
Antimicrobial and Antiviral (SARS-CoV-2) Potential of Cannabinoids and Cannabis sativa: A Comprehensive Review

 , , , and
, , , and
Abstract
1. Introduction
2. Antibacterial Activity of Cannabinoids and C. sativa
2.1. Historical Overview
2.2. Antibacterial Activities of Cannabinoids against Pathogens in the WHO’s Priority List
2.3. Antibacterial Activities of Cannabinoids against Pathogenic Bacteria Not on the WHO Priority List
3. Antifungal Activity
4. Antiviral Activity: Special Focus on SARS-CoV-2
4.1. Antiviral Efficacy against Viral Pathogens
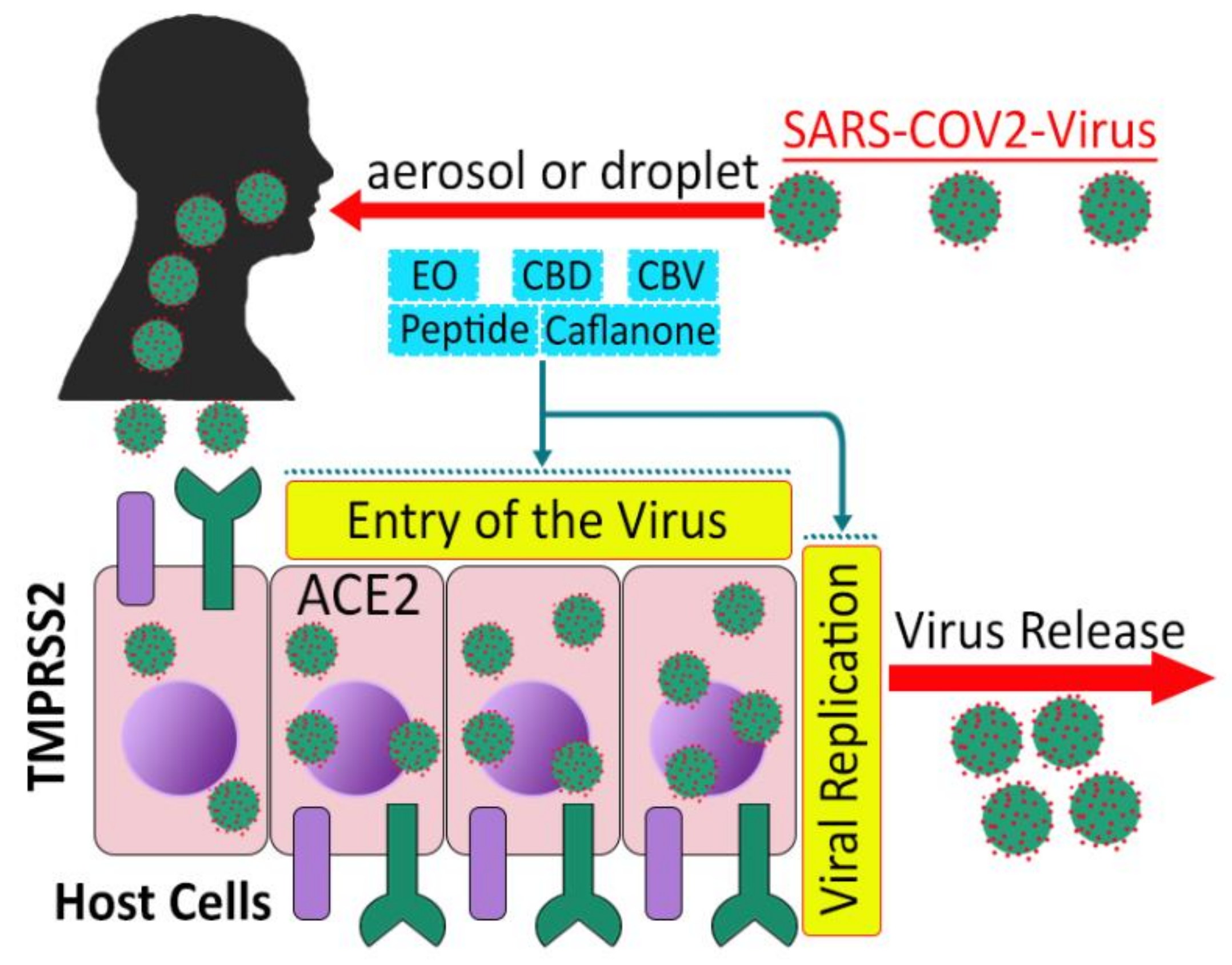
4.2. SARS-CoV-2
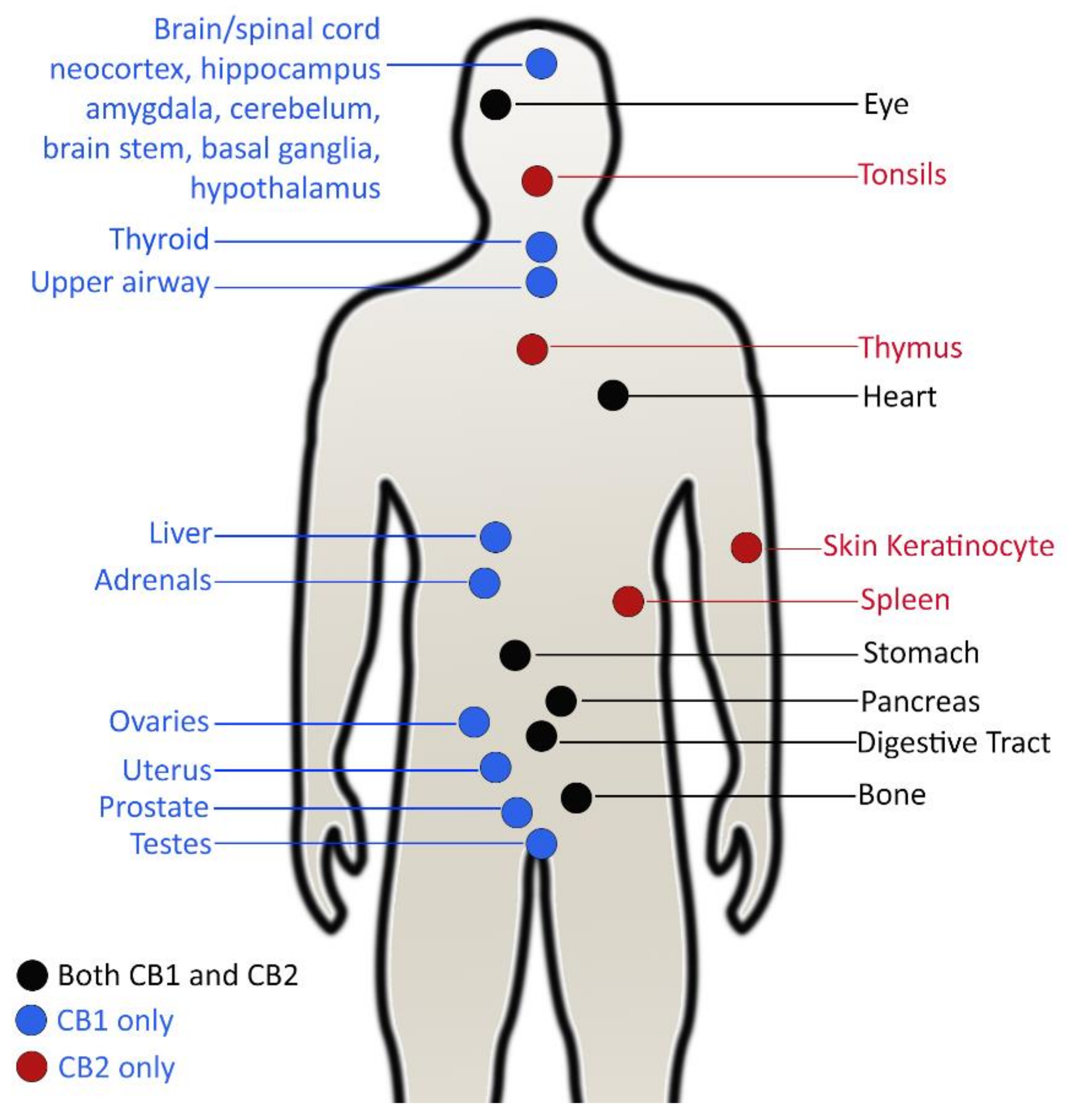
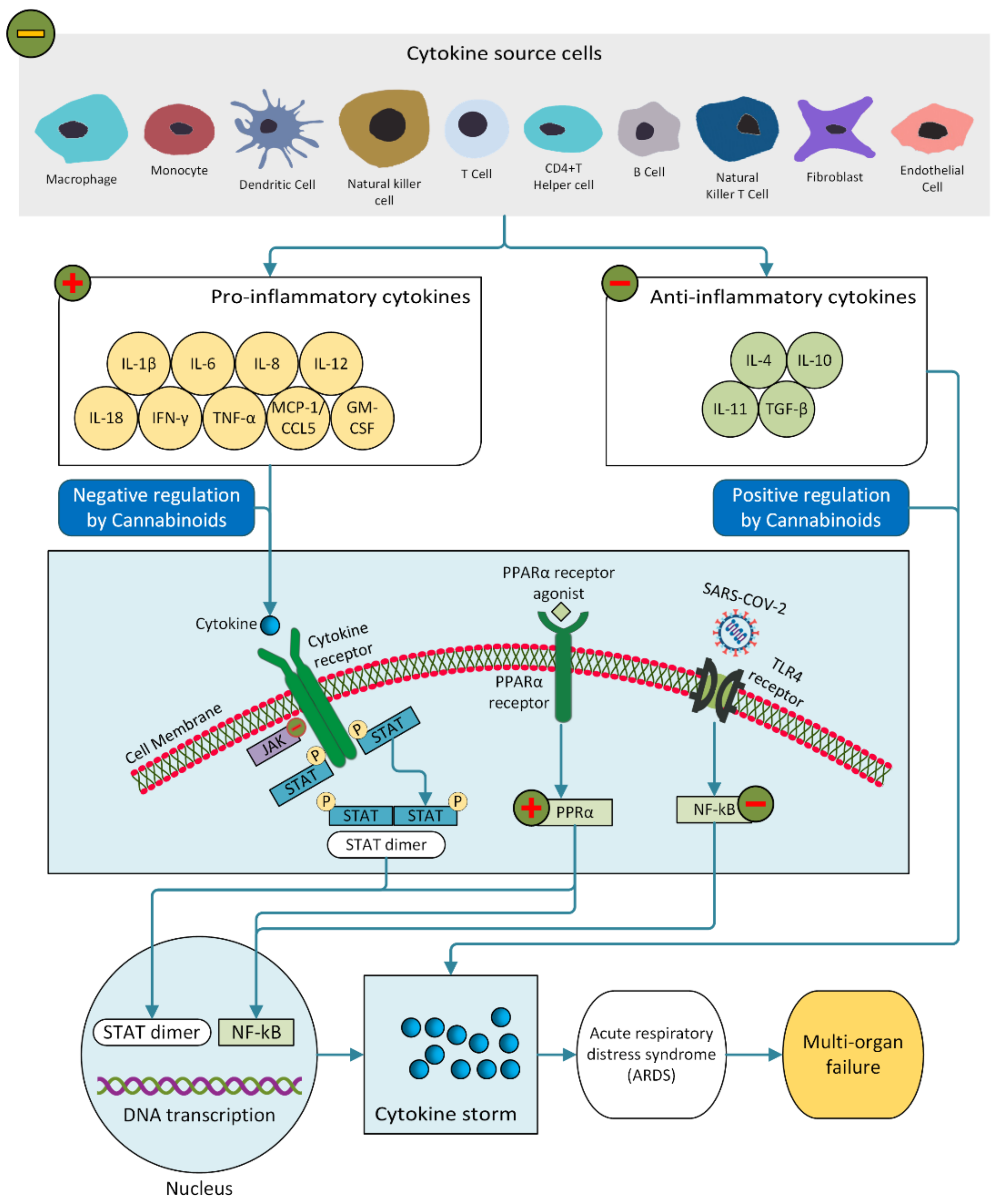
5. Mode of Action of Cannabinoids
6. Factors Affecting Antimicrobial and Antiviral Activities of Phytocompounds, EOs and Extracts
6.1. Physical Factors
6.2. Structure
6.3. Synergism
7. Potential Application of Antimicrobial Properties of Cannabinoids in Non-Drug Agents
7.1. Cosmetics (Toothpaste)
7.2. Food Plants
7.3. Crop Protection
7.4. Others Application
8. Challenge vs. Opportunity as a Pharmaceutical Drug
9. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
| ACE2 | Angiotensin-converting enzyme 2 |
| AIDS | Acquired immunodeficiency syndrome |
| ARDS | Acute respiratory distress syndrome |
| CBC | Cannabichromene |
| CBCA | Cannabichromenic acid |
| CBD | Cannabidiol |
| CBDA | Cannabidiolic acid |
| CBDV | Cannabidivarin |
| CBDVM | Cannabidivarin methyl ester |
| CBE | Cannabielsoin |
| CBG | Cannabigerol |
| CBL | Cannabicyclol |
| CBN | Cannabinol |
| CBT | Cannabitriol |
| CBV | Cannabivarin |
| CDC | Centers for disease control and prevention |
| CNS | Central nervous system |
| DNA | Deoxyribonucleic acid |
| EC | Endocannabinoid |
| EO | Essential oil |
| FDA | Food and drug administration |
| GNB | Gram-negative bacteria |
| GPB | Gram-positive bacteria |
| HMVEC | Human microvascular endothelial cells |
| HOMO | Highest occupied molecular orbital |
| IL | Interleukin |
| KSHV | Kaposi sarcoma associated herpesvirus |
| LPS | Lipopolysaccharide |
| LUMO | Lowest unoccupied molecular orbital |
| MBEC | Minimum biofilm eradication concentration |
| MD | Molecular dynamic |
| MIC | Minimum inhibitory concentration |
| MRSA | Methicillin-resistant Staphylococcus aureus |
| MSSA | Methicillin-susceptible Staphylococcus aureus |
| MV | Membrane vesicle |
| PBP | Penicillin-binding proteins |
| QIDP | Qualified infectious disease product |
| QSAR | Quantitative structure-activity relationship |
| RNA | Ribonucleic acid |
| R&D | Research and Development |
| SARS-CoV | Severe acute respiratory syndrome coronavirus |
| THC | Tetrahydrocannabinol |
| THCA | Tetrahydrocannabinolic acid |
| TMPRSS2 | Transmembrane protease, serine 2 |
| TNF-α | Tumor necrosis factor |
| VISA | Vancomycin-intermediate Staphylococcus aureus |
| VRSA | Vancomycin-resistant Staphylococcus aureus |
| WHO | World health organization |
References
- Burnett-Boothroyd, S.; McCarthy, B. Antimicrobial treatments of textiles for hygiene and infection control applications: An industrial perspective. In Textiles for Hygiene and Infection Control; Elsevier: Amsterdam, The Netherlands, 2011; pp. 196–209. [Google Scholar]
- Shahid, M.; Sobia, F.; Sahai, A.; Tripathi, T.; Singh, A.; Shahzad, A.; Khan, H.M. Umesh Plant Natural Products as a Potential Source for Antibacterial Agents: Recent Trends. Anti-Infect. Agents Med. Chem. 2009, 8, 211–225. [Google Scholar] [CrossRef]
- Ashkenazi, S. Beginning and possibly the end of the antibiotic era. J. Paediatr. Child Health 2012, 49, E179–E182. [Google Scholar] [CrossRef]
- Hutchings, M.I.; Truman, A.; Wilkinson, B. Editorial overview: Antimicrobials: Tackling AMR in the 21st century. Curr. Opin. Microbiol. 2019, 51, 3–5. [Google Scholar] [CrossRef] [PubMed]
- Prestinaci, F.; Pezzotti, P.; Pantosti, A. Antimicrobial resistance: A global multifaceted phenomenon. Pathog. Glob. Health 2015, 109, 309–318. [Google Scholar] [CrossRef] [PubMed]
- Abraham, E.P.; Chain, E. An Enzyme from Bacteria able to Destry penicillin. Nature 1940, 146, 837. [Google Scholar] [CrossRef]
- Ventola, C.L. The Antibiotic Resistance Crisis Part 1: Causes and Threats. Pharm. Ther. 2015, 40, 277–283. [Google Scholar]
- Antimicrobial Resistance. Available online: https://www.who.int/news-room/fact-sheets/detail/antimicrobial-resistance (accessed on 13 October 2020).
- CDC 2019 AR Threats Report. Available online: https://www.cdc.gov/drugresistance/biggest_threats.html (accessed on 9 August 2021).
- O’Neill, J. Review on Antimicrobial Resistance. Available online: https://amr-review.org/background.html (accessed on 1 July 2021).
- Ledingham, K.; Hinchliffe, S.; Jackson, M.; Thomas, F.; Tomson, G. Antibiotic resistance: Using a cultural contexts of health approach to address a global health challenge. WHO Reg. Off. Eur. 2019, 29, 166. [Google Scholar] [CrossRef]
- Aminov, R.I. A Brief History of the Antibiotic Era: Lessons Learned and Challenges for the Future. Front. Microbiol. 2010, 1, 134. [Google Scholar] [CrossRef] [PubMed]
- Wright, G.D. Something old, something new: Revisiting natural products in antibiotic drug discovery. Can. J. Microbiol. 2014, 60, 147–154. [Google Scholar] [CrossRef] [PubMed]
- Piddock, L. The crisis of no new antibiotics—What is the way forward? Lancet Infect. Dis. 2012, 12, 249–253. [Google Scholar] [CrossRef]
- WHO. Global Priority List of Antibiotic-Resistant Bacteria to Guide Research, Discovery, and Development of New Antibiotics; WHO: Geneva, Switzerland, 2017. [Google Scholar]
- John Hopkins University. Available online: https://coronavirus.jhu.edu/ (accessed on 3 July 2021).
- Zuardi, A.W. History of cannabis as a medicine: A review. Rev. Bras. Psiquiatr. 2006, 28, 153–157. [Google Scholar] [CrossRef] [PubMed]
- Kabelik, J.; Krejci, Z.; Santavy, F. Hemp as a medicament. Bull. Narc. 1960, 12, 5–23. [Google Scholar]
- Pisanti, S.; Malfitano, A.M.; Ciaglia, E.; Lamberti, A.; Ranieri, R.; Cuomo, G.; Abate, M.; Faggiana, G.; Proto, M.C.; Fiore, D.; et al. Cannabidiol: State of the art and new challenges for therapeutic applications. Pharmacol. Ther. 2017, 175, 133–150. [Google Scholar] [CrossRef]
- Gonçalves, J.; Rosado, T.; Soares, S.; Simão, A.Y.; Caramelo, D.; Luís, Â.; Fernández, N.; Barroso, M.; Gallardo, E.; Duarte, A.P. Cannabis and Its Secondary Metabolites: Their Use as Therapeutic Drugs, Toxicological Aspects, and Analytical Determination. Medicines 2019, 6, 31. [Google Scholar] [CrossRef] [PubMed]
- Nagarkatti, P.; Pandey, R.; Rieder, S.A.; Hegde, V.L.; Nagarkatti, M. Cannabinoids as novel anti-inflammatory drugs. Future Med. Chem. 2009, 1, 1333–1349. [Google Scholar] [CrossRef] [PubMed]
- Downer, E.J. Anti-inflammatory Potential of Terpenes Present in Cannabis sativa L. ACS Chem. Neurosci. 2020, 11, 659–662. [Google Scholar] [CrossRef]
- Gallily, R.; Yekhtin, Z.; Hanuš, L.O. The Anti-Inflammatory Properties of Terpenoids from Cannabis. Cannabis Cannabinoid Res. 2018, 3, 282–290. [Google Scholar] [CrossRef] [PubMed]
- Bonini, S.A.; Premoli, M.; Tambaro, S.; Kumar, A.; Maccarinelli, G.; Memo, M.; Mastinu, A. Cannabis sativa: A comprehensive ethnopharmacological review of a medicinal plant with a long history. J. Ethnopharmacol. 2018, 227, 300–315. [Google Scholar] [CrossRef] [PubMed]
- Paland, N.; Pechkovsky, A.; Aswad, M.; Hamza, H.; Popov, T.; Shahar, E.; Louria-Hayon, I. The Immunopathology of COVID-19 and the Cannabis Paradigm. Front. Immunol. 2021, 12, 631233. [Google Scholar] [CrossRef]
- Hanuš, L.O.; Hod, Y. Terpenes/Terpenoids in Cannabis: Are They Important? Med. Cannabis Cannabinoids 2020, 3, 25–60. [Google Scholar] [CrossRef]
- Tahamtan, A.; Tavakoli-Yaraki, M.; Rygiel, T.P.; Mokhtari-Azad, T.; Salimi, V. Effects of Cannabinoids and their Receptors on Viral Infections. J. Med. Virol. 2016, 88, 1–12. [Google Scholar] [CrossRef]
- Beji, C.; Loucif, H.; Telittchenko, R.; Olagnier, D.; Dagenais-Lussier, X.; Van Grevenynghe, J. Cannabinoid-Induced Immunomodulation during Viral Infections: A Focus on Mitochondria. Viruses 2020, 12, 875. [Google Scholar] [CrossRef]
- Reiss, C.S. Cannabinoids and Viral Infections. Pharmaceuticals 2010, 3, 1873–1886. [Google Scholar] [CrossRef] [PubMed]
- Klein, T.W.; Friedman, H.; Specter, S. Marijuana, immunity and infection. J. Neuroimmunol. 1998, 83, 102–115. [Google Scholar] [CrossRef]
- Cabral, G.A. Marijuana and Cannabinoids: Effects on infections, immunity, and AIDS. In Cannabis Therapeutics in HIV/AIDS; Russo, E.B., Ed.; Routledge: New York, USA, 2002; pp. 61–86. ISBN 9780203049105. [Google Scholar]
- Tagne, A.M.; Pacchetti, B.; Sodergren, M.; Cosentino, M.; Marino, F. Cannabidiol for Viral Diseases: Hype or Hope? Cannabis Cannabinoid Res. 2020, 5, 121–131. [Google Scholar] [CrossRef]
- Chianese, G.; Taglialatela-Scafati, O. Cannabinoids: Occurrence and Medicinal Chemistry. Curr. Med. Chem. 2011, 18, 1085–1099. [Google Scholar] [CrossRef]
- Nascimento, G.G.F.; Locatelli, J.; Freitas, P.C.; Silva, G.L. Antibacterial activity of plant extracts and phytochemicals on antibiotic-resistant bacteria. Braz. J. Microbiol. 2000, 31, 247–256. [Google Scholar] [CrossRef]
- Marcu, J.P. An Overview of Major and Minor Phytocannabinoids; Elsevier Inc.: Amsterdam, The Netherlands, 2016; Volume 1, ISBN 9780128003763. [Google Scholar]
- Krejci, Z. Hemp as a Medicament. Ph.D. Thesis, Faculty of Natural Sciences, Brno, Czech Republic, 1950. [Google Scholar]
- Krejci, Z. Antibacterial action of Canabis indica. Lek. List. 1952, 7, 500–503. [Google Scholar]
- Krejci, Z. Hanf (Cannabis sativa) -Antibiotisches Heilmittel. 2. Mitteilung: Methodik und Ergebnisse der bakteriologischen Untersuchungen und vorläufige klinische Erfahrungen. Pharmazie 1959, 14, 155–166. [Google Scholar]
- Ferenczy, L. Antbacterial substances in seeds. Nature 1956, 178, 639–640. [Google Scholar] [CrossRef]
- Ferenczy, L.; Gracza, L.; Jakobey, I. An antibacterial preparatum from hemp (Cannabis sativa L.). Naturwissenschaften 1958, 45, 188. [Google Scholar] [CrossRef]
- Boucher, H.W.; Talbot, G.H.; Bradley, J.S.; Edwards, J.E.; Gilbert, D.; Rice, L.B.; Scheld, M.; Spellberg, B.; Bartlett, J. Bad Bugs, No Drugs: No ESKAPE! An Update from the Infectious Diseases Society of America. Clin. Infect. Dis. 2009, 48, 1–12. [Google Scholar] [CrossRef]
- Nannini, E.; Murray, B.E.; Arias, C.A. Resistance or decreased susceptibility to glycopeptides, daptomycin, and linezolid in methicillin-resistant Staphylococcus aureus. Curr. Opin. Pharmacol. 2010, 10, 516–521. [Google Scholar] [CrossRef]
- Farha, M.A.; El-Halfawy, O.M.; Gale, R.T.; MacNair, C.R.; Carfrae, L.A.; Zhang, X.; Jentsch, N.G.; Magolan, J.; Brown, E.D. Uncovering the Hidden Antibiotic Potential of Cannabis. ACS Infect. Dis. 2020, 6, 338–346. [Google Scholar] [CrossRef] [PubMed]
- Lewis, K. Persister cells, dormancy and infectious disease. Nat. Rev. Genet. 2006, 5, 48–56. [Google Scholar] [CrossRef] [PubMed]
- Otto, M. Staphylococcal Infections: Mechanisms of Biofilm Maturation and Detachment as Critical Determinants of Pathogenicity. Annu. Rev. Med. 2013, 64, 175–188. [Google Scholar] [CrossRef]
- Mah, T.-F.C.; O’Toole, G.A. Mechanisms of biofilm resistance to antimicrobial agents. Trends Microbiol. 2001, 9, 34–39. [Google Scholar] [CrossRef]
- Conlon, B.P.; Rowe, S.E.; Gandt, A.B.; Nuxoll, A.S.; Donegan, N.P.; Zalis, E.A.; Clair, G.; Adkins, J.N.; Cheung, A.L.; Lewis, K. Persister formation in Staphylococcus aureus is associated with ATP depletion. Nat. Microbiol. 2016, 1, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Severin, A.; Tabei, K.; Tenover, F.; Chung, M.; Clarke, N.; Tomasz, A. High Level Oxacillin and Vancomycin Resistance and Altered Cell Wall Composition in Staphylococcus aureus Carrying the Staphylococcal mecA and the Enterococcal vanA Gene Complex. J. Biol. Chem. 2004, 279, 3398–3407. [Google Scholar] [CrossRef]
- Blaskovich, M.A.T.; Kavanagh, A.M.; Elliott, A.G.; Zhang, B.; Ramu, S.; Amado, M.; Lowe, G.J.; Hinton, A.O.; Pham, D.M.T.; Zuegg, J.; et al. The antimicrobial potential of cannabidiol. Commun. Biol. 2021, 4, 1–18. [Google Scholar] [CrossRef] [PubMed]
- Martinenghi, L.D.; Jønsson, R.; Lund, T.; Jenssen, H. Isolation, Purification, and Antimicrobial Characterization of Cannabidiolic Acid and Cannabidiol from Cannabis sativa L. Biomolecules 2020, 10, 900. [Google Scholar] [CrossRef] [PubMed]
- Wassmann, C.S.; Højrup, P.; Klitgaard, J.K. Cannabidiol is an effective helper compound in combination with bacitracin to kill Gram-positive bacteria. Sci. Rep. 2020, 10, 1–12. [Google Scholar] [CrossRef]
- Galletta, M.; Reekie, T.; Nagalingam, G.; Bottomley, A.; Harry, E.; Kassiou, M.; Triccas, J. Rapid Antibacterial Activity of Cannabichromenic Acid against Methicillin-Resistant Staphylococcus aureus. Antibiotics 2020, 9, 523. [Google Scholar] [CrossRef] [PubMed]
- Mascio, C.T.M.; Alder, J.D.; Silverman, J.A. Bactericidal Action of Daptomycin against Stationary-Phase and Nondividing Staphylococcus aureus Cells. Antimicrob. Agents Chemother. 2007, 51, 4255–4260. [Google Scholar] [CrossRef]
- Russo, E.B. Taming THC: Potential cannabis synergy and phytocannabinoid-terpenoid entourage effects. Br. J. Pharmacol. 2011, 163, 1344–1364. [Google Scholar] [CrossRef]
- Chakraborty, S.; Afaq, N.; Singh, N.; Majumdar, S. Antimicrobial activity of Cannabis sativa, Thuja orientalis and Psidium guajava leaf extracts against methicillin-resistant Staphylococcus aureus. J. Integr. Med. 2018, 16, 350–357. [Google Scholar] [CrossRef] [PubMed]
- Nashra, A.; Sujatha, R.; Sameer, D.; Bagoliwal, A.; Mishra, V.; Kumar, A.; Majid, A. Comparative Evaluation of Antibacterial Efficacy of Cannabis Sativa, Allium Sativum, Allium Cepa, Thuja Orientalis and Psidium Guajava against Drug Resistance Pathogens. Int. J. Health Sci. Res. 2018, 8, 89–97. [Google Scholar]
- Torres, J.A.; Villegas, M.V.; Quinn, J.P. Current concepts in antibiotic-resistant Gram-negative bacteria. Expert Rev. Anti-Infect. Ther. 2007, 5, 833–843. [Google Scholar] [CrossRef] [PubMed]
- Exner, M.; Bhattacharya, S.; Christiansen, B.; Gebel, J.; Goroncy-Bermes, P.; Hartemann, P.; Heeg, P.; Ilschner, C.; Kramer, A.; Larson, E.; et al. Antibiotic resistance: What is so special about multidrug-resistant Gram-negative bacteria? Antibiotikaresistenz: Was ist so besonders an den Gram-negativen. GMS Hyg. Infect. Control 2017, 12, 1–24. [Google Scholar]
- Miller, S.I. Antibiotic Resistance and Regulation of the Gram-Negative Bacterial Outer Membrane Barrier by Host Innate Immune Molecules. mBio 2016, 7, e01541-16. [Google Scholar] [CrossRef]
- Nafis, A.; Kasrati, A.; Jamali, C.A.; Mezrioui, N.; Setzer, W.; Abbad, A.; Hassani, L. Antioxidant activity and evidence for synergism of Cannabis sativa (L.) essential oil with antimicrobial standards. Ind. Crops Prod. 2019, 137, 396–400. [Google Scholar] [CrossRef]
- Zengin, G.; Menghini, L.; Di Sotto, A.; Mancinelli, R.; Sisto, F.; Carradori, S.; Cesa, S.; Fraschetti, C.; Filippi, A.; Angiolella, L.; et al. Chromatographic Analyses, In Vitro Biological Activities, and Cytotoxicity of Cannabis sativa L. Essential Oil: A Multidisciplinary Study. Molecules 2018, 23, 3266. [Google Scholar] [CrossRef]
- Shah, S.B.; Sartaj, L.; Hussain, S.; Ullah, N.; Idrees, M.; Shaheen, A.; Javed, M.S.; Aslam, M.K. In-vitro evaluation of antimicrobial, antioxidant, alpha-amylase inhibition and cytotoxicity properties of Cannabis sativa. Adv. Tradit. Med. 2019, 20, 181–187. [Google Scholar] [CrossRef]
- Kosgodage, U.S.; Matewele, P.; Awamaria, B.; Kraev, I.; Warde, P.; Mastroianni, G.; Nunn, A.V.; Guy, G.W.; Bell, J.D.; Inal, J.; et al. Cannabidiol Is a Novel Modulator of Bacterial Membrane Vesicles. Front. Cell. Infect. Microbiol. 2019, 9, 324. [Google Scholar] [CrossRef]
- Delcour, A.H. Outer membrane permeability and antibiotic resistance. Biochim. et Biophys. Acta (BBA)—Proteins Proteom. 2009, 1794, 808–816. [Google Scholar] [CrossRef]
- Nagakubo, T.; Nomura, N.; Toyofuku, M. Cracking Open Bacterial Membrane Vesicles. Front. Microbiol. 2019, 10, 3026. [Google Scholar] [CrossRef] [PubMed]
- Pang, Z.; Raudonis, R.; Glick, B.R.; Lin, T.-J.; Cheng, Z. Antibiotic resistance in Pseudomonas aeruginosa: Mechanisms and alternative therapeutic strategies. Biotechnol. Adv. 2018, 37, 177–192. [Google Scholar] [CrossRef]
- Ali, E.M.M.; Almagboul, A.Z.I.; Khogali, S.M.E.; Gergeir, U.M.A. Antimicrobial Activity of Cannabis sativa L. Chin. Med. 2012, 03, 61–64. [Google Scholar] [CrossRef]
- Anjum, M.; Arooj, Z.-E.-; Azam, S.; Rehman, P.; Khadim, J.; Anjum, M. Evaluation of antimicrobial activity and ethnobotanical study of Cannabis sativa L. Pure Appl. Biol. 2018, 7, 706–713. [Google Scholar] [CrossRef]
- Kourmouli, A.; Valenti, M.; Van Rijn, E.; Beaumont, H.J.E.; Kalantzi, O.-I.; Schmidt-Ott, A.; Biskos, G. Can disc diffusion susceptibility tests assess the antimicrobial activity of engineered nanoparticles? J. Nanoparticle Res. 2018, 20, 2–7. [Google Scholar] [CrossRef] [PubMed]
- Klančnik, A.; Piskernik, S.; Jeršek, B.; Možina, S.S. Evaluation of diffusion and dilution methods to determine the antibacterial activity of plant extracts. J. Microbiol. Methods 2010, 81, 121–126. [Google Scholar] [CrossRef] [PubMed]
- Mathur, P.; Singh, A.; Srivastava, V.R.; Singh, D.; Mishra, Y. Antimicrobial activity of indigenous wildly growing plants: Potential source of green antibiotics. Afr. J. Microbiol. Res. 2013, 7, 3807–3815. [Google Scholar] [CrossRef]
- Naveed, M.; Khan, T.A.; Ali, I.; Hassan, A.; Ali, H.; Ud, Z.; Hassan, Z.; Tabassum, S.; Majid, A.; Rehman, M.U. In vitro antibacterial activity of Cannabis sativa leaf extracts to some selective pathogenicbacterial strains. Int. J. Biosci. (IJB) 2014, 4, 65–70. [Google Scholar] [CrossRef]
- Lone, T.A.; Lone, R.A. Extraction of cannabinoids from Cannabis sativa L plant and its potential antimicrobial activity. Univers. J. Med. Dent. 2012, 1, 51–55. [Google Scholar]
- Mkpenie, V.N.; Essien, E.E.; Udoh, I.I. Effect of extraction conditions on total polyphenol contents, antioxidant and antimicrobial activities of Cannabis sativa L. Electron. J. Environ. Agric. Food Chem. 2012, 11, 300–307. [Google Scholar]
- Elsohly, H.N.; Turner, C.E.; Clark, A.M.; Elsohly, M.A. Synthesis and Antimicrobial Activities of Certain Cannabichromene and Cannabigerol Related Compounds. J. Pharm. Sci. 1982, 71, 1319–1323. [Google Scholar] [CrossRef]
- Ullah, S.; Jan, G.; Gul, F.; Khan, S.; Husna, H.; Sher, J.; Abidullah, S. Phytochemistry and antibacterial activities of some selected plants of war affected area of bajaur agency, pakistan. J. Pharmacogn. Phytochem. 2018, 7, 416–417. [Google Scholar]
- Nadir, I.; Rana, N.F.; Ahmad, N.M.; Tanweer, T.; Batool, A.; Taimoor, Z.; Riaz, S.; Ali, S.M. Cannabinoids and Terpenes as an Antibacterial and Antibiofouling Promotor for PES Water Filtration Membranes. Molecules 2020, 25, 691. [Google Scholar] [CrossRef]
- Kim, S.-Y.; Kang, D.-H.; Kim, J.-K.; Ha, Y.-G.; Hwang, J.Y.; Kim, T.; Lee, S.-H. Antimicrobial Activity of Plant Extracts Against Salmonella typhimurium, Escherichia coli O157:H7, and Listeria monocytogenes on Fresh Lettuce. J. Food Sci. 2010, 76, M41–M46. [Google Scholar] [CrossRef] [PubMed]
- Novak, J.; Zitterl-Eglseer, K.; Deans, S.G.; Franz, C.M. Essential oils of different cultivars of Cannabis sativa L. and their antimicrobial activity. Flavour Fragr. J. 2001, 16, 259–262. [Google Scholar] [CrossRef]
- Nasrullah, S.; Rahman, K.; Ikram, M.; Nisar, M.; Khan, I. Screening of antibacterial activity of medicinal plants. Int. J. Pharm. Sci. Rev. Res. 2012, 14, 25–29. [Google Scholar]
- Sharma, C.; Kaur, S.; Chaudhry, S.; Aman, R. Antimicrobial Potential of Three Common Weeds of Kurukshetra: An in vitro Study. Res. J. Microbiol. 2015, 10, 280–287. [Google Scholar] [CrossRef]
- Ali, M.; Romman, M.; Parvez, R.; Shuaib, M.; Bahadur, S.; Khalil, A.A.K.; Khan, M.; Haq, F.; Jan, S.; Hayat, S.S.S.; et al. Anti-bacterial activity of Cannabis sativa Linn. leaf extracts against different pathogenic bacterial strains. Biosci. Res. 2020, 17, 2730–2735. [Google Scholar]
- Zheljazkov, V.D.; Sikora, V.; Dincheva, I.; Kačániová, M.; Astatkie, T.; Semerdjieva, I.B.; Latkovic, D. Industrial, CBD, and Wild Hemp: How Different Are Their Essential Oil Profile and Antimicrobial Activity? Molecules 2020, 25, 4631. [Google Scholar] [CrossRef] [PubMed]
- Mikulcová, V.; Kašpárková, V.; Humpolíček, P.; Buňková, L. Formulation, Characterization and Properties of Hemp Seed Oil and Its Emulsions. Molecules 2017, 22, 700. [Google Scholar] [CrossRef] [PubMed]
- Oyedemi, B.M. Antiplasmid and Antimicrobial Activities of Synthetic and Natural Products from Selected Medicinal Plants; UCL School of Pharmacy London: London, UK, 2014. [Google Scholar]
- Raleigh, E.; Low, K. Conjugation. Brenner’s Encycl. Genet. Second Ed. 2013, 1, 144–151. [Google Scholar] [CrossRef]
- Spengler, G.; Molnar, A.; Schelz, Z.; Amaral, L.; Sharples, D.; Molnar, J. The Mechanism of Plasmid Curing in Bacteria. Curr. Drug Targets 2006, 7, 823–841. [Google Scholar] [CrossRef]
- Molnár, J.; Csiszár, K.; Nishioka, I.; Shoyama, Y. The effects of cannabispiro compounds and tetrahydrocannabidiolic acid on the plasmid transfer and maintenance in Escherichia coli. Acta Microbiol. Hung. 1986, 33, 221–231. [Google Scholar]
- Feldman, M.; Smoum, R.; Mechoulam, R.; Steinberg, D. Antimicrobial potential of endocannabinoid and endocannabinoid-like compounds against methicillin-resistant Staphylococcus aureus. Sci. Rep. 2018, 8, 17696. [Google Scholar] [CrossRef]
- Feldman, M.; Smoum, R.; Mechoulam, R.; Steinberg, D. Potential combinations of endocannabinoid/endocannabinoid-like compounds and antibiotics against methicillin-resistant Staphylococcus aureus. PLoS ONE 2020, 15, e0231583. [Google Scholar] [CrossRef]
- Veringa, E.M.; Ferguson, D.A.; Lambe, D.W.; Verhoef, J. The role of glycocalyx in surface phagocytosis of Baeteroides spp., in the presence and absence of clindamycin. J. Antimicrob. Chemother. 1989, 23, 711–720. [Google Scholar] [CrossRef]
- Nazir, R.; Rehman, S.; Nisa, M.; Baba, U. ali Exploring bacterial diversity: From cell to sequence. In Freshwater Microbiology: Perspectives of Bacterial Dynamics in Lake Ecosystems; Elsevier Inc.: Amsterdam, The Netherlands, 2019; pp. 263–306. ISBN 9780128174951. [Google Scholar]
- Nissen, L.; Zatta, A.; Stefanini, I.; Grandi, S.; Sgorbati, B.; Biavati, B.; Monti, A. Characterization and antimicrobial activity of essential oils of industrial hemp varieties (Cannabis sativa L.). Fitoterapia 2010, 81, 413–419. [Google Scholar] [CrossRef]
- Iseppi, R.; Brighenti, V.; Licata, M.; Lambertini, A.; Sabia, C.; Messi, P.; Pellati, F.; Benvenuti, S. Chemical Characterization and Evaluation of the Antibacterial Activity of Essential Oils from Fibre-Type Cannabis sativa L. (Hemp). Molecules 2019, 24, 2302. [Google Scholar] [CrossRef] [PubMed]
- Appendino, G.; Gibbons, S.; Giana, A.; Pagani, A.; Grassi, G.; Stavri, M.; Smith, E.; Rahman, M. Antibacterial Cannabinoids from Cannabis sativa: A Structure−Activity Study. J. Nat. Prod. 2008, 71, 1427–1430. [Google Scholar] [CrossRef] [PubMed]
- Radwan, M.M.; ElSohly, M.A.; Slade, D.; Ahmed, S.A.; Khan, I.A.; Ross, S.A. Biologically Active Cannabinoids from High-Potency Cannabis sativa. J. Nat. Prod. 2009, 72, 906–911. [Google Scholar] [CrossRef] [PubMed]
- Nalli, Y.; Arora, P.; Riyaz-Ul-Hassan, S.; Ali, A. Chemical investigation of Cannabis sativa leading to the discovery of a prenylspirodinone with anti-microbial potential. Tetrahedron Lett. 2018, 59, 2470–2472. [Google Scholar] [CrossRef]
- Wanas, A.S.; Radwan, M.M.; Mehmedic, Z.; Jacob, M.; Khan, I.A.; Elsohly, M.A. Antifungal activity of the volatiles of high potency Cannabis sativa L. Against Cryptococcus neoformans. Rec. Nat. Prod. 2015, 10, 214–220. [Google Scholar]
- Ferrante, C.; Recinella, L.; Ronci, M.; Menghini, L.; Brunetti, L.; Chiavaroli, A.; Leone, S.; Di Iorio, L.; Carradori, S.; Tirillini, B.; et al. Multiple pharmacognostic characterization on hemp commercial cultivars: Focus on inflorescence water extract activity. Food Chem. Toxicol. 2019, 125, 452–461. [Google Scholar] [CrossRef]
- Elhendawy, M.A.; Wanas, A.S.; Radwan, M.M.; Azzaz, N.A.; Toson, E.S.; ElSohly, M.A. Chemical and Biological Studies of Cannabis sativa Roots. Med. Cannabis Cannabinoids 2018, 1, 104–111. [Google Scholar] [CrossRef]
- Frassinetti, S.; Gabriele, M.; Moccia, E.; Longo, V.; Di Gioia, D. Antimicrobial and antibiofilm activity of Cannabis sativa L. seeds extract against Staphylococcus aureus and growth effects on probiotic Lactobacillus spp. LWT 2020, 124, 109149. [Google Scholar] [CrossRef]
- Nocera, F.P.; Mancini, S.; Najar, B.; Bertelloni, F.; Pistelli, L.; De Filippis, A.; Fiorito, F.; De Martino, L.; Fratini, F. Antimicrobial Activity of Some Essential Oils against Methicillin-Susceptible and Methicillin-Resistant Staphylococcus pseudintermedius-Associated Pyoderma in Dogs. Animals 2020, 10, 1782. [Google Scholar] [CrossRef]
- Radošević, A.; Kupinić, M.; Grlić, L.; Kupini, M. Antibiotic Activity of Various Types of Cannabis Resin. Nature 1962, 195, 1007–1009. [Google Scholar] [CrossRef] [PubMed]
- Kabelik, V.J. Hanf (Cannabis sativa)—Antibiotisches Heilmittel. 1. Mitteilung: Hanf in der Alt- und Volksmedizin. Pharmazie 1958, 12, 439–443. [Google Scholar]
- Wasim, K.; Haq, I.; Ashraf, M. Antimicrobial studies of the leaf of Cannabis sativa L. Pak. J. Pharm. Sci. 1995, 8, 29–38. [Google Scholar]
- Borchardt, J.R.; Wyse, D.L.; Sheaffer, C.C.; Kauppi, K.L.; Fulcher, R.G.; Ehlke, N.J.; Biesboer, D.D.; Bey, R.F. Antimicrobial activity of native and naturalized plants of Minnesota and Wisconsin. J. Med. Plants Res. 2008, 2, 98–110. [Google Scholar]
- Zheljazkov, V.D.; Sikora, V.; Semerdjieva, I.B.; Kačániová, M.; Astatkie, T.; Dincheva, I. Grinding and Fractionation during Distillation Alter Hemp Essential Oil Profile and Its Antimicrobial Activity. Molecules 2020, 25, 3943. [Google Scholar] [CrossRef] [PubMed]
- Lelario, F.; Scrano, L.; De Franchi, S.; Bonomo, M.G.; Salzano, G.; Milan, S.; Milella, L.; Bufo, S.A. Identification and antimicrobial activity of most representative secondary metabolites from different plant species. Chem. Biol. Technol. Agric. 2018, 5, 13. [Google Scholar] [CrossRef]
- Viswanath, H.; Bhat, K.A.; Bhat, N.; Wani, T.; Mughal, M.N. Antibacterial Efficacy of Aqueous Plant Extracts against Storage Soft Rot of Potato Caused by Erwinia carotovora. Int. J. Curr. Microbiol. Appl. Sci. 2018, 7, 2630–2639. [Google Scholar] [CrossRef][Green Version]
- Pellegrini, M.; Palmieri, S.; Ricci, A.; Serio, A.; Paparella, A.; Sterzo, C.L. In vitro antioxidant and antimicrobial activity of Cannabis sativa L. cv ‘Futura 75’ essential oil. Nat. Prod. Res. 2020, 1–5. [Google Scholar] [CrossRef]
- Marini, E.; Magi, G.; Ferretti, G.; Bacchetti, T.; Giuliani, A.; Pugnaloni, A.; Rippo, M.R.; Facinelli, B. Attenuation of Listeria monocytogenes Virulence by Cannabis sativa L. Essential Oil. Front. Cell. Infect. Microbiol. 2018, 8, 293. [Google Scholar] [CrossRef]
- Turner, C.E.; Elsohly, M.A. Biological Activity of Cannabichromene, its Homologs and Isomers. J. Clin. Pharmacol. 1981, 21, 283S–291S. [Google Scholar] [CrossRef]
- Anumudu, C.K.; Akpaka, M.N.; Anumudu, I.C. Antimicrobial activity of Cannabis sativa extracts on Lancefield Group A Streptococcus species associated with streptococcal pharyngitis (strep throat). Afr. J. Biol. Sci. 2020, 2, 9. [Google Scholar] [CrossRef]
- Radwan, M.M.; Ross, S.A.; Slade, D.; Ahmed, S.A.; Zulfiqar, F.; ElSohly, M.A. Isolation and Characterization of New Cannabis Constituents from a High Potency Variety. Planta Med. 2008, 74, 267–272. [Google Scholar] [CrossRef] [PubMed]
- Van Klingeren, B.; Ham, M.T. Antibacterial activity of Δ9-tetrahydrocannabinol and cannabidiol. Antonie van Leeuwenhoek 1976, 42, 9–12. [Google Scholar] [CrossRef]
- Nola, I.; Kostović, K.; Oremović, L.; Soldo-Belić, A.; Lugović, L. Candida infections today—How big is the problem? Acta Dermatovenerol. Croat. 2003, 11, 171–177. [Google Scholar]
- Aleksic, V.; Knezevic, P. Antimicrobial and antioxidative activity of extracts and essential oils of Myrtus communis L. Microbiol. Res. 2014, 169, 240–254. [Google Scholar] [CrossRef] [PubMed]
- Dabur, R.; Singh, H.; Chhillar, A.; Ali, M.; Sharma, G. Antifungal potential of Indian medicinal plants. Fitoterapia 2004, 75, 389–391. [Google Scholar] [CrossRef] [PubMed]
- Kuete, V. Potential of Cameroonian Plants and Derived Products against Microbial Infections: A Review. Planta Med. 2010, 76, 1479–1491. [Google Scholar] [CrossRef]
- Nobile, C.J.; Johnson, A.D. Candida albicans Biofilms and Human Disease. Annu. Rev. Microbiol. 2015, 69, 71–92. [Google Scholar] [CrossRef]
- Tsui, C.; Kong, E.F.; Jabra-Rizk, M.A. Pathogenesis ofCandida albicansbiofilm. Pathog. Dis. 2016, 74, ftw018. [Google Scholar] [CrossRef]
- Khan, I.H.; Javaid, A. Antifungal activity of leaf extract of Cannabis sativa against Aspergillus flavipes. Pak. J. Weed Sci. Res. 2020, 26, 447–453. [Google Scholar] [CrossRef]
- Orlando, G.; Adorisio, S.; Delfino, D.; Chiavaroli, A.; Brunetti, L.; Recinella, L.; Leone, S.; D’Antonio, M.; Zengin, G.; Acquaviva, A.; et al. Comparative Investigation of Composition, Antifungal, and Anti-Inflammatory Effects of the Essential Oil from Three Industrial Hemp Varieties from Italian Cultivation. Antibiotics 2021, 10, 334. [Google Scholar] [CrossRef] [PubMed]
- Aboul-ela, M.A.; Bahaa, N.; Din, E. Antimicrobial Evaluation of Extracts from some Yemeni Plants. Alexander J. Pharm. Sci. 1995, 9, 35–37. [Google Scholar]
- Ahmed, S.A.; Ross, S.A.; Slade, D.; Radwan, M.M.; Zulfiqar, F.; ElSohly, M.A. Cannabinoid Ester Constituents from High-Potency Cannabis sativa. J. Nat. Prod. 2008, 71, 536–542. [Google Scholar] [CrossRef] [PubMed]
- Žitek, T.; Leitgeb, M.; Golle, A.; Dariš, B.; Knez, Ž.; Hrnčič, M.K. The Influence of Hemp Extract in Combination with Ginger on the Metabolic Activity of Metastatic Cells and Microorganisms. Molecules 2020, 25, 4992. [Google Scholar] [CrossRef]
- Miller, A.M.; Stella, N. CB2 receptor-mediated migration of immune cells: It can go either way. Br. J. Pharmacol. 2008, 153, 299–308. [Google Scholar] [CrossRef] [PubMed]
- Yao, B.; Mackie, K. Endocannabinoid Receptor Pharmacology. Curr. Top. Behav. Neurosci. 2009, 1, 37–63. [Google Scholar] [CrossRef]
- Reddy, P.M.; Maurya, N.; Velmurugan, B.K. Medicinal Use of Synthetic Cannabinoids—A Mini Review. Curr. Pharmacol. Rep. 2019, 5, 1–13. [Google Scholar] [CrossRef]
- Herrera, R.A.; Oved, J.H.; Reiss, C.S. Disruption of IFN-γ–Mediated Antiviral Activity in Neurons: The Role of Cannabinoids. Viral Immunol. 2008, 21, 141–152. [Google Scholar] [CrossRef]
- Rice, W.; Shannon, J.M.; Burton, F.; Fiedeldey, D. Expression of a brain-type cannabinoid receptor (CB1) in alveolar Type II cells in the lung: Regulation by hydrocortisone. Eur. J. Pharmacol. 1997, 327, 227–232. [Google Scholar] [CrossRef]
- Van Der Poorten, D.; Shahidi, M.; Tay, E.; Sesha, J.; Tran, K.; McLeod, D.; Milliken, J.S.; Ho, V.; Hebbard, L.W.; Douglas, M.W.; et al. Hepatitis C Virus Induces the Cannabinoid Receptor 1. PLoS ONE 2010, 5, e12841. [Google Scholar] [CrossRef] [PubMed]
- Nichols, J.M.; Kaplan, B.L. Immune Responses Regulated by Cannabidiol. Cannabis Cannabinoid Res. 2020, 5, 12–31. [Google Scholar] [CrossRef]
- Walter, L.; Stella, N. Cannabinoids and neuroinflammation. Br. J. Pharmacol. 2004, 141, 775–785. [Google Scholar] [CrossRef] [PubMed]
- D’Addario, C.; Di Francesco, A.; Pucci, M.; Agrò, A.F.; Maccarrone, M. Epigenetic mechanisms and endocannabinoid signalling. FEBS J. 2013, 280, 1905–1917. [Google Scholar] [CrossRef] [PubMed]
- Rock, R.B.; Gekker, G.; Hu, S.; Sheng, W.S.; Cabral, G.A.; Martin, B.R.; Peterson, P.K. WIN55,212-2-Mediated Inhibition of HIV-1 Expression in Microglial Cells: Involvement of Cannabinoid Receptors. J. Neuroimmune Pharmacol. 2006, 2, 178–183. [Google Scholar] [CrossRef]
- Ramirez, S.H.; Reichenbach, N.L.; Fan, S.; Rom, S.; Merkel, S.F.; Wang, X.; Ho, W.-Z.; Persidsky, Y. Attenuation of HIV-1 replication in macrophages by cannabinoid receptor 2 agonists. J. Leukoc. Biol. 2013, 93, 801–810. [Google Scholar] [CrossRef]
- Molina, P.E.; Winsauer, P.; Zhang, P.; Walker, E.; Birke, L.; Amedee, A.; Stouwe, C.V.; Troxclair, D.; McGoey, R.; Varner, K.; et al. Cannabinoid Administration Attenuates the Progression of Simian Immunodeficiency Virus. AIDS Res. Hum. Retrovir. 2011, 27, 585–592. [Google Scholar] [CrossRef]
- Abubakar, Y.U.; Taura, D.W.; Yushau, M.; Muhammad, A.U. An in ovo investigation on antiviral activity of Cannabis sativa extracts against Newcastle Disease Virus (NDV). Adv. Pharm. J. 2020, 5, 21–30. [Google Scholar] [CrossRef]
- Medveczky, M.M.; Sherwood, T.A.; Klein, T.W.; Friedman, H.; Medveczky, P.G. Delta-9 tetrahydrocannabinol (THC) inhibits lytic replication of gamma oncogenic herpesviruses in vitro. BMC Med. 2004, 2, 34. [Google Scholar] [CrossRef]
- Lancz, G.; Specter, S.; Brown, H.K. Suppressive Effect of -9-Tetrahydrocannabinol on Herpes Simplex Virus Infectivity In Vitro. Exp. Biol. Med. 1991, 196, 401–404. [Google Scholar] [CrossRef]
- Blevins, R.D.; Dumic, M.P. The Effect of -9-Tetrahydrocannabinol on Herpes Simplex Virus Replication. J. Gen. Virol. 1980, 49, 427–431. [Google Scholar] [CrossRef]
- Toyang, N.J.; Lowe, H.I.C.; McLaughlin, W. Potential of cannabidiol for the treatment of viral hepatitis. Pharmacogn. Res. 2017, 9, 116–118. [Google Scholar] [CrossRef] [PubMed]
- Maor, Y.; Yu, J.; Kuzontkoski, P.M.; Dezube, B.J.; Zhang, X.; Groopman, J.E. Cannabidiol Inhibits Growth and Induces Programmed Cell Death in Kaposi Sarcoma-Associated Herpesvirus-Infected Endothelium. Genes Cancer 2012, 3, 512–520. [Google Scholar] [CrossRef] [PubMed]
- Lowe, H.; Steele, B.; Bryant, J.; Fouad, E.; Toyang, N.; Ngwa, W. Antiviral Activity of Jamaican Medicinal Plants and Isolated Bioactive Compounds. Molecules 2021, 26, 607. [Google Scholar] [CrossRef]
- FDA. FDA and Cannabis: Research and Drug Approval Process. Available online: https://www.fda.gov/news-events/public-health-focus/fda-and-cannabis-research-and-drug-approval-process (accessed on 10 May 2021).
- Wu, Y.; Ho, W.; Huang, Y.; Jin, D.-Y.; Li, S.; Liu, S.-L.; Liu, X.; Qiu, J.; Sang, Y.; Wang, Q.; et al. SARS-CoV-2 is an appropriate name for the new coronavirus. Lancet 2020, 395, 949–950. [Google Scholar] [CrossRef]
- Hojyo, S.; Uchida, M.; Tanaka, K.; Hasebe, R.; Tanaka, Y.; Murakami, M.; Hirano, T. How COVID-19 induces cytokine storm with high mortality. Inflamm. Regen. 2020, 40, 1–7. [Google Scholar] [CrossRef]
- Mahmudpour, M.; Roozbeh, J.; Keshavarz, M.; Farrokhi, S.; Nabipour, I. COVID-19 cytokine storm: The anger of inflammation. Cytokine 2020, 133, 155151. [Google Scholar] [CrossRef]
- Xu, X.; Han, M.; Li, T.; Sun, W.; Wang, D.; Fu, B.; Zhou, Y.; Zheng, X.; Yang, Y.; Li, X.; et al. Effective treatment of severe COVID-19 patients with tocilizumab. Proc. Natl. Acad. Sci. USA 2020, 117, 10970–10975. [Google Scholar] [CrossRef] [PubMed]
- Del Valle, D.M.; Kim-Schulze, S.; Huang, H.-H.; Beckmann, N.D.; Nirenberg, S.; Wang, B.; Lavin, Y.; Swartz, T.H.; Madduri, D.; Stock, A.; et al. An inflammatory cytokine signature predicts COVID-19 severity and survival. Nat. Med. 2020, 26, 1636–1643. [Google Scholar] [CrossRef]
- Huang, Q.; Wu, X.; Zheng, X.; Luo, S.; Xu, S.; Weng, J. Targeting inflammation and cytokine storm in COVID-19. Pharmacol. Res. 2020, 159, 105051. [Google Scholar] [CrossRef]
- Song, Y.; Zhang, M.; Yin, L.; Wang, K.; Zhou, Y.; Zhou, M.; Lu, Y. COVID-19 treatment: Close to a cure? A rapid review of pharmacotherapies for the novel coronavirus (SARS-CoV-2). Int. J. Antimicrob. Agents 2020, 56, 106080. [Google Scholar] [CrossRef]
- Lucaciu, O.; Aghiorghiesei, O.; Petrescu, N.B.; Mirica, I.C.; Benea, H.R.C.; Apostu, D. In quest of a new therapeutic approach in COVID-19: The endocannabinoid system. Drug Metab. Rev. 2021, 53, 478–490. [Google Scholar] [CrossRef]
- Wang, B.; Kovalchuk, A.; Li, D.; Rodriguez-Juarez, R.; Ilnytskyy, Y.; Kovalchuk, I.; Kovalchuk, O. In search of preventative strategies: Novel high-CBD Cannabis sativa extracts modulate ACE2 expression in COVID-19 gateway tissues. Aging 2020, 12, 22425–22444. [Google Scholar] [CrossRef]
- Zhou, P.; Yang, X.-L.; Wang, X.-G.; Hu, B.; Zhang, L.; Zhang, W.; Si, H.-R.; Zhu, Y.; Li, B.; Huang, C.-L.; et al. A pneumonia outbreak associated with a new coronavirus of probable bat origin. Nature 2020, 579, 270–273. [Google Scholar] [CrossRef] [PubMed]
- Mollica, V.; Rizzo, A.; Massari, F. The pivotal role of TMPRSS2 in coronavirus disease 2019 and prostate cancer. Futur. Oncol. 2020, 16, 2029–2033. [Google Scholar] [CrossRef]
- Hoffmann, M.; Kleine-Weber, H.; Schroeder, S.; Krüger, N.; Herrler, T.; Erichsen, S.; Schiergens, T.S.; Herrler, G.; Wu, N.-H.; Nitsche, A.; et al. SARS-CoV-2 Cell Entry Depends on ACE2 and TMPRSS2 and Is Blocked by a Clinically Proven Protease Inhibitor. Cell 2020, 181, 271–280.e8. [Google Scholar] [CrossRef] [PubMed]
- Li, M.; Chen, L.; Zhang, J.; Xiong, C.; Li, X. The SARS-CoV-2 receptor ACE2 expression of maternal-fetal interface and fetal organs by single-cell transcriptome study. PLoS ONE 2020, 15, e0230295. [Google Scholar] [CrossRef]
- Malinowska, B.; Baranowska-kuczko, M.; Kicman, A.; Schlicker, E. Opportunities, Challenges and Pitfalls of Using Cannabidiol as an Adjuvant Drug in COVID-19. Int. J. Mol. Sci. 2021, 22, 1986. [Google Scholar] [CrossRef] [PubMed]
- Ragia, G.; Manolopoulos, V.G. Inhibition of SARS-CoV-2 entry through the ACE2/TMPRSS2 pathway: A promising approach for uncovering early COVID-19 drug therapies. Eur. J. Clin. Pharmacol. 2020, 76, 1623–1630. [Google Scholar] [CrossRef] [PubMed]
- Jin, Z.; Du, X.; Xu, Y.; Deng, Y.; Liu, M.; Zhao, Y.; Zhang, B.; Li, X.; Zhang, L.; Peng, C.; et al. Structure of Mpro from SARS-CoV-2 and discovery of its inhibitors. Nature 2020, 582, 289–293. [Google Scholar] [CrossRef]
- Zhang, L.; Lin, D.; Sun, X.; Curth, U.; Drosten, C.; Sauerhering, L.; Becker, S.; Rox, K.; Hilgenfeld, R. Crystal structure of SARS-CoV-2 main protease provides a basis for design of improved α-ketoamide inhibitors. Science 2020, 368, 409–412. [Google Scholar] [CrossRef] [PubMed]
- COVID-19 Treatment Guidelines Panel. Coronavirus Disease 2019 (COVID-19) Treatment Guidelines. Available online: https://www.covid19treatmentguidelines.nih.gov (accessed on 3 July 2021).
- Rossi, F.; Tortora, C.; Argenziano, M.; Di Paola, A.; Punzo, F. Cannabinoid Receptor Type 2: A Possible Target in SARS-CoV-2 (CoV-19) Infection? Int. J. Mol. Sci. 2020, 21, 3809. [Google Scholar] [CrossRef] [PubMed]
- Hill, K.P. Cannabinoids and the Coronavirus. Cannabis Cannabinoid Res. 2020, 5, 118–120. [Google Scholar] [CrossRef] [PubMed]
- Onaivi, E.S.; Sharma, V. Cannabis for COVID-19: Can cannabinoids quell the cytokine storm? Future Sci. OA 2020, 6, FSO625. [Google Scholar] [CrossRef]
- Esposito, G.; Pesce, M.; Seguella, L.; Sanseverino, W.; Lu, J.; Corpetti, C.; Sarnelli, G. The potential of cannabidiol in the COVID-19 pandemic. Br. J. Pharmacol. 2020, 177, 4967–4970. [Google Scholar] [CrossRef] [PubMed]
- Mamber, S.W.; Krakowka, S.; Osborn, J.; Saberski, L.; Rhodes, R.G.; Dahlberg, A.E.; Pond-Tor, S.; Fitzgerald, K.; Wright, N.; Beseme, S.; et al. Can Unconventional Immunomodulatory Agents Help Alleviate COVID-19 Symptoms and Severity? mSphere 2020, 5, e00288-20. [Google Scholar] [CrossRef]
- Costiniuk, C.T.; Jenabian, M.-A. Acute inflammation and pathogenesis of SARS-CoV-2 infection: Cannabidiol as a potential anti-inflammatory treatment? Cytokine Growth Factor Rev. 2020, 53, 63–65. [Google Scholar] [CrossRef] [PubMed]
- Byrareddy, S.N.; Mohan, M. SARS-CoV2 induced respiratory distress: Can cannabinoids be added to anti-viral therapies to reduce lung inflammation? Brain Behav. Immun. 2020, 87, 120–121. [Google Scholar] [CrossRef]
- Zhu, W.; Newton, C.; Daaka, Y.; Friedman, H.; Klein, T.W. delta 9-Tetrahydrocannabinol enhances the secretion of interleukin 1 from endotoxin-stimulated macrophages. J. Pharmacol. Exp. Ther. 1994, 270, 1334–1339. [Google Scholar]
- Srivastava, M.D.; Srivastava, B.; Brouhard, B. Δ9 Tetrahydrocannabinol and cannabidiol alter cytokine production by human immune cells. Immunopharmacology 1998, 40, 179–185. [Google Scholar] [CrossRef]
- Kishimoto, S.; Kobayashi, Y.; Oka, S.; Gokoh, M.; Waku, K.; Sugiura, T. 2-Arachidonoylglycerol, an endogenous cannabinoid receptor ligand, induces accelerated production of chemokines in HL-60 cells. J. Biochem. 2004, 135, 517–524. [Google Scholar] [CrossRef]
- Anil, S.M.; Shalev, N.; Vinayaka, A.C.; Nadarajan, S.; Namdar, D.; Belausov, E.; Shoval, I.; Mani, K.A.; Mechrez, G.; Koltai, H. Cannabis compounds exhibit anti-inflammatory activity in vitro in COVID-19-related inflammation in lung epithelial cells and pro-inflammatory activity in macrophages. Sci. Rep. 2021, 11, 1–14. [Google Scholar] [CrossRef]
- Mormina, M.; Thakur, S.; Molleman, A.; Whelan, C.J.; Baydoun, A. Cannabinoid signalling in TNF-α induced IL-8 release. Eur. J. Pharmacol. 2006, 540, 183–190. [Google Scholar] [CrossRef] [PubMed]
- Abani, O.; Abbas, A.; Abbas, F.; Abbas, M.; Abbasi, S.; Abbass, H.; Abbott, A.; Abdallah, N.; Abdelaziz, A.; Abdelfattah, M.; et al. Tocilizumab in patients admitted to hospital with COVID-19 (RECOVERY): Preliminary results of a randomised, controlled, open-label, platform trial. Lancet 2021, 397, 1637–1645. [Google Scholar] [CrossRef]
- Gremese, E.; Cingolani, A.; Bosello, S.L.; Alivernini, S.; Tolusso, B.; Perniola, S.; Landi, F.; Pompili, M.; Murri, R.; Santoliquido, A.; et al. Sarilumab use in severe SARS-CoV-2 pneumonia. EClinicalMedicine 2020, 27, 100553. [Google Scholar] [CrossRef]
- Gritti, G.; Raimondi, F.; Ripamonti, D.; Riva, I.; Landi, F.; Alborghetti, L.; Frigeni, M.; Damiani, M.; Micò, C.; Fagiuoli, S.; et al. IL-6 signalling pathway inactivation with siltuximab in patients with COVID-19 respiratory failure: An observational cohort study. medRxiv 2018, 1–3. [Google Scholar] [CrossRef]
- FDA ok for Phase III Trial of Siltuximab for COVID-19. Available online: https://www.thepharmaletter.com/article/fda-ok-for-phase-iii-trial-of-siltuximab-for-covid-19 (accessed on 8 May 2021).
- Khodadadi, H.; Salles, L.; Jarrahi, A.; Chibane, F.; Costigliola, V.; Yu, J.C.; Vaibhav, K.; Hess, D.C.; Dhandapani, K.M.; Baban, B. Cannabidiol Modulates Cytokine Storm in Acute Respiratory Distress Syndrome Induced by Simulated Viral Infection Using Synthetic RNA. Cannabis Cannabinoid Res. 2020, 5, 197–201. [Google Scholar] [CrossRef]
- Salles, L.; Khodadadi, H.; Jarrahi, A.; Ahluwalia, M.; Paffaro, V.A.; Costigliola, V.; Yu, J.C.; Hess, D.C.; Dhandapani, K.M.; Baban, B. Cannabidiol (CBD) modulation of apelin in acute respiratory distress syndrome. J. Cell. Mol. Med. 2020, 24, 12869–12872. [Google Scholar] [CrossRef] [PubMed]
- Lowe, H.; Steele, B.; Bryant, J.; Toyang, N.; Ngwa, W. Non-Cannabinoid Metabolites of Cannabis sativa L. with Therapeutic Potential. Plants 2021, 10, 400. [Google Scholar] [CrossRef] [PubMed]
- Ngwa, W.; Kumar, R.; Thompson, D.; Lyerly, W.; Moore, R.; Reid, T.-E.; Lowe, H.; Toyang, N. Potential of Flavonoid-Inspired Phytomedicines against COVID-19. Molecules 2020, 25, 2707. [Google Scholar] [CrossRef]
- Mohammed, A.; Alghetaa, H.K.; Zhou, J.; Chatterjee, S.; Nagarkatti, P.; Nagarkatti, M. Protective effects of Δ9-tetrahydrocannabinol against enterotoxin-induced acute respiratory distress syndrome are mediated by modulation of microbiota. Br. J. Pharmacol. 2020, 177, 5078–5095. [Google Scholar] [CrossRef]
- Zgair, A.; Lee, J.B.; Wong, J.C.M.; Taha, D.; Aram, J.; Di Virgilio, D.; McArthur, J.W.; Cheng, Y.-K.; Hennig, I.M.; Barrett, D.A.; et al. Oral administration of cannabis with lipids leads to high levels of cannabinoids in the intestinal lymphatic system and prominent immunomodulation. Sci. Rep. 2017, 7, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Chatow, L.; Nudel, A.; Nesher, I.; Hemo, D.H.; Rozenberg, P.; Voropaev, H.; Winkler, I.; Levy, R.; Kerem, Z.; Yaniv, Z.; et al. In Vitro Evaluation of the Activity of Terpenes and Cannabidiol against Human Coronavirus E229. Life 2021, 11, 290. [Google Scholar] [CrossRef] [PubMed]
- Crossney, J. A Storm of Research Activity into Cannabis and Coronavirus a Closer Look at Expanding Research of Cannabinoids and Terpenes in COVID-19. Available online: https://www.cannabissciencetech.com/view/a-storm-of-research-activity-into-cannabis-and-coronavirus-a-closer-look-at-expanding-research-of-cannabinoids-and-terpenes-in-COVID-19 (accessed on 5 May 2021).
- Raj, V.; Park, J.G.; Cho, K.-H.; Choi, P.; Kim, T.; Ham, J.; Lee, J. Assessment of antiviral potencies of cannabinoids against SARS-CoV-2 using computational and in vitro approaches. Int. J. Biol. Macromol. 2020, 168, 474–485. [Google Scholar] [CrossRef] [PubMed]
- Mishra, D.; Maurya, R.R.; Kumar, K.; Munjal, N.S.; Bahadur, V.; Sharma, S.; Singh, P.; Bahadur, I. Structurally modified compounds of hydroxychloroquine, remdesivir and tetrahydrocannabinol against main protease of SARS-CoV-2, a possible hope for COVID-19: Docking and molecular dynamics simulation studies. J. Mol. Liq. 2021, 335, 116185. [Google Scholar] [CrossRef]
- Sarkar, I.; Sen, G.; Bhattyachariya, M.; Bhattacharyya, S.; Sen, A. In silico inquest reveals the efficacy of Cannabis in the treatment of post-Covid-19 related neurodegeneration. J. Biomol. Struct. Dyn. 2021, 1–10. [Google Scholar] [CrossRef]
- Kielian, M. Enhancing host cell infection by SARS-CoV-2. Science 2020, 370, 765–766. [Google Scholar] [CrossRef]
- Orio, L.P.; Boschin, G.; Recca, T.; Morelli, C.F.; Ragona, L.; Francescato, P.; Arnoldi, A.; Speranza, G. New ACE-Inhibitory Peptides from Hemp Seed (Cannabis sativa L.) Proteins. J. Agric. Food Chem. 2017, 65, 10482–10488. [Google Scholar] [CrossRef]
- Tadayon, N.; Ramazani, A. A review on the syntheses of Dronabinol and Epidiolex as classical cannabinoids with various biological activities including those against SARS-COV2. J. Iran. Chem. Soc. 2021, 18, 2517–2534. [Google Scholar] [CrossRef]
- Nguyen, L.C.; Yang, D.; Nicolaescu, V.; Best, T.J.; Ohtsuki, T.; Chen, S.-N.; Friesen, J.B.; Drayman, N.; Mohamed, A.; Dann, C.; et al. Cannabidiol Inhibits SARS-CoV-2 Replication and Promotes the Host Innate Immune Response. bioRxiv 2021. [Google Scholar] [CrossRef]
- Bilal, M.; Rasheed, T.; Iqbal, H.M.; Hu, H.; Wang, W.; Zhang, X. Macromolecular agents with antimicrobial potentialities: A drive to combat antimicrobial resistance. Int. J. Biol. Macromol. 2017, 103, 554–574. [Google Scholar] [CrossRef] [PubMed]
- Cortes, E.; Mora, J.; Márquez, E. Modelling the Anti-Methicillin-Resistant Staphylococcus Aureus (MRSA) Activity of Cannabinoids: A QSAR and Docking Study. Crystals 2020, 10, 692. [Google Scholar] [CrossRef]
- Hurdle, J.G.; O’neill, A.J.; Chopra, I.; Lee, R.E. Targeting bacterial membrane function: An underexploited mechanism for treating persistent infections. Nat. Rev. Microbiol. 2011, 9, 62–75. [Google Scholar] [CrossRef]
- Hyldgaard, M.; Mygind, T.; Meyer, R.L. Essential Oils in Food Preservation: Mode of Action, Synergies, and Interactions with Food Matrix Components. Front. Microbiol. 2012, 3, 12. [Google Scholar] [CrossRef] [PubMed]
- Helander, I.M.; Alakomi, H.-L.; Latva-Kala, K.; Mattila-Sandholm, T.; Pol, I.; Smid, E.J.; Gorris, L.G.M.; von Wright, A. Characterization of the Action of Selected Essential Oil Components on Gram-Negative Bacteria. J. Agric. Food Chem. 1998, 46, 3590–3595. [Google Scholar] [CrossRef]
- Nazzaro, F.; Fratianni, F.; DE Martino, L.; Coppola, R.; De Feo, V. Effect of Essential Oils on Pathogenic Bacteria. Pharmaceuticals 2013, 6, 1451–1474. [Google Scholar] [CrossRef]
- Djilani, A.; Dicko, A. The Therapeutic Benefits of Essential Oils. In Nutrition, Well-Being and Health; Bouayed, J., Ed.; InTech: London, UK, 2012. [Google Scholar] [CrossRef]
- Jassim, S.; Naji, M. Novel antiviral agents: A medicinal plant perspective. J. Appl. Microbiol. 2003, 95, 412–427. [Google Scholar] [CrossRef] [PubMed]
- Leizer, C.; Ribnicky, D.; Poulev, A.; Dushenkov, S.; Raskin, I. The Composition of Hemp Seed Oil and Its Potential as an Important Source of Nutrition. J. Nutraceuticals Funct. Med. Foods 2000, 2, 35–53. [Google Scholar] [CrossRef]
- Cowan, M.M. Plant Products as Antimicrobial Agents. Clin. Microbiol. Rev. 1999, 12, 564–582. [Google Scholar] [CrossRef]
- Nostro, A.; Germano, M.; D’Angelo, V.; Marino, A.; Cannatelli, M. Extraction methods and bioautography for evaluation of medicinal plant antimicrobial activity. Lett. Appl. Microbiol. 2000, 30, 379–384. [Google Scholar] [CrossRef]
- Valgas, C.; De Souza, S.M.; Smânia, E.F.A.; Smânia, A., Jr. Screening methods to determine antibacterial activity of natural products. Braz. J. Microbiol. 2007, 38, 369–380. [Google Scholar] [CrossRef]
- Novak, J.; Franz, C. Composition of the Essential Oils and Extracts of Two Populations ofCannabis sativaL. ssp.spontaneafrom Austria. J. Essent. Oil Res. 2003, 15, 158–160. [Google Scholar] [CrossRef]
- Hao, X.M.; Yang, Y.; An, L.X.; Wang, J.M.; Han, L. Study on Antibacterial Mechanism of Hemp Fiber. Adv. Mater. Res. 2014, 887–888, 610–613. [Google Scholar] [CrossRef]
- Khan, B.A.; Wang, J.; Warner, P.; Wang, H. Antibacterial properties of hemp hurd powder against E. coli. J. Appl. Polym. Sci. 2014, 132, 41588. [Google Scholar] [CrossRef]
- Karas, J.A.; Wong, L.J.M.; Paulin, O.K.A.; Mazeh, A.C.; Hussein, M.H.; Li, J.; Velkov, T. The Antimicrobial Activity of Cannabinoids. Antibiotics 2020, 9, 406. [Google Scholar] [CrossRef] [PubMed]
- Feldman, M.; Sionov, R.; Smoum, R.; Mechoulam, R.; Ginsburg, I.; Steinberg, D. Comparative Evaluation of Combinatory Interaction between Endocannabinoid System Compounds and Poly-L-lysine against Streptococcus mutans Growth and Biofilm Formation. BioMed Res. Int. 2020, 2020, 7258380–7258387. [Google Scholar] [CrossRef]
- ZhengHai, Z.; Yan, D.; YanRu, J.; QingLi, Y.; ZhenWei, L. Antibacterial activity and stability of extract from hemp (Cannabis sativa L.) leaves. J. Food Saf. Qual. 2019, 10, 927–933. [Google Scholar]
- Dhifi, W.; Bellili, S.; Jazi, S.; Bahloul, N.; Mnif, W. Essential Oils’ Chemical Characterization and Investigation of Some Biological Activities: A Critical Review. Medicines 2016, 3, 25. [Google Scholar] [CrossRef] [PubMed]
- Vasudevan, K.; Stahl, V. Cannabinoids infused mouthwash products are as effective as chlorhexidine on inhibition of total-culturable bacterial content in dental plaque samples. J. Cannabis Res. 2020, 2, 1–9. [Google Scholar] [CrossRef]
- Raina, S.; Thakur, A.; Sharma, A.; Pooja, D.; Minhas, A.P. Bactericidal activity of Cannabis sativa phytochemicals from leaf extract and their derived Carbon Dots and Ag@Carbon Dots. Mater. Lett. 2019, 262, 127122. [Google Scholar] [CrossRef]
- Stahl, V.; Vasudevan, K. Comparison of Efficacy of Cannabinoids versus Commercial Oral Care Products in Reducing Bacterial Content from Dental Plaque: A Preliminary Observation. Cureus 2020, 12, e6809. [Google Scholar] [CrossRef]
- Guimaraes, A.R.D.; Peres, M.A.; Vieira, R.D.S.; Ferreira, R.M.; Ramos-Jorge, M.L.; Apolinario, S.; Debom, A. Self-perception of side effects by adolescents in a chlorhexidine-fluoride-based preventive oral health program. J. Appl. Oral Sci. 2006, 14, 291–296. [Google Scholar] [CrossRef][Green Version]
- Schirone, M.; Visciano, P.; Tofalo, R.; Suzzi, G. Editorial: Foodborne Pathogens: Hygiene and Safety. Front. Microbiol. 2019, 10, 1974. [Google Scholar] [CrossRef]
- Cabeça, T.K.; Pizzolitto, A.C.; Pizzolitto, E.L. Activity of disinfectants against foodborne pathogens in suspension and adhered to stainless steel surfaces. Braz. J. Microbiol. 2012, 43, 1112–1119. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Langsrud, S.; Sidhu, M.S.; Heir, E.; Holck, A.L. Bacterial disinfectant resistance—a challenge for the food industry. Int. Biodeterior. Biodegrad. 2003, 51, 283–290. [Google Scholar] [CrossRef]
- Park, K.M.; Yoon, S.-G.; Choi, T.-H.; Kim, H.J.; Park, K.J.; Koo, M. The Bactericidal Effect of a Combination of Food-Grade Compounds and their Application as Alternative Antibacterial Agents for Food Contact Surfaces. Foods 2020, 9, 59. [Google Scholar] [CrossRef] [PubMed]
- Colagiorgi, A.; Bruini, I.; Di Ciccio, P.A.; Zanardi, E.; Ghidini, S.; Ianieri, A. Listeria monocytogenes Biofilms in the Wonderland of Food Industry. Pathogens 2017, 6, 41. [Google Scholar] [CrossRef]
- Krawczyk-Balska, A.; Markiewicz, Z. The intrinsic cephalosporin resistome of Listeria monocytogenes in the context of stress response, gene regulation, pathogenesis and therapeutics. J. Appl. Microbiol. 2015, 120, 251–265. [Google Scholar] [CrossRef]
- Doulgeraki, A.I.; Di Ciccio, P.; Ianieri, A.; Nychas, G.-J. Methicillin-resistant food-related Staphylococcus aureus: A review of current knowledge and biofilm formation for future studies and applications. Res. Microbiol. 2017, 168, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Kurtzman, C.P. Torulaspora Lindner (1904). In The Yeasts; Elsevier B.V.: Amsterdam, The Netherlands, 2011; Volume 2, pp. 867–874. ISBN 9780444521491. [Google Scholar]
- Kuanyshev, N.; Adamo, G.M.; Porro, D.; Branduardi, P. The spoilage yeastZygosaccharomyces bailii: Foe or friend? Yeast 2017, 34, 359–370. [Google Scholar] [CrossRef] [PubMed]
- Pasquali, F.; Schinzari, M.; Lucchi, A.; Mandrioli, M.; Toschi, T.G.; De Cesare, A.; Manfreda, G. Preliminary data on the antimicrobial effect of Cannabis sativa L. variety Futura 75 against food-borne pathogens in vitro as well as against naturally occurring microbial populations on minced meat during storage. Ital. J. Food Saf. 2020, 9, 8581. [Google Scholar] [CrossRef]
- Doehlemann, G.; Ökmen, B.; Zhu, W.; Sharon, A. Plant Pathogenic Fungi. Fungal Kingd. 2017, 701–726. [Google Scholar] [CrossRef]
- Ingold, C.T.; Hudson, H.J. Fungi as Plant Pathogens. In The Biology of Fungi; Ingold, C.T., Ed.; Springer: Dordrecht, The Netherlands, 1993; pp. 159–182. [Google Scholar]
- Yang, J.; Hsiang, T.; Bhadauria, V.; Chen, X.-L.; Li, G. Plant Fungal Pathogenesis. BioMed Res. Int. 2017, 2017, 1–2. [Google Scholar] [CrossRef] [PubMed]
- Wightwick, A.; Walters, R.; Allinson, G.; Reichman, S.; Menzies, S.R.A.N. Environmental Risks of Fungicides Used in Horticultural Production Systems. In Fungicides; Carisse, O., Ed.; BoD—Books on Demand: Norderstedt, Germany, 2010. [Google Scholar] [CrossRef]
- Zubrod, J.P.; Bundschuh, M.; Arts, G.; Brühl, C.A.; Imfeld, G.; Knäbel, A.; Payraudeau, S.; Rasmussen, J.J.; Rohr, J.; Scharmüller, A.; et al. Fungicides: An Overlooked Pesticide Class? Environ. Sci. Technol. 2019, 53, 3347–3365. [Google Scholar] [CrossRef] [PubMed]
- Tapwal, A.; Nisha; Garg, S.; Gautam, N.; Kumar, R. In Vitro antifungal potency of plant extracts against five phytopathogens. Braz. Arch. Biol. Technol. 2011, 54, 1093–1098. [Google Scholar] [CrossRef]
- Garcia-Aroca, T.; Doyle, V.; Singh, R.; Price, T.; Collins, K. First Report of Curvularia Leaf Spot of Corn, Caused by Curvularia lunata, in the United States. Plant Health Prog. 2018, 19, 140–142. [Google Scholar] [CrossRef]
- Haroun, N.E.; Elamin, S.E.; Mahgoub, B.M.; Elssidig, M.A.; Mohammed, E.H. Leaf blight: A new disease of Xanthium strumarium L. caused by Curvularia lunata and Drechslera spicifera in Sudan. Int. J. Curr. Microbiol. Appl. Sci. 2015, 4, 511–515. [Google Scholar]
- Msikita, W.; Baimey, H.; James, B.D. Severity of Curvularia Stem Blight Disease of Cassava in West Africa. Plant Dis. 2007, 91, 1430–1435. [Google Scholar] [CrossRef]
- Verma, V.S.; Gupta, V.K. First Report of Curvularia lunata Causing Root Rot of Strawberry in India. Plant Dis. 2010, 94, 477. [Google Scholar] [CrossRef]
- Farooq, M.A.; Iqbal, U.; Iqbal, S.M.; Afzal, R.; Rasool, A. Invitro evaluation of different plant extracts on mycelial growth of sclerotium rolfsii the cause of root rot of sugar beet. Mycopath 2010, 8, 8184. [Google Scholar]
- Pal, G.K.; Kumar, B.; Shani, S.K. Antifungal activity of some seed extracts against seed-borne phytopathogenic fungi Alternaria Spp. Int. J. Univers. Pharm. Life Sci. 2013, 3, 6–14. [Google Scholar]
- Aphajal, M.; Jaish, B.M. In vitro Antifungal activity of somee higher plant extracts against Alternaria brassicae (Berk.) sacc. and A. brassicicola (Schw.) Wiltsh. Bull. Pure Appl. Sci. Bot. 2018, 37b, 108. [Google Scholar] [CrossRef]
- Thomma, B.P.H.J. Alternariaspp.: From general saprophyte to specific parasite. Mol. Plant Pathol. 2003, 4, 225–236. [Google Scholar] [CrossRef] [PubMed]
- Charkowski, A.O. Biology and control of Pectobacterium in potato. Am. J. Potato Res. 2015, 92, 223–229. [Google Scholar] [CrossRef]
- Bao, Q.; Liu, H.; Fu, K.; Zhang, C.; Wang, C.; Feng, Y.; Yang, L.; Lao, H.; Ren, Q. Hemp Bast Fiber Extract with Antibacterial Activity, Preparation Method and Application of Hemp Bast Fiber Extract 2014. CN201410249611.XA, 2 March 2016. [Google Scholar]
- Cassano, R.; Trombino, S.; Ferrarelli, T.; Nicoletta, F.P.; Mauro, M.V.; Giraldi, C.; Picci, N. Hemp fiber (Cannabis sativa L.) derivatives with antibacterial and chelating properties. Cellulose 2013, 20, 547–557. [Google Scholar] [CrossRef]
- Fang, G.L. Surgical Sewing Free Zipper Made of Antibiotic Material Hemp Fiber 2005. CN2829641Y, 25 October 2006. [Google Scholar]
- Andre, C.M.; Hausman, J.-F.; Guerriero, G. Cannabis sativa: The Plant of the Thousand and One Molecules. Front. Plant Sci. 2016, 7, 19. [Google Scholar] [CrossRef] [PubMed]
- Khan, B.A.; Warner, P.; Wang, H. Antibacterial Properties of Hemp and Other Natural Fibre Plants: A Review. BioResources 2014, 9, 3642–3659. [Google Scholar] [CrossRef]
- Stasiłowicz, A.; Tomala, A.; Podolak, I.; Cielecka-Piontek, J. Cannabis sativa L. as a Natural Drug Meeting the Criteria of a Multitarget Approach to Treatment. Int. J. Mol. Sci. 2021, 22, 778. [Google Scholar] [CrossRef]
- Stott, C.G.; Guy, G.W. Cannabinoids for the pharmaceutical industry. Euphytica 2004, 140, 83–93. [Google Scholar] [CrossRef]
- McKenna, G.J. The Current Status of Medical Marijuana in the United States. Hawaii J. Med. Public Health 2014, 73, 105–108. [Google Scholar]
- Koltai, H.; Poulin, P.; Namdar, D. Promoting cannabis products to pharmaceutical drugs. Eur. J. Pharm. Sci. 2019, 132, 118–120. [Google Scholar] [CrossRef]
- Benito, S.B.; Seijo-Vila, M.; Caro-Villalobos, M.; Tundidor, I.; Andradas, C.; García-Taboada, E.; Wade, J.; Smith, S.; Guzmán, M.; Pérez-Gómez, E.; et al. Appraising the “entourage effect”: Antitumor action of a pure cannabinoid versus a botanical drug preparation in preclinical models of breast cancer. Biochem. Pharmacol. 2018, 157, 285–293. [Google Scholar] [CrossRef]
- Nahtigal, I.; Blake, A.; Hand, A.; Florentinus-Mefailoski, A.; Sohi, H.H.; Friedberg, J. The pharmacological properties of cannabis. In Pain Management Yearbook 2016; Merrick, J., Ed.; Nova Science Publishers, Inc.: New York, NY, USA, 2016; pp. 481–491. ISBN 978-1-53610-949-8. [Google Scholar]
- Sharma, P.; Murthy, P.; Bharath, M.S. Chemistry, Metabolism, and Toxicology of Cannabis: Clinical Implications. Iran. J. Psychiatry 2012, 7, 149–156. [Google Scholar]
- Tamilselvan, N.; Thirumalai, T.; Shyamala, P.; David, E. A review on some poisonous plants and their medicinal values. J. Acute Dis. 2014, 3, 85–89. [Google Scholar] [CrossRef]
- Borgelt, L.M.; Franson, K.L.; Nussbaum, A.; Wang, G.S. The Pharmacologic and Clinical Effects of Medical Cannabis. Pharmacother. J. Hum. Pharmacol. Drug Ther. 2013, 33, 195–209. [Google Scholar] [CrossRef] [PubMed]
- Pertwee, R.G.; Howlett, A.C.; Abood, M.E.; Alexander, S.P.; Di Marzo, V.; Elphick, M.R.; Greasley, P.J.; Hansen, H.S.; Kunos, G.; Mackie, K.; et al. International Union of Basic and Clinical Pharmacology. LXXIX. Cannabinoid Receptors. Pharmacol. Rev. 2010, 62, 588–631. [Google Scholar] [CrossRef] [PubMed]
- Uliss, D.B.; Dalzell, H.C.; Handrick, G.R.; Howes, J.F.; Razdan, R.K. Hashish. Importance of the phenolic hydroxyl group in tetrahydrocannabinols. J. Med. Chem. 1975, 18, 213–215. [Google Scholar] [CrossRef]
- Vaughn, D.; Kulpa, J.; Paulionis, L. Preliminary Investigation of the Safety of Escalating Cannabinoid Doses in Healthy Dogs. Front. Vet. Sci. 2020, 7, 51. [Google Scholar] [CrossRef]
- Deiana, S.; Watanabe, A.; Yamasaki, Y.; Amada, N.; Arthur, M.; Fleming, S.; Woodcock, H.; Dorward, P.; Pigliacampo, B.; Close, S.; et al. Plasma and brain pharmacokinetic profile of cannabidiol (CBD), cannabidivarine (CBDV), Δ9-tetrahydrocannabivarin (THCV) and cannabigerol (CBG) in rats and mice following oral and intraperitoneal administration and CBD action on obsessive–compulsive behaviour. Psychopharmacology 2011, 219, 859–873. [Google Scholar] [CrossRef]
- Satyal, P.; Setzer, W.N. Chemotyping and Determination of Antimicrobial, Insecticidal, and Cytotoxic Properties of Wild- Grown Cannabis sativa from Nepal. J. Med. Act. Plants J. Med. Act. Plants J. Med. Act. Plants 2014, 3, 9–169. [Google Scholar] [CrossRef]
- Zuardi, A.W.; Crippa, J.A.S.; Hallak, J.E.; Bhattacharyya, S.; Atakan, Z.; Martin-Santos, R.; McGuire, P.; Guimaraes, F.S. A Critical Review of the Antipsychotic Effects of Cannabidiol: 30 Years of a Translational Investigation. Curr. Pharm. Des. 2012, 18, 5131–5140. [Google Scholar] [CrossRef]
- Thompson, G.R.; Rosenkrantz, H.; Schaeppi, U.H.; Braude, M.C. Comparison of acute oral toxicity of cannabinoids in rats, dogs and monkeys. Toxicol. Appl. Pharmacol. 1973, 25, 363–372. [Google Scholar] [CrossRef]
- Beaulieu, P. Toxic effects of cannabis and cannabinoids: Animal data. Pain Res. Manag. 2005, 10, 23A–26A. [Google Scholar] [CrossRef]
- Di, L.; Kerns, E.; Carter, G. Drug-Like Property Concepts in Pharmaceutical Design. Curr. Pharm. Des. 2009, 15, 2184–2194. [Google Scholar] [CrossRef] [PubMed]
- Kerns, E.H.; Di, L. Advantages of Good Drug-like Properties. In Drug-Like Properties: Concepts, Structure Design and Methods from ADME to Toxicity Optimization; Kerns, E.H., Di, L., Eds.; Academic Press: Burlington, VT, USA, 2016; pp. 6–16. ISBN 978-0-1236-9520-8. [Google Scholar]
- Landmark, C.J.; Brandl, U. Pharmacology and drug interactions of cannabinoids. Epileptic Disord. 2020, 22, 16–22. [Google Scholar]
- Prandi, C.; Blangetti, M.; Namdar, D.; Koltai, H. Structure-Activity Relationship of Cannabis Derived Compounds for the Treatment of Neuronal Activity-Related Diseases. Molecules 2018, 23, 1526. [Google Scholar] [CrossRef] [PubMed]
- Furqan, T.; Batool, S.; Habib, R.; Shah, M.; Kalasz, H.; Darvas, F.; Kuca, K.; Nepovimova, E.; Batool, S.; Nurulain, S.M. Cannabis Constituents and Acetylcholinesterase Interaction: Molecular Docking, In Vitro Studies and Association with CNR1 rs806368 and ACHE rs17228602. Biomolecules 2020, 10, 758. [Google Scholar] [CrossRef]
- Filloux, F.M. Cannabinoids for pediatric epilepsy? Up in smoke or real science? Transl. Pediatr. 2015, 4, 271–282. [Google Scholar] [CrossRef]
- Desa, S.; Osman, A.; Hyslop, R. In Silico Assessment of Drug-Like Properties of Phytocannabinoids in Cannabis Sativa. Educ. J. Sci. Math. Technol. 2017, 4, 1–7. [Google Scholar] [CrossRef]
- Wiley, J.L.; Martin, B.R. Cannabinoid pharmacological properties common to other centrally acting drugs. Eur. J. Pharmacol. 2003, 471, 185–193. [Google Scholar] [CrossRef]
- Anonymous. New Antimicrobial Data and Conference Presentation. Available online: https://www.asx.com.au/asxpdf/20200908/pdf/44mf662gw8dbkd.pdf (accessed on 28 July 2021).
- Kocis, P.; Vrana, K.E. Delta-9-Tetrahydrocannabinol and Cannabidiol Drug-Drug Interactions. Med Cannabis Cannabinoids 2020, 3, 61–73. [Google Scholar] [CrossRef] [PubMed]
- Parmar, J.R.; Forrest, B.D.; Freeman, R.A. Medical marijuana patient counseling points for health care professionals based on trends in the medical uses, efficacy, and adverse effects of cannabis-based pharmaceutical drugs. Res. Soc. Adm. Pharm. 2015, 12, 638–654. [Google Scholar] [CrossRef] [PubMed]
- Cabral, G.A.; Pettit, D.A.D. Drugs and immunity: Cannabinoids and their role in decreased resistance to infectious disease. J. Neuroimmunol. 1998, 83, 116–123. [Google Scholar] [CrossRef]
- Anonymous. FDA Grants BTX 1801 Qualified Infectious Disease Product Designation Status. Available online: https://yourir.info/resources/3f148bb5dfccdf8f/announcements/bot.asx/6A976285/BOT_FDA_Grants_BTX_1801_QIDP_Designation_Status.pdf?embed=1 (accessed on 28 July 2021).





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Pathogen | Compound/Extract/EO | Activity | Reference Antibiotic | Ref | |
|---|---|---|---|---|---|
| Antibiotic | Activity | ||||
| Gram+ve | |||||
| Enterococcus faecium | EO, α-humulene, α-pinene, β-pinene, myrcene | MIC 0.75–1.87 (%v/v) MBC 1.39–2.83 (%v/v) | [93] | ||
| E. faecium | EO, α-humulene, α-pinene, β-pinene, myrcene | MIC 1–4 µg/mL | Ciprofloxacin | MIC 8 µg/mL | [94] |
| EMRSA 15 and EMRSA 16 | CBD, THC, CBG, CBC, CBN | MIC 0.5–2.0 µg/mL | [95] | ||
| MRSA | 4-acetoxy-2-geranyl-5-hydroxy-3-n-pentylphenol and 8-hydroxycannabinolic acid A | IC50 6.7 µM | Ciprofloxacin | IC50 0.4 µM | [96] |
| MRSA | CVDVM | MIC 15.6 µM | [52] | ||
| MRSA | CBCA | MIC 3.9 µM | [52] | ||
| MRSA | CBD | MIC 1 µg/mL | Tobramycin, Meropenem, Ofloxacin | MIC 1, 16, 64 µg/mL (respectively) | [50] |
| MRSA | CBD | MBEC 2–4 µg/mL | [49] | ||
| MRSA | CBD analogs | MIC 0.25–64.0 µg/mL | Vancomycin, Daptomycin, Mupirocin | MIC 0.125–2.0 µg/mL | [49] |
| MRSA | CBD, CBN, CBC, CBDV and Δ1 & 9-THC | IC50 5.8–10.6 µM | Ciprofloxacin | IC50 9.33 µM | [97] |
| MRSA | CBDA | MIC 4 µg/mL | Tobramycin, Meropenem, Ofloxacin | MIC 1, 16, 64 µg/mL (respectively) | [50] |
| MRSA | CBG | MIC 2 µg/mL and MBEC 4 µg/mL | [43] | ||
| MRSA | EO | IC50 0.82–4.22 µg/mL | [98] | ||
| MRSA, VISA, VRSA, E. faecium | CBD | MIC 1–2 µg/mL | Vancomycin, Daptomycin, Trimethoprim, Mupirocin, Clindamycin | MIC 0.125 to >64 µg/mL | [49] |
| Streptococcus pneumoniae | CBD | MIC 1–4 µg/mL | Vancomycin, Daptomycin, Trimethoprim, Mupirocin, Clindamycin | MIC 0.25 to >64 µg/mL | [49] |
| VRE | CBCA | MIC 7.8 µM | [52] | ||
| Gram -ve | |||||
| Escherichia coli | Aqueous extract | MIC 7.14 mg/mL | Ciprofloxacin | MIC < 0.12 mg/mL | [99] |
| E. coli | N-p-trans-coumaroyl-tyramine | IC50 0.8 µg/mL | Ciprofloxacin | IC50 0.01 µg/mL | [100] |
| E. coli | Seed extract | MIC 25 µg/mL | [67] | ||
| E. coli and Salmonella typhimurium | Seed extract | Growth inhibition at 1 mg/mL | [101] | ||
| E. coli, and Pseudomonas aeruginosa | EO | MIC 1.2 mg/mL | MIC 0.062–1.0 mg/mL | [60] | |
| Enterobacter aerogenes | Seed extract | MIC 2.5 mg/mL | [101] | ||
| Neisseria gonorrhoeae | CBD | MIC 1–2 µg/mL | Vancomycin, Levofloxacin, Meropenem, Gentamicin | MIC 0.002–4.0 µg/mL | [49] |
| N. gonorrhoeae | CBD analogs | MIC 0.03–16.0 µg/mL | Mupirocin Colistin | MIC 1–32 µg/mL | [49] |
| P. aeruginosa | Aqueous extract | MIC 7.14 mg/mL | Ciprofloxacin | MIC 1.23 mg/mL | [99] |
| P. aeruginosa | Whole plant extract | MIC 12.5 µg/mL | [67] | ||
| Pathogen | Compound/Extract/EO | Activity | Reference Antibiotic | Ref | |
|---|---|---|---|---|---|
| Antibiotic | Activity | ||||
| Gram+ve | |||||
| Bacillus subtilis and Staphylococcus aureus | Leaf extract | MIC 1.56 mg/mL | [81] | ||
| B. subtilis, S. aureus and Micrococcus luteus | EO | MIC 1.2–4.7 mg/mL | Ciprofloxacin | MIC 0.015–0.031 mg/mL | [60] |
| B. subtilis, S. aureus, Mycobacterium smegmatis | CBC, its homologs and isomers | MIC 0.39–3.12 µg/mL | [112] | ||
| Clostridium species *, Enterococcus hirae *, Streptococcus salivarius * | EO, α-humulene, α-pinene, β-pinene, myrcene | MIC ≥ 0.8 (%v/v) | [93] | ||
| Enterococcus *, Staphylococcus *, and Bacillus species * | EO | MIC ≥ 0.5 µg/mL | Ampicillin, Ciprofloxacin | MIC ≥ 0.25 µg/mL | [94] |
| Listeria monocytogenes strains * | EO | MIC/MBC 2.5–5.0 μL/mL | [110] | ||
| L. monocytogenes * | EO | MIC ≥ 1 µg/mL | Ampicillin | MIC ≥ 0.25 µg/mL | [94] |
| L. monocytogenes * | EO, α-pinene, Myrcene | MBC ≥ 1024 µg/mL | [111] | ||
| Lancefield Group A Streptococcus sp. | Leaf extract | MIC 20 mg/mL MBC 30 mg/mL | [113] | ||
| MRSA biofilms * | Seed extract | MIC 1 mg/mL | [101] | ||
| MSSA | CBCA | MIC 7.8 µM | [52] | ||
| MSSA, VISE, Staphylococcus epidermidis, Staphylococcus pyogenes, Enterococcus faecalis, Cutibacterium acnes, Clostridioides difficile | CBD | MIC 0.5–4.0 µg/mL | Vancomycin, Daptomycin, Trimethoprim, Mupirocin, Clindamycin, Levofloxacin, Meropenem, Gentamicin, Erythromycin, Tetracycline, Mupirocin | MIC 0.03–64.0 µg/mL | [49] |
| Mycobacterium intracellulare | CBG | IC50 15 µg/mL | [114] | ||
| S. aureus | 4-acetoxy-2-geranyl-5-hydroxy-3-n-pentylphenol, 8-hydroxycannabinolic acid A | IC50 3.5 µM | Ciprofloxacin | IC50 0.4 µM | [96] |
| S. aureus | Aqueous extract | MIC 3.57 mg/mL | Ciprofloxacin | MIC 0.62 µg/mL | [99] |
| S. aureus | Methanol extract | MIC 25 µg/mL | [67] | ||
| S. aureus (including multi drug resistant S. aureus 104) | EO | MIC 8 mg/mL | [61] | ||
| S. aureus (mature and pre-formed biofilms) | EO | MBEC 24 mg/mL | [61] | ||
| S. aureus and E. faecalis | Seed extract | MIC 1 mg/mL | [101] | ||
| S. aureus biofilm * | EO | MIC 0.5 mg/mL | [101] | ||
| S. aureus planktonic cells * | EO | MIC 1 mg/mL | [101] | ||
| S. aureus * | EO | MIC 1.25–5.0 µg/mL | [110] | ||
| S. aureus * | EO | MIC 1–4 µg/mL | Ciprofloxacin | MIC 0.5–16.0 µg/mL | [94] |
| S. aureus, S. epidermidis | CBD, CBDA | MIC 1–4 µg/mL | Torbamycin, Meropenem, Ofloxacin | MIC 0.06–0.5 µg/mL | [50] |
| SA-1199B (MDR), RN4220 (Macrolide-resistant), XU212 (Tetracycline-resistant) | CBD, CBC, THC, CBG, CBN, Carboxylated versions, Abnormal cannabinoids | MIC 0.5–4.0 µg/mL | [95] | ||
| Staphylococcus species | THC, CBD | MIC 1–5 µg/mL | [115] | ||
| Staphylococcus, Lactococcus and Bacillus species | CBD, CBN, CBC, CBDV and Δ1 & 9-THC | IC50 2.6–9.2 µM | Ciprofloxacin | IC50 0.003–2.4 µM | [97] |
| Gram-ve | |||||
| Moraxella catarrhalis, Neisseria meningitidis and Legionella pneumophila | CBD | MIC 0.25–1.0 µg/mL | Vancomycin, Levofloxacin, Meropenem, Gentamicin | MIC 0.03–32 µg/mL | [49] |
| Pectobacterium carotovorum subsp. carotovorum * | EO, α-humulene, α-pinene, β-pinene, myrcene | MIC ≥ 1.24 (%v/v) | [93] | ||
| Pseudomonas fluorescens and Xanthobacter flavus | CBD, CBN, CBC, CBDV and Δ1 & 9-THC | IC50 3.1–9.3 µM | Ciprofloxacin | IC50 0.15–2.3 µM | [97] |
| Pseudomonas species | EO(s) and Terpenes | MIC 1.05–1.97 (%v/v) | [93] | ||
| Pathogen | Compound/Extract/EO | Activity | Reference Antibiotic | Ref | |
|---|---|---|---|---|---|
| Antibiotic | Activity | ||||
| Candida albicans | Extract | MIC 0.25 mg/mL | [124] | ||
| C. albicans | Extract | MIC 1.42 mg/mL | Fluconazole | MIC 2 mg/mL | [99] |
| C. albicans | 4-terpenyl cannabinolate | MIC 8.5 µg/mL | [125] | ||
| C. albicans | 8-hydroxycannabinol | IC50 4.6 µM | Amphotericin B | IC50 0.3 µM | [96] |
| C. albicans | Cannabis and ginger blend | MIC 4.69 mg/mL | [126] | ||
| C. albicans | CBDV | IC50 11.9 mM | Nystatin | IC50 1.50 mM | [97] |
| C. albicans | CBNA | IC50 8.5 µg/mL | [125] | ||
| Candida krusei | Cannabinoids | IC50 53.4–60.5 µM | amphotericin B | IC50 0.7 µM | [96] |
| Candida neoformans | β-caryophyllene/oxide | IC50 1.18–19.4 µg/mL | [98] | ||
| Candida species | β-caryophyllene | MIC 1.45–10.0 µg/mL | [98] | ||
| Plasmodium falciparum | Cannabinoids | IC50 4.0–6.7 µM | Chloroquine | IC50 0.1–0.5 µM | [96] |
| P. falciparum | CBNA | IC50 2.4–2.7 µg/mL | [125] | ||
| Trichophyton and Arthroderma species | EO | MIC 0.312–6.3 µg/mL | Griseofulvin | MIC 1.26 to >8.0 µg/mL | [123] |
| Virus | Compound | Activity (µM) | Ref |
|---|---|---|---|
| Epstein–Barr virus (EBV) | THC | IC50 3.0 | [140] |
| Hepatitis C virus (HCV) | CBD | EC50 3.16 | [143] |
| Herpes simplex virus (HSV) | THC | IC50 1.9 | [140] |
| Kaposi sarcoma associated herpesvirus (KSHV) | THC | IC50 3.3 | [140] |
| KSHV | CBD | IC50 2.08 | [144] |
| Murine gamma herpesvirus 68 (MHV) | THC | IC50 1.9 | [140] |
| Corona Virus Group | Compound | Activity (µM) | Reference Drug | Activity (μM) | Ref |
|---|---|---|---|---|---|
| hCov-OC43 | Caflanone | EC50 0.42 | [184] | ||
| SARS-CoV-2 (spike positive) | CBD | EC50 0.64–1.79 | [195] | ||
| SARS-CoV-2 in A549-ACE2 | 7-OH-CBD | EC50 3.6 | [195] | ||
| SARS-CoV-2 | CBD, THC, CBN, CBDA, THCA | IC50 7.91–37.61 | Remdesivir, Lopinavir and Chloroquine | IC50 8.17–13.16 | [189] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mahmud, M.S.; Hossain, M.S.; Ahmed, A.T.M.F.; Islam, M.Z.; Sarker, M.E.; Islam, M.R. Antimicrobial and Antiviral (SARS-CoV-2) Potential of Cannabinoids and Cannabis sativa: A Comprehensive Review. Molecules 2021, 26, 7216. https://doi.org/10.3390/molecules26237216
Mahmud MS, Hossain MS, Ahmed ATMF, Islam MZ, Sarker ME, Islam MR. Antimicrobial and Antiviral (SARS-CoV-2) Potential of Cannabinoids and Cannabis sativa: A Comprehensive Review. Molecules. 2021; 26(23):7216. https://doi.org/10.3390/molecules26237216
Chicago/Turabian StyleMahmud, Md Sultan, Mohammad Sorowar Hossain, A. T. M. Faiz Ahmed, Md Zahidul Islam, Md Emdad Sarker, and Md Reajul Islam. 2021. "Antimicrobial and Antiviral (SARS-CoV-2) Potential of Cannabinoids and Cannabis sativa: A Comprehensive Review" Molecules 26, no. 23: 7216. https://doi.org/10.3390/molecules26237216
APA StyleMahmud, M. S., Hossain, M. S., Ahmed, A. T. M. F., Islam, M. Z., Sarker, M. E., & Islam, M. R. (2021). Antimicrobial and Antiviral (SARS-CoV-2) Potential of Cannabinoids and Cannabis sativa: A Comprehensive Review. Molecules, 26(23), 7216. https://doi.org/10.3390/molecules26237216