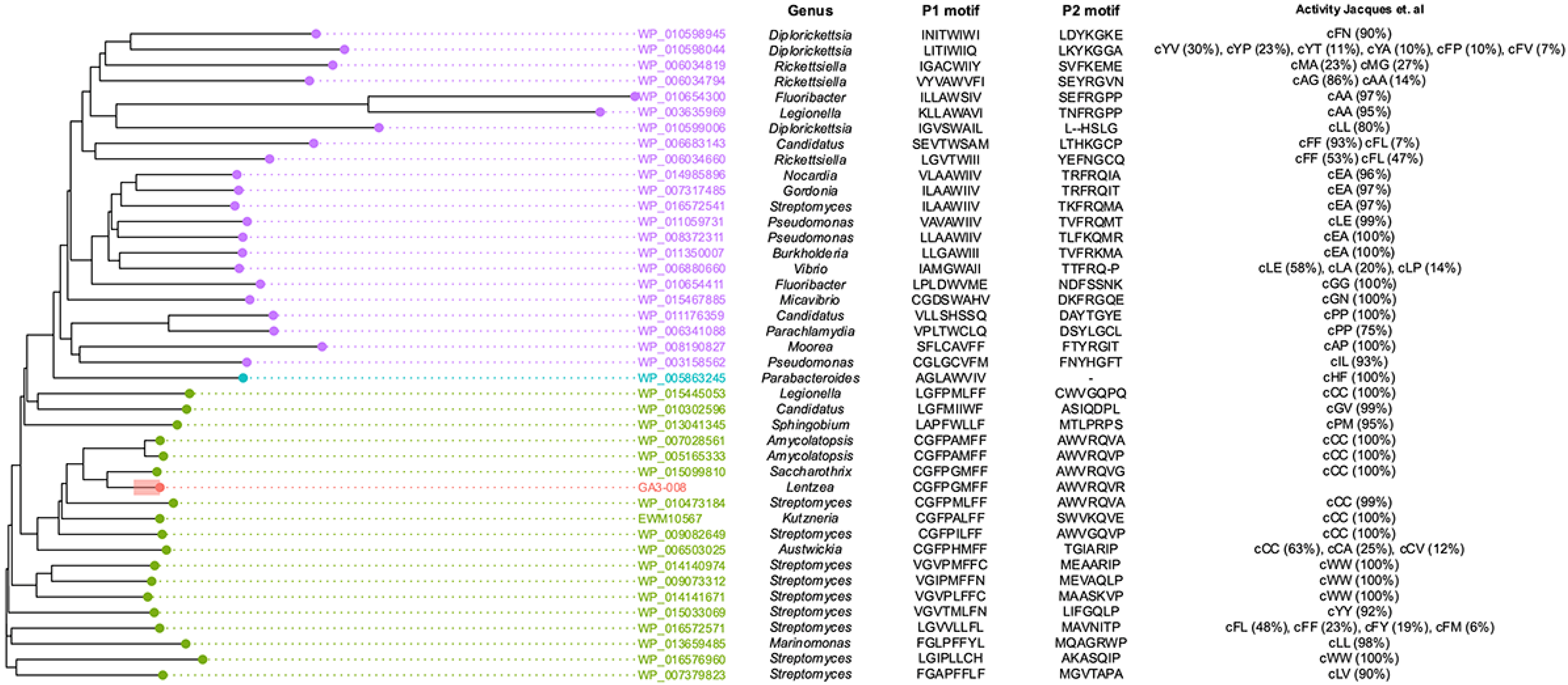
2.2. Analysis of the GA3-008 Genome for Pyrazine Biosynthesis
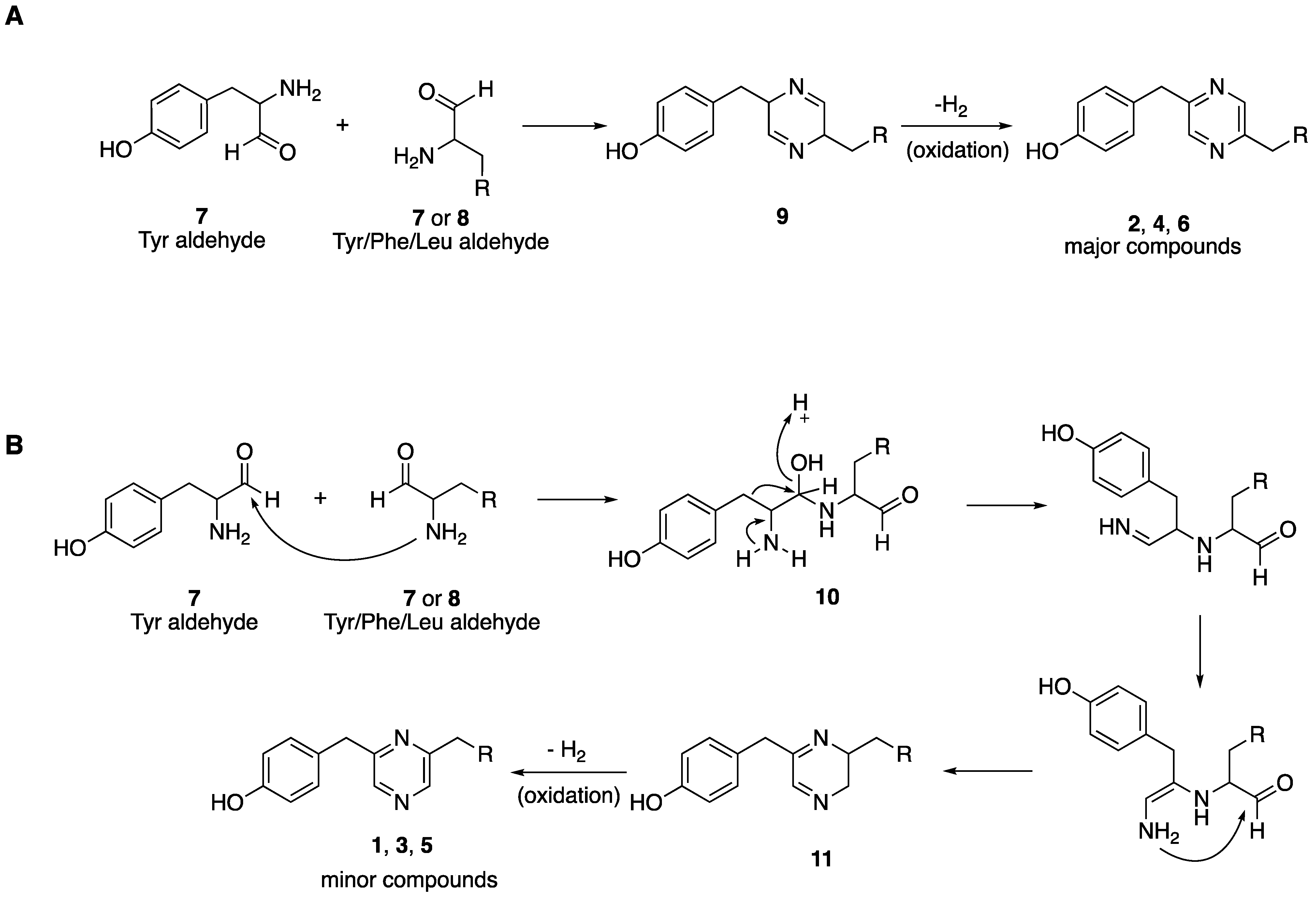
3. Discussion
4. Materials and Methods
4.1. General Experimental Procedures
4.2. Isolation of Lentzea sp. GA3-008
4.3. Solid Agar Screening for Antimicrobial Activity of Actinomycete Colonies
4.4. Genome Sequencing
4.5. Fermentation, Extraction and Isolation
4.6. Antimicrobial Assays
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Mortzfeld, F.B.; Hashem, C.; Vranková, K.; Winkler, M.; Rudroff, F. Pyrazines: Synthesis and Industrial Application of These Valuable Flavor and Fragrance Compounds. Biotechnol. J. 2020, 15, 2000064. [Google Scholar] [CrossRef]
- Barlin, G.B. Chemistry of Heterocyclic Compounds: The Pyrazines.; John Wiley and Sons, Inc.: New York, NY, USA, 1982; Volume 41, pp. 1–10. [Google Scholar]
- Mattey, M.; Harle, E.M. Aerobic Metabolism of Pyrazine Compounds by a Pseudomonas Species. Biochem. Soc. Trans. 1976, 4, 492–494. [Google Scholar] [CrossRef][Green Version]
- Müller, R.; Rappert, S. Pyrazines: Occurrence, formation and biodegradation. Appl. Microbiol. Biotechnol. 2010, 85, 1315–1320. [Google Scholar] [CrossRef]
- Adams, J.; Kauffman, M. Development of the Proteasome Inhibitor Velcade™ (Bortezomib). Cancer Investig. 2004, 22, 304–311. [Google Scholar] [CrossRef]
- Goetz, M.A.; Zhang, C.; Zink, D.L.; Arocho, M.; Vicente, F.; Bills, G.F.; Polishook, J.; Dorso, K.; Onishi, R.; Gill, C.; et al. Coelomycin, a highly substituted 2,6-dioxo-pyrazine fungal metabolite antibacterial agent discovered by Staphylococcus aureus fitness test profiling. J. Antibiot. 2010, 63, 512–518. [Google Scholar] [CrossRef] [PubMed]
- Belin, P.; Moutiez, M.; Lautru, S.; Seguin, J.; Pernodet, J.-L.; Gondry, M. The Nonribosomal Synthesis of Diketopiperazines in tRNA-Dependent Cyclodipeptide Synthase Pathways. Nat. Prod. Rep. 2012, 29, 961–979. [Google Scholar] [CrossRef]
- Borgman, P.; Lopez, R.D.; Lane, A.L. The Expanding Spectrum of Diketopiperazine Natural Product Biosynthetic Pathways Containing Cyclodipeptide Synthases. Org. Biomol. Chem. 2019, 17, 2305–2314. [Google Scholar] [CrossRef] [PubMed]
- Skinnider, M.A.; Johnston, C.W.; Merwin, N.J.; Dejong, C.A.; Magarvey, N.A. Global Analysis of Prokaryotic tRNA-Derived Cyclodipeptide Biosynthesis. BMC Genom. 2018, 19, 45. [Google Scholar] [CrossRef]
- Johnson, E.G.; Krasnoff, S.B.; Bignell, D.R.; Chung, W.C.; Tao, T.; Parry, R.J.; Loria, R.; Gibson, D.M. 4-Nitrotryptophan is a Substrate for the Non-Ribosomal Peptide Synthetase Txtb in the Thaxtomin a Biosynthetic Pathway. Mol. Microbiol. 2009, 73, 409–418. [Google Scholar] [CrossRef]
- Balibar, C.J.; Walsh, C.T. GliP, a Multimodular Nonribosomal Peptide Synthetase in Aspergillus fumigatus, Makes the Diketopiperazine Scaffold of Gliotoxin. Biochemistry 2006, 45, 15029–15038. [Google Scholar] [CrossRef] [PubMed]
- Huang, S.-X.; Powell, E.; Rajski, S.R.; Zhao, L.-X.; Jiang, C.-L.; Duan, Y.; Xu, W.; Shen, B. Discovery and Total Synthesis of a New Estrogen Receptor Heterodimerizing Actinopolymorphol A from Actinopolymorpha rutilus. Org. Lett. 2010, 12, 3525–3527. [Google Scholar] [CrossRef]
- Yuan, B.; Liu, D.; Guan, X.; Yan, Y.; Zhang, J.; Zhang, Y.; Yang, D.; Ma, M.; Lin, W. Piperazine Ring Formation by a Single-Module NRPS and Cleavage by an A-Kg-Dependent Nonheme Iron Dioxygenase in Brasiliamide Biosynthesis. Appl. Microbiol. Biotechnol. 2020, 104, 6149–6159. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Lacey, A.E.; Shu, S.; Kalaitzis, J.A.; Vuong, D.; Crombie, A.; Hu, J.; Gilchrist, C.L.M.; Lacey, E.; Piggott, A.M.; et al. Hancockiamides: Phenylpropanoid Piperazines from Aspergillus hancockii are Biosynthesised by a Versatile Dual Single-module NRPS Pathway. Org. Biomol. Chem. 2020, 19, 587–595. [Google Scholar] [CrossRef]
- Guo, C.J.; Chang, F.Y.; Wyche, T.P.; Backus, K.M.; Acker, T.M.; Funabashi, M.; Taketani, M.; Donia, M.S.; Nayfach, S.; Pollard, K.S.; et al. Discovery of Reactive Microbiota-Derived Metabolites That Inhibit Host Proteases. Cell 2017, 168, 517–526. [Google Scholar] [CrossRef] [PubMed]
- Kim, C.S.; Gatsios, A.; Cuesta, S.; Lam, Y.C.; Wei, Z.; Chen, H.; Russell, R.M.; Shine, E.E.; Wang, R.; Wyche, T.P.; et al. Characterization of Autoinducer-3 Structure and Biosynthesis in E. coli. ACS Central Sci. 2020, 6, 197–206. [Google Scholar] [CrossRef] [PubMed]
- Wyatt, M.A.; Mok, M.C.Y.; Junop, M.; Magarvey, N.A. Heterologous Expression and Structural Characterisation of a Pyrazinone Natural Product Assembly Line. Chembiochem 2012, 13, 2408–2415. [Google Scholar] [CrossRef] [PubMed]
- Fill, T.P.; dos Santos, R.M.G.; Barisson, A.; Rodrigues-Filho, E.; Souza, A.Q. Co-Production of Bisphenylpropanoid Amides and Meroterpenes by an Endophytic Penicillium brasilianum Found in the Root Bark of Melia zedarach. Z. Naturforsch. C 2009, 64, 355–360. [Google Scholar] [CrossRef]
- Nawrath, T.; Dickschat, J.S.; Kunze, B.; Schulz, S. The Biosynthesis of Branched Dialkylpyrazines in Myxobacteria. Chem. Biodivers. 2010, 7, 2129–2144. [Google Scholar] [CrossRef] [PubMed]
- Toriya, M.; Yaginuma, S.; Murofushi, S.; Ogawa, K.; Muto, N.; Hayashi, M.; Matsumoto, K. Zincphyrin, a Novel Coproporphyrin III with Zinc from Streptomyces sp. J. Antibiot. 1993, 46, 196–200. [Google Scholar] [CrossRef]
- Abraham, R.J.; Hawkes, G.E.; Smith, K.M. The nuclear magnetic resonance spectra of porphyrins. Part VIII. The 13C nuclear magnetic resonance spectra of some porphyrins and metalloporphyrins. J. Chem. Soc. Perkin Trans. 2 1974, 627–634. [Google Scholar] [CrossRef]
- Wyatt, M.A.; Magarvey, N.A. Optimizing Dimodular Nonribosomal Peptide Synthetases and Natural Dipeptides in an Escherichia coli Heterologous Host. Biochem. Cell Biol. 2013, 91, 203–208. [Google Scholar] [CrossRef]
- Blin, K.; Shaw, S.; Steinke, K.; Villebro, R.; Ziemert, N.; Lee, S.Y.; Medema, M.H.; Weber, T. antiSMASH 5.0: Updates to the Secondary Metabolite Genome Mining Pipeline. Nucleic Acids Res. 2019, 47, W81–W87. [Google Scholar] [CrossRef]
- Jacques, I.B.; Moutiez, M.; Witwinowski, J.; Darbon, E.; Martel, C.; Seguin, J.; Favry, E.; Thai, R.; Lecoq, A.; Dubois, S.; et al. Analysis of 51 Cyclodipeptide Synthases Reveals the Basis for Substrate Specificity. Nat. Chem. Biol. 2015, 11, 721–727. [Google Scholar] [CrossRef] [PubMed]
- Forseth, R.R.; Amaike, S.; Schwenk, D.; Affeldt, K.J.; Hoffmeister, D.; Schroeder, F.; Keller, N.P. Homologous NRPS-like Gene Clusters Mediate Redundant Small-Molecule Biosynthesis in Aspergillus flavus. Angew. Chem. Int. Ed. 2013, 52, 1590–1594. [Google Scholar] [CrossRef] [PubMed]
- Tanifuji, R.; Koketsu, K.; Takakura, M.; Asano, R.; Minami, A.; Oikawa, H.; Oguri, H. Chemo-enzymatic Total Syntheses of Jorunnamycin A, Saframycin A and N-Fmoc Saframycin Y3. J. Am. Chem. Soc. 2018, 140, 10705–10709. [Google Scholar] [CrossRef]
- Tietze, A.; Shi, Y.N.; Kronenwerth, M.; Bode, H.B. Nonribosomal Peptides Produced by Minimal and Engineered Synthetases with Terminal Reductase Domains. ChemBioChem 2020, 21, 2750–2754. [Google Scholar] [CrossRef]
- Yu, X.; Liu, F.; Zou, Y.; Tang, M.-C.; Hang, L.; Houk, K.N.; Tang, Y. Biosynthesis of Strained Piperazine Alkaloids: Uncovering the Concise Pathway of Herquline A. J. Am. Chem. Soc. 2016, 138, 13529–13532. [Google Scholar] [CrossRef] [PubMed]
- Gahloth, D.; Dunstan, M.S.; Quaglia, D.; Klumbys, E.; Lockhart-Cairns, M.P.; Hill, A.M.; Derrington, S.R.; Scrutton, N.S.; Turner, N.J.; Leys, D. Structures of Carboxylic Acid Reductase Reveal Domain Dynamics Underlying Catalysis. Nat. Chem. Biol. 2017, 13, 975–981. [Google Scholar] [CrossRef]
- Hai, Y.; Huang, A.M.; Tang, Y. Structure-Guided Function Discovery of an NRPS-Like Glycine Betaine Reductase for Choline Biosynthesis in Fungi. Proc. Natl. Acad. Sci. USA. 2019, 116, 10348–10353. [Google Scholar] [CrossRef]
- Schulz, S.; Fuhlendorff, J.; Reichenbach, H. Identification and Synthesis of Volatiles Released by the Myxobacterium Chondromyces crocatus. Tetrahedron 2004, 60, 3863–3872. [Google Scholar] [CrossRef]
- Maricic, N.; Dawid, S. Using the overlay assay to qualitatively measure bacterial production of and sensitivity to pneumococcal bacteriocins. J. Vis. Exp. 2014, e51876. [Google Scholar] [CrossRef] [PubMed]
- Koren, S.; Walenz, B.P.; Berlin, K.; Miller, J.R.; Bergman, N.H.; Phillippy, A.M. Canu: Scalable and Accurate Long-Read Assembly Via Adaptive K-Mer Weighting and Repeat Separation. Genome Res. 2017, 27, 722–736. [Google Scholar] [CrossRef] [PubMed]
- Seemann, T. Prokka: Rapid Prokaryotic Genome Annotation. Bioinformatics 2014, 30, 2068–2069. [Google Scholar] [CrossRef] [PubMed]
- Cockerill, F.; Wikler, M.; Alder, J.; Dudley, M.; Eliopoulos, G.; Ferraro, M.; Hardy, D.; Hecht, D.; Hindler, J.; Patel, J. Methods for Dilution Antimicrobial Susceptibility Tests for Bacteria That Grow Aerobically: Approved Standard. Clin. Lab. Stand. Inst. 2006, 26, M7–A7. [Google Scholar]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| 1 A | 2 A | 3 B | 4 B | 5 C | 6 C | |||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| δC | δH, multi. (J in Hz) | δC | δH, multi. (J in Hz) | δC | δH, multi. (J in Hz) | δC | δH, multi. (J in Hz) | δC | δH, multi. (J in Hz) | δC | δH, multi. (J in Hz) | |
| 1 | 142.7 | 8.27 (s) | 144.7 | 8.38 (s) | 141.6 | 8.33 (s) | 143.3 | 8.43 (s) | 142.5 | 8.26 (s) | 144.6 | 8.37 (s) |
| 2 | 158.1 | 156.1 | 155.7 | 154.0 | 157.8 D | 155.8 | ||||||
| 3 | 41.6 | 4.06 (s, 2H) | 41.1 | 4.04 (s, 2H) | 40.7 | 3.96 (s, 2H) | 39.6 | 3.94 (s, 2H) | 41.6 | 4.05 (s, 2H) | 41.1 | 4.05 (s, 2H) |
| 4 | 130.5 | 130.5 | 129.0 | 129.1 | 130.6 | 130.6 | ||||||
| 5 | 131.0 | 7.08 (d, 8.1) | 131.0 | 7.07 (d, 8.4) | 129.8 | 7.06 (d, 8.4) | 129.8 | 7.04 (d, 8.5) | 130.9 | 7.08 (d, 8.5) | 131.0 | 7.08 (d, 8.5) |
| 6 | 116.4 | 6.71 (d, 8.1) | 116.5 | 6.71 (d, 8.4) | 115.3 | 6.66 (d, 8.4) | 115.3 | 6.66 (d, 8.5) | 116.5 | 6.72 (d, 8.5) | 116.5 | 6.72 (d, 8.5) |
| 7 | 157.28 | 157.3 | 155.9 | 155.8 | 157.1 D | 157.3 | ||||||
| 8 | 116.4 | 6.71 (d, 8.1) | 116.5 | 6.71 (d, 8.4) | 115.3 | 6.66 (d, 8.4) | 115.3 | 6.66 (d, 8.5) | 116.5 | 6.72 (d, 8.5) | 116.5 | 6.72 (d, 8.5) |
| 9 | 131.0 | 7.08 (d, 8.1) | 131.0 | 7.07 (d, 8.4) | 129.8 | 7.06 (d, 8.4) | 129.8 | 7.04 (d, 8.5) | 130.9 | 7.08 (d, 8.5) | 131.0 | 7.08 (d, 8.5) |
| 1′ | 142.6 | 8.29 (s) | 144.6 | 8.40 (s) | 141.6 | 8.33 (s) | 143.3 | 8.43 (s) | 143.0 | δ 8.28 (s) | 145.0 | 8.38 (s) |
| 2′ | 157.34 | 155.3 | 155.7 | 154.0 | 157.5 D | 155.5 | ||||||
| 3′ | 42.3 | 4.16 (s, 2H) | 41.9 | 4.14 (s, 2H) | 40.7 | 3.96 (s, 2H) | 39.6 | 3.94 (s, 2H) | 45.0 | 2.68 (d, 7.3, 2H) | 44.6 | 2.66 (d, 7.2, 2H) |
| 4′ | 139.8 | 139.9 | 129.0 | 129.1 | 30.3 | 2.10 (m) | 30.3 | 2.05 (m) | ||||
| 5′ | 130.0 | 7.26 (d, 6.9) | 130.0 | 7.25 (d, 7.0) | 129.8 | 7.06 (d, 8.4) | 129.8 | 7.04 (d, 8.5) | 22.6 | 0.93 (d, 6.6, 3H) | 22.6 | 0.93 (d, 6.6, 3H) |
| 6′ | 129.7 | 7.28 (t, 6.9) | 129.8 | 7.28 (t, 7.0) | 115.3 | 6.66 (d, 8.4) | 115.3 | 6.66 (d, 8.5) | 22.6 | 0.93 (d, 6.6, 3H) | 22.6 | 0.93 (d, 6.6, 3H) |
| 7′ | 127.7 | 7.21 (t, 6.9) | 127.7 | 7.20 (t, 7.0) | 155.9 | 155.8 | ||||||
| 8′ | 129.7 | 7.28 (t, 6.9) | 129.8 | 7.28 (t, 7.0) | 115.3 | 6.66 (d, 8.4) | 115.3 | 6.66 (d, 8.5) | ||||
| 9′ | 130.0 | 7.26 (d, 6.9) | 130.0 | 7.25 (d, 7.0) | 129.8 | 7.06 (d, 8.4) | 129.8 | 7.04 (d, 8.5) | ||||
| N-1 E | 325.4 | 333.6 | 336.3 | 333.0 | 326.3 | |||||||
| N-2 E | 326.7 | 334.0 | 329.3 | 333.0 | 324.5 | |||||||
| -OH | 9.25 (s) | 9.23 (s) | ||||||||||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liu, H.-B.; Davison, J.R.; Rajwani, R.; Zhao, G.; Ohlemacher, S.I.; O’Connor, R.D.; Bewley, C.A. Lentzeacins A-E, New Bacterial-Derived 2,5- and 2,6-Disubstituted Pyrazines from a BGC-Rich Soil Bacterium Lentzea sp. GA3-008. Molecules 2021, 26, 7197. https://doi.org/10.3390/molecules26237197
Liu H-B, Davison JR, Rajwani R, Zhao G, Ohlemacher SI, O’Connor RD, Bewley CA. Lentzeacins A-E, New Bacterial-Derived 2,5- and 2,6-Disubstituted Pyrazines from a BGC-Rich Soil Bacterium Lentzea sp. GA3-008. Molecules. 2021; 26(23):7197. https://doi.org/10.3390/molecules26237197
Chicago/Turabian StyleLiu, Hong-Bing, Jack R. Davison, Rahim Rajwani, Gengxiang Zhao, Shannon I. Ohlemacher, Robert D. O’Connor, and Carole A. Bewley. 2021. "Lentzeacins A-E, New Bacterial-Derived 2,5- and 2,6-Disubstituted Pyrazines from a BGC-Rich Soil Bacterium Lentzea sp. GA3-008" Molecules 26, no. 23: 7197. https://doi.org/10.3390/molecules26237197
APA StyleLiu, H.-B., Davison, J. R., Rajwani, R., Zhao, G., Ohlemacher, S. I., O’Connor, R. D., & Bewley, C. A. (2021). Lentzeacins A-E, New Bacterial-Derived 2,5- and 2,6-Disubstituted Pyrazines from a BGC-Rich Soil Bacterium Lentzea sp. GA3-008. Molecules, 26(23), 7197. https://doi.org/10.3390/molecules26237197