
GABAA Receptor-Mediated Sleep-Promoting Effect of Saaz–Saphir Hops Mixture Containing Xanthohumol and Humulone

 , , and
, , and
Abstract
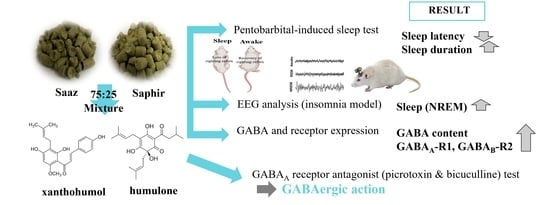
:
1. Introduction
2. Results
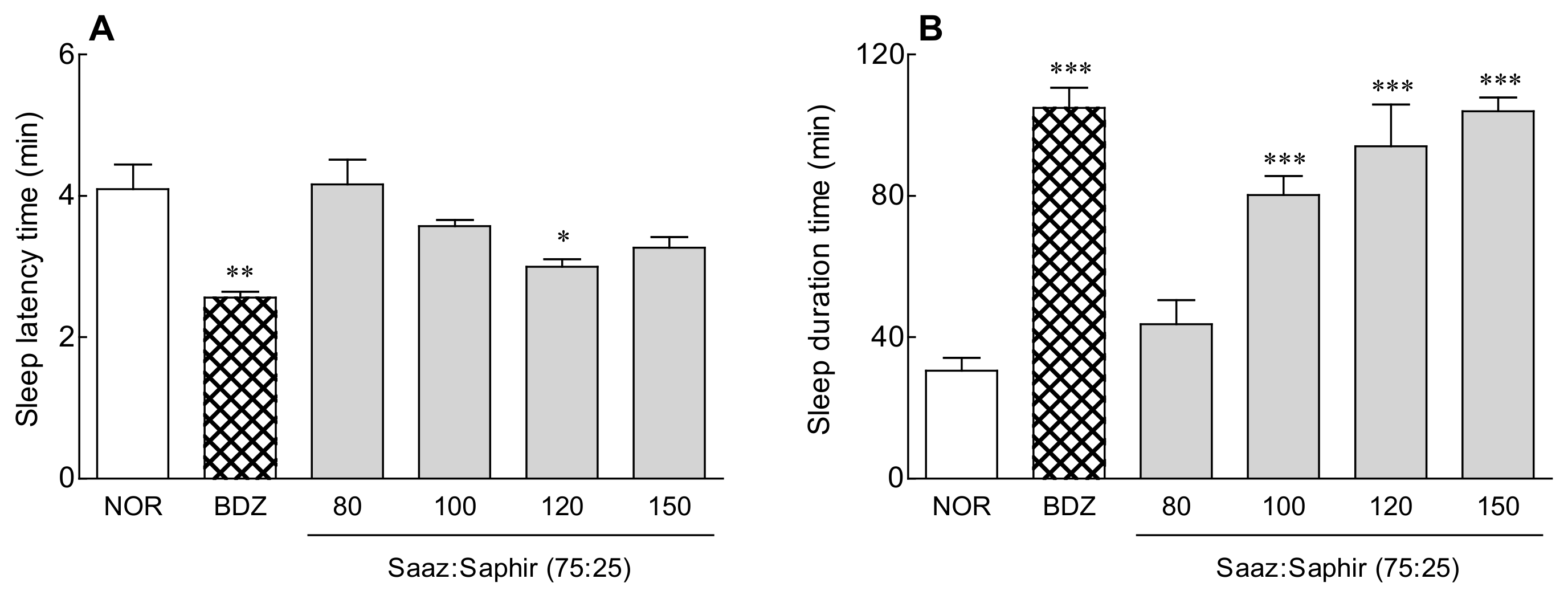
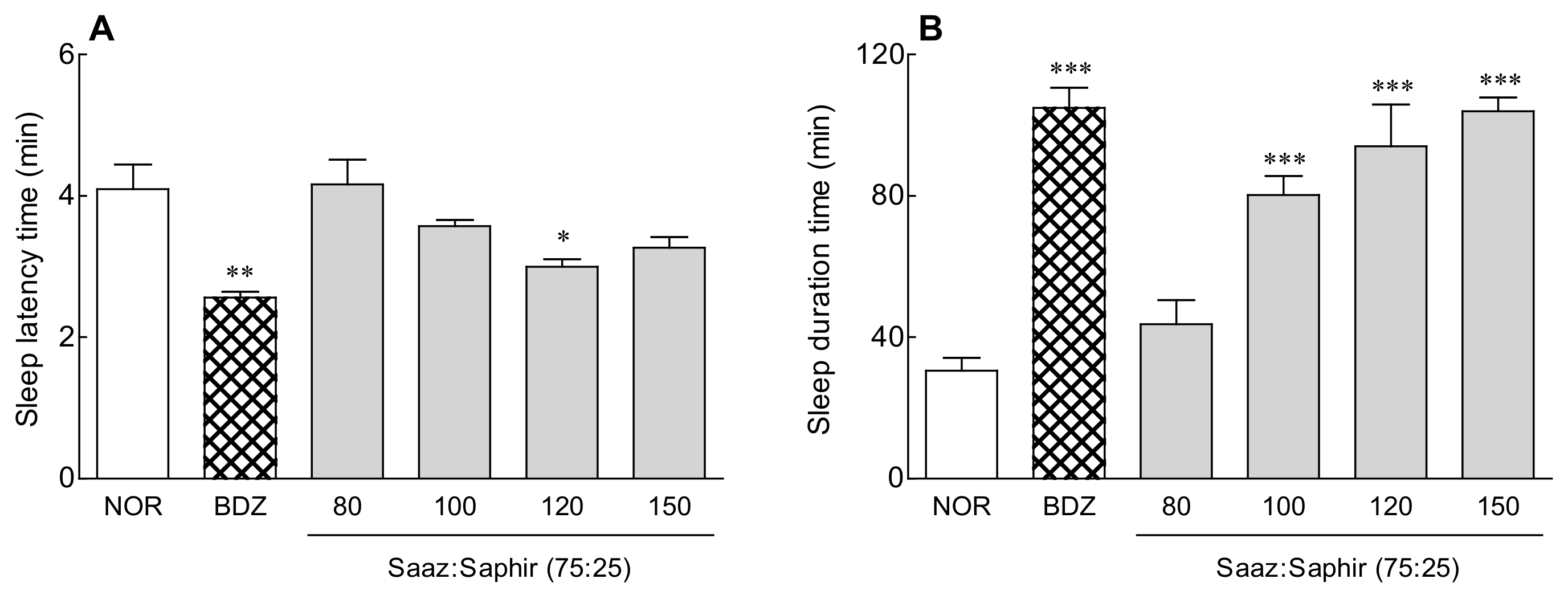
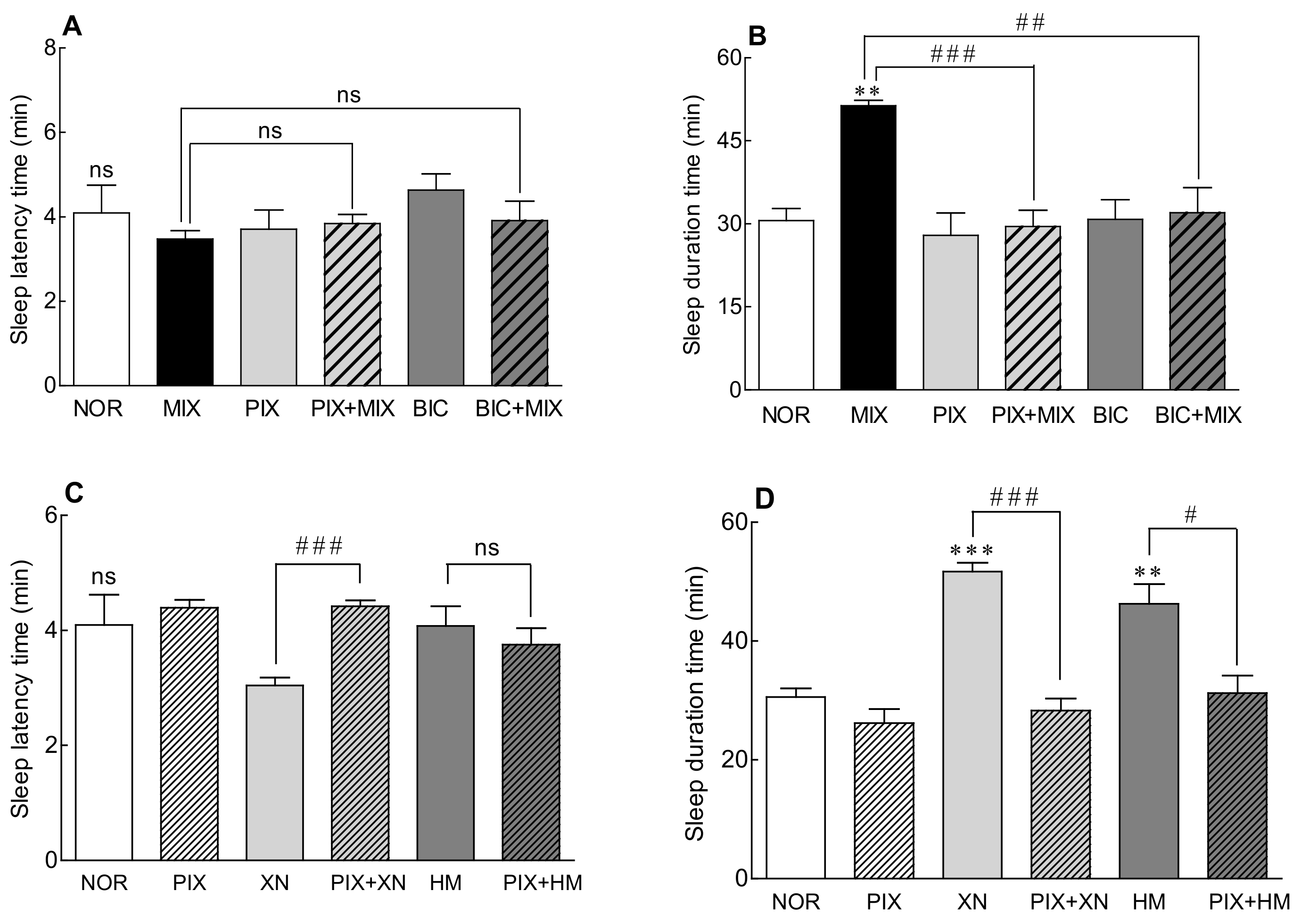
2.1. Sleep Latency and Sleep Duration of the Saaz–Saphir Mixture
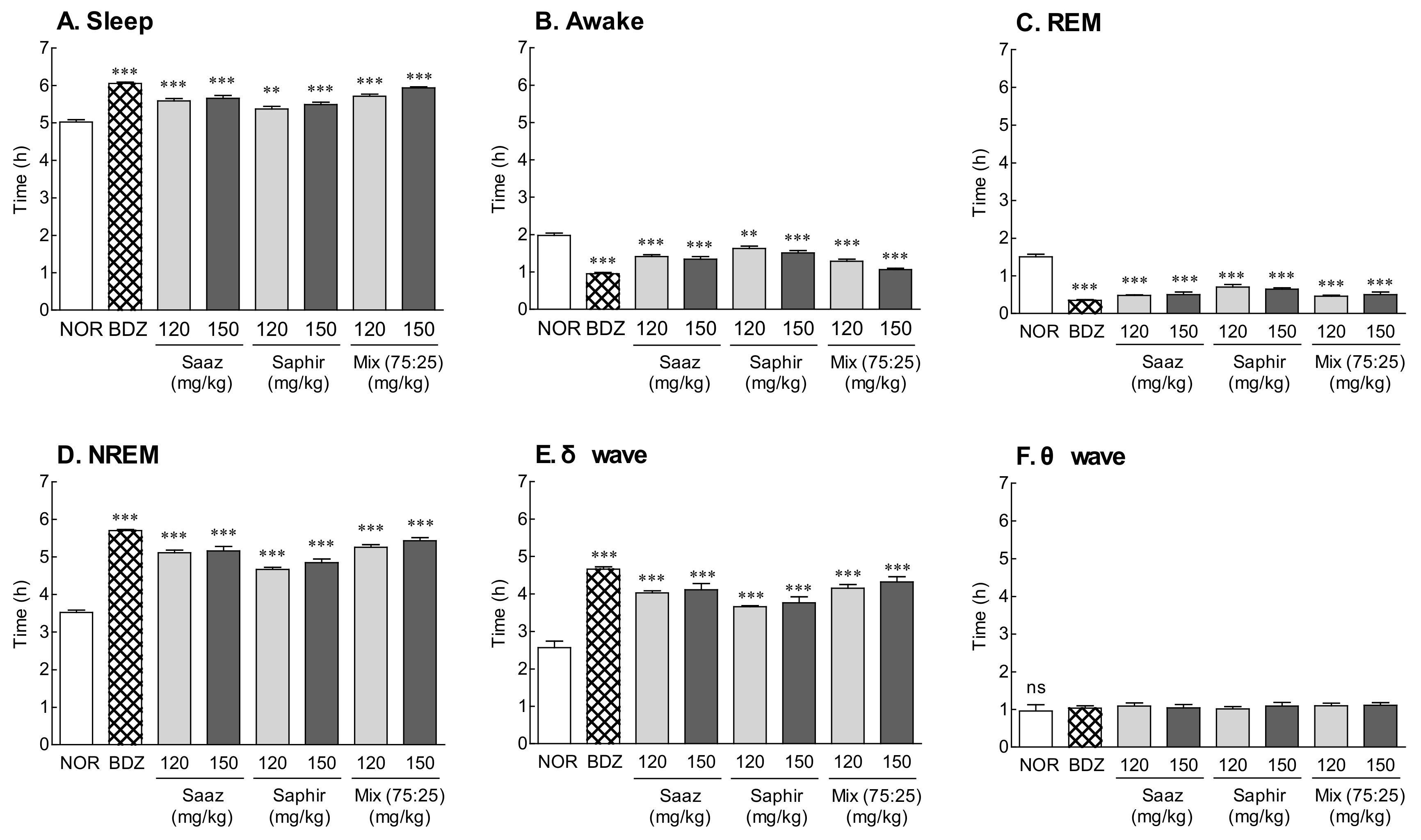
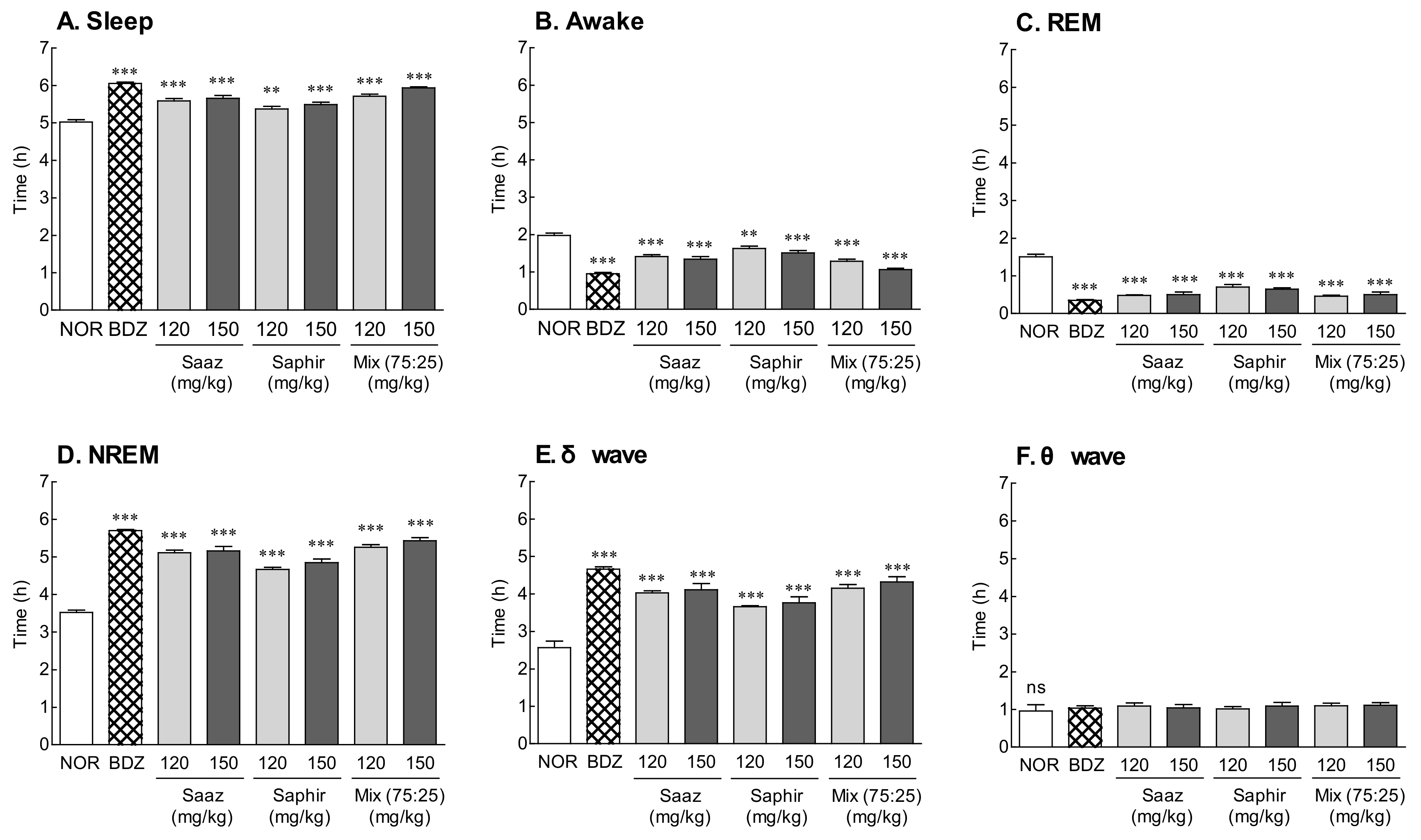
2.2. EEG Analysis by Administration of the Saaz–Saphir Mixture
2.3. EEG Analysis by Administration of the Saaz–Saphir Mixture in the Caffeine-Induced Insomnia Model
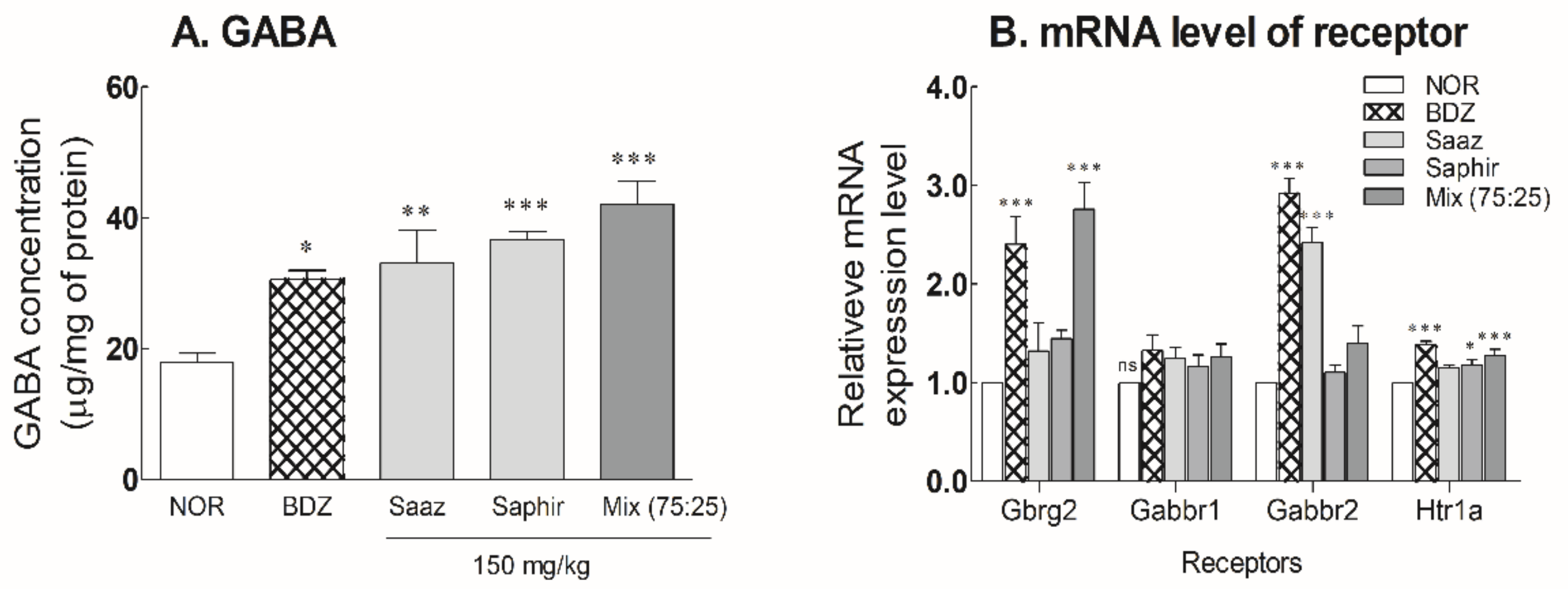
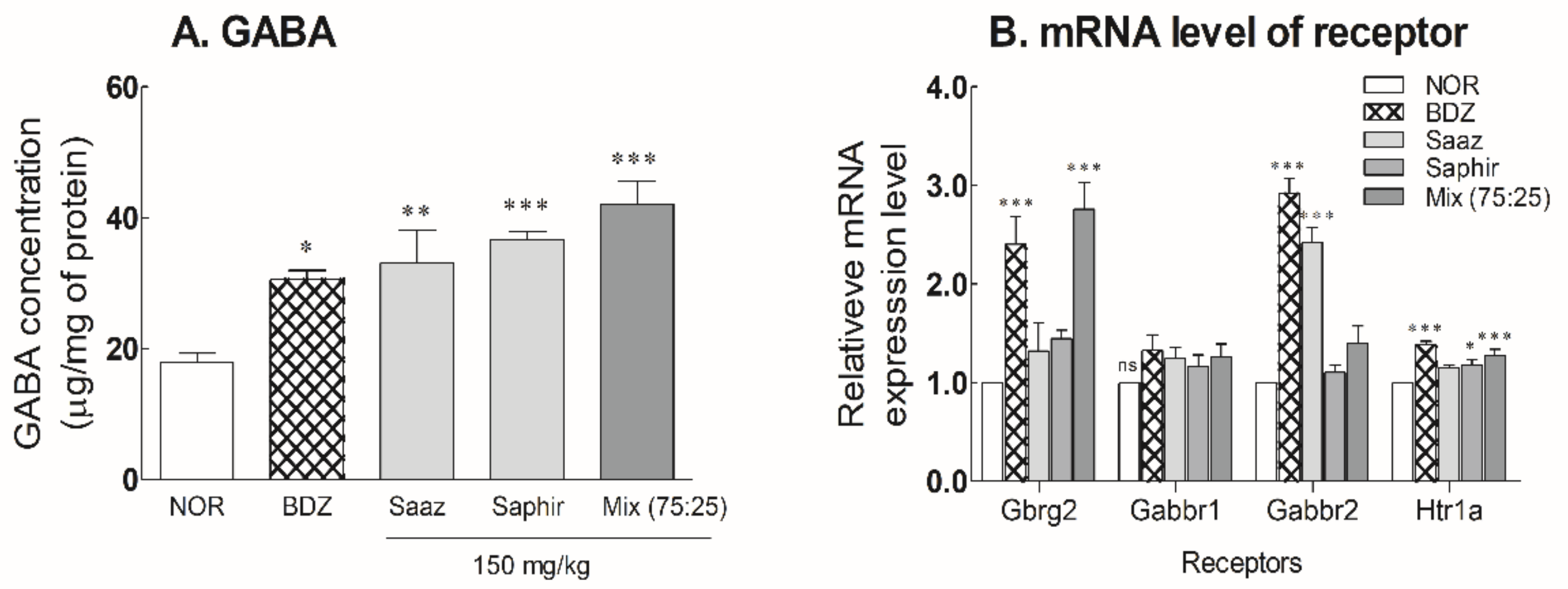
2.4. Changes of GABA and Receptor Expression Levels in Mouse Brain by Oral Administration of the Saaz–Saphir Mixture
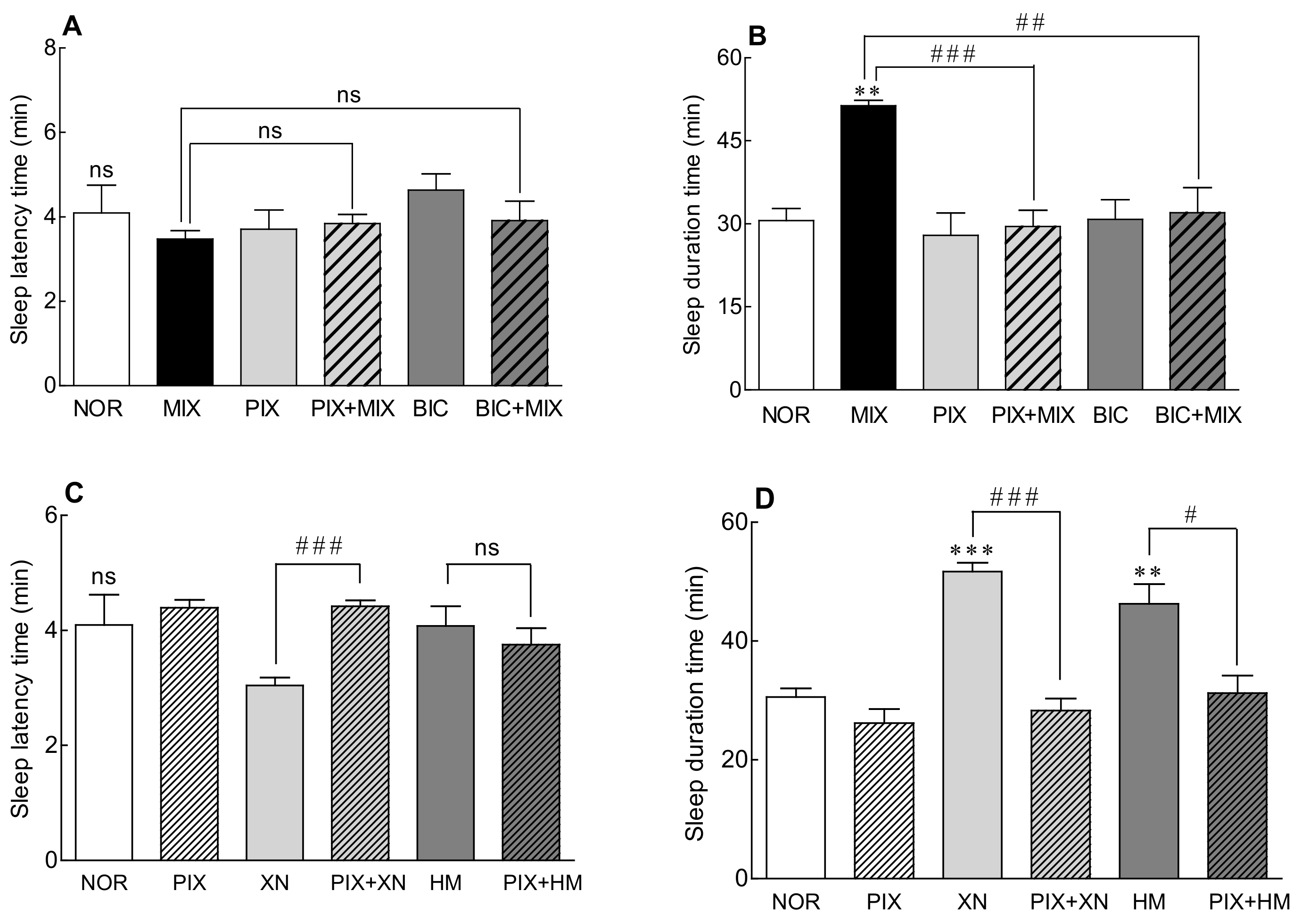
2.5. Receptor Binding Mode of the Saaz–Saphir Mixture by Antagonists
3. Discussion
4. Materials and Methods
4.1. Preparation of Hops Alcoholic Extract
4.2. Flavonoids Assay Using HPLC
4.3. Pentobarbital-Induced Sleep Test in Mouse
4.4. Electroencephalogram (EEG) Analysis
4.5. Analysis of the Receptor Binding Mode Using an Antagonist
4.6. Receptor Gene Expression Level and GABA Content Analysis
4.7. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Sample Availability
References
- Miyama, H.; Shimura, A.; Furuichi, W.; Seki, T.; Ono, K.; Masuya, J.; Odagiri, Y.; Inoue, S.; Inoue, T. Association of chronotypes and sleep disturbance with perceived job stressors and stress response: A covariance structure analysis. Neuropsychiatr. Dis. Treat. 2020, 16, 1997–2005. [Google Scholar] [CrossRef]
- Grimaldi, D.; Reid, K.J.; Papalambros, N.A.; Braun, R.I.; Malkani, R.G.; Abbott, S.M.; Ong, J.C.; Zee, P.C. Autonomic dysregulation and sleep homeostasis in insomnia. Sleep 2021, 44, zsaa274. [Google Scholar] [CrossRef]
- Makarem, N.; Alcántara, C.; Williams, N.; Bello, N.A.; Abdalla, M. Effect of sleep disturbances on blood pressure. Hypertension 2021, 77, 1036–1046. [Google Scholar] [CrossRef]
- Lee, M.; Choh, A.; Demerath, E.; Knutson, K.; Duren, D.; Sherwood, R.; Sun, S.; Chumlea, W.C.; Towne, B.; Siervogel, R. Sleep disturbance in relation to health-related quality of life in adults: The fels longitudinal study. J. Nutr. Health Aging 2009, 13, 576–583. [Google Scholar] [CrossRef] [Green Version]
- Freeman, D.; Sheaves, B.; Waite, F.; Harvey, A.G.; Harrison, P.J. Sleep disturbance and psychiatric disorders. Lancet Psychiatry 2020, 7, 628–637. [Google Scholar] [CrossRef]
- Donoghue, J.; Lader, M. Usage of benzodiazepines: A review. Int. J. Psychiatry Clin. Pract. 2010, 14, 78–87. [Google Scholar] [CrossRef] [PubMed]
- Beer, B.; Clody, D.; Mangano, R.; Levner, M.; Mayer, P.; Barrett, J. A review of the preclinical development of zaleplon, a novel non-benzodiazepine hypnotic for the treatment of insomnia. CNS Drug Rev. 1997, 3, 207–224. [Google Scholar] [CrossRef]
- Liu, M.; Hansen, P.E.; Wang, G.; Qiu, L.; Dong, J.; Yin, H.; Qian, Z.; Yang, M.; Miao, J. Pharmacological profile of xanthohumol, a prenylated flavonoid from hops (Humulus lupulus). Molecules 2015, 20, 754–779. [Google Scholar] [CrossRef]
- Zanoli, P.; Zavatti, M. Pharmacognostic and pharmacological profile of Humulus lupulus L. J. Ethnopharmacol. 2008, 116, 383–396. [Google Scholar] [CrossRef] [PubMed]
- Arruda, T.R.; Pinheiro, P.F.; Silva, P.I.; Bernardes, P.C. A new perspective of a well-recognized raw material: Phenolic content, antioxidant and antimicrobial activities and α-and β-acids profile of Brazilian hop (Humulus lupulus L.) extracts. LWT—Food Sci. Technol. 2021, 141, 110905. [Google Scholar] [CrossRef]
- Benkherouf, A.Y.; Eerola, K.; Soini, S.L.; Uusi-Oukari, M. Humulone modulation of GABAA receptors and its role in hops sleep-promoting activity. Front. Neurosci. 2020, 14, 594708. [Google Scholar] [CrossRef]
- Meissner, O.; Häberlein, H. Influence of xanthohumol on the binding behavior of GABAA receptors and their lateral mobility at hippocampal neurons. Planta Med. 2006, 72, 656–658. [Google Scholar] [CrossRef]
- Vazquez-Cervantes, G.I.; Ortega, D.R.; Blanco Ayala, T.; Pérez de la Cruz, V.; Esquivel, D.F.G.; Salazar, A.; Pineda, B. Redox and anti-inflammatory properties from hop components in beer-related to neuroprotection. Nutrients 2021, 13, 2000. [Google Scholar] [CrossRef]
- Bonnet, M.H.; Arand, D.L. Caffeine use as a model of acute and chronic insomnia. Sleep 1992, 15, 526–536. [Google Scholar]
- Nehlig, A.; Daval, J.L.; Debry, G. Caffeine and the central nervous system: Mechanisms of action, biochemical, metabolic and psychostimulant effects. Brain Res. Rev. 1992, 17, 139–170. [Google Scholar] [CrossRef]
- Winkelman, J.W.; Buxton, O.M.; Jensen, J.E.; Benson, K.L.; O’Connor, S.P.; Wang, W.; Renshaw, P.F. Reduced brain GABA in primary insomnia: Preliminary data from 4T proton magnetic resonance spectroscopy (1H-MRS). Sleep 2008, 31, 1499–1506. [Google Scholar] [CrossRef] [Green Version]
- Plante, D.T.; Jensen, J.E.; Schoerning, L.; Winkelman, J.W. Reduced gamma-aminobutyric acid in occipital and anterior cingulate cortices in primary insomnia: A link to major depressive disorder? Neuropsychopharmacology 2012, 37, 1548–1557. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kakeda, S.; Korogi, Y.; Moriya, J.; Ohnari, N.; Sato, T.; Ueno, S.; Yanagihara, N.; Harada, M.; Matsuda, T. Influence of work shift on glutamic acid and gamma-aminobutyric acid (GABA): Evaluation with proton magnetic resonance spectroscopy at 3T. Psychiatry Res. Neuroimaging 2011, 192, 55–59. [Google Scholar] [CrossRef] [PubMed]
- Mignot, E.; Taheri, S.; Nishino, S. Sleeping with the hypothalamus: Emerging therapeutic targets for sleep disorders. Nat. Neurosci. 2002, 5, 1071–1075. [Google Scholar] [CrossRef] [PubMed]
- Bravo, R.; Matito, S.; Cubero, J.; Paredes, S.D.; Franco, L.; Rivero, M.; Rodríguez, A.B.; Barriga, C. Tryptophan-enriched cereal intake improves nocturnal sleep, melatonin, serotonin, and total antioxidant capacity levels and mood in elderly humans. Age 2013, 35, 1277–1285. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sieghart, W. Structure and pharmacology of gamma-aminobutyric acid(a) receptor subtypes. Pharmacol. Rev. 1995, 47, 181–234. [Google Scholar] [PubMed]
- Aguayo, L.G.; Peoples, R.W.; Yeh, H.H.; Yevenes, G.E. GABA-A receptors as molecular sites of ethanol action. Direct or indirect actions? Curr. Top. Med. Chem. 2002, 2, 869–885. [Google Scholar] [CrossRef] [PubMed]
- Förstera, B.; Castro, P.A.; Moraga-Cid, G.; Aguayo, L.G. Potentiation of g aminobutyric acid receptors (GABAAR) by ethanol: How are inhibitory receptors affected? Front. Cell. Neurosci. 2016, 10, 114. [Google Scholar] [CrossRef] [Green Version]
- Sieghart, W.; Fuchs, K.; Tretter, V.; Ebert, V.; Jechlinger, M.; Höger, H.; Adamiker, D. Structure and subunit composition of GABAA receptors. Neurochem. Int. 1999, 34, 379–385. [Google Scholar] [CrossRef]
- Kerr, D.I.; Ong, J. GABA agonists and antagonists. Med. Res. Rev. 1992, 12, 593–636. [Google Scholar] [CrossRef]
- Khawaled, R.; Bruening-Wright, A.; Adelman, J.P.; Maylie, J. Bicuculline block of small-conductance calcium-activated potassium channels. Pflugers Arch. 1999, 438, 314–321. [Google Scholar] [CrossRef] [PubMed]
- Winsky-Sommerer, R. Role of GABAA receptors in the physiology and pharmacology of sleep. Eur. J. Neurosci. 2009, 29, 1779–1794. [Google Scholar] [CrossRef] [Green Version]
- Kim, C.S.; Han, J.Y.; Kim, S.H.; Hong, J.T.; Oh, K.W. Herbs for the treatment of insomnia. Biomol. Ther. 2011, 19, 274–281. [Google Scholar] [CrossRef] [Green Version]
- Choi, H.S.; Hong, K.B.; Han, S.H.; Suh, H.J. Valerian/Cascade mixture promotes sleep by increasing non-rapid eye movement (NREM) in rodent model. Biomed. Pharmacother. 2018, 99, 913–920. [Google Scholar] [CrossRef] [PubMed]
- Buckee, G.; Baker, C. Estimation of α-and β-acids in hop extracts by HPLC. J. Inst. Brew. 1987, 93, 468–471. [Google Scholar] [CrossRef]
- Salazar, C.; Armenta, J.M.; Cortés, D.F.; Shulaev, V. Combination of an AccQ Tag-ultra performance liquid chromatographic method with tandem mass spectrometry for the analysis of amino acids. In Amino Acid Analysis; Alterman, M.A., Ahunziker, P., Eds.; Springer: Hertfordshire, UK, 2012; pp. 13–28. [Google Scholar]
- Liu, W.L.; Wu, B.F.; Shang, J.H.; Zhao, Y.L.; Huang, A.X. Moringa oleifera Lam seed oil augments pentobarbital-induced sleeping behaviors in mice via GABAergic systems. J. Agric. Food Chem. 2020, 68, 3149–3162. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Hop | Xanthohumol | α-Acids | β-Acids | |||
|---|---|---|---|---|---|---|
| Humulone | Cohumulone | Adhumulone | Colupulone | Adlupulone | ||
| Saaz | 2.51 ± 0.13 | 47.28 ± 0.23 | 5.41 ± 0.01 | 17.91 ± 0.05 | 20.68 ± 0.05 | 28.83 ± 0.53 |
| Saphir | 4.23 ± 0.14 | 53.13 ± 0.20 | 3.14 ± 0.09 | 27.67 ± 0.24 | 27.12 ± 0.07 | 39.24 ± 0.10 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Min, B.; Ahn, Y.; Cho, H.-J.; Kwak, W.-K.; Suh, H.J.; Jo, K. GABAA Receptor-Mediated Sleep-Promoting Effect of Saaz–Saphir Hops Mixture Containing Xanthohumol and Humulone. Molecules 2021, 26, 7108. https://doi.org/10.3390/molecules26237108
Min B, Ahn Y, Cho H-J, Kwak W-K, Suh HJ, Jo K. GABAA Receptor-Mediated Sleep-Promoting Effect of Saaz–Saphir Hops Mixture Containing Xanthohumol and Humulone. Molecules. 2021; 26(23):7108. https://doi.org/10.3390/molecules26237108
Chicago/Turabian StyleMin, Byungjick, Yejin Ahn, Hyeok-Jun Cho, Woong-Kwon Kwak, Hyung Joo Suh, and Kyungae Jo. 2021. "GABAA Receptor-Mediated Sleep-Promoting Effect of Saaz–Saphir Hops Mixture Containing Xanthohumol and Humulone" Molecules 26, no. 23: 7108. https://doi.org/10.3390/molecules26237108
APA StyleMin, B., Ahn, Y., Cho, H.-J., Kwak, W.-K., Suh, H. J., & Jo, K. (2021). GABAA Receptor-Mediated Sleep-Promoting Effect of Saaz–Saphir Hops Mixture Containing Xanthohumol and Humulone. Molecules, 26(23), 7108. https://doi.org/10.3390/molecules26237108